Nuisance Algae in Ballast Water Facing International Conventions. Insights from DNA Metabarcoding in Ships Arriving in Bay of Biscay

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Analysed
2.2. DNA Analysis
2.3. Bioinformatics and Statistical Analysis
3. Results
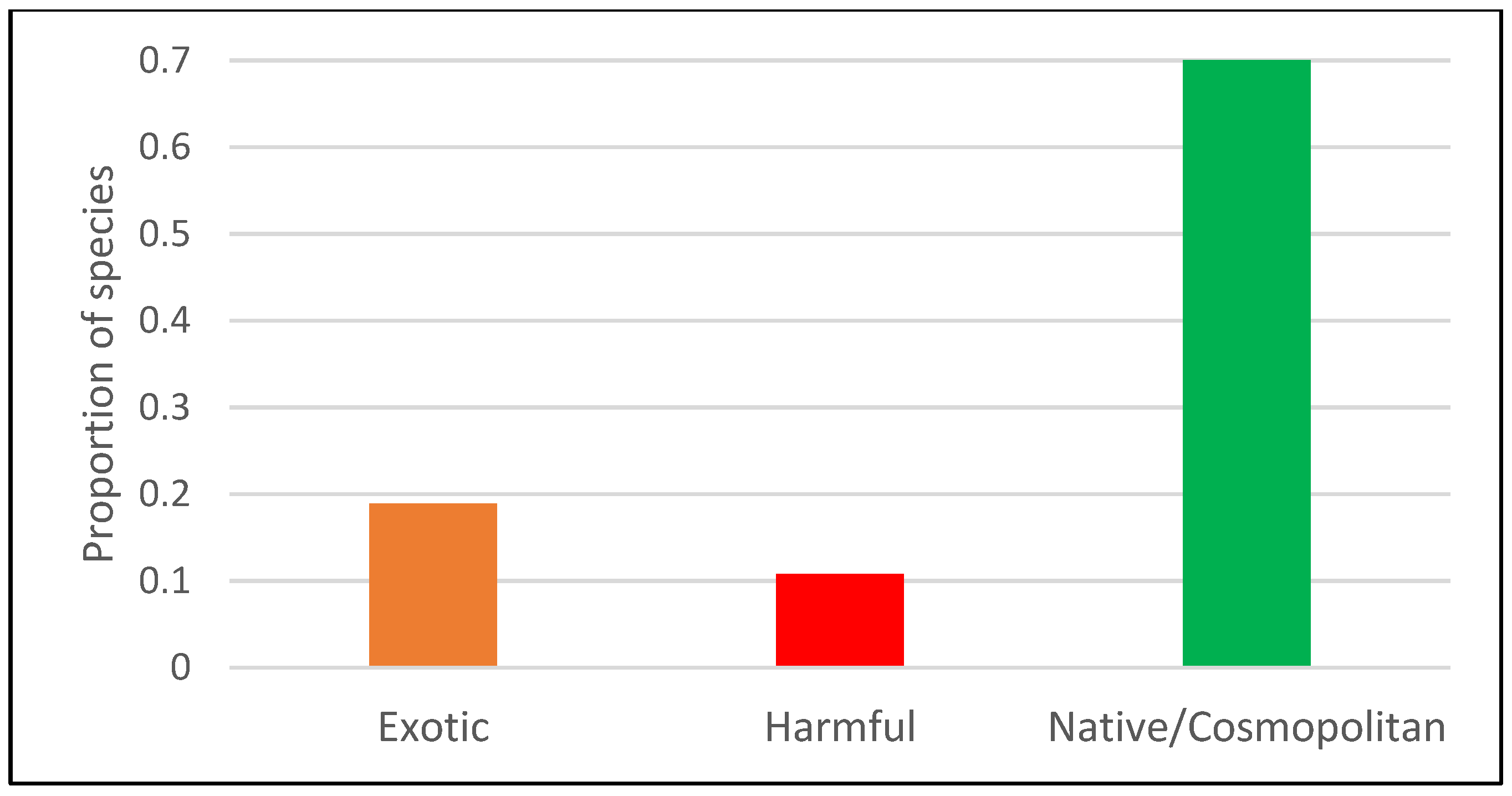
3.1. Algae Community in BW of Ships Arriving in Gijon Port
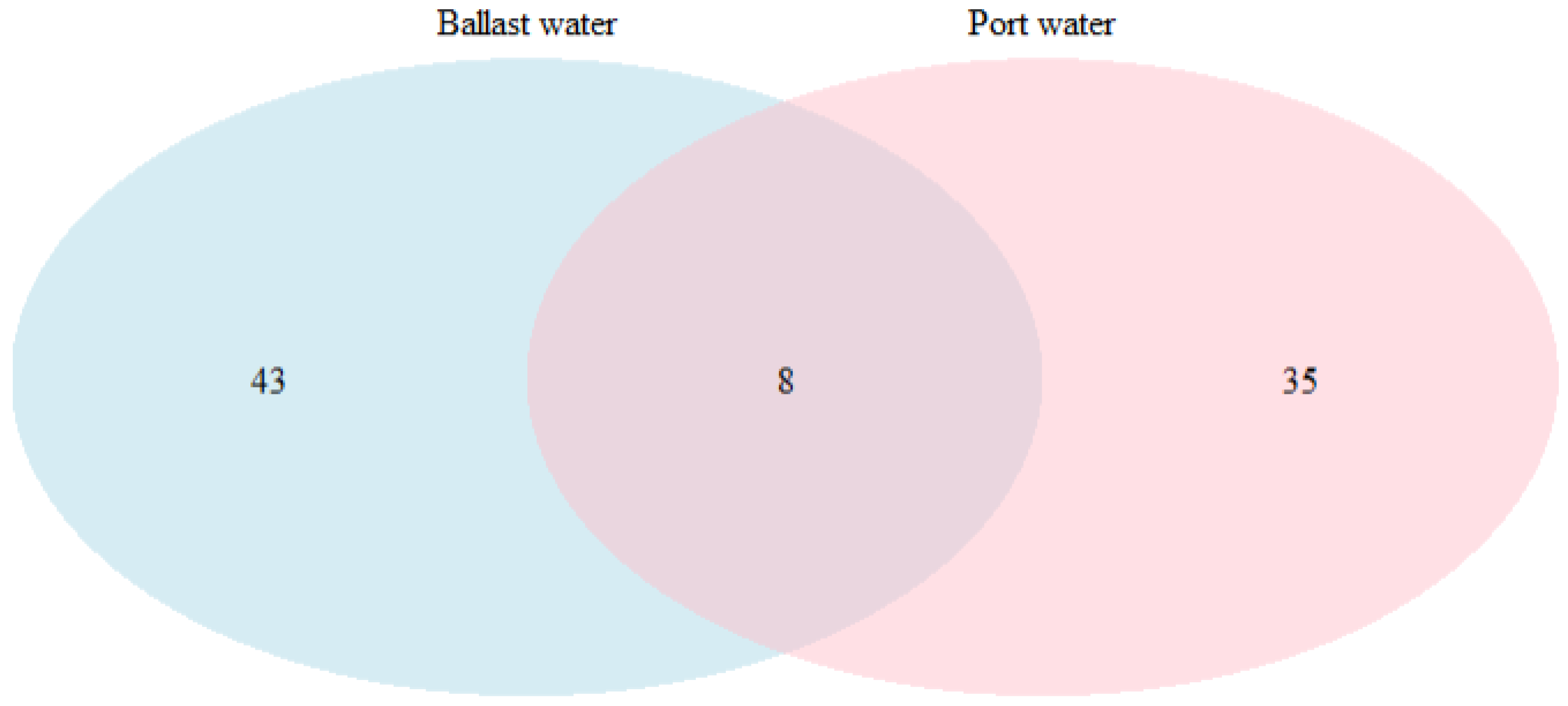
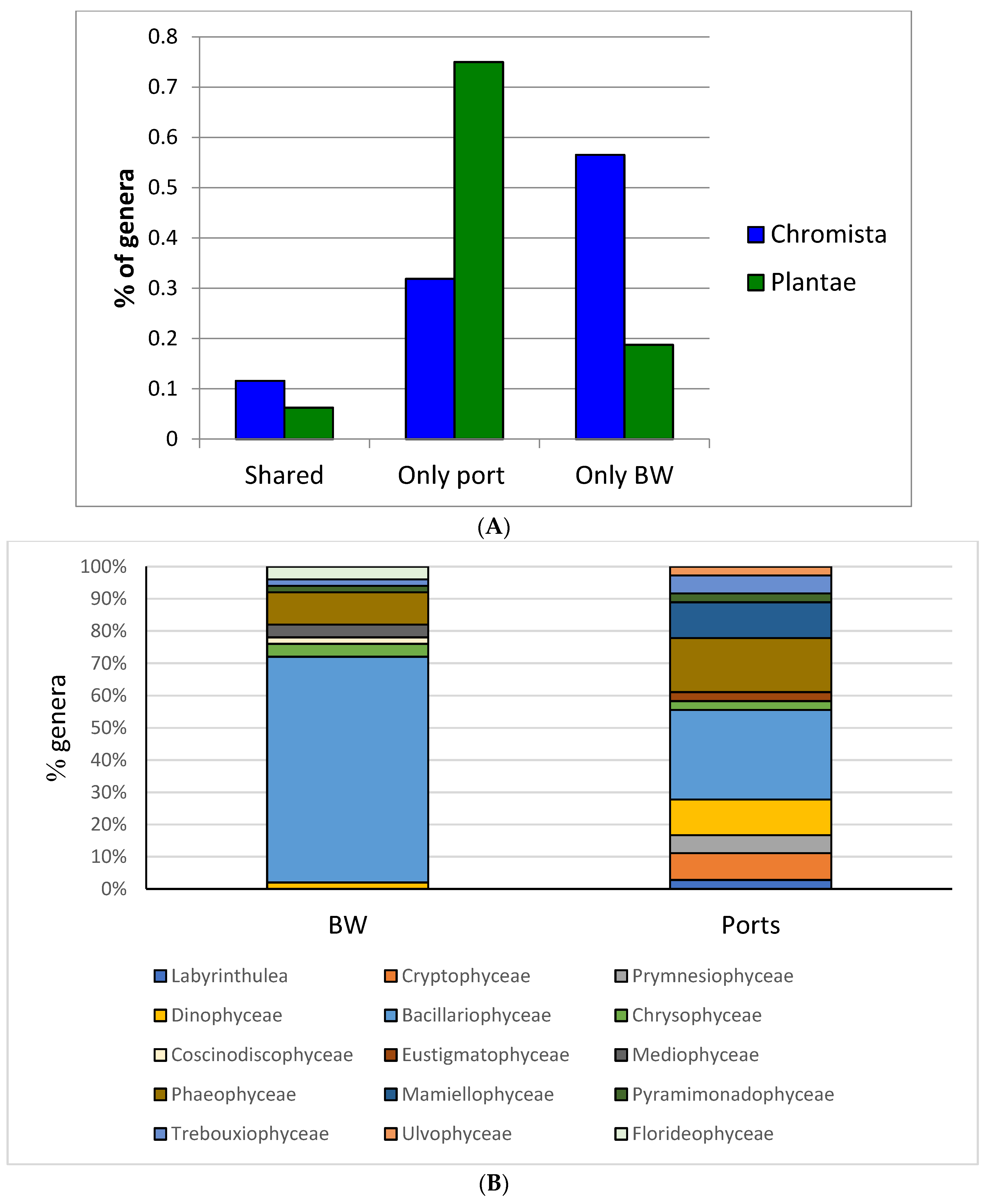
3.2. Comparison of BW and Port Phytoplankton Detected Through NGS in the Region
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethics statement
Accessibility of molecular data
References
- Cohen, N.J.; Slaten, D.D.; Marano, N.; Tappero, J.W.; Wellman, M.; Albert, R.; Hill, V.R.; Espey, D.; Handzel, T.; Henry, A.; et al. Preventing maritime transfer of toxigenic vibrio cholerae. Emerg. Infect. Dis. 2012, 18, 1680–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, C.; Lourenço, N.; Lopes, T.; Rall, V.L.; Lopes, C. Ballast water: A review of the impact on the world public health. J. Venom. Anim. Toxins Incl. Trop. Dis. 2008, 14, 393–408. [Google Scholar] [CrossRef]
- Frazier, M.; Miller, A.W.; Ruiz, G.M. Linking science and policy to prevent the spread of invasive species from the ballast water discharge of ships. Ecol. Appl. 2013, 23, 287–289. [Google Scholar] [CrossRef] [PubMed]
- Lilly, E.L.; Kulis, D.M.; Gentien, P.; Anderson, D.M. Paralytic shellfish poisoning toxins in France linked to a human-introduced strain of Alexandrium catenella from the western Pacific: Evidence from DNA and toxin analysis. J. Plankton Res. 2002, 24, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Butrón, A.; Orive, E.; Madariaga, I. Potential risk of harmful algae transport by ballast waters: The case of Bilbao Harbour. Mar. Pollut. Bull. 2011, 62, 747–757. [Google Scholar] [CrossRef] [PubMed]
- van den Bergh, J.C.J.M.; Nunes, A.L.D.; Dotinga, H.M.; Kooistra, W.H.C.M.; Vrieling, W.F.; Peperzak, L. Exotic harmful algae in marine ecosystems: An integrated biological–economic– legal analysis of impacts and policies. Mar. Policy 2002, 26, 59–74. [Google Scholar] [CrossRef]
- Pertola, S.; Faust, M.A.; Kuosa, H. Survey on germination and species composition of dinoflagellates from ballast tanks and recent sediments in ports on the South Coast of Finland, North-Eastern Baltic Sea. Mar. Pollut. Bull. 2006, 52, 900–911. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, K.; Rohit, M.; Mohan, S.V. Microalgae-based carotenoids production. In Algal Green Chemistry; Elsevier: Amsterdam, The Netherlands, 2017; pp. 139–147. [Google Scholar]
- David, M.; Gollasch, S. Global Maritime Transport and Ballast Water Management; Springer: Dordrecht, The Netherlands, 2015. [Google Scholar]
- Casas-Monroy, O.; Linley, R.D.; Adams, J.K.; Chan, F.T.; Drake, D.A.R.; Bailey, S.A. Relative invasion risk for plankton across marine and freshwater systems: Examining efficacy of proposed international ballast water discharge standards. PLoS ONE 2015, 10, e0118267. [Google Scholar] [CrossRef] [Green Version]
- Rey, A.; Basurko, O.C.; Rodriguez-Ezpeleta, N.; Rey, A.; Rodríguez-Ezpeleta, N. The challenges and promises of genetic approaches for ballast water management. J. Sea Res. 2018, 133, 134–145. [Google Scholar] [CrossRef] [Green Version]
- Ardura, A.; Zaiko, A.; Martínez, J.L.; Samuiloviene, A.; Borrell, Y.; Garcia-Vazquez, E. Environmental DNA evidence of transfer of North Sea molluscs across tropical waters through ballast water. J. Molluscan Stud. 2015, 81, 495–501. [Google Scholar] [CrossRef]
- Zaiko, A.; Martinez, J.L.; Schmidt-Petersen, J.; Ribicic, D.; Samulioviene, A.; Vazquez, E.G. Metabarcoding approach for the ballast water surveillance—An advantageous solution or an awkward challenge? Mar. Pollut. Bull. 2015, 92, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Zaiko, A.; Martínez, J.L.; Ardura, A.; Clusa, L.; Borrell, Y.J.; Samuiloviene, A.; Roca, A.; Vazquez, E.G. Detecting nuisance species using NGST: Methodology shortcomings and possible application in ballast water monitoring. Mar. Environ. Res. 2015, 112, 64–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Yu, S.-X.; Wang, Y.-C.; Li, J.-L.; Li, F.-C.; Qin, S. Diversity of phototrophic phytoplankton in Northern South China Sea indicated by rbcL analysis. Environ. Biol. Fishes 2015, 28, 773–781. [Google Scholar] [CrossRef]
- Borrell, Y.J.; Miralles, L.; Huu, H.D.; Mohammed-Geba, K.; Garcia-Vazquez, E. DNA in a bottle—Rapid metabarcoding survey for early alerts of invasive species in ports. PLoS ONE 2017, 12, e0183347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leray, M.; Yang, J.Y.; Meyer, C.; Mills, S.C.; Agudelo, N.; Ranwez, V.; Boehm, J.T.; Machida, R.J. A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity: Application for characterizing coral reef fish gut contents. Front. Zool. 2013, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Geller, J.; Meyer, C.; Parker, M.; Hawk, H. Redesign of PCR primers for mitochondrial cytochromecoxidase subunit I for marine invertebrates and application in all-taxa biotic surveys. Mol. Ecol. Resour. 2013, 13, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Rivera, S.F.; Vasselon, V.; Jacquet, S.; Bouchez, A.; Ariztegui, D.; Rimet, F. Metabarcoding of lake benthic diatoms: From structure assemblages to ecological assessment. Hydrobiologia 2017, 807, 37–51. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Scott, R.; Zhan, A.; Brown, E.A.; Chain, F.J.J.; Cristescu, M.E.; Gras, R.; MacIsaac, H.J. Optimization and performance testing of a sequence processing pipeline applied to detection of nonindigenous species. Evol. Appl. 2018, 11, 891–905. [Google Scholar] [CrossRef] [Green Version]
- Von Ammon, U.; Wood, S.A.; Laroche, O.; Zaiko, A.; Tait, L.W.; Lavery, S.; Inglis, G.J.; Pochon, X. Combining morpho-taxonomy and metabarcoding enhances the detection of non-indigenous marine pests in biofouling communities. Sci. Rep. 2018, 8, 16290. [Google Scholar] [CrossRef] [Green Version]
- Helleren, S. The Diatom Chaetoceros spp. as A Potential Contributing Factor to Fish Mortality Events in Cockburn Sound. Dalcon Environmental Report DE00000.R1; 2016. Available online: https://www.fish.wa.gov.au/Documents/corporate_publications/chaetoceros-report-cockburn-sound-nov-2015.pdf (accessed on 2 June 2020).
- Dittami, S.M.; Hostyeva, V.; Egge, E.S.; Kegel, J.; Eikrem, W.; Edvardsen, B. Seasonal dynamics of harmful algae in outer Oslofjorden monitored by microarray, qPCR, and microscopy. Environ. Sci. Pollut. Res. 2013, 20, 6719–6732. [Google Scholar] [CrossRef]
- Bates, S.S.; Richard, D.J.A. Shellfish harvest area closure due to domoic acid—Mill river, Prince Edward Island. Harmful Algae News 2000, 21, 6–7. [Google Scholar]
- Lassus, P.; Chomérat, N.; Hess, P.; Nézan, E. Toxic and harmful micro algae of the world ocean/Micro-algues toxiques et nuisibles de l’océan Mondial; International society for the study of harmful algae: Copenhagen, Denmark and United Nations Educational, Scientific and Cultural Organi statio: Paris, France, 2016; p. 68. [Google Scholar]
- Twiner, M.J.; Dixon, S.J.; Trick, C.G. Toxic effects of Heterosigma akashiw o do not appear to be mediated by hydrogen peroxide. Limnol. Oceanogr. 2001, 46, 1400–1405. [Google Scholar] [CrossRef] [Green Version]
- Cerino, F.; Orsini, L.; Sarn, D.; Dell’Aversano, C.; Tartaglione, L.; Zingone, A. The alternation of different morphotypes in the seasonal cycle of the toxic diatom Pseudo-nitzschia galaxiae. Harmful Algae 2005, 4, 33–48. [Google Scholar] [CrossRef]
- Murphy, K.R.; Ritz, D.; Hewitt, C. Heterogeneous zooplankton distribution in a ship’s ballast tanks. J. Plankton Res. 2002, 24, 729–734. [Google Scholar] [CrossRef]
- Rajakaruna, H.; Vandenbyllaardt, J.; Kydd, J.; Bailey, S.A. Modeling the distribution of colonial species to improve estimation of plankton concentration in ballast water. J. Sea Res. 2018, 133, 166–176. [Google Scholar] [CrossRef]
- Sutton, C.A.; Murphy, K.R.; Martin, R.B.; Hewitt, C.L. A Review and Evaluation of Ballast Water Sampling Protocols; CSIRO Division of Marine Research: Hobart, Australia, 1998. [Google Scholar]
- Zaiko, A.; Pochon, X.; Garcia-Vazquez, E.; Olenin, S.; Wood, S.A. Advantages and Limitations of Environmental DNA/RNA Tools for Marine Biosecurity: Management and Surveillance of Non-indigenous Species. Front. Mar. Sci. 2018, 5, 322. [Google Scholar] [CrossRef] [Green Version]
- Miralles, L.; Ardura, A.; Arias, A.; Borrell, Y.; Clusa, L.; Dopico, E.; De Rojas, A.H.; Lopez, B.; Colmenero, A.M.M.; Roca, A.; et al. Barcodes of marine invertebrates from north Iberian ports: Native diversity and resistance to biological invasions. Mar. Pollut. Bull. 2016, 112, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Shea, K. Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
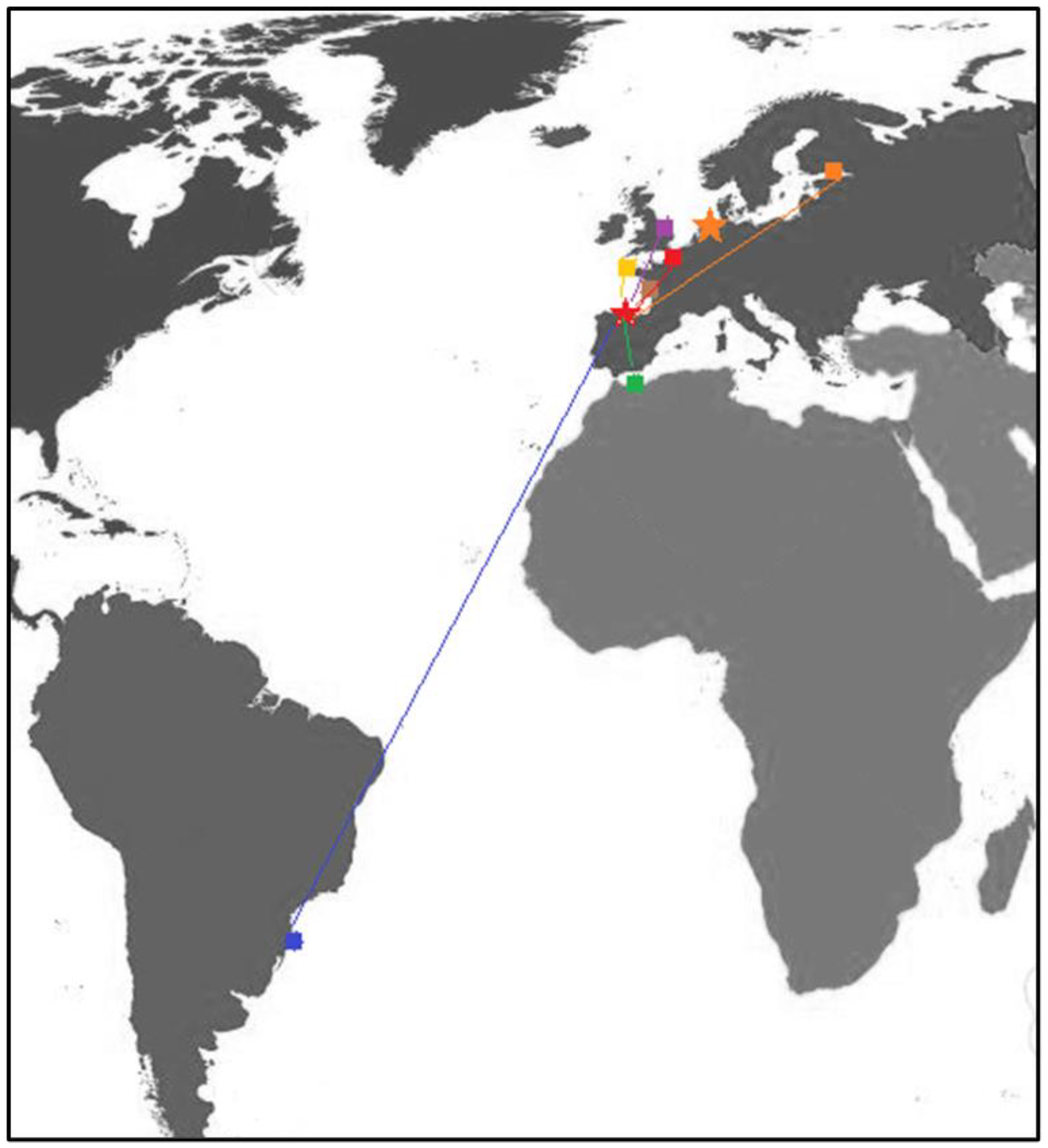
| Ship | Type of Ship | Origin Port | Country | Region | Sampling Date | Days since BW Upload | Species Detected | Risk Species |
|---|---|---|---|---|---|---|---|---|
| Ship#1 | General cargo | Brest | France | English Channel/Atlantic Ocean | 31/08/2017 | 3 | 1 | 0 |
| Ship#2 | LPG tanker | Nador | Morocco | Mediterranean Sea | 09/10/2017 | 39 | 4 | 1 |
| Ship#3 | Bulk carrier | Port of Tubarao | Brazil | SW Atlantic Ocean | 19/10/2017 | 3 | 1 | 1 |
| Ship#4 | General cargo | Gunnes Warf | United Kingdom | Fluvial port (River Trent), North Sea | 12/12/2017 | 12 | 66 | 20 |
| Ship#5 | Oil tanker | La Pallice (La Rochelle) | France | NW Bay of Biscay/Atlantic Ocean | 05/07/2017 | 1 | 1 | 1 |
| Ship#6 | Bulk carrier | Vysotsk | Russia | Baltic Sea | 11/07/2017 | 4 | 0 | 0 |
| Ship#7 | General cargo | Rotterdam | Netherlands | Fluvial port, North Sea | 13/07/17 | 1 | 0 | 0 |
| COI | rbcL | ||||||
|---|---|---|---|---|---|---|---|
| Ship | Sample | QC Reads | Assigned | After Expert Check | QC Reads | Assigned | After Expert Check |
| Ship#1 | 1 | 130 | 0 | 0 | 538 | 0 | 0 |
| 2 | 26 | 2 | 0 | 244 | 1 | 1 | |
| Ship#2 | 1 | 138,174 | 9.234 | 3.027 | 73,400 | 372 | 372 |
| 2 | 6 | 0 | 0 | 72,450 | 0 | 0 | |
| Ship#3 | 1 | 634 | 98 | 47 | 14 | 0 | 0 |
| 2 | 24 | 0 | 0 | 18 | 1 | 1 | |
| Ship#4 | 1 | 131,180 | 16.585 | 14.281 | 50,044 | 10.234 | 8.997 |
| 2 | 4 | 0 | 0 | 62 | 0 | 0 | |
| Ship#5 | 1 | 32 | 0 | 0 | 1280 | 1 | 1 |
| 2 | 80 | 1 | 0 | 1160 | 0 | 0 | |
| Ship#6 | 1 | 24 | 0 | 0 | 1694 | 0 | 0 |
| 2 | 22 | 0 | 0 | 1020 | 0 | 0 | |
| Ship#7 | 1 | 28 | 1 | 0 | 22 | 0 | 0 |
| 2 | 2906 | 4 | 1 | 10 | 0 | 0 | |
| Species | Cause of Concern | Origin | |
|---|---|---|---|
| #2, Nador | #4, Trent | ||
| Chaetoceros septentrionalis | Harmful algae, HAB (23) | X | |
| Blixaea quinquecornis | Baltic, Bloom forming species (28) | X | |
| Cylindrotheca closterium | Cosmopolitan, Bloom forming (29) | X | |
| Halamphora sp. | HAB (4) | X | |
| Heterosigma akashiwo | NW Pacific, HAB (30) | X | |
| Pseudo-nitzschia galaxiae | NW Atlantic HAB (30) | X | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ardura, A.; Borrell, Y.J.; Fernández, S.; González Arenales, M.; Martínez, J.L.; Garcia-Vazquez, E. Nuisance Algae in Ballast Water Facing International Conventions. Insights from DNA Metabarcoding in Ships Arriving in Bay of Biscay. Water 2020, 12, 2168. https://doi.org/10.3390/w12082168
Ardura A, Borrell YJ, Fernández S, González Arenales M, Martínez JL, Garcia-Vazquez E. Nuisance Algae in Ballast Water Facing International Conventions. Insights from DNA Metabarcoding in Ships Arriving in Bay of Biscay. Water. 2020; 12(8):2168. https://doi.org/10.3390/w12082168
Chicago/Turabian StyleArdura, Alba, Yaisel J. Borrell, Sara Fernández, Mónica González Arenales, José L. Martínez, and Eva Garcia-Vazquez. 2020. "Nuisance Algae in Ballast Water Facing International Conventions. Insights from DNA Metabarcoding in Ships Arriving in Bay of Biscay" Water 12, no. 8: 2168. https://doi.org/10.3390/w12082168