Habitats and Diversity of Subterranean Macroscopic Freshwater Invertebrates: Main Gaps and Future Trends



1
Institute of Nature Conservation, Polish Academy of Sciences, 31-120 Kraków, Poland
2
Karst Research Institute, Research Centre of the Slovenian Academy of Sciences and Arts, 1000 Ljubljana, Slovenia
3
Department of Environmental Science, American University, Washington, DC 20016, USA
*
Author to whom correspondence should be addressed.
Water 2020, 12(8), 2170; https://doi.org/10.3390/w12082170
Submission received: 15 June 2020
/
Revised: 23 July 2020
/
Accepted: 24 July 2020
/
Published: 31 July 2020
(This article belongs to the Special Issue Freshwater Macroinvertebrates: Main Gaps and Future Trends)
Abstract
:Caves are the best studied aquatic subterranean habitat, but there is a wide variety of these habitats, ranging in depth below the surface and size of the spaces (pore or habitat size). Both factors are important in setting limits to species composition and richness. In addition to caves, among the most important shallow aquatic subterranean habitats are the hyporheal (underflow of rivers and streams), the hypotelminorheal (very superficial drainages with water exiting in seeps), epikarst, and calcrete aquifers. Although it is little studied, both body size and species composition in the different habitats is different. Because of high levels of endemism and difficulty in access, no subterranean habitats are well sampled, even caves. However, there are enough data for robust generalizations about some geographic patterns. Individual hotspot caves are concentrated in the Dinaric region of southern Europe, and overall, tropical regions have fewer obligate aquatic cave dwellers (stygobionts). In all subterranean aquatic habitats, regional diversity is much higher than local diversity, but local diversity (especially single cave diversity) may be a useful predictor of regional species richness. In Europe there is a ridge of high aquatic subterranean species richness basically extending east from the French–Spanish border. Its cause may be either high productivity or that long-term temperature oscillations are at a minimum. With increased collecting and analysis, global and continental trends should become clearer.
1. Introduction
The existence of eyeless and depigmented animals in the darkness of cave streams has been known since at least 1537 [1]. Of course, caves themselves have been known since at least the Paleolithic [2]. Groundwater also has a long history of human knowledge and use, and the first known well dates back to the Neolithic [3]. On the biological side, in 1907 Racovitza [4] mentioned studies from the 1890s on eyeless, depigmented species from artesian wells in Texas and wells in the Canterbury Plain of New Zealand.
It is probably Racovitza [4] who first pointed to the potential advantages of considering all terrestrial and freshwater aphotic habitats and their inhabitants together. Earlier, North American neo-Lamarckians had a strong interest in the evolution of eyelessness due to its connection to evolution by disuse (e.g., Packard [5]). While there was a general recognition of the unity of the subterranean domain, especially by European scientists, e.g., Ginet and Decou [6], it was not until the publication of the groundbreaking book “Groundwater Ecology” [7] that at least the aquatic side of subterranean biology became a distinct discipline with its own tenets.
In this review, we consider three areas. The first is the range of aquatic subterranean habitats and what unites and divides them. There have been two overarching classifications of freshwater subterranean habitats. One, due to Botosaneanu [8] who divided aquatic subterranean habitats (he included the marine interstitial) into two branches—“milieu perméables en grande” and “milieu perméables en petit”. He included more than 30 habitats, and the two most familiar are caves and the hyporheic, the underflow of streams and rivers. The other is due to Culver and Pipan [9,10] in which they attach equal, if not greater, weight to the vertical depth of the habitat compared to habitat size. We reassess these divisions and revisit the range of aquatic subterranean habitats.
The second area we review is the vexing question: how complete is our knowledge of the subterranean biota? In addition to the nearly universal shortfall of taxonomists to describe new species that have already been collected (the Linnean impediment), there are regions of the Earth’s surface that have been little sampled for aquatic subterranean fauna, especially in the tropics (the Wallacean impediment), and there are subterranean habitats that are poorly sampled due to sampling difficulties (the Racovitzan impediment [11]). All of these problems are especially severe in aquatic subterranean habitats because of the high levels of endemism of the species [12,13]. What can we say about overall fauna patterns in light of this uncertainty?
The third area we review is: what is the global pattern of richness of aquatic subterranean invertebrates? After reviewing the pattern of individual hotspot sites globally, we review some of the potential “fixes” to incomplete data, especially functions such as those of Chao [14] that estimate missing species, as well as assessing the potential impact of missing data (e.g., Zagmajster et al. [15]).
2. Aquatic Subterranean Habitats
Botosaneanu’s [8] compilation of subterranean habitats, produced as an addendum to his extensive review of the subterranean fauna is an appropriate place to begin (Table 1). We separated out from his dichotomy of “perméable en grand” and “perméable en petit” the categories of springs, which he included in “perméable en petit”. Springs are both habitats themselves (ecotones between surface and subterranean waters), but also convenient collecting points for subterranean species from more inaccessible (often unknown) subterranean habitats. These habitats may have fine or coarse sediments and may be shallow or deep.
Botosaneanu suggested only species limited to subterranean habitats be considered, but in practice it makes more sense to consider all species in springs (e.g., [16,17]), especially since not only species limited to subterranean habitats are blind and eyeless [18]. Springs themselves can be classified in a number of different ways, the oldest being based on discharge rate [19]. Other classifications, which can be quite elaborate (see Springer and Stevens [20]), are based on characteristics of the hydraulic head, geomorphologic structure, and water quality and temperature [21]. All of these are more elaborate than Botosaneanu’s [8] (Table 1). He listed springs as porous habitats [8] but we give them a separate category. Perhaps the reason that speleobiologists have not taken up more elaborate subdivisions of springs is that the fauna of springs often has few, if any, species showing the characteristics of subterranean life such as reduced eyes and pigment (see Botosaneanu [22]). However, this is not always the case. Dumnicka and Galas [23] show that a significant fraction of the subterranean fauna of Poland can be found in springs. Certainly, the classification of springs from the point of view of the groundwater fauna needs more attention. For example, Dumnicka et al. [17] show that substrate in the spring has a major effect on faunal composition.
Botosaneanu’s [8] division of cave habitats vertically (percolating (epikarst), vadose, epiphreatic, phreatic) follows the convention of hydrogeology. A similar classification, based on Leruth [28], was used by Howarth and Moldovan [29]. They identified five aquatic cave habitats:
- Highly dynamic flowing waters (sinking streams);
- Slow-flowing waters and lakes;
- Gours or pools formed on flowstone;
- Small pools on clay or mud;
- Dripping or percolation water.
There can be more elaborate subdivisions. For example, Poulson [30] distinguishes several types of cave streams, such as shallow streams and moderately deep master shaft drain streams. While no doubt useful in the context of Mammoth Cave, where he worked, their generality seems very limited. Each of the cave subdivisions has its own strengths and weaknesses, and it seems counterproductive to a priori decide how detailed such a subdivision should be.
Small pore habitats, categorized by Botosaneanu [8] (Table 1), were largely divided on the basis of water flow and salinity. An interesting feature of the classification is the hypotelminorheal—a persistent wet spot, a kind of perched aquifer, fed by subsurface water in a slight depression in an area of low to moderate slope, rich in organic matter, underlain by an impermeable clay layer typically less than 50 cm below the surface—see Culver and Pipan [9] and Meštrov [31]. It appears in the classification both as a porous habitat and as a spring (called seepage springs by Keany et al. [32]). If more were known about the drainage area of all springs, this duality of classification should occur for all spring types. The fauna collected at seepage springs seems to be a mixture of species primarily found in the hypotelminorheal and species primarily found in the seepage spring itself [33].
Just as caves dominate large pore (diameter) habitats, the hyporheal dominates small pore (diameter) habitats, at least in terms of the amount of research done. Orghidan [34], (see Käser [35] for an English translation) coined the term and defined it as the zone of interstitial spaces constituted by the sediments of the stream bed. To our knowledge there have not been formally proposed subdivisions but even at the scale of a few meters there are differences, especially due to upwelling and downwelling [36]. In different streams, the connections between the hyporheic, the groundwater zone, and underlying impermeable substratum vary and have a profound effect on the composition and abundance of the fauna. Malard et al. [36] list five cases but do not formally name them:
- No hyporheic zone.
- Hyporheic zone created only by advected channel water (no ground water).
- Hyporheic zone created by advection by both channel water and ground water.
- Hyporheic zone created only by infiltration of channel water beneath the stream bed (no parafluvial flow).
- A perched hyporheic zone created only by infiltration of channel water beneath the stream bed.
Their fifth category would appear to include seepage springs (outlet of the hypotelminorheic).
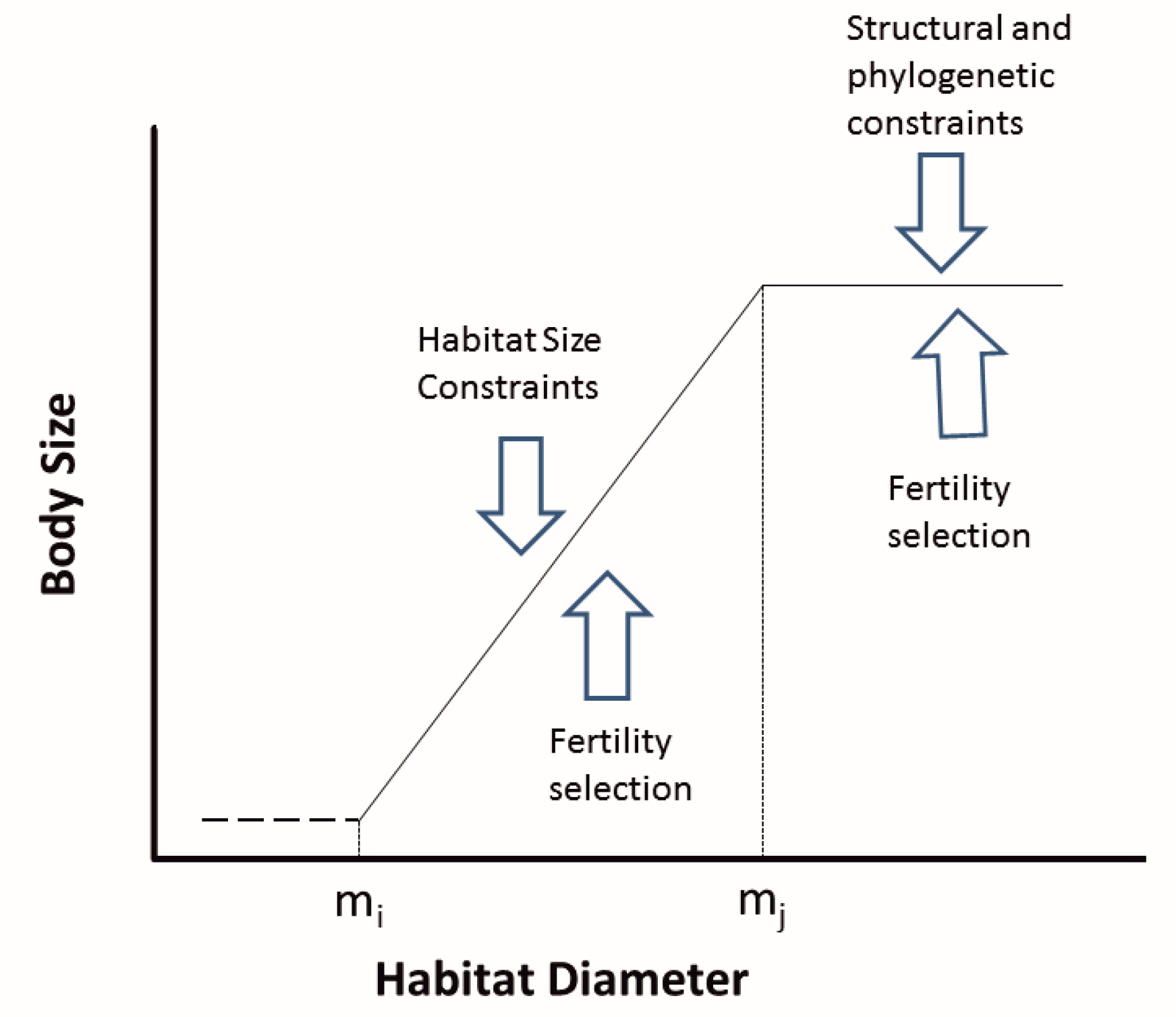
As knowledge of subterranean habitats has grown, the dichotomy between large and small pore habitats has grown increasingly problematic. Habitats such as calcrete aquifers and the hypotelminorheic are likely intermediate in pore size, at least based on the size of the organisms found in these habitats. Although epikarst and percolating water are part of karst, they are small pore habitats, based on the size of the inhabitants [10]. There is accumulating evidence [24] that the size of the habitat spaces has an impact on body size. Their model of how this works is shown in Figure 1 and the overall pattern in relationship to habitat categories is shown in Figure 2.
Given these size differences in both habitat and organism, one would expect the community composition in different habitats to be quite different. Dumnicka et al. [37], basing on 280 records for interstitial habitat, 150 for cave waters and 50 for wells, looked at this question for the Polish groundwater annelids fauna. Somewhat surprisingly, relatively little separation of habitats was found, although there was a tendency for the interstitial fauna to differ from the cave fauna along the first axis of their correspondence analysis (Figure 3) (Supplementary Materials). The well fauna, collected in various kinds of wells, overlapped broadly with both. Hahn and Fuchs [38] found a similar pattern for the German fauna. It is curious that there have not been more analyses of this question, and it is worth pursuing.
An interesting question is how many species are found in the different subterranean habitats in a region. In Poland, which was largely glaciated, Dumnicka and Galas [23] demonstrate that there are more records per sample in interstitial waters and wells than in caves. In fact, there are more records per sample in springs than in caves (Figure 4). As far as we can determine, these are the only data of this type available. It would be interesting to compare the Polish data with data from an unglaciated area, areas which generally have a much richer cave fauna.
Culver and Pipan [10,39], Pipan and Culver [18] and Blatnik et al. [40] suggested that there was a third major category of subterranean habitats, close to the surface and with intermediate-sized habitat spaces—shallow subterranean habitats (see Table 1). In their book-length treatment of the topic, Culver and Pipan [10] emphasized the vertical division—the distance from the surface and defined shallow subterranean habitats as occurring less than 10 m from the surface. They suggested several unifying features:
- Absence of light;
- Close surface-subsurface connections (except for calcrete aquifers);
- Availability of organic carbon and nutrients;
- Generally small habitat (pore) size.
Habitats include the hypotelminorheal, epikarst, hyporheal, and calcrete aquifers. Halse [25] calls calcrete aquifers deep subterranean habitats but gives no criteria for this choice. Calcretes in the Pilbara region are deeper than 10 m [13] while those in the Yilgarn are often less than 10 m [41]. Of course, all caves with natural entrances occur in part at depths of less than 10 m. Culver and Pipan [10] included only habitats that were typically less than 10 m in depth. Whether calcretes are included or not, the grouping of habitats into shallow versus deep has proven to be frequently used, although it remains to be seen how useful the distinction is.
3. The Struggle to Measure Aquatic Subterranean Biodiversity
Aquatic subterranean biodiversity surveys share with most other invertebrate surveys both a Linnean shortfall (not enough taxonomists) and a Wallacean shortfall (not all areas sampled). However, there are a number of taxonomists, especially in Europe, actively working on major subterranean groups, including amphipods, copepods, isopods, and oligochaetes. With respect to the Wallacean shortfall, tropical caves, if not other subterranean habitats, are being increasingly well sampled (e.g., [42,43,44]).
However, there are a number of aspects of the subterranean fauna that make unbiased sampling very difficult. Nearly all the analyses and datasets are based on those species limited to subterranean habitats—stygobionts for aquatic habitats. However, in most subterranean habitats there are species that show little morphological modification for subterranean life and/or are also found in surface habitats [18]. Such stygophiles are part of the subterranean community and reproduce in subterranean habitats. Analysis of stygobionts is an analysis of the highly evolved component of subterranean communities specialized and limited to subterranean habitats, not the entire subterranean community.
A second issue is the omnipresence of undescribed species. Some of these species that have been collected are awaiting taxonomic analysis, some have been observed but not collected, and some have not even been observed but are considered likely to be present in a region by expert taxonomists. Obviously the last two are fraught with the likelihood of overestimation or exaggeration of numbers [45]. Bolded letters spell out PASCALIS and it is usually written this way. Collected, undescribed species present a special problem. If they are ignored, then some regions, like Brazil—see Trajano and Bichuette [46]—will appear to be species-poor when in fact they are species rich. One possible solution to the problem is to apply a “discount rate”. For example, Culver et al. [45] report that of 19 species listed as undescribed by Holsinger et al. [47] in their enumeration of the West Virginia cave fauna, six turned out to be previously described species. This results in a discount rate of 0.68. The omnipresence of undescribed species can also lead to inappropriate comparisons between regions based on data taken from different time frames—what Culver et al. [45] term the fallacy of provincialism.
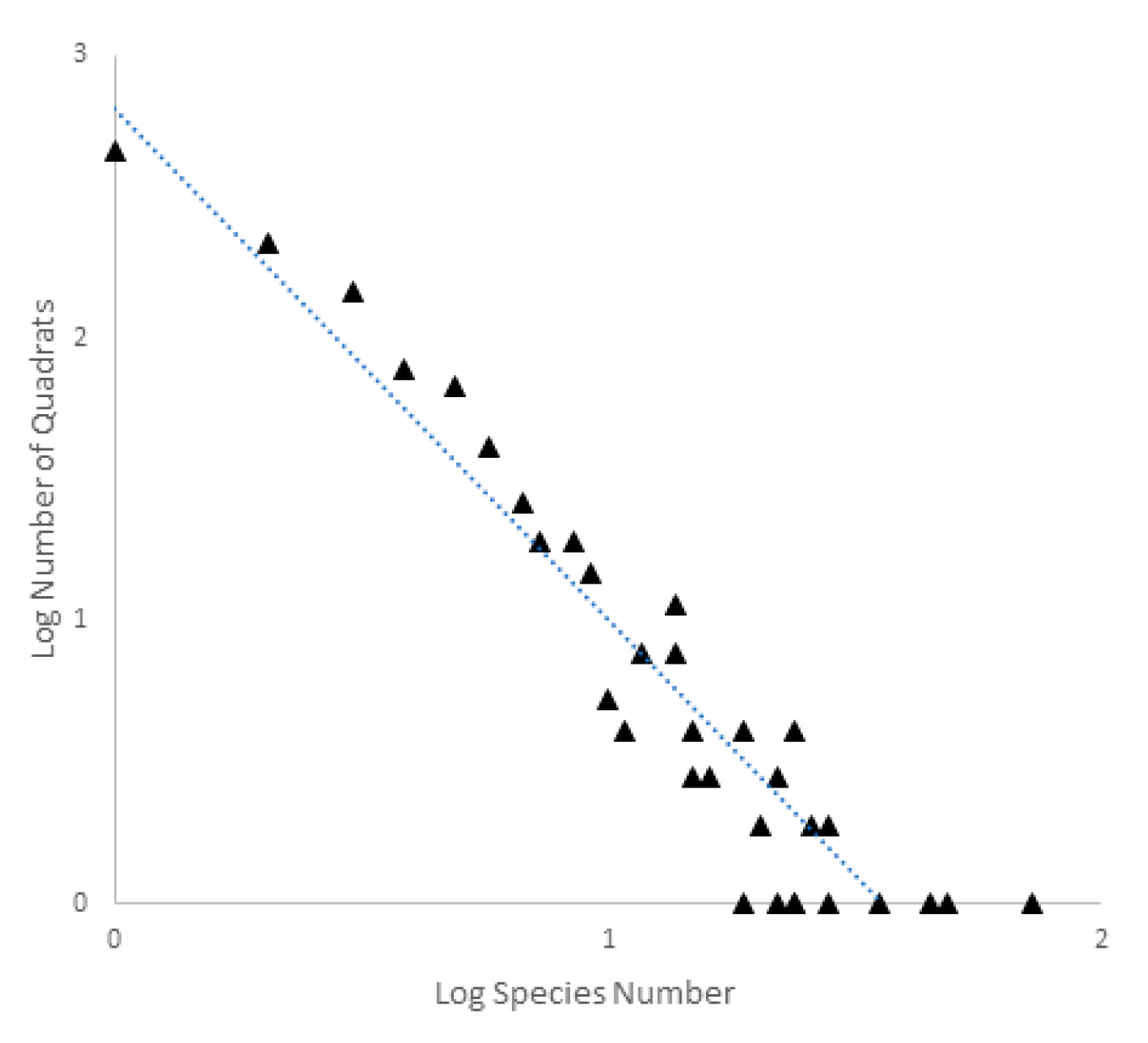
A third and related issue is the high frequency of local endemism, often single site endemism. An example of this is from the large-scale European study of subterranean biodiversity, PASCALIS (Protocols for the Assessment and Conservation of Aquatic Life in the Subsurface) [48]. In this study of all known stygobionts from six European countries, there was a strong negative relationship between number of species and number of 0.2° × 0.2° grid cells occupied by a species (Figure 5). Of the 930 described stygobionts, 396 were known from a single grid cell.
This means that unless all cells are sampled, many single cell endemics will be missed. In practice it appears that the pattern of species richness is unaffected if single cell and other narrow endemics are not included [50]. More generally, missing species do not seem to affect geographic pattern. Deharveng et al. [49] measured these missing species using a jackknife procedure that resamples and takes into account the number of narrow endemics. Their results, by country, are shown in Table 2.
The final impediment is the Racovitzan impediment, the incompleteness of sampling of subterranean habitats [11]. Caves are incompletely sampled because new caves are constantly being discovered and the number of known caves is very large (e.g., more than 10,000 in Slovenia). While sampling all caves is neither possible nor necessary, the high levels of endemism makes thorough sampling important. Nevertheless, there are some very large datasets for subterranean animals. Probably the largest is that of Zagmajster et al. [51], who assembled data for 21,000 occurrences of 1570 European aquatic subterranean species. The situation for non-cave aquatic subterranean species is particularly difficult because neither epikarst, the hyporheal, or the hypotelminorheal can be sampled easily. For the hyporheal, most samples are taken by pumping water out of the habitat through a fine mesh filter, and then sorting the samples. The Bou–Rouch pump [52,53] in many ways made the sampling of hyporheal possible. Prior to the development of a continuous filtering device [54], epikarst could only be sampled very indirectly by sampling drip pools, themselves biased samples of the organisms in dripping water [55]. The situation is even more dire for the hypotelminorheal. No good sampling device exists, and the habitat must be destructively sampled. However, Niemiller et al. [56] were able to show that eDNA of seep amphipods (genus Stygobromus) could be detected, allowing for the possibility of non-destructive sampling.
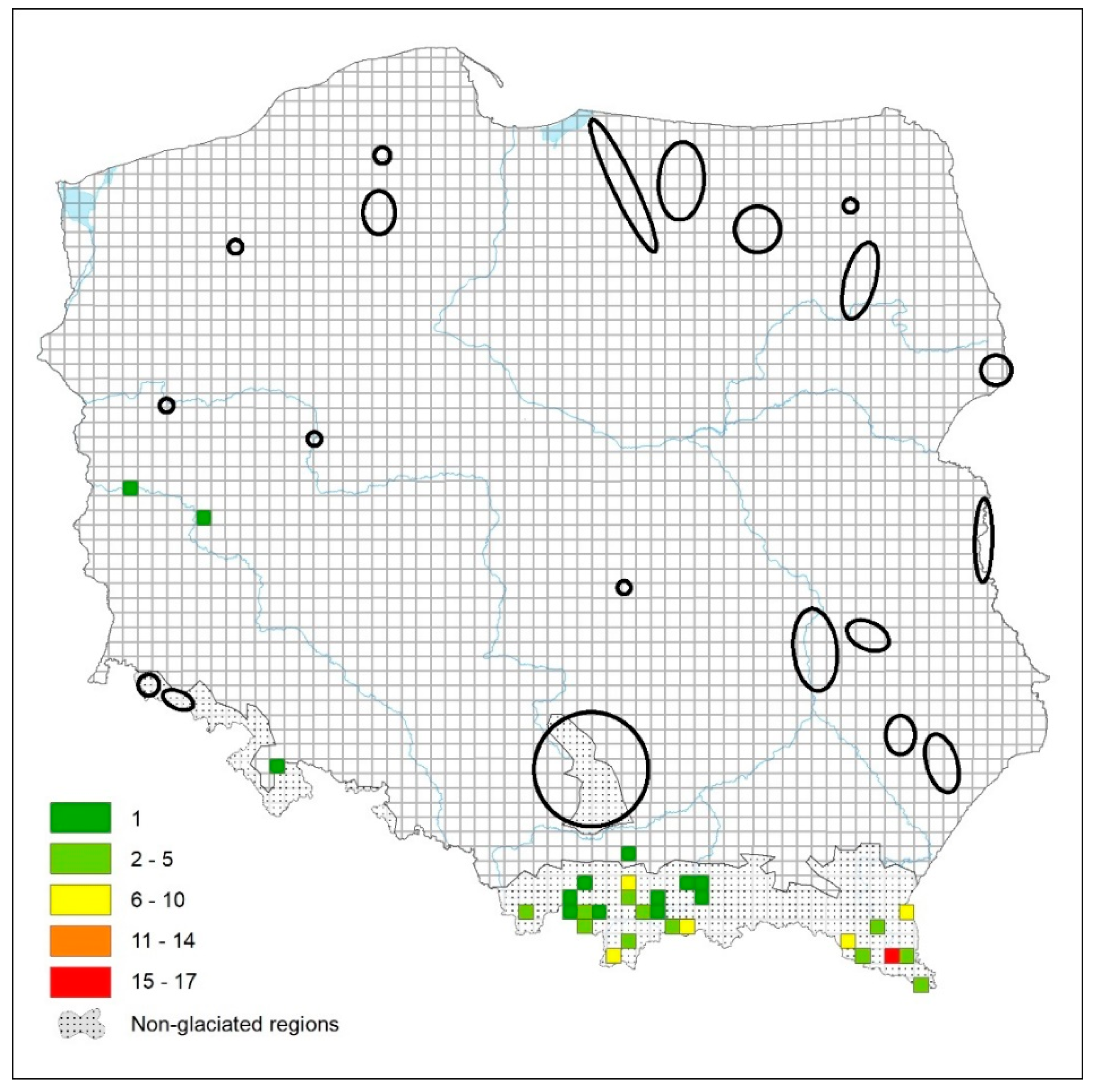
There is one way that at least the extent of undersampling can be better understood and that is to report on sites where no specimens were found. There has been a reluctance to do this, perhaps based on the sense that empty samples are failed samples. However, such reporting can be extremely informative. Dumnicka et al. [57] report those quadrats in Poland where they failed to find any water mites specialized for subterranean life, and almost all of these sites were in the glaciated areas of Poland, where the specialized subterranean fauna should be rare (Figure 6).
4. Geographic Patterns of Species Richness in Aquatic Subterranean Invertebrates
The easiest, but certainly not the best, measure of species richness is the species richness at a single subterranean site, α-diversity. It is an inadequate measure because α- diversity is a small fraction of the species richness of a region, γ-diversity [58]. However, the data are much easier to accumulate since the regional analysis requires data on many sites [59]. Culver and Sket [60] published the first list, which included 20 caves and wells with 20 or more obligate subterranean species, including both aquatic and terrestrial species. Since then, the number of such sites has at least doubled, and most recently Culver and Pipan [61] published a list of sites with 25 or more stygobiotic species (Table 3).
Several points emerge. First, with the exception of the Robe River in western Australia, none of the sites are in the tropics or sub-tropics. Second, there is a concentration of hotspots in the Dinaric karst, with four of ten sites in this region.
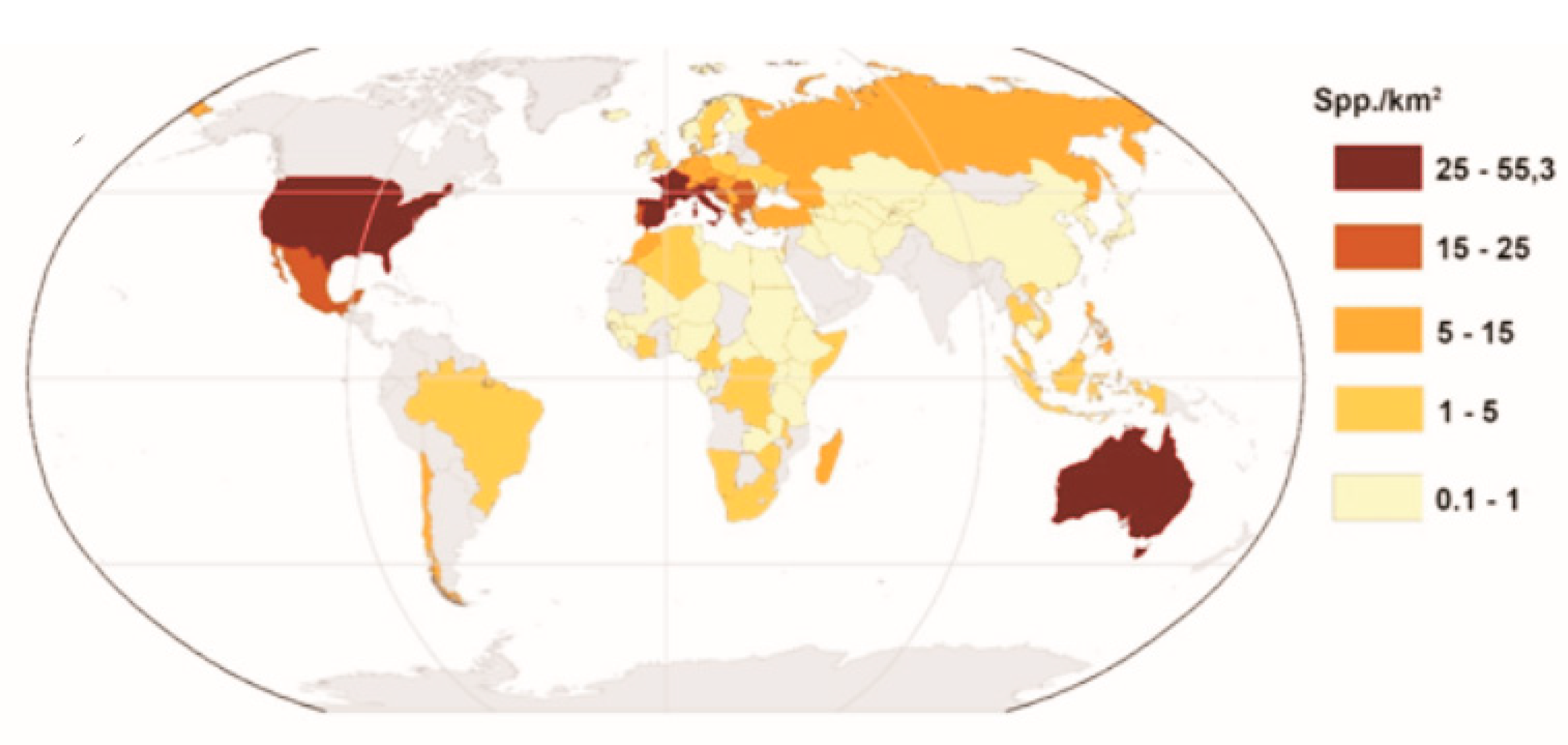
Counts of numbers of species by country (corrected for size) give a similar picture to Table 3. There is a broad band of high species richness in the north temperate zone (China is relatively unknown) as well as in Australia (Figure 7), where aquatic subterranean diversity is largely found in calcrete aquifers, accessible only by wells [25,26].
Calcrete aquifers are carbonate deposits that form in the vicinity of the water table as a result of evaporation of groundwater—see Culver and Pipan [10]. Aquatic subterranean species richness is very high in both the Yilgarn and Pilbara aquifers of western Australia, but it remains difficult to compare with other areas because of the distinctive and different way researchers have measured species richness in Australia [45,63].
More detailed analyses at the global scale are not available, but there is more detailed information available for Europe [48]. Zagmajster et al. [51] showed that there is a ridge of high species richness in southern Europe (Figure 8), one that corresponds to a similar ridge in terrestrial species richness [59]. The explanation for this ridge may be that it corresponds to a ridge of primary productivity or that long term temperature oscillations are at a minimum.
The above analyses of the patterns of aquatic subterranean species richness is an analysis of only the specialists for cave life. e.g., stygobionts. Many cave streams, especially in glaciated areas, have functioning, reproducing communities but without any specialized species. A good example of this is the aquatic fauna in Swildon’s Hole, a nearly 10 km long cave in the Mendip Hills of the United Kingdom. Knight [64] found 38 taxa in the main stream, mainly Trichoptera and Diptera. There were no stygobionts, yet a functioning community. The situation in the tributaries was different where three of ten species were specialized for subterranean life.
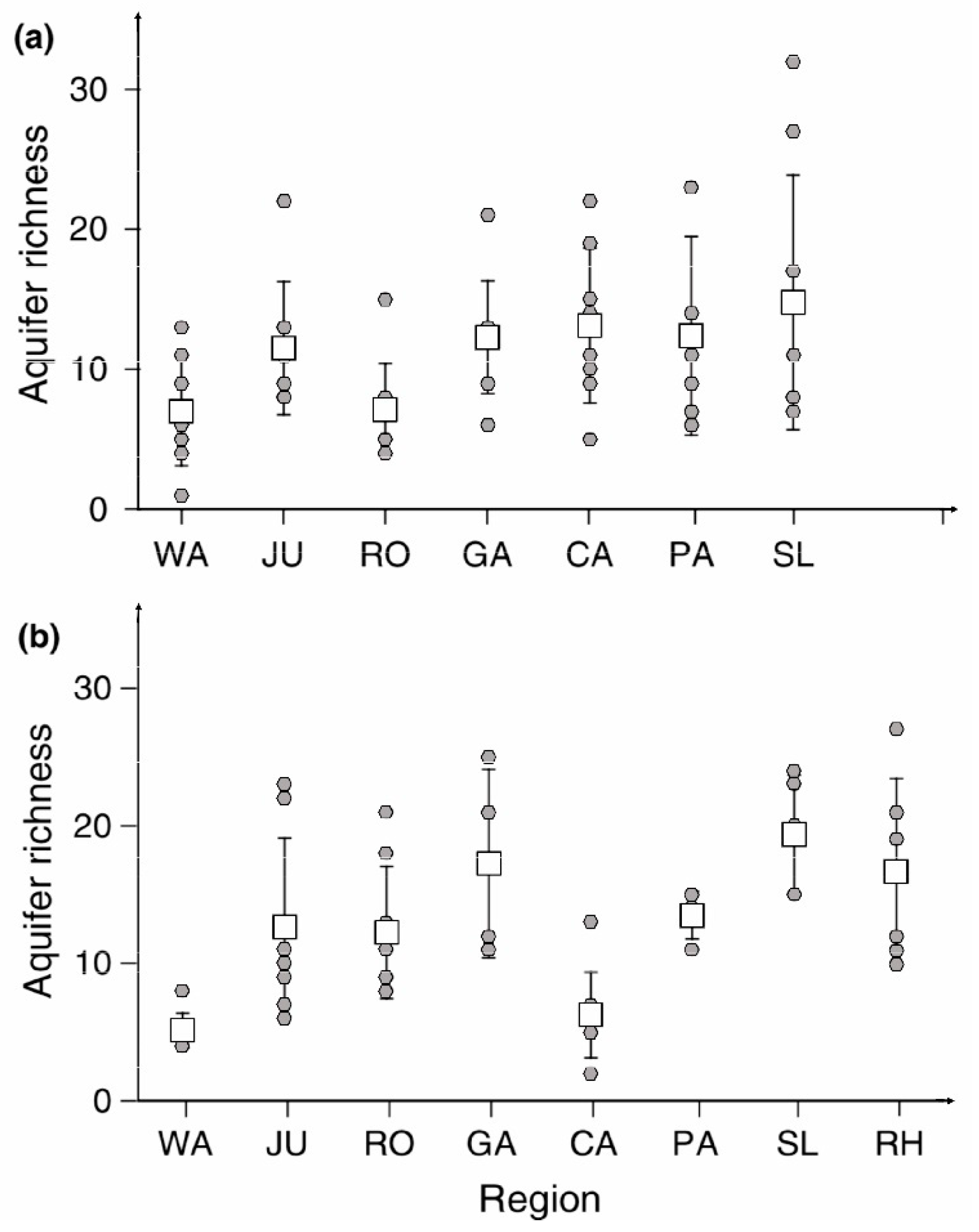
Very little is known about the geographic pattern of species richness in non-cave aquatic subterranean habitats. The PASCALIS project [48] included extensive sampling of the hyporheal, but only eight relatively small regions were included in the study (Figure 8): Wallonia (Belgium), Jura (France), Rousillon (France), Cantabria (Spain), Padano–Alpine region (Italy), Slovenia, Rhône valley (France), and Garonne (France). Malard et al. [58] showed that species richness was highest in Slovenia, followed by the Rhone and the Garonne (Figure 9). Slovenia and the Garonne are on the ridge of aquatic species richness (Figure 8). One of the features of porous aquifers in general and hyporheal in particular is its fine scale heterogeneity, which is evident in Figure 9.
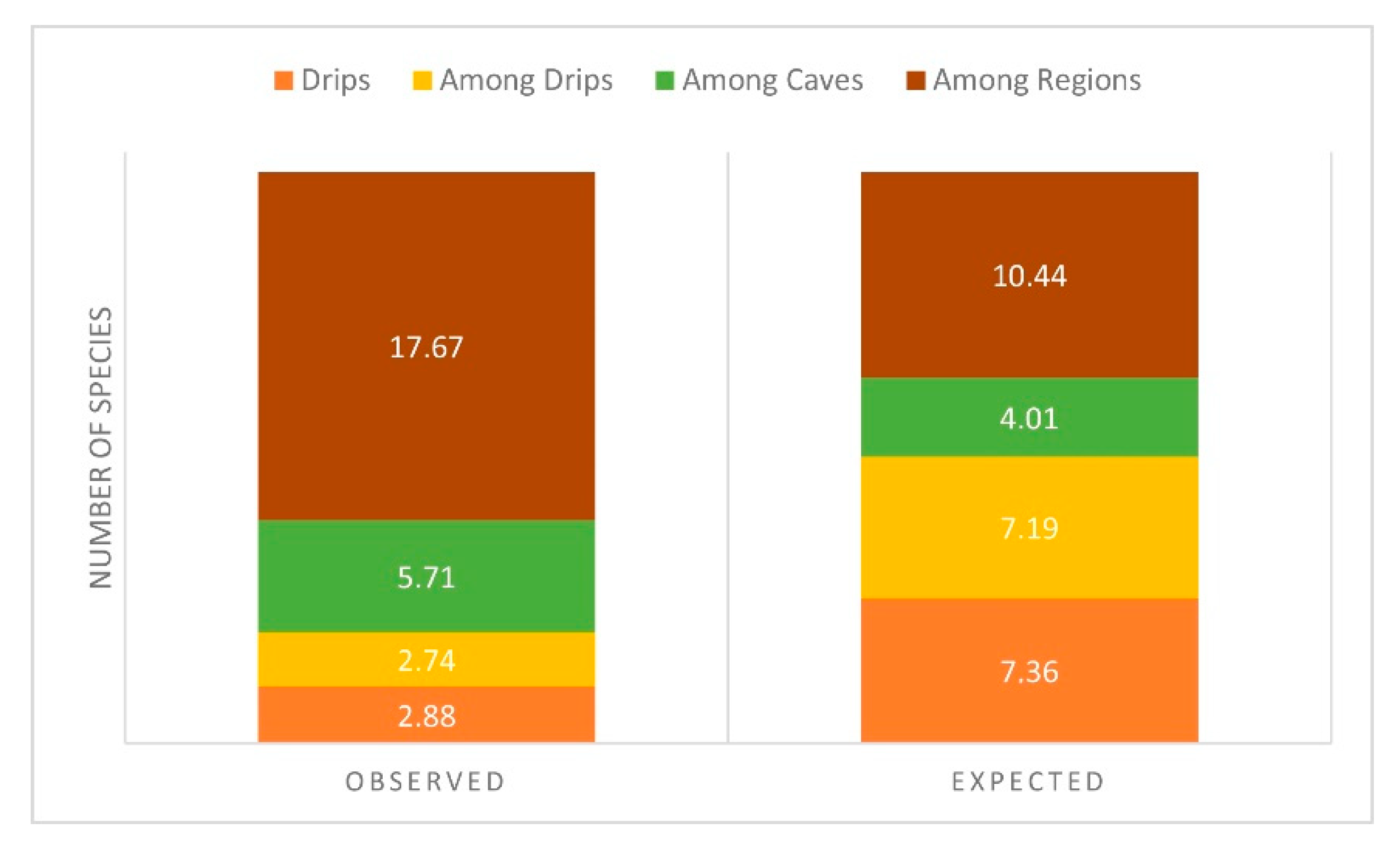
Pipan et al. [65] did a small regional study of epikarst copepods in Slovenia. They analyzed 81 drips from13 caves in three karst areas of Slovenia. As with Malard et al.’s analysis, they found small scale differences and that α diversity (within drip species richness) was small compared to β diversity. Of the 18 species, only three were accounted for by α diversity, three by differences within a cave, six by differences between caves in a region, and 18 by differences among regions (Figure 10). When the data are viewed in another way, one that emphasizes the occurrence of “hotspot” drips, a different pattern emerges.
A few drips contribute a disproportionate share of species diversity. The most species-rich drip in the Dinaric karst has 10 species and the entire Županova jama has 13 species, so this drip contributes 40 percent of the species diversity known from the entire Dinaric karst! The task of assessing epikarst species diversity would be considerably simplified if we had a method of determining which drips were hotspots prior to sampling, but we do not.
5. Conclusions
Obligate aquatic subterranean organisms (stygobionts) occur in a variety of subterranean habitats that vary both in depth and in pore size. This fauna is relatively well known in Europe and North America, and poorly known elsewhere. Sampling this fauna, particularly the non-cave fauna remains difficult, resulting in a Racovitzan shortfall for these habitats. Species richness is highest in mid-temperate latitudes and the Dinaric karst in southern Europe is a hotspot of stygobiotic species richness. Endemism is high and consequently β diversity is much higher than α diversity.
With increased amounts of data and new analytical tools, both continental and global patterns of species richness, and its explanation, should become clearer.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4441/12/8/2170/s1, Table S1: Full names of species cited in Figure 3 and the text.
Author Contributions
Conceptualization, E.D.; methodology, D.C.C., E.D., T.P.; validation, D.C.C., E.D., T.P.; formal analysis, D.C.C., E.D., T.P.; investigation, D.C.C., E.D., T.P.; resources, E.D.; data curation, D.C.C., E.D., T.P.; writing—original draft preparation, D.C.C.; writing—review and editing, D.C.C., E.D., T.P.; visualization, E.D., T.P.; supervision, E.D.; project administration, D.C.C., E.D., T.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Romero, A. Cave Biology: Life in Darkness; Cambridge University Press: Cambridge, UK, 2009; ISBN 978-0-521-82846-8. [Google Scholar]
- Shaw, T. History of Cave Science: The Exploration and Study of Limestone Caves, to 1900, 2nd ed.; Sydney Speleological Society: Sydney, Australia, 1992; ISBN 0-646-12503-6. [Google Scholar]
- Tegel, W.; Elburg, R.; Hakelberg, D.; Stäuble, H.; Büntgen, U. Early Neolithic water wells reveal the world’s oldest wood architecture. PLoS ONE 2012, 7, e51374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Racovitza, E.G. Essai sur les problems biospéologiques. Arch. Zool. Exp. Gén. IV 1907, 6, 371–488. [Google Scholar]
- Packard, A.S. The cave fauna of North America, with remarks on the anatomy of brain and the origin of the blind species. Mem. Nat. Acad. Sci. USA 1888, 4, 1–156. [Google Scholar]
- Ginet, R.; Decou, V. Initiation à la Biologie et à L’écologie Souterraines; Delarge, J-P.: Paris, France, 1977; ISBN 2-7113-0073-0. [Google Scholar]
- Gibert, J.; Danielopol, D.L.; Stanford, J.A. (Eds.) Groundwater Ecology; Academic Press: San Diego, CA, USA, 1994; ISBN 978-0-08-050762-0. [Google Scholar]
- Botosaneanu, L. (Ed.) Stygofauna Mundi; EJ Brill: Leiden, The Netherlands, 1986; ISBN 90-04-075712. [Google Scholar]
- Culver, D.C.; Pipan, T. Redefining the extent of the aquatic subterranean biotope-shallow subterranean habitats. Ecohydrolgy 2011, 4, 721–730. [Google Scholar] [CrossRef]
- Culver, D.C.; Pipan, T. Shallow Subterranean Habitats. Ecology, Evolution, and Conservation; Oxford University Press: Oxford, UK, 2014; ISBN 978-0-19-964617-3. [Google Scholar]
- Ficetola, G.F.; Canedoli, C.; Stoch, F. The Racovitzan impediment and the hidden biodiversity of unexplored environments. Conserv. Biol. 2019, 33, 214–216. [Google Scholar] [CrossRef]
- Trontelj, P.; Douady, C.J.; Fišer, C.; Gibert, J.; Gorički, Š.; Lefebure, T.B.; Zakšek, V. A molecular test for cryptic diversity in ground water: How large are the ranges of macro-stygobionts? Freshw. Biol. 2009, 54, 727–744. [Google Scholar] [CrossRef]
- Eberhard, S.M.; Halse, S.A.; Williams, M.R.; Scanlon, M.D.; Cocking, J.; Barron, H.J. Exploring the relationship between sampling efficiency and short-range endemism for groundwater fauna in the Pilbara region. Freshw. Biol. 2009, 54, 885–901. [Google Scholar] [CrossRef]
- Chao, A. Estimating the population size for capture-recapture data with unequal catchability. Biometrics 1987, 43, 783–791. [Google Scholar] [CrossRef]
- Zagmajster, M.; Culver, D.C.; Christman, M.C.; Sket, B. Evaluating the sampling bias in pattern of subterranean species richness: Combining approaches. Biodivers. Conserv. 2010, 19, 3035–3048. [Google Scholar] [CrossRef]
- Dumnicka, E. Stygofauna associated with springfauna in southern Poland. Subterr. Biol. 2005, 3, 29–36. [Google Scholar]
- Dumnicka, E.; Galas, J.; Koperski, P. Benthic invertebrates in karst springs: Does substratum or location define communities? Int. Rev. Hydrobiol. 2007, 92, 452–464. [Google Scholar] [CrossRef]
- Pipan, T.; Culver, D.C. Convergence and divergence in the subterranean realm: A reassessment. Biol. J. Linn. Soc. 2012, 107, 1–14. [Google Scholar] [CrossRef]
- Meinzer, O.E. The Occurrence of Ground Water in the United States with a Discussion of Principles; US 89 Survey Water-Supply Paper No. 489; U.S. Department of the Interior: Washington, DC, USA, 1923. [CrossRef]
- Springer, A.E.; Stevens, L.E. Spheres of discharge of springs. Hydrogeol. J. 2009, 17, 83–93. [Google Scholar] [CrossRef]
- Kresic, N. Types and classifications of springs. In Groundwater Hydrology of Springs: Engineering, Theory, Management, and Sustainability; Kresic, N., Stevanovic, Z., Eds.; Elsevier: Amsterdam, The Netherlands, 2010; pp. 31–86. ISBN 978-1-85617-502-9. [Google Scholar]
- Botosaneanu, L. (Ed.) Studies in Crenobiology: The Biology of Springs and Springbrooks; Backhuys Publishers: Leiden, The Netherlands, 1998; ISBN 90-73348-04-8. [Google Scholar] [CrossRef]
- Dumnicka, E.; Galas, J. An overview of stygobiontic invertebrates of Poland based on published data. Subterr. Biol. 2017, 23, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Pipan, T.; Culver, D.C. The unity and diversity of the subterranean realm with respect to invertebrate body size. J. Cave Karst Stud. 2017, 79, 1–9. [Google Scholar] [CrossRef]
- Halse, S. Research in calcretes and other deep subterranean habitats outside of caves. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer Nature: Gland, Switzerland, 2018; pp. 415–434. ISBN 978-3-319-98850-4. [Google Scholar]
- Humphreys, W.F. Groundwater calcrete aquifers in the Australian arid zone: The context of an unfolding plethora of stygal biodiversity. Rec. West. Aust. Mus. 2001, 64, 63–83. [Google Scholar] [CrossRef] [Green Version]
- Pentecost, A. Travertine; Springer: Berlin/Heidelberg, Germany, 2005; ISBN 978-1-4020-3523-4. [Google Scholar]
- Leruth, R. La biologie du domaine souterrain et la faune cavernicole de la Belgique. Mém. Mus. Hist. Nat. Belgique 1939, 87, 1–506. [Google Scholar]
- Howarth, F.G.; Moldovan, O.T. Where cave animals live. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer Nature: Gland, Switzerland, 2018; pp. 41–68. ISBN 978-3-319-98850-4. [Google Scholar]
- Poulson, T.L. The Mammoth Cave ecosystem. In The Natural History of Biospeleology; Camacho, A.I., Ed.; Museo Nactional de Ciencias Naturales: Madrid, Spain, 1992; pp. 569–612. ISBN 84-00-07280-4. [Google Scholar]
- Meštrov, M. Un nouveau milieu aquatique souterrain: Le biotope hypotelminorheique. C. R. Acad. Sci. 1962, 254, 2677–2679. [Google Scholar]
- Keany, J.M.; Christman, M.C.; Milton, M.; Knee, K.L.; Gilbert, H.; Culver, D.C. Distribution and structure of shallow subterranean aquatic arthropod communities in the parklands of Washington, D.C. Ecohydrolgy 2018, 12, e2044. [Google Scholar] [CrossRef]
- Culver, D.C.; Trontelj, P.; Zagmajster, M.; Pipan, T. Paving the way for standardized and comparable subterranean biodiversity studies. Subterr. Biol. 2012, 10, 43–50. [Google Scholar] [CrossRef]
- Orghidan, T. Un nou domeniu de viata acvatica subterana: “Biotopul hiporeic”. Buletin Stiintific Sectia de Biologie si Stiinte Agricole si Sectia de Geologie si Geografie 1955, 7, 657–676. [Google Scholar]
- Käser, D.H. A new habitat of subsurface waters: The hyporheic biotope, by Traian Orghidan (1959). Fundam. Appl. Limnol. 2010, 176, 291–302. [Google Scholar]
- Malard, F.; Tockner, K.; Dole-Oliver, M.-J.; Ward, J.V. A landscape perspective of surface-subsurface hydrological exchanges in river corridors. Freshw. Biol. 2002, 47, 621–640. [Google Scholar] [CrossRef]
- Dumnicka, E.; Galas, J.; Krodkiewska, M.; Pociecha, A. The diversity of annelids in subterranean waters: A case study from Poland. Knowl. Manag. Aquat. Ecosyst. 2020, 421, 16. [Google Scholar] [CrossRef]
- Hahn, H.J.; Fuchs, A. Distribution patterns of groundwater communities across aquifer types in south-western Germany. Freshw. Biol. 2009, 54, 848–860. [Google Scholar] [CrossRef]
- Culver, D.C.; Pipan, T. Superficial subterranean habitats-gateway to the subterranean realm? Cave Karst Sci. 2008, 35, 5–12. [Google Scholar]
- Blatnik, M.; Culver, D.C.; Gabrovšek, F.; Knez, M.; Kogovšek, B.; Kogovšek, J.; Liu, H.; Mayaud, C.; Mihevc, A.; Mulec, J.; et al. Changing perspectives on subterranean habitats. In Karstology in the Classical Karst; Knez, M., Otoničar, B., Petrič, M., Pipan, T., Slabe, T., Eds.; Springer: Cham, Switzerland, 2020; pp. 183–205. ISBN 978-3-030-26827-5. [Google Scholar]
- Bradford, T.; Adams, M.; Humphreys, W.F.; Austin, A.D.; Cooper, S.J.B. DNA barcoding of stygofauna uncovers cryptic amphipod diversity in a calcrete aquifer in Western Australia’s arid zone. Mol. Ecol. Resour. 2010, 10, 41–50. [Google Scholar] [CrossRef]
- Brancelj, A.; Boonyanusith, C.; Watiroyram, S.; Senamuang, L.O. The groundwater-dwelling fauna of Southeast Asia. J. Limnol. 2013, 72, 327–344. [Google Scholar] [CrossRef] [Green Version]
- Nola, M.; Togouet, S.H.Z.; Marmonier, P.; Kayo, R.T.; Piscart, C. An annotated checklist of freshwater stygobiotic crustaceans of Africa and Madagascar. Crustaceana 2012, 85, 1613–1631. [Google Scholar] [CrossRef]
- Ríos-Escalante, P.; Parra-Coloma, L.; Peralta, M.A.; Pérez-Schultheiss, J.; Rudolph, E.H. A checklist of subterranean water crustaceans from Chile (South America). Proc. Biol. Soc. Wash. 2016, 129, 114–128. [Google Scholar] [CrossRef] [Green Version]
- Culver, D.C.; Holsinger, J.R.; Feller, D.J. The fauna of seepage springs and other shallow subterranean habitats in the mid-Atlantic Piedmont and Coastal Plain, U.S.A. Northeast. Nat. 2012, 19, 1–42. [Google Scholar] [CrossRef]
- Trajano, E.; Bichuette, M.E. Diversity of Brazilian subterranean invertebrates, with a list of troglomorphic taxa. Subter. Biol. 2010, 7, 1–16. [Google Scholar]
- Holsinger, J.R.; Baroody, R.A.; Culver, D.C. The Invertebrate Cave Fauna of West Virginia; Bulletin 7 WV Speleological Survey; West Virginia Speleological Survey: Barrackville, WV, USA, 1976. [Google Scholar]
- Gibert, J. Protocols for the assessment and conservation of aquatic life in the subsurface (PASCALIS), a European project. In Mapping Subterranean Biodiversity/Cartographie de la Biodiversité Souterrain; Culver, D.C., Deharveng, L., Gibert, J., Sasowsky, I., Eds.; Karst Waters Institute Special Publication 6; Karst Waters Institute: Charles Town, WV, USA, 2001; pp. 19–21. ISBN 0-9640258-5-x. [Google Scholar]
- Deharveng, L.; Stoch, F.; Gibert, J.; Bedos, A.; Galassi, D.; Zagmajster, M.; Brancelj, A.; Camacho, A.; Fiers, F.; Martin, P.; et al. Groundwater biodiversity in Europe. Freshw. Biol. 2009, 54, 709–726. [Google Scholar] [CrossRef]
- Eme, D.; Zagmajster, M.; Delić, T.; Fišer, C.; Flot, J.-F.; Konecny-Dupré, L.; Pálsson, S.; Stoch, F.; Zakšek, V.; Douady, C.J.; et al. Do cryptic species matter in macroecology? Sequencing European groundwater crustaceans yields smaller ranges but does not challenge biodiversity determinants. Ecography 2017, 40, 424–436. [Google Scholar] [CrossRef] [Green Version]
- Zagmajster, M.; Eme, D.; Fišer, C.; Galassi, D.; Marmonier, P.; Stoch, F.; Cornu, J.-F.; Malard, F. Geographic variation in range size and beta diversity of groundwater crustaceans: Inputs from habitats with low thermal seasonality. Glob. Ecol. Biogeogr. 2014, 23, 1135–1145. [Google Scholar] [CrossRef]
- Bou, C.; Rouch, R. Un nouveau champ de recherches sur la faune aquatiques souterraine. C. R. Acad. Sci. Paris 1967, 265, 369–370. [Google Scholar]
- Malard, F.; Dole-Olivier, M.-J.; Mathieu, J.; Stoch, F. Sampling Manual for the Assessment of Regional Groundwater Biodiversity; European Project PASCALIS: Lyon, France, 2002. [Google Scholar]
- Pipan, T. Epikarst-A Promising Habitat; Založba, ZRC: Ljlubljana, Slovenia, 2005; ISBN 961-6500-90-2. [Google Scholar]
- Pipan, T.; Holt, N.; Culver, D.C. How to protect a diverse, poorly known, inaccessible fauna: Identification and protection of source and sink habitats in the epikarst. Aquat. Conserv. Mar. Freshw. Ecosyst. 2010, 20, 748–755. [Google Scholar] [CrossRef]
- Niemiller, M.L.; Porter, M.L.; Keany, J.; Gilbert, H.; Culver, D.C.; Fong, D.W.; Hobson, C.S.; Kendall, K.D.; Taylor, S.J. Evaluation of eDNA for groundwater invertebrate detection and monitoring: A case study with endangered Stygobromus (Amphipoda: Crangonyctidae). Conserv. Genet. Resour. 2017, 10, 247–257. [Google Scholar] [CrossRef]
- Dumnicka, E.; Galas, J.; Najberek, K.; Urban, J. The influence of Pleistocene glaciations on the distribution of obligate aquatic subterranean invertebrate fauna in Poland. Zool. Anz. 2020, 286, 90–99. [Google Scholar] [CrossRef]
- Malard, F.; Boutin, C.; Camacho, A.I.; Ferreira, D.; Michel, G.; Sket, B.; Stoch, F. Diversity patterns of stygobiotic crustaceans across multiple spatial scales in Europe. Freshw. Biol. 2009, 54, 756–776. [Google Scholar] [CrossRef]
- Culver, D.C.; Deharveng, L.; Bedos, A.; Lewis, J.J.; Madden, M.; Reddell, J.R.; Sket, B.; Trontelj, P.; White, W. The mid-latitude biodiversity ridge in terrestrial cave fauna. Ecography 2006, 29, 120–128. [Google Scholar] [CrossRef]
- Culver, D.C.; Sket, B. Hotspots of subterranean biodiversity in caves and wells. J. Cave Karst Stud. 2000, 62, 11–17. [Google Scholar]
- Culver, D.C.; Pipan, T. The Biology of Caves and Other Subterranean Habitats; Oxford University Press: Oxford, UK, 2019; ISBN 978-0-19-882076-5. [Google Scholar]
- Zagmajster, M.; Malard, F.; Eme, D.; Culver, D.C. Subterranean biodiversity patterns from global to regional scales. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer Nature: Gland, Switzerland, 2018; pp. 195–228. ISBN 978-3-319-98850-4. [Google Scholar]
- Guzik, M.T.; Austin, A.D.; Cooper, S.J.B.; Harvey, M.S.; Humphreys, W.F.; Bradford, T.; Eberhard, S.M.; King, R.A.; Leys, R.; Muirhead, K.A.; et al. Is the Australian subterranean fauna uniquely diverse? Invertebr. Syst. 2010, 24, 407–418. [Google Scholar] [CrossRef] [Green Version]
- Knight, L.R.F.D. The aquatic macro-invertebrate fauna of Swildon’s Hole, Mendip Hills, Somerset, UK. Cave Karst Sci. 2011, 38, 81–92. [Google Scholar]
- Pipan, T.; Culver, D.C.; Papi, F.; Kozel, P. Partitioning diversity in subterranean invertebrates: The epikarst of Slovenia. PLoS ONE 2018, 13, e0195991. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Hypothetical relationship between pore size (habitat diameter) in subterranean habitats and body size, with selective forces indicated by arrows. Below a minimum (mi), there is not sufficient space for animals to occur without burrowing. Above a maximum (mj), body size is likely constrained by other factors, such as phylogenetic and structural constraints. The relationship need not be linear but is presented as such for simplicity. From Pipan and Culver [24]. Used with permission of the National Speleological Society (www.caves.org).
Figure 1.
Hypothetical relationship between pore size (habitat diameter) in subterranean habitats and body size, with selective forces indicated by arrows. Below a minimum (mi), there is not sufficient space for animals to occur without burrowing. Above a maximum (mj), body size is likely constrained by other factors, such as phylogenetic and structural constraints. The relationship need not be linear but is presented as such for simplicity. From Pipan and Culver [24]. Used with permission of the National Speleological Society (www.caves.org).

Figure 2.
Histograms of body lengths of inhabitants (bottom) and log of pore size (top) for different subterranean habitats. From Pipan and Culver [24], modified. Log pore (habitat) size was used because the range of habitat sizes ranged over several magnitudes. The MSS (milieu souterrain superficiel) is the habitat of interconnected cracks and crevices of scree slopes, especially covered ones. Used with permission of the National Speleological Society (www.caves.org).
Figure 2.
Histograms of body lengths of inhabitants (bottom) and log of pore size (top) for different subterranean habitats. From Pipan and Culver [24], modified. Log pore (habitat) size was used because the range of habitat sizes ranged over several magnitudes. The MSS (milieu souterrain superficiel) is the habitat of interconnected cracks and crevices of scree slopes, especially covered ones. Used with permission of the National Speleological Society (www.caves.org).

Figure 3.
Correspondence analysis (CA) ordination diagram with both samples and species plotted. Abbreviations of species names are explained in Table S1. From Dumnicka et al. [37], modified.

Figure 4.
Number of studies performed in particular habitats (green bar) and number of records of specialized subterranean species found in them (light green bar) in Poland. From Dumnicka and Galas [23], modified.
Figure 4.
Number of studies performed in particular habitats (green bar) and number of records of specialized subterranean species found in them (light green bar) in Poland. From Dumnicka and Galas [23], modified.

Figure 5.
Relationship between the log of the numbers of quadrats and log of the numbers of species for the PASCALIS project. The relationship between the two is Y (number of quadrats) = 677 × X (Number of Species)−1.76. Data from Deharveng et al. [49].
Figure 5.
Relationship between the log of the numbers of quadrats and log of the numbers of species for the PASCALIS project. The relationship between the two is Y (number of quadrats) = 677 × X (Number of Species)−1.76. Data from Deharveng et al. [49].

Figure 6.
The number of stygobiotic water mites recorded in various squares in Poland. Ellipses (or circles) represent areas in which water mite fauna was studied in various types of surface/subterranean waters but stygobionts were not found. From Dumnicka et al. [57], modified.
Figure 6.
The number of stygobiotic water mites recorded in various squares in Poland. Ellipses (or circles) represent areas in which water mite fauna was studied in various types of surface/subterranean waters but stygobionts were not found. From Dumnicka et al. [57], modified.

Figure 7.
Distribution of obligate subterranean species by country, corrected for area. See Zagmajster et al. [62] for details. With permission of Springer Nature.
Figure 7.
Distribution of obligate subterranean species by country, corrected for area. See Zagmajster et al. [62] for details. With permission of Springer Nature.

Figure 8.
Map of species richness patterns of European stygobionts. (A) Species richness of 10,000 km2 cells. (B) Relationship between the cell average of species richness per 0.09′ latitudinal band and latitude. Black horizontal bars and boxes show the median and interquartile range, respectively, for latitudinal banks. The maximum length of each whisker is 1.5 times the interquartile range and open circles represent outliers. The thick red line is the fit of generalized additive model to the averages of latitudinal bands. From Zagmajster et al. [51]. With permission of Springer Nature.
Figure 8.
Map of species richness patterns of European stygobionts. (A) Species richness of 10,000 km2 cells. (B) Relationship between the cell average of species richness per 0.09′ latitudinal band and latitude. Black horizontal bars and boxes show the median and interquartile range, respectively, for latitudinal banks. The maximum length of each whisker is 1.5 times the interquartile range and open circles represent outliers. The thick red line is the fit of generalized additive model to the averages of latitudinal bands. From Zagmajster et al. [51]. With permission of Springer Nature.

Figure 9.
Differences in species richness (+SD) for porous aquifers from Wallonia (WA), Jura (JU), Roussilon (RO), Cantabria (CA), Padano–Alpine region (PA), Slovenia (SL), Rhône River valley (RH) and Garonne (GA) regions. (a)—karst, (b)—interstitial. From Malard et al. [58], modified.
Figure 9.
Differences in species richness (+SD) for porous aquifers from Wallonia (WA), Jura (JU), Roussilon (RO), Cantabria (CA), Padano–Alpine region (PA), Slovenia (SL), Rhône River valley (RH) and Garonne (GA) regions. (a)—karst, (b)—interstitial. From Malard et al. [58], modified.

Figure 10.
Relative contribution of within drip (α-diversity), among drip, among cave, and among region diversity (all β-diversity) to the overall diversity of 30 epikarst copepod species, relative to random expectation. From Pipan et al. [65].
Figure 10.
Relative contribution of within drip (α-diversity), among drip, among cave, and among region diversity (all β-diversity) to the overall diversity of 30 epikarst copepod species, relative to random expectation. From Pipan et al. [65].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Aquatic subterranean habitats: after Botosaneanu [8]—simplified and modified. The shallow/deep dichotomy is based on a diving line at 10 m [10].
| Major Categories | Botosaneanu’s [8] Divisions, Supplemented | Pore Size (after [8]) | Corrections to Pore Size | Depth | Replicate of Surface Habitat |
|---|---|---|---|---|---|
| Large habitats (caves in karst or pseudokarst) | cave water in general | large | variable | variable | |
| percolation water (rimstone pools) | large | small 1 | shallow | No | |
| epiphreatic (streams) | large | variable | Yes | ||
| phreatic lakes | large | deep | Yes | ||
| cenotes | large | deep | Yes | ||
| anchialine | large | variable | No | ||
| lava tubes, mines, etc. | large | shallow | Yes | ||
| artesian wells | large | deep | No | ||
| calcrete aquifers 2 | large | variable | No | ||
| Porous habitats | Alluvial wells | small | deep | No | |
| hyporheal | small | shallow | No | ||
| hyporheal in caves | small | variable | No | ||
| Water on border of fw stagnant water | small | shallow | No | ||
| hypotelminorheal | small | shallow | No | ||
| artificial filters of sand or gravel | small | shallow | No | ||
| interstitial water of marine beaches | small | shallow | No | ||
| interstitial water of marine sublitoral | small | shallow | No | ||
| interstitial of brackish or hypersaline water bodies | small | shallow | No | ||
| Ecotones (springs) | general | small | variable | n/a | |
| karst | small | variable | n/a | ||
| phreatic | small | variable | n/a | ||
| hypotelminorheic | small | shallow | n/a | ||
| thermal springs | small | variable | n/a | ||
| travertine springs 3 | small | shallow | n/a |
Table 2.
Groundwater biodiversity in six European countries—from Deharveng et al. [49], modified.
Table 2.
Groundwater biodiversity in six European countries—from Deharveng et al. [49], modified.
| Country | No. Sampled Cells | No. Sampled Sites | No. Observed Species | No. Predicted ssp. (Jackknife) |
|---|---|---|---|---|
| Belgium | 17 | 155 | 33 | 43 |
| France | 566 | 1712 | 320 | 434 |
| Italy | 337 | 1580 | 288 | 394 |
| Portugal | 24 | 34 | 48 | 88 |
| Slovenia | 54 | 491 | 183 | 246 |
| Spain | 241 | 737 | 216 | 308 |
| ALL | 1228 | 4709 | 930 | 1291 |
Table 3.
Caves and other karst sites with 25 or more obligate aquatic subterranean species (stygobionts). From Culver and Pipan [61], modified.
Table 3.
Caves and other karst sites with 25 or more obligate aquatic subterranean species (stygobionts). From Culver and Pipan [61], modified.
| Site Name | Country | No. of Species |
|---|---|---|
| Postojna–Planina Cave System | Slovenia | 48 |
| Vjetrenica | Bosnia and Hercegovina | 40 |
| Walsingham Cave | Bermuda | 37 |
| Triadou wells | France | 34 |
| Robe River | Australia | 32 |
| Jameos del Aqua | Lanzarote, Canary Islands, Spain | 32 |
| Križna Jama | Slovenia | 29 |
| Logarček | Slovenia | 28 |
| Edwards Aquifer | Texas, USA | 27 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dumnicka, E.; Pipan, T.; Culver, D.C. Habitats and Diversity of Subterranean Macroscopic Freshwater Invertebrates: Main Gaps and Future Trends. Water 2020, 12, 2170. https://doi.org/10.3390/w12082170
AMA Style
Dumnicka E, Pipan T, Culver DC. Habitats and Diversity of Subterranean Macroscopic Freshwater Invertebrates: Main Gaps and Future Trends. Water. 2020; 12(8):2170. https://doi.org/10.3390/w12082170
Chicago/Turabian StyleDumnicka, Elzbieta, Tanja Pipan, and David C. Culver. 2020. "Habitats and Diversity of Subterranean Macroscopic Freshwater Invertebrates: Main Gaps and Future Trends" Water 12, no. 8: 2170. https://doi.org/10.3390/w12082170
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.