Drought-Induced Regulatory Cascades and Their Effects on the Nutritional Quality of Developing Potato Tubers

 ,
, 
 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
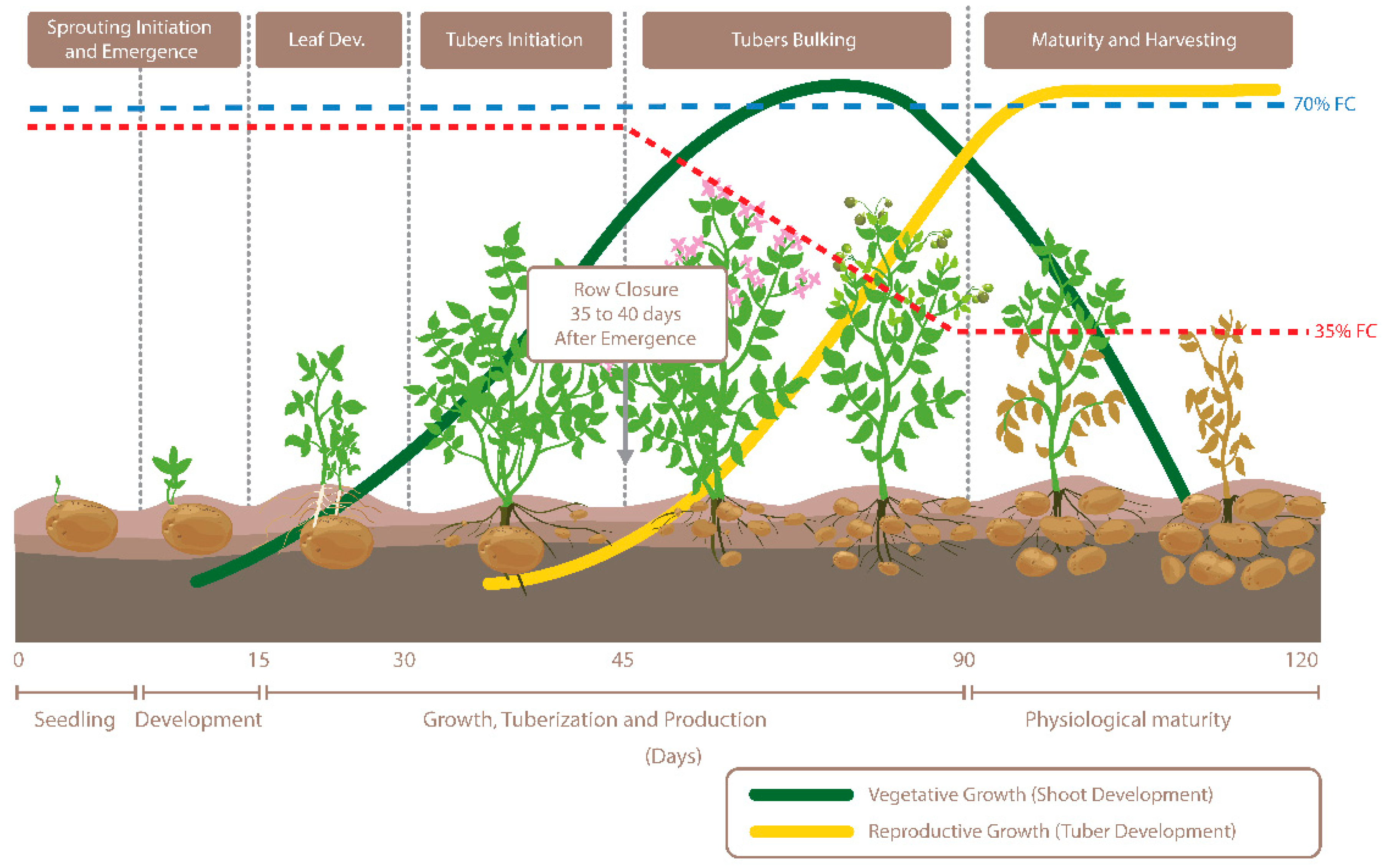
2.1. Experimental Design and Plant Growth Conditions
2.2. Physiological Measurements
2.3. Amino Acid Profiling and Abscisic Acid Content
2.4. Transcriptome and Small RNA Sequencing
2.5. RNA and Small RNA Read Mapping and Analysis
2.6. Differential RNA Expression Analysis
3. Results
3.1. Physiological Response
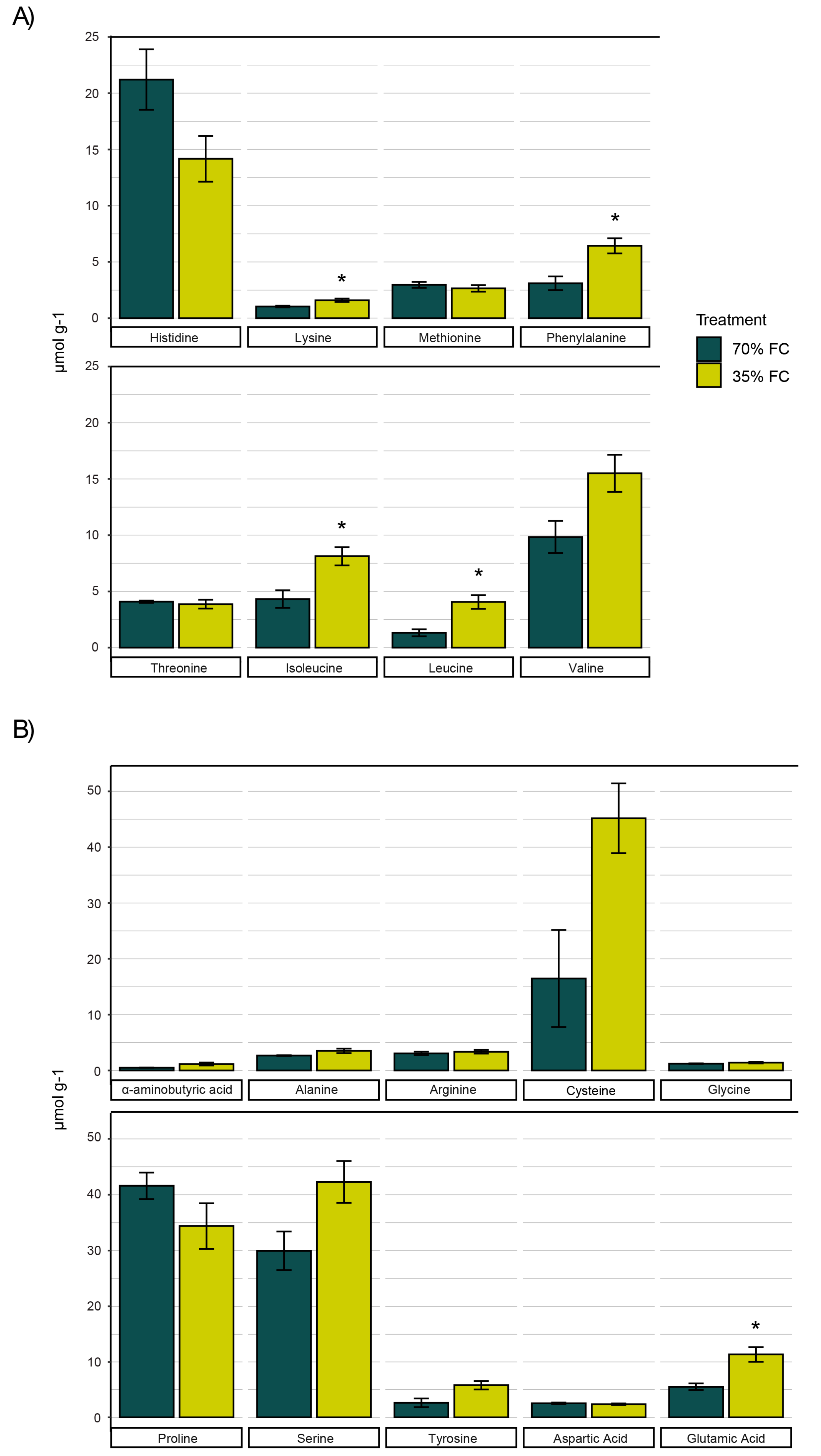
3.2. Tuber Amino Acid Fluctuations in Response to Soil Moisture Deficit
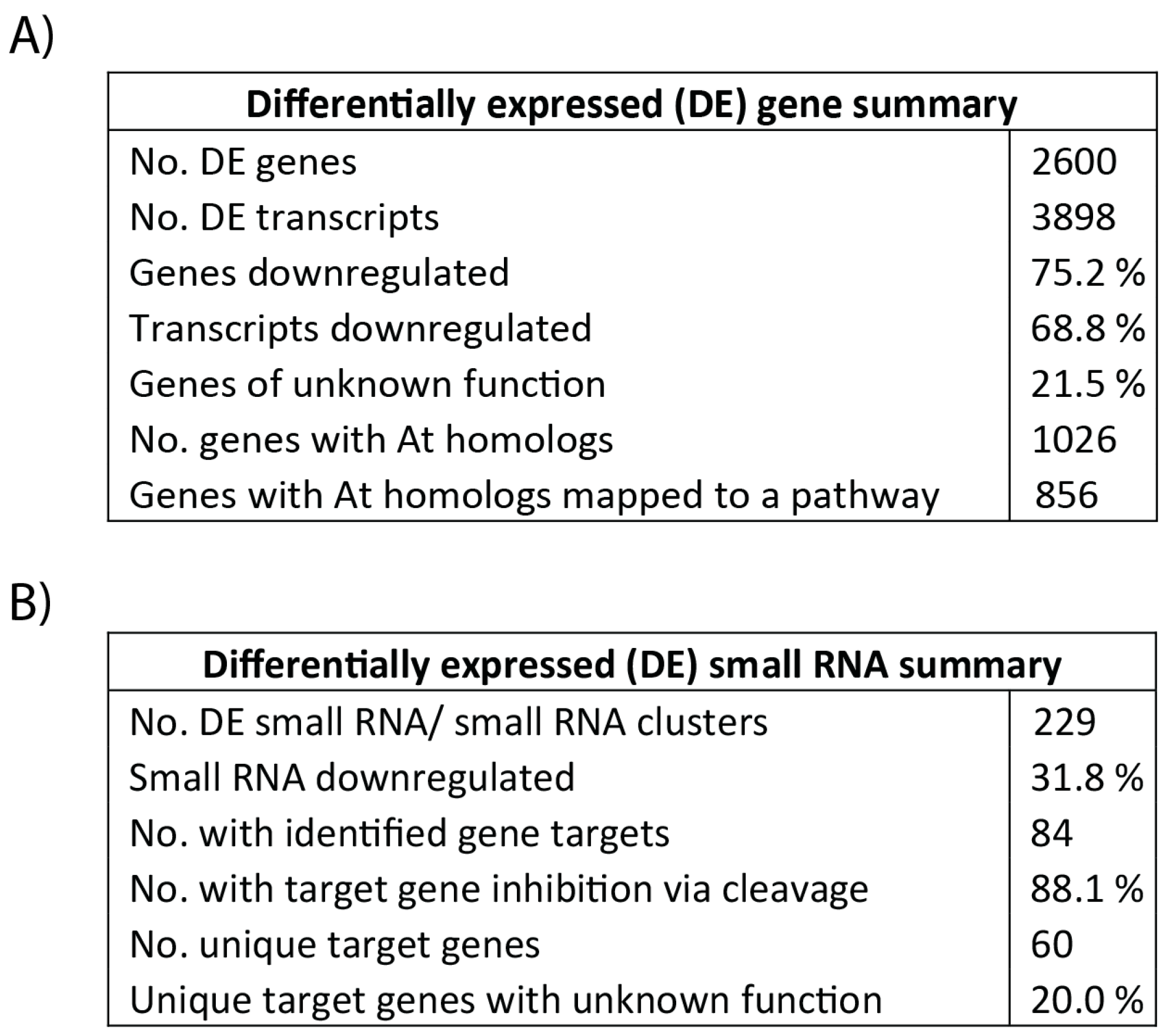
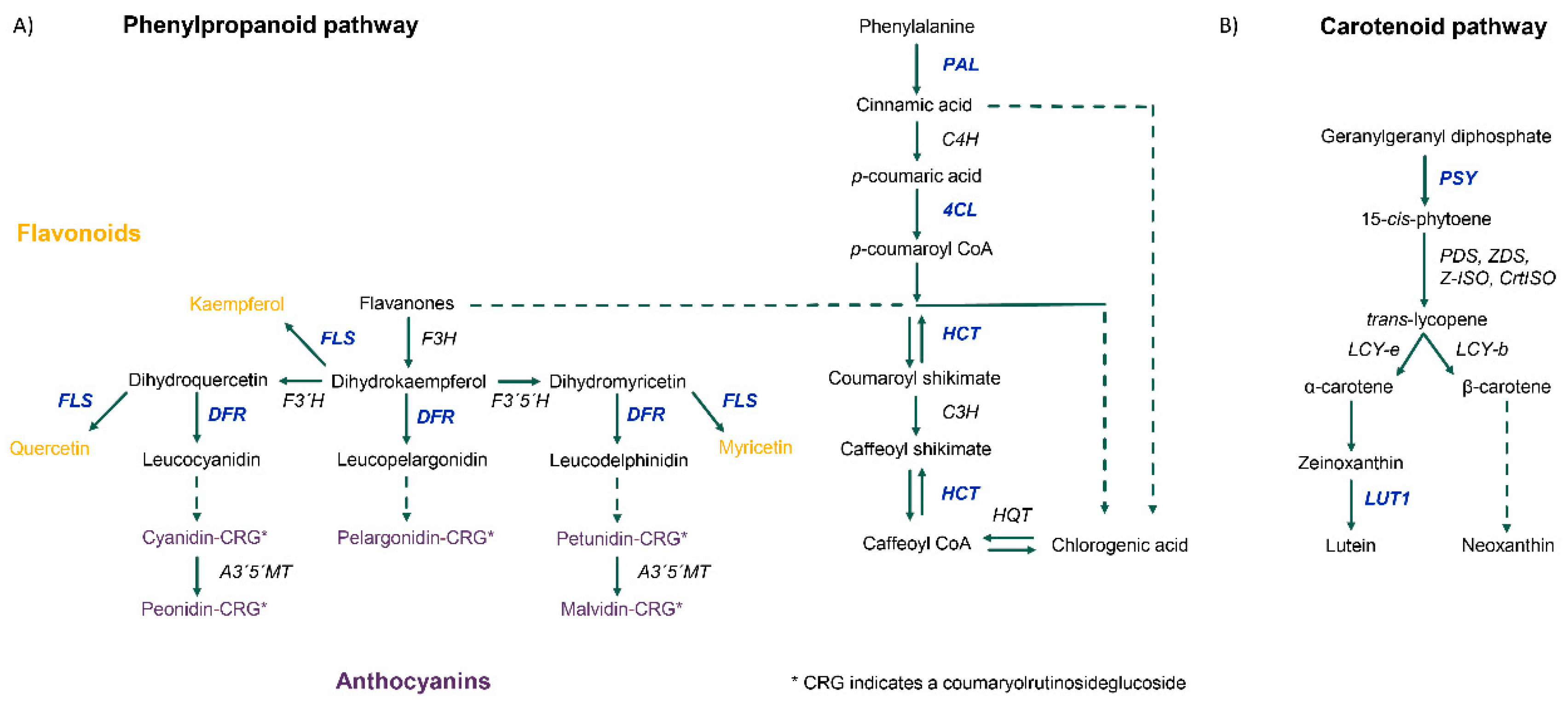
3.3. Differential Gene Expression and Regulatory Cascades in Developing Tubers Under Drought Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- King, J.C.; Slavin, J.L. White potatoes, human health, and dietary guidance. Adv. Nutr. 2013, 4, 393S–401S. [Google Scholar] [CrossRef] [Green Version]
- Chun, O.K.; Kim, D.O.; Smith, N.; Schroeder, D.; Han, J.T.; Lee, C.Y. Daily consumption of phenolics and total antioxidant capacity from fruit and vegetables in the American diet. J. Sci. Food Agric. 2005, 85, 1715–1724. [Google Scholar] [CrossRef]
- Andre, C.M.; Oufir, M.; Guignard, C.; Hoffmann, L.; Hausman, J.F.; Evers, D.; Larondelle, Y. Antioxidant profiling of native Andean potato tubers (Solanum tuberosum L.) reveals cultivars with high levels of β-carotene, α-tocopherol, chlorogenic acid, and petanin. J. Agric. Food Chem. 2007, 55, 10839–10849. [Google Scholar] [CrossRef]
- Rosenthal, S.; Jansky, S. Effect of production site and storage on antioxidant levels in specialty potato (Solanum tuberosum L.) tubers. J. Sci. Food Agric. 2008, 88, 2087–2092. [Google Scholar] [CrossRef]
- Payyavula, R.S.; Navarre, D.A.; Kuhl, J.C.; Pantoja, A.; Pillai, S.S. Differential effects of environment on potato phenylpropanoid and carotenoid expression. BMC Plant Biol. 2012, 12, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezekiel, R.; Singh, N.; Sharma, S.; Kaur, A. Beneficial phytochemicals in potato—A review. Food Res. 2013, 50, 487–496. [Google Scholar] [CrossRef]
- Deng, Y.; Lu, S. Biosynthesis and regulation of phenylpropanoids in plants. Crit. Rev. Plant Sci. 2017, 36, 257–290. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, C.J. Multifaceted regulations of gateway enzyme phenylalanine ammonia-lyase in the biosynthesis of phenylpropanoids. Mol. Plant 2015, 8, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zeng, Z.; Liu, Z.; Xia, R. Small RNAs, emerging regulators critical for the development of horticultural traits. Hort Res. 2018, 5, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Yang, J.; Wang, Z.; Wen, Y.; Wang, J.; He, W.; Liu, B.; Si, H.; Wang, D. Identification of novel and conserved microRNAs related to drought stress in potato by deep sequencing. PLoS ONE 2014, 9, e95489. [Google Scholar] [CrossRef]
- Pęksa, A.; Kita, A.; Kułakowska, K.; Aniołowska, M.; Hamouz, K.; Nemś, A. The quality of protein of coloured fleshed potatoes. Food Chem. 2013, 141, 2960–2966. [Google Scholar] [CrossRef]
- Bártová, V.; Bárta, J.; Brabcová, A.; Zdráhal, Z.; Horáčková, V. Amino acid composition and nutritional value of four cultivated South American potato species. J. Food Compos. Anal. 2015, 40, 78–85. [Google Scholar] [CrossRef]
- Galili, G.; Amir, R.; Fernie, A.R. The regulation of essential amino acid synthesis and accumulation in plants. Annu. Rev. Plant Biol. 2016, 67, 153–178. [Google Scholar] [CrossRef] [PubMed]
- Rykaczewska, K. The effect of high temperature occurring in subsequent stages of plant development on potato yield and tuber physiological defects. Am. Potato J. 2015, 92, 339. [Google Scholar] [CrossRef] [Green Version]
- Boguszewska-Mańkowska, D.; Pieczyński, M.; Wyrzykowska, A.; Kalaji, H.M.; Sieczko, L.; Szweykowska-Kulińska, Z.; Zagdańska, B. Divergent strategies displayed by potato (Solanum tuberosum L.) cultivars to cope with soil drought. J. Agron. Crop Sci. 2018, 204, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Wishart, J.; George, T.S.; Brown, L.K.; White, P.J.; Ramsay, G.; Jones, H.; Gregory, P.J. Field phenotyping of potato to assess root and shoot characteristics associated with drought tolerance. Plant Soil 2014, 378, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Pieczynski, M.; Wyrzykowska, A.; Milanowska, K.; Boguszewska-Mankowska, D.; Zagdanska, B.; Karlowski, W.; Jarmolowski, A.; Szweykowska-Kulinska, Z. Genomewide identification of genes involved in the potato response to drought indicates functional evolutionary conservation with Arabidopsis plants. Plant Biotechnol. J. 2018, 16, 603–614. [Google Scholar] [CrossRef] [Green Version]
- Sprenger, H.; Erban, A.; Seddig, S.; Rudack, K.; Thalhammer, A.; Le, M.Q.; Walther, D.; Zuther, E.; Köhl, K.I.; Kopka, J.; et al. Metabolite and transcript markers for the prediction of potato drought tolerance. Plant Biotechnol. J. 2018, 16, 939–950. [Google Scholar] [CrossRef] [Green Version]
- Inaba, K.; Fujiwara, T.; Hayashi, H.; Chino, M.; Komeda, Y.; Naito, S. Isolation of an Arabidopsis thaliana mutant, mto1 that overaccumulates soluble methionine (Temporal and spatial patterns of soluble methionine accumulation). Plant Physiol. 1994, 104, 881–887. [Google Scholar] [CrossRef] [Green Version]
- Cohen, S.A.; Michaud, D.P. Synthesis of a fluorescent derivatizing reagent, 6-aminoquinolyl-N-hydroxysuccinimidyl carbamate, and its application for the analysis of hydrolysate amino acids via high-performance liquid chromatography. Anal. Biochem. 1993, 211, 279–287. [Google Scholar] [CrossRef]
- Yan, D.; Easwaran, V.; Chau, V.; Okamoto, M.; Ierullo, M.; Kimura, M.; Endo, A.; Yano, R.; Pasha, A.; Gong, Y.; et al. NIN-like protein 8 is a master regulator of nitrate-promoted seed germination in Arabidopsis. Nat. Commun. 2016, 7, 13179. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinform. 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrew, S. FastQC: A quality control tool for high throughput sequence data. 2015. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 5 September 2018).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. Embnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Johnson, N.R.; Yeoh, J.M.; Coruh, C.; Axtell, M.J. Improved placement of multi-mapping small RNAs. G3 (Bethesda) 2016, 6, 2103–2111. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Zhuang, Z.; Zhao, X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Prashar, A.; Yildiz, J.; McNicol, J.W.; Bryan, G.J.; Jones, H.G. Infra-red thermography for high throughput field phenotyping in Solanum tuberosum. PLoS ONE 2013, 8, e65816. [Google Scholar] [CrossRef]
- Ren, H.; Gray, W.M. SAUR proteins as effectors of hormonal and environmental signals in plant growth. Mol. Plant 2015, 8, 1153–1164. [Google Scholar] [CrossRef] [Green Version]
- Sprenger, H.; Kurowsky, C.; Horn, R.; Erban, A.; Seddig, S.; Rudack, K.; Fischer, A.; Walther, D.; Zuther, E.; Köhl, K.; et al. The drought response of potato reference cultivars with contrasting tolerance. Plant Cell Environ. 2016, 39, 2370–2389. [Google Scholar] [CrossRef]
- Obidiegwu, J.E.; Bryan, G.J.; Jones, H.G.; Prashar, A. Coping with drought: Stress and adaptive responses in potato and perspectives for improvement. Front. Plant Sci. 2015, 6, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, L.; Zhang, H.; Gan, X.; Zhang, L.; Chen, Y.; Nie, F.; Shi, L.; Li, M.; Guo, Z.; Zhang, G.; et al. Transcriptome profiling of the potato (Solanum tuberosum L.) plant under drought stress and water-stimulus conditions. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Vasquez-Robinet, C.; Mane, S.P.; Ulanov, A.V.; Watkinson, J.I.; Stromberg, V.K.; De Koeyer, D.; Schafleitner, R.; Willmot, D.B.; Bonierbale, M.; Bohnert, H.J.; et al. Physiological and molecular adaptations to drought in Andean potato genotypes. J. Exp. Bot. 2008, 59, 2109–2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dezfulian, M.; Foreman, C.; Jalili, E.; Pal, M.; Dhaliwal, R.; Roberto, D.; Imre, K.; Kohalmi, S.; Crosby, W. Acetolactate synthase regulatory subunits play divergent and overlapping roles in branched-chain amino acid synthesis and Arabidopsis development. BMC Plant Biol. 2017, 17, 71. [Google Scholar] [CrossRef] [Green Version]
- Dhondt, S.; Geoffroy, P.; Stelmach, B.A.; Legrand, M.; Heitz, T. Soluble phospholipase A2 activity is induced before oxylipin accumulation in tobacco mosaic virus-infected tobacco leaves and is contributed by patatin-like enzymes. Plant J. 2000, 23, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R. Tuber storage proteins. Ann. Bot. 2003, 91, 755–769. [Google Scholar] [CrossRef] [Green Version]
- Reyes, L.F.; Miller, J.C.; Cisneros-Zevallos, L. Antioxidant capacity, anthocyanins and total phenolics in purple-and red-fleshed potato (Solanum tuberosum L.) genotypes. Am. Potato. J. 2005, 82, 271. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Yield (g) | No. of Tubers | CCI | Fv/Fm | Canopy Temp (°C) |
|---|---|---|---|---|---|
| 35% FC | 1381.7 (248.1) | 12.8 (0.8) | 19.5 (0.2) | 0.24 (0.05) | 28.0 (0.3) |
| 70% FC | 1737.3 (198.1) | 18.1 (3.5) | 15.4 (1.6) | 0.38 (0.03) | 24.1 (0.9) |
| Gene Regulation | Pathway | Gene Name | Description | Log2 Fold Change | At Homologs | Descriptor | % ID |
|---|---|---|---|---|---|---|---|
| Down- | ABA signaling | PGSC0003DMG400002100 | Abscisic acid receptor PYR1 | −2.07 | AT4G17,870 | PYR1 | 72.2 |
| regulated | AT5G46,790 | PYL1 | 61.0 | ||||
| Auxin | PGSC0003DMG400001589 | Amino acid transporter | −5.30 | AT2G21050 | LAX2 | 86.3 | |
| biosynthesis | PGSC0003DMG400024978 | Indole-3-acetic acid-amido | −5.06 | AT2G14960 | GH3.1 | 77.3 | |
| and signaling | synthetase GH3.3 | AT2G23170 | GH3.3 | 74.1 | |||
| AT4G37390 | GH3.2 | 73.3 | |||||
| AT1G59500 | GH3.4 | 69.8 | |||||
| PGSC0003DMG400024997 | Indole-3-acetic acid-amido synthetase GH3.6 | −2.17 | AT5G54510 | GH3.6 | 70.8 | ||
| PGSC0003DMG400014707 | Flavin monooxygenase | −3.42 | AT4G28720 | YUC8 | 68.3 | ||
| AT5G43890 | YUC5 | 67.2 | |||||
| PGSC0003DMG400026087 | Flavin monooxygenase | −3.09 | AT5G11320 | YUC4 | 57.4 | ||
| AT4G32540 | YUC | 54.3 | |||||
| PGSC0003DMG400003773 | SAUR family protein | −8.34 | AT1G75580 | SAUR51 | 72.2 | ||
| AT1G19830 | SAUR54 | 61.5 | |||||
| PGSC0003DMG400001667 | SAUR family protein | −7.40 | AT4G38860 | SAUR16 | 64.8 | ||
| AT4G34760 | SAUR50 | 64.5 | |||||
| AT2G21220 | SAUR12 | 63.5 | |||||
| AT2G16580 | SAUR8 | 63.0 | |||||
| PGSC0003DMG400001614 | SAUR family protein | −3.75 | AT4G34760 | SAUR50 | 75.7 | ||
| AT4G38860 | SAUR16 | 73.3 | |||||
| AT2G16580 | SAUR8 | 71.3 | |||||
| AT2G21220 | SAUR12 | 71.1 | |||||
| PGSC0003DMG400001668 | SAUR family protein | −3.71 | AT4G38860 | SAUR16 | 77.1 | ||
| AT4G34760 | SAUR50 | 76.6 | |||||
| AT2G21220 | SAUR12 | 75.0 | |||||
| AT2G16580 | SAUR8 | 70.4 | |||||
| PGSC0003DMG400001655 | SAUR family protein | −2.98 | AT4G34750 | SAUR49 | 54.0 | ||
| PGSC0003DMG400022233 | SAUR family protein ARG7 | −2.93 | AT3G12830 | SAUR72 | 64.4 | ||
| AT1G16510 | SAUR41 | 55.1 | |||||
| PGSC0003DMG400001615 | SAUR family protein | −2.06 | AT4G34760 | SAUR50 | 73.8 | ||
| AT4G38860 | SAUR16 | 71.4 | |||||
| AT2G21220 | SAUR12 | 69.2 | |||||
| AT2G16580 | SAUR8 | 68.5 | |||||
| Carotenoid biosynthesis | PGSC0003DMG400028180 | Cytochrome P450-type monooxygenase 97C11 | −2.07 | AT3G53130 | LUT1 | 77.2 | |
| PGSC0003DMG400024063 | Phytoene synthase 1, chloroplastic | −5.07 | AT5G17230 | PSY | 64.3 | ||
| Ethylene signaling | PGSC0003DMG400014204 | Transcription factor TSRF1 | −3.57 | AT3G23240 | ERF1 | 51.4 | |
| Phenylpropanoid | PGSC0003DMG400003605 | Dihydroflavonol 4-reductase | −5.19 | AT5G42800 | DFR | 59.2 | |
| biosynthesis | PGSC0003DMG400014093 | Flavonol synthase | −2.19 | AT5G08640 | FLS1 | 62.5 | |
| AT5G63590 | FLS3 | 50.3 | |||||
| PGSC0003DMG400014152 | Hydroxycinnamoyl transferase | −2.00 | AT5G48930 | HCT | 77.8 | ||
| PGSC0003DMG400023458 | Phenylalanine ammonia- | −4.68 | AT3G10340 | PAL4 | 79.9 | ||
| lyase | AT5G04230 | PAL3 | 73.2 | ||||
| PGSC0003DMG400014223 | 4-coumarate--CoA ligase 2 | −2.30 | AT3G21240 | 4CL2 | 68.5 | ||
| AT1G51680 | 4CL1 | 67.9 | |||||
| AT3G21230 | 4CL4 | 58.9 | |||||
| PGSC0003DMG400028929 | 4-coumarate--CoA ligase 2 | −2.00 | AT3G21240 | 4CL2 | 69.2 | ||
| AT1G51680 | 4CL1 | 68.8 | |||||
| AT3G21230 | 4CL4 | 59.8 | |||||
| Up-regulated | Amino acid biosynthesis | PGSC0003DMG400034102 | Acetolactate synthase | 2.20 | AT3G48560 | CSR1 | 76.9 |
| Protein folding | PGSC0003DMG400008223 | Heat shock factor protein HSF30 | 4.44 | AT2G26150 | HSFA2 | 51.0 | |
| PGSC0003DMG400003219 | Small heat shock protein, chloroplastic | 4.11 | AT4G27670 | Heat shock protein 21 | 53.7 | ||
| PGSC0003DMG400030341 | Small heat shock protein-Class I 17.6kD | 3.99 | AT2G29500 | HSP17.6B | 77.8 | ||
| PGSC0003DMG400024707 | Small heat shock protein | 2.90 | AT1G09080 | Heat shock protein 70 | 75.1 | ||
| PGSC0003DMG402028907 | Small heat shock protein 90 | 2.72 | AT5G52640 | Heat shock protein 90 | 52.0 | ||
| PGSC0003DMG400030426 | Small heat shock protein-Class I 17.6kD | 2.50 | AT2G29500 | HSP17.6B | 74.5 | ||
| Proteolysis | PGSC0003DMG400006185 | Skp1 1 | 2.56 | AT1G75950 | SKP1 | 74.4 | |
| PGSC0003DMG400006184 | Skp1 | 2.20 | AT1G75950 | SKP1 | 75.0 |
| Small RNA Cluster | Log2 Fold Change | Target Alignment | Target Gene | Log2-Fold Change | Protein Description |
|---|---|---|---|---|---|
| Cluster 34023 | 5.03 | AGCUCAUUAAUCUCUUCGAUA | PGSC0003DMG400009921 | −6.24 | Cysteine protease 14 |
| Cluster 23921 | 4.68 | AGGGUUCAAGAAAAUGCAUUA | PGSC0003DMG400029247 | −4.75 | Patatin group O |
| Cluster 15144 | 4.62 | AGGGUUCAAGAAAAUGCAUUA | |||
| Cluster 41775 | 4.49 | ACCUCAGGGUUCAAGAAAAUG | |||
| Cluster 83189 | 5.49 | AGGCACUGGCACUACUUCAGA | PGSC0003DMG400017091 | −4.25 | Patatin-01; Probable lipolytic |
| Cluster 83175 | 4.98 | AGCCAGUAAUAUUCACCAAGU | acyl hydrolase | ||
| Cluster 83174 | 3.45 | AGGCACUGGCACUACUUCAGA | |||
| Cluster 7920 | 4.95 | GGCAGCAAGUUCUUACAUGAC | PGSC0003DMG400008749 | −4.06 | Patatin-05; Probable lipolytic |
| Cluster 68384 | 3.01 | AUCAUUCCGGGUAUCAUUCUC | acyl hydrolase | ||
| Cluster 83190 | 2.87 | UUCCGGGUAUCAUUCUCGAAU | |||
| Cluster 83166 | 2.66 | UCCGGGUAUCAUUCUCGAAU | |||
| Cluster 68380 | 5.49 | AGGCACUGGCACUAAUUCAGA | PGSC0003DMG400014104 | −4.47 | Patatin-2-Kuras 4; Probable |
| Cluster 83164 | 5.49 | AGGCAGCUAAAUGGGGUCCUC | lipolytic acyl hydrolase | ||
| Cluster 20497 | 5.38 | CUGUUGGUGAUCCGGCGUUA | |||
| Cluster 68397 | 5.36 | GUUGCUACUGUUGGUGAUCCG | |||
| Cluster 83182 | 4.97 | GGCACUACUUCAGAGUUUGAU | PGSC0003DMG401017090 | −4.91 | Patatin-3-Kuras 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Da Ros, L.; Elferjani, R.; Soolanayakanahally, R.; Kagale, S.; Pahari, S.; Kulkarni, M.; Wahab, J.; Bizimungu, B. Drought-Induced Regulatory Cascades and Their Effects on the Nutritional Quality of Developing Potato Tubers. Genes 2020, 11, 864. https://doi.org/10.3390/genes11080864
Da Ros L, Elferjani R, Soolanayakanahally R, Kagale S, Pahari S, Kulkarni M, Wahab J, Bizimungu B. Drought-Induced Regulatory Cascades and Their Effects on the Nutritional Quality of Developing Potato Tubers. Genes. 2020; 11(8):864. https://doi.org/10.3390/genes11080864
Chicago/Turabian StyleDa Ros, Letitia, Raed Elferjani, Raju Soolanayakanahally, Sateesh Kagale, Shankar Pahari, Manoj Kulkarni, Jazeem Wahab, and Benoit Bizimungu. 2020. "Drought-Induced Regulatory Cascades and Their Effects on the Nutritional Quality of Developing Potato Tubers" Genes 11, no. 8: 864. https://doi.org/10.3390/genes11080864