Toll-Like Receptor 2 Expression as a New Hallmark of Advanced Endometriosis

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Peripheral Blood
2.2. Flow Cytometry and Sample Preparation: Monocytes and Dendritic Cells
2.3. Flow Cytometry and Sample Preparation: Lymphocytes, Natural Killer, and Natural Killer T-Like Cells
2.4. Statistical Analysis
3. Results
3.1. Characteristics of the Patients and Controls
3.2. Dendritic Cells, Monocytes, and Basic Peripheral Blood Lymphocyte Subsets and Expression of TLR2 Antigen in Healthy Individuals and Patients with Endometriosis
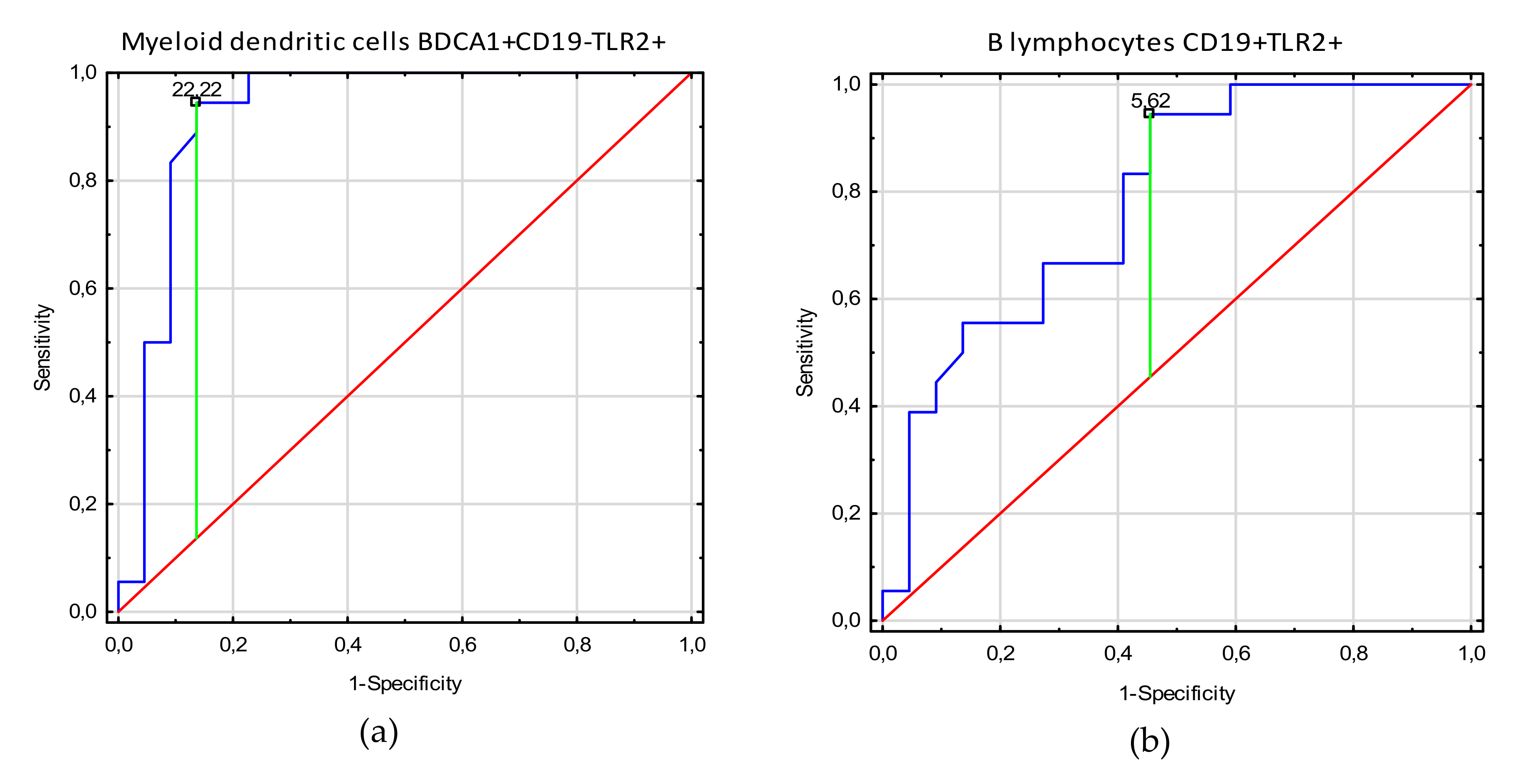
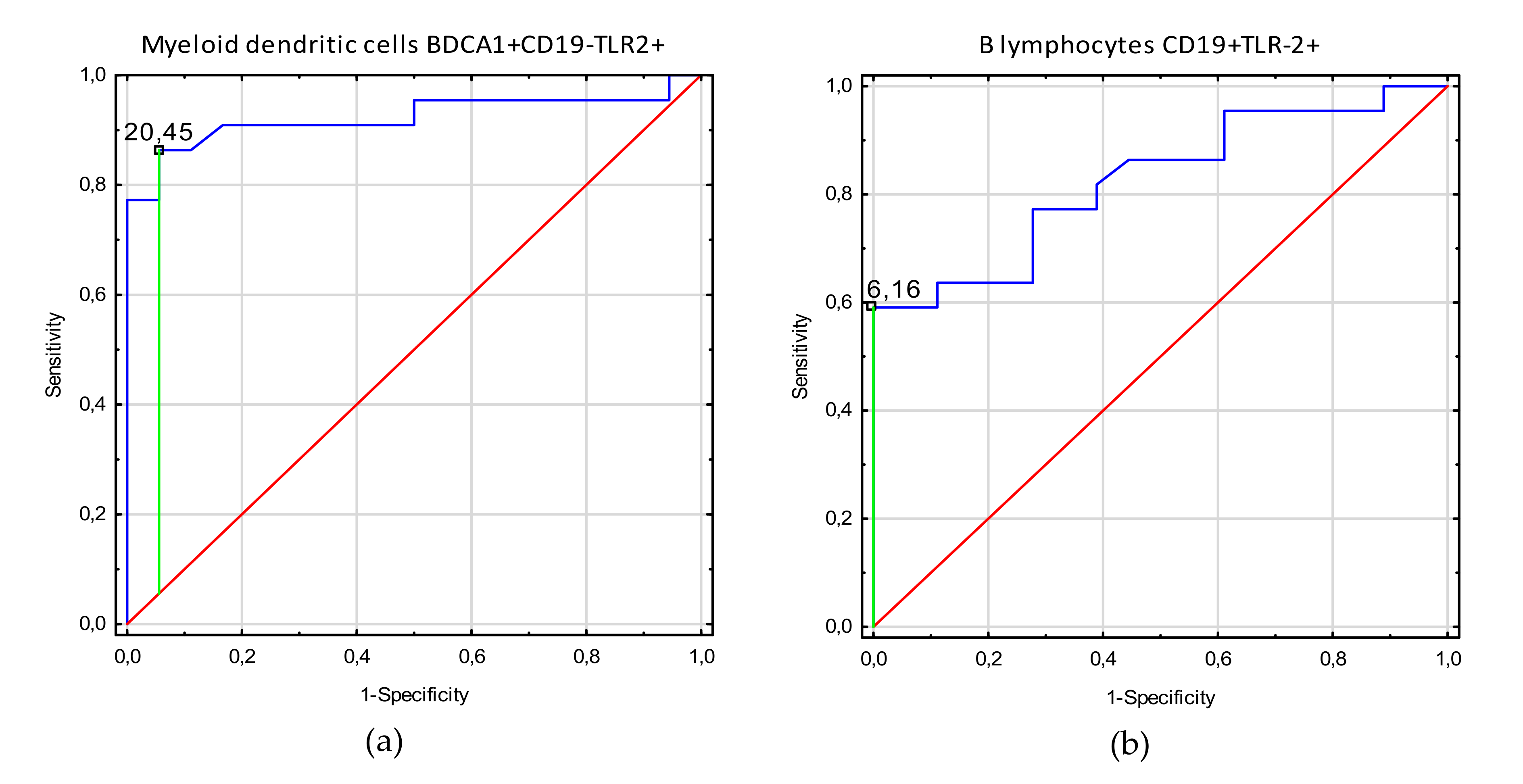
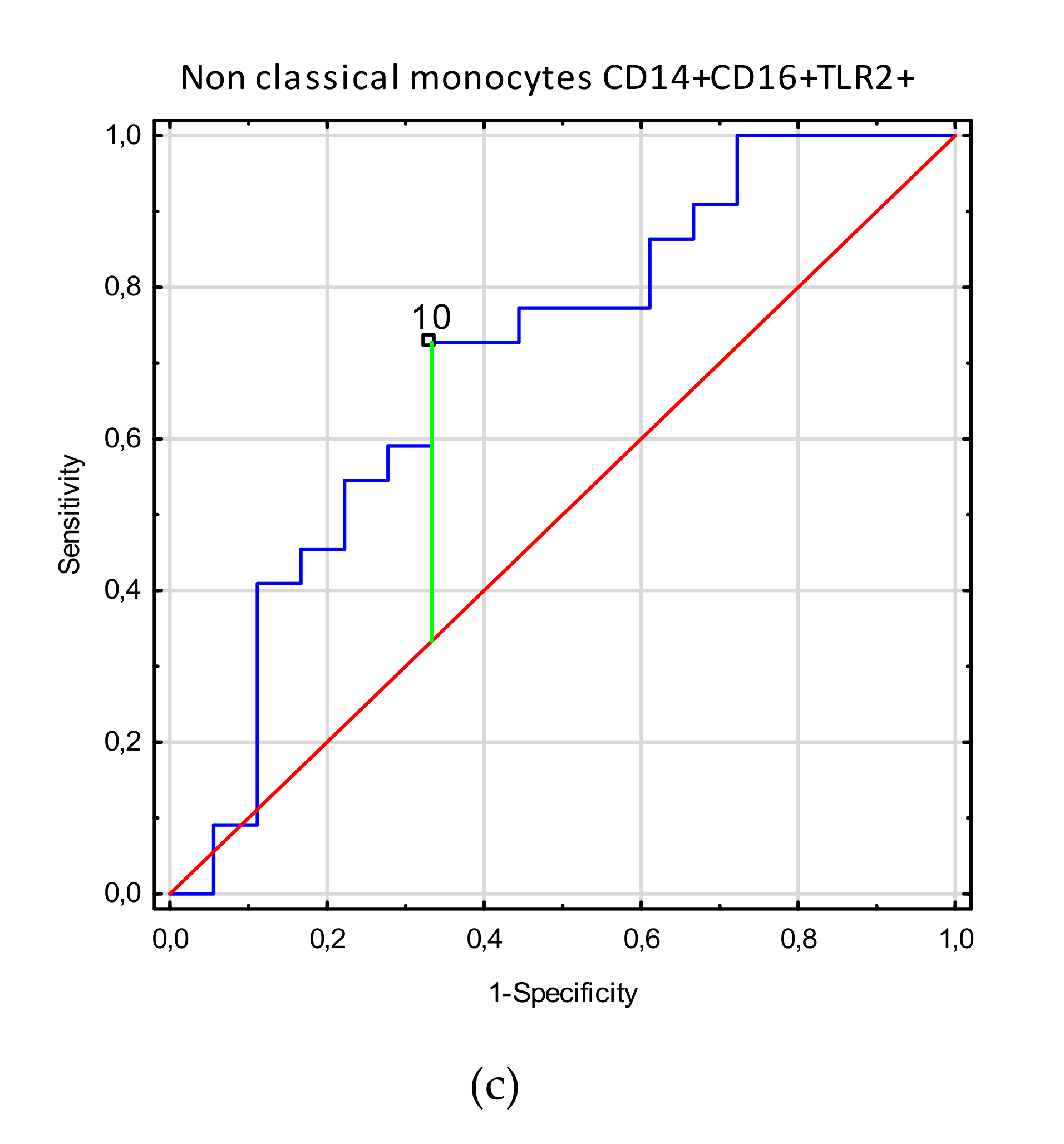
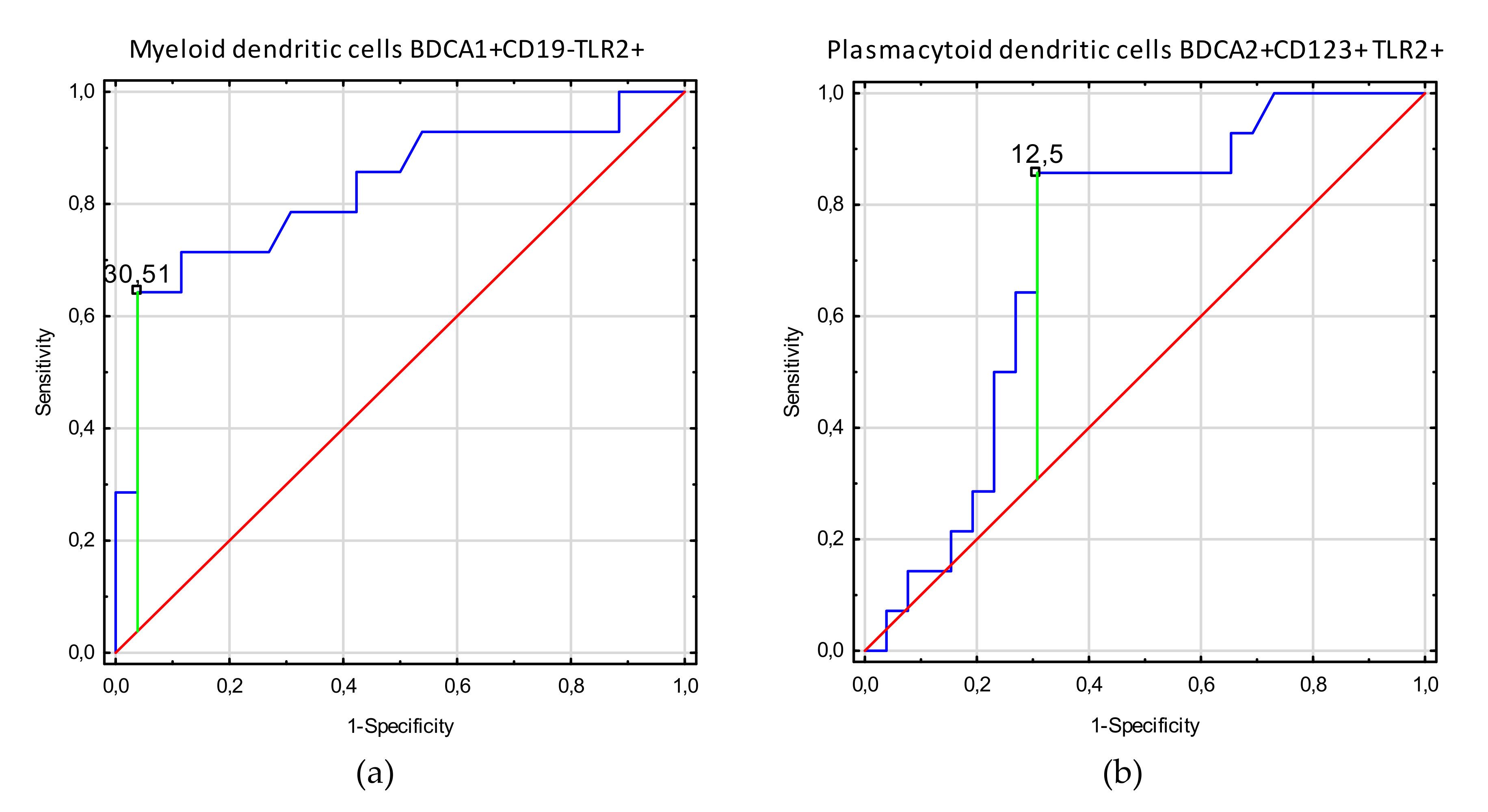
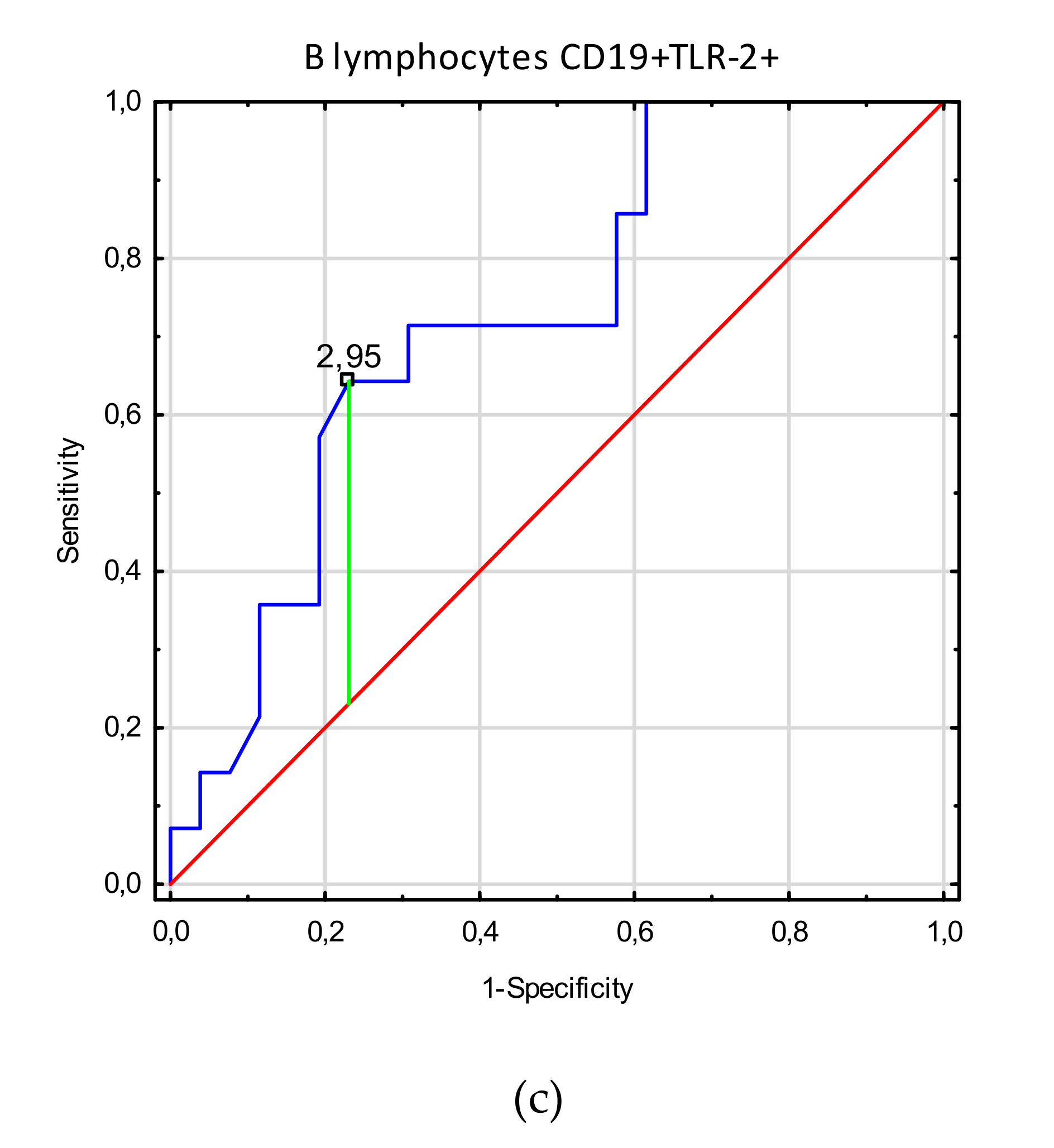
3.3. Receiver Operating Characteristic (ROC) Curve Analysis to Determine the Diagnostic Accuracy of TLR2 Expression on Myeloid Dendritic Cells, Plasmacytoid Dendritic Cells, Monocytes, and T and B Lymphocytes in Patients with Endometriosis vs. Controls
4. Discussion
4.1. The Amount of mDCs Increases Significantly in the Course of Endometriosis
4.2. Monocytes and Basic Peripheral Blood Lymphocytes and their Clinical Relation to Endometriosis
4.3. TLR2 Expression is Implicated in the Pathogenesis of Endometriosis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BDCA | Blood Dendritic Cell Antigen |
| DC | Dendritic Cell |
| CD | Cluster of Differentiation |
| TLR | Toll-like receptor |
References
- Khan, K.N.; Kitajima, M.; Fujishita, A.; Nakashima, M.; Masuzaki, H. Toll-like receptor system and endometriosis. J. Obstet. Gynaecol. Res. 2013, 39, 1281–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulun, S.E. Endometriosis. N. Engl. J. Med. 2009, 360, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Dunselman, G.A.J.; Vermeulen, N.; Becker, C.; Calhaz-Jorge, C.; D’Hooghe, T.; De Bie, B.; Heikinheimo, O.; Horne, A.W.; Kiesel, L.; Nap, A.; et al. ESHRE guideline: Management of women with endometriosis. Hum. Reprod. 2014, 29, 400–412. [Google Scholar] [CrossRef] [PubMed]
- May, K.E.; Villar, J.; Kirtley, S.; Kennedy, S.H.; Becker, C.M. Endometrial alterations in endometriosis: A systematic review of putative biomarkers. Hum. Reprod. Update 2011, 17, 637–653. [Google Scholar] [CrossRef]
- Fassbender, A.; Burney, R.O.; FO, D.; D’Hooghe, T.; Giudice, L. Update on biomarkers for the detection of endometriosis. Biomed Res. Int. 2015, 2015, 130854. [Google Scholar] [CrossRef] [Green Version]
- May, K.E.; Conduit-Hulbert, S.A.; Villar, J.; Kirtley, S.; Kennedy, S.H.; Becker, C.M. Peripheral biomarkers of endometriosis: A systematic review. Hum. Reprod. Update 2010, 16, 651–674. [Google Scholar] [CrossRef]
- Nisenblat, V.; Prentice, L.; Bossuyt, P.M.M.; Farquhar, C.; Hull, M.L.; Johnson, N. Combination of the non-invasive tests for the diagnosis of endometriosis. Cochrane Database Syst. Rev. 2016, 2016, CD012281. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; De Carolis, C.; Man, G.C.W.; Wang, C.C. The link between immunity, autoimmunity and endometriosis: A literature update. Autoimmun. Rev. 2018, 17, 945–955. [Google Scholar] [CrossRef]
- Dmowski, W.P.; Gebel, H.M.; Braun, D.P. The role of cell-mediated immunity in pathogenesis of endometriosis. Acta Obstet. Gynecol. Scand. Suppl. 1994, 159, 7–14. [Google Scholar]
- Bouet, P.-E.; Chao de la Barca, J.-M.; Boucret, L.; Descamps, P.; Legendre, G.; Hachem, H.E.; Blanchard, S.; Jeannin, P.; Reynier, P.; May-Panloup, P. Elevated Levels of Monocyte Chemotactic Protein-1 in the Follicular Fluid Reveals Different Populations among Women with Severe Endometriosis. J. Clin. Med. 2020, 9, 1306. [Google Scholar] [CrossRef]
- Marchionni, E.; Porpora, M.G.; Megiorni, F.; Piacenti, I.; Giovannetti, A.; Marchese, C.; Benedetti Panici, P.; Pizzuti, A. TLR4 T399I Polymorphism and Endometriosis in a Cohort of Italian Women. Diagnostics 2020, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Merad, M.; Sathe, P.; Helft, J.; Mortha, A. The dendritic cell lineage: Ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu. Rev. Immunol. 2013, 31, 563–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reizis, B.; Bunin, A.; Ghosh, H.S.; Lewis, K.L.; Sisirak, V. Plasmacytoid dendritic cells: Recent progress and open questions. Annu. Rev. Immunol. 2011, 29, 163–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izumi, G.; Koga, K.; Takamura, M.; Makabe, T.; Nagai, M.; Urata, Y.; Harada, M.; Hirata, T.; Hirota, Y.; Fujii, T.; et al. Mannose receptor is highly expressed by peritoneal dendritic cells in endometriosis. Fertil. Steril. 2017, 107, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fainaru, O.; Adini, A.; Benny, O.; Adini, I.; Short, S.; Bazinet, L.; Nakai, K.; Pravda, E.; Hornstein, M.D.; D’Amato, R.J.; et al. Dendritic cells support angiogenesis and promote lesion growth in a murine model of endometriosis. FASEB J. 2008, 22, 522–529. [Google Scholar] [CrossRef]
- Pencovich, N.; Luk, J.; Hantisteanu, S.; Hornstein, M.D.; Fainaru, O. The development of endometriosis in a murine model is dependent on the presence of dendritic cells. Reprod. Biomed. Online 2014, 28, 515–521. [Google Scholar] [CrossRef] [Green Version]
- Stanic, A.K.; Kim, M.; Styer, A.K.; Rueda, B.R. Dendritic cells attenuate the early establishment of endometriosis-like lesions in a murine model. Reprod. Sci. 2014, 21, 1228–1236. [Google Scholar] [CrossRef]
- Bacci, M.; Capobianco, A.; Monno, A.; Cottone, L.; Di Puppo, F.; Camisa, B.; Mariani, M.; Brignole, C.; Ponzoni, M.; Ferrari, S.; et al. Macrophages are alternatively activated in patients with endometriosis and required for growth and vascularization of lesions in a mouse model of disease. Am. J. Pathol. 2009, 175, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Ancuta, P.; Liu, K.Y.; Misra, V.; Wacleche, V.S.; Gosselin, A.; Zhou, X.; Gabuzda, D. Transcriptional profiling reveals developmental relationship and distinct biological functions of CD16_ and Cd16- monocyte subsets. BMC Genom. 2009, 10, 403. [Google Scholar] [CrossRef] [Green Version]
- Dale, D.; Boxer, L.; Liles, C. The phagocytes: Neutrophils and monocytes. Blood 2008, 112, 935–945. [Google Scholar] [CrossRef]
- Chuang, P.; Wu, M.; Shoji, Y.; Tsai, S. Downregulation of CD36 results in reduced phagocytic ability of peritoneal macrophages of women with endometriosis. J. Pathol. 2009, 219, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.V.; Connors, T.J.; Farber, D.L. Human T cell development, localization, and function throughout life. Immunity 2018, 48, 202–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguschi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T cells and immune tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takamura, M.; Koga, K.; Izumi, G.; Hirata, T.; Harada, M.; Hirota, Y.; Hiraike, O.; Fujii, T.; Osuga, Y. Simultaneous Detection and Evaluation of Four Subsets of CD4+ T Lymphocyte in Lesions and Peripheral Blood in Endometriosis. Am. J. Reprod. Immunol. 2015, 74, 480–486. [Google Scholar] [CrossRef]
- Olkowska-Truchanowicz, J.; Bocian, K.; Maksym, R.B.; Białoszewska, A.; Włodarczyk, D.; Baranowski, W.; Za̧bek, J.; Korczak-Kowalska, G.; Malejczyk, J. CD4+ CD25+ FOXP3+ regulatory T cells in peripheral blood and peritoneal fluid of patients with endometriosis. Hum. Reprod. 2013, 28, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Kurosaki, T. B-lymphocyte biology. Immunol. Rev. 2010, 237, 5–9. [Google Scholar] [CrossRef]
- Fernàndez-Shaw, S.; Hicks, B.R.; Yudkin, P.L.; Kennedy, S.; Barlow, D.H.; Starkey, P.M. Anti-endometrial and anti-endothelial auto-antibodies in women with endometriosis. Hum. Reprod. 1993, 8, 310–315. [Google Scholar] [CrossRef]
- Zhang, C.; Maeda, N.; Izumiya, C.; Yamamoto, Y.; Kusume, T.; Oguri, H.; Yamashita, C.; Nishimori, Y.; Hayashi, K.; Luo, J.; et al. Killer immunoglobulin-like receptor and human leukocyte antigen expression as immunodiagnostic parameters for pelvic endometriosis. Am. J. Reprod. Immunol. 2006, 55, 106–114. [Google Scholar] [CrossRef]
- Lavagna, A.; Auger, J.-P.; Giradin, S.E.; Gisch, N.; Segura, M.; Gottschalk, M. Recognition of Lipoproteins by Toll-like Receptor 2 and DNA by the AIM2 Inflammasome Is Responsible for Production of Interleukin-1β by Virulent Suilysin-Negative Streptococcus suis Serotype 2. Pathogens 2020, 9, 147. [Google Scholar] [CrossRef] [Green Version]
- Lillico, D.M.E.; Pemberton, J.G.; Stafford, J.L. Selective Regulation of Cytoskeletal Dynamics and Filopodia Formation by Teleost Leukocyte Immune-Type Receptors Differentially Contributes to Target Capture During the Phagocytic Process. Front. Immunol. 2018, 9, 1144. [Google Scholar] [CrossRef]
- Khan, K.N.; Kitajima, M.; Hiraki, K.; Fujishita, A.; Ishimaru, T.; Masuzaki, H. Toll-like receptors in innate immunity: Role of bacterial endotoxin and toll-like receptor 4 in endometrium and endometriosis. Gynecol. Obstet. Investig. 2009, 68, 40–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, K.-Y.; Liu, H.-Y.; Yan, W.-L.; Wu, C.-C.; Lee, M.-H.; Leng, C.-H.; Liu, S.-J. Liposomal TLR9 Agonist Combined with TLR2 Agonist-Fused Antigen Can Modulate Tumor Microenvironment through Dendritic Cells. Cancers 2020, 12, 810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira-Nascimento, L.; Massari, P.; Wetzler, L.M. The role of TLR2 in infection and immunity. Front. Immunol. 2012, 3, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, A.; Pergolizzi, S.; Cimino, F.; Lauriano, E.R.; Speciale, A.; D’Angelo, V.; Sicurella, M.; Argnani, R.; Manservigi, R.; Marconi, P. Role of Herpes Simplex Envelope Glycoprotein B and Toll-Like Receptor 2 in Ocular Inflammation: An Ex Vivo Organotypic Rabbit Corneal Model. Viruses 2019, 11, 819. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Han, Z.; Oppenheim, J.J. Alarmins and immunity. Immunol. Rev. 2017, 280, 41–56. [Google Scholar] [CrossRef]
- Costa Mendonça-Natividade, F.; Duque Lopes, C.; Ricci-Azevedo, R.; Sardinha-Silva, A.; Figueiredo Pinzan, C.; Paiva Alegre-Maller, A.C.; L. Nohara, L.; B. Carneiro, A.; Panunto-Castelo, A.; C. Almeida, I.; et al. Receptor Heterodimerization and Co-Receptor Engagement in TLR2 Activation Induced by MIC1 and MIC4 from Toxoplasma gondii. Int. J. Mol. Sci. 2019, 20, 5001. [Google Scholar] [CrossRef] [Green Version]
- McKinnon, B.; Mueller, M.D.; Nirgianakis, K.; Bersinger, N.A. Comparison of ovarian cancer markers in endometriosis favours HE4 over CA125. Mol. Med. Rep. 2015, 12, 5179–5184. [Google Scholar] [CrossRef] [Green Version]
- Van Gorp, T.; Cadron, I.; Despierre, E.; Daemen, A.; Leunen, K.; Amant, F.; Timmerman, D.; De Moor, B.; Vergote, I. HE4 and CA125 as a diagnostic test in ovarian cancer: Prospective validation of the Risk of Ovarian Malignancy Algorithm. Br. J. Cancer 2011, 104, 863–870. [Google Scholar] [CrossRef] [Green Version]
- Canis, M.; Donnez, J.G.; Guzick, D.S.; Halme, J.K.; Rock, J.A.; Schenken, R.S.; Vernon, M.W. Revised American Society for Reproductive Medicine classification of endometriosis: 1996. Fertil. Steril. 1997, 67, 817–821. [Google Scholar] [CrossRef]
- Schulke, L.; Berbic, M.; Manconi, F.; Tokushige, N.; Markhan, R.; Fraser, I.S. Dendriti cell populations in the eutopic and ectopic endometrium of women with endometriosis. Hum. Reprod. 2009, 24, 1695–16703. [Google Scholar] [CrossRef] [Green Version]
- Berbic, M.; Schulke, L.; Markham, R.; Tokushige, N.; Russell, P.; Fraser, I.S. Macrophage expression in endometrium of women with and without endometriosis. Hum. Reprod. 2009, 24, 325–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, P.C.; Lin, Y.J.; Wu, M.H.; Wing, L.Y.; Shoji, Y.; Tsai, S.J. Inhibition of CD36-dependent phagocytosis by prostaglandin E2 contributes to the development of endometriosis. Am. J. Pathol. 2010, 176, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Montagna, P.; Capellino, S.; Villaggio, B.; Remorgida, V.; Ragni, N.; Cutolo, M.; Ferrero, S. Peritoneal fluid macrophages in endometriosis: Correlation between expression of estrogen receptors and inflammation. Fertil. Steril. 2008, 90, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Lousse, J.C.; Vam Langendonckt, A.; González-Ramos, R.; Defrére, S.; Renkin, E.; Donnez, J. Increased activation of nuclear factor-kappa B (NF-kappaB) in isolated peritoneal macrophages of patients with endometriosis. Fertil. Steril. 2008, 90, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Akoum, A.; Kong, J.; Metz, C.; Beaumont, M.C. Spontaneous and stimulated secretion of monocyte chemotactic protein-1 and macrophage migration inhibitory factor by peritoneal macrophages with and without endometriosis. Fertil. Steril. 2002, 77, 989–994. [Google Scholar] [CrossRef]
- Hallamaa, M.; Huhtienen, K.; Suvitie, P.; Perheentupa, A. Serum concentrations of HE4 change little during in vitro fertilization. Acta Obstet. Gynecol. Scand. 2014, 93, 640–646. [Google Scholar] [CrossRef]
- Gogacz, M.; Winkler, I.; Bojarska-Junak, A.; Tabarkiewicz, J.; Semczuk, A.; Rechberger, T.; Adamiak, A. T regulatory lymphocytes in patients with endometriosis. Mol. Med. Rep. 2014, 10, 1072–1076. [Google Scholar] [CrossRef]
- Greaves, E.; Temp, J.; Esnal-Zufiurre, A.; Mechsner, S.; Horne, A.W.; Saunders, P.T. Estradiol is a criticalmediator of macrophage-nerve cross talk in peritoneal endometriosis. Am. J. Pathol. 2015, 185, 2286–2297. [Google Scholar] [CrossRef]
- Kobayashi, H.; Higashiura, Y.; Shigetomi, H.; Kajihara, H. Pathogenesis of endometriosis: The role of initial infection and subsequent sterile inflammation (Review). Mol. Med. Rep. 2014, 9, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Izawa, M.; Harada, T.; Deura, I.; Taniguchi, F.; Iwabe, T.; Terakawa, N. Drug-induced apoptosis was markedly attenuated in endometriotic stromal cells. Hum. Reprod. 2006, 21, 600–604. [Google Scholar] [CrossRef] [Green Version]
- Kajihara, H.; Yamada, Y.; Kanayama, S.; Furukawa, N.; Noguchi, T.; Haruta, S.; Yoshida, S.; Sado, T.; Oi, H.; Kobayashi, H. New insights into the pathophysiology of endometriosis: From chronic inflammation to danger signal. Gynecol. Endocrinol. 2011, 27, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Fazeli, A.; Bruce, C.; Anumba, D.O. Characterization of Toll-like receptors in the female reproductive tract in humans. Hum. Reprod. 2005, 20, 1372–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allhorn, S.; Böing, C.; Koch, A.A.; Kimming, R.; Gashaw, I. TLR3 and TLR4 expression in healthy and diseased human endometrium. Reprod. Biol. Endocrinol. 2008, 6, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, K.N.; Kitajima, M.; Hiraki, K.; Yamaguchi, N.; Katamine, S.; Matsuyama, T.; Nakashima, M.; Fujishita, A.; Ishimaru, T.; Masuzaki, H. Escherichia coli contamination of menstrual blood and effect of bacterial endotoxin on endometriosis. Fertil. Steril. 2010, 94, 2860–2863. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, T.M.; Desouza, K.; Fahey, J.V.; Beagley, K.W.; Wira, C.R. Toll-like receptor (TLR) expression and TLR-mediated cytokine/chemokine production by human uterine epithelial cells. Immunology 2004, 112, 428–436. [Google Scholar] [CrossRef]
- Piccinini, A.M.; Midwood, K.S. DAMPening inflammation by modulating TLR signaling. Mediat. Inflamm. 2010, 2010, 672395. [Google Scholar] [CrossRef] [Green Version]
- Matzinger, P. Tolerance, danger, and the extended family. Annu. Rev. Immunol. 1994, 12, 991–1045. [Google Scholar] [CrossRef]
- Zubor, P.; Hatok, J.; Galo, S.; Dokus, K.; Klobusiakova, D.; Danko, J.; Racay, P. Anti-apoptotic and pro-apoptotic gene expression evaluated from eutopic endometrium in the proliferative phase of the menstrual cycle among women with endometriosis and healthy controls. Eur. J. Obstet. Gynecol. Reprod. Biol. 2009, 145, 172–176. [Google Scholar] [CrossRef]
- Dmowski, W.P.; Ding, J.; Shen, J.; Rana, N.; Fernandez, B.B.; Braun, D.P. Apoptosis in endometrial glandular and stromal cells in women with and without endometriosis. Hum. Reprod. 2001, 16, 1802–1808. [Google Scholar] [CrossRef] [Green Version]
- Braun, D.P.; Ding, J.; Shen, J.; Rana, N.; Fernandez, B.B.; Dmowski, W.P. Relationship between apoptosis and the number of macrophages in eutopic endometrium from women with and without endometriosis. Fertil. Steril. 2002, 78, 830–835. [Google Scholar] [CrossRef]
- Bokor, A.; Kyama, C.M.; Vercruysse, L.; Fassbender, A.; Gevaert, O.; Vodolazkaia, A.; De Moor, B.; Fülöp, V.; D’Hooghe, T. Density of small diameter sensory nerve fibres in endometrium: A semi-invasive diagnostic test for minimal to mild endometriosis. Hum. Reprod. 2009, 24, 3025–3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Jefout, M.; Dezarnaulds, G.; Cooper, M.; Tokushige, N.; Luscombe, G.M.; Markham, R.; Fraser, I.S. Diagnosis of endometriosis by detection of nerve fibres in an endometrial biopsy: A double blind study. Hum. Reprod. 2009, 24, 3019–3024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mol, B.W.J.; Bayram, N.; Lijmer, J.G.; Wiegerinck, M.A.H.M.; Bongers, M.Y.; Van Der Veen, F.; Bossuyt, P.M.M. The performance of CA-125 measurement in the detection of endometriosis: A meta-analysis. Fertil. Steril. 1998, 70, 1101–1108. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Endometriosis (n = 40) | Control Group (n = 40) | p-Value |
|---|---|---|---|
| Mean ± SD/Median (Range) | Mean ± SD/Median (Range) | ||
| Age [years] | 35.5 ± 8.0 | 36.1 ± 10.7 | NS |
| Ca-125 [U/mL] | 31.8 (10.1–126.1) | 13.5 (2.0–26.3) | <0.0001 |
| HE4 [pmol/L] | 38.7 (24.0–73.0) | 32.6 (11.0–55.7) | 0.025 |
| Clinical features | N (%) | N (%) | |
| Stage of endometriosis: | N/A | ||
| I | 10 (25) | N/A | |
| II | 12 (30) | N/A | |
| III | 7 (17.5) | N/A | |
| IV | 11 (27.5) | N/A | |
| Adhesions | 18 (45) | N/A | |
| Dysmenorrhea | 26 (65) | N/A | |
| Infertility | 14 (35) | N/A |
| Parameter [%] | Endometriosis Group (n = 40) | Healthy Control Group (n = 40) | t/Z Value | p-Value |
|---|---|---|---|---|
| Mean ± SD/Median (Range) | Mean ± SD/Median (Range) | |||
| Myeloid Dendritic Cells BDCA1+CD19- | 0.28 ± 0.2 | 0.41 ± 0.15 | −3.20 | 0.0014 |
| Plasmacytoid dendritic cells BDCA2+CD123+ | 0.32 ± 0.2 | 0.29 ± 0.17 | 0.39 | 0.69 |
| Myeloid dendritic cells BDCA1+CD19−/Plasmacytoid dendritic cells BDCA2+CD123+ ratio | 0.88 (0.04–5.0) | 1.7 (0.31–6.3) | −3.13 | 0.0017 |
| Classical monocytes CD14+CD16− | 85.1 ± 5.5 | 90.9 ± 3.5 | −4.76 | <0.0001 |
| Non classical monocytes CD14+CD16+ | 10.7 ± 4.9 | 5.3 ± 2.5 | 5.00 | <0.0001 |
| T lymphocytes CD3+ | 73.0 (61.9−78.8) | 72.4 (61.6−79.7) | 0.51 | 0.61 |
| B lymphocytes CD19+ | 11.5 ± 3.2 | 11.7 ± 2.7 | −0.48 | 0.63 |
| NK cells CD3−CD16+CD56+ | 11.0 ± 4.2 | 14.7 ± 3.4 | −3.80 | 0.0001 |
| NKT-like cells CD3+CD16+CD56+ | 1.7 (0.24–11.3) | 3.3 (1.2–4.9) | −1.66 | 0.096 |
| T lymphocytes CD3+CD4+ | 40.2 (26.6–53.1) | 40.3 (33.4–57.4) | −0.37 | 0.71 |
| T lymphocytes CD3+CD8+ | 29.3 ± 5.5 | 30.2 ± 4.9 | −1.20 | 0.23 |
| T lymphocytes ratio CD3+CD4+/T CD3+CD8+ | 1.4 (0.73–2.7) | 1.3 (0.93–4.5) | 0.54 | 0.59 |
| Myeloid dendritic cells BDCA1+CD19-TLR2+ | 21.3 (6.1–48.0) | 3.5 (1.1–9.3) | 7.45 | <0.0001 |
| Plasmacytoid dendritic cells BDCA2+CD123+ TLR2+ | 12.7 (2.5–43.9) | 5.8 (0.98–14.3) | 4.13 | <0.0001 |
| Classical monocytes CD14+CD16-TLR2+ | 8.3 ± 4.8 | 4.7 ± 2.1 | 3.37 | 0.0007 |
| Non classical monocytes CD14+CD16+TLR2+ | 10.1 (1.5–45.6) | 4.4 (0.66–11.6) | 4.16 | <0.0001 |
| T lymphocytes CD4+TLR-2+ | 3.8 (0.47–40.1) | 0.92 (0.04–4.1) | 5.47 | <0.0001 |
| T lymphocytes CD8+TLR-2+ | 2.4 (0.1–59.4) | 1.0 (0.08–6.5) | 2.78 | 0.0054 |
| B lymphocytes CD19+TLR-2+ | 4.2 (0.42–23.5) | 2.6 (0.14–7.0) | 2.67 | 0.0077 |
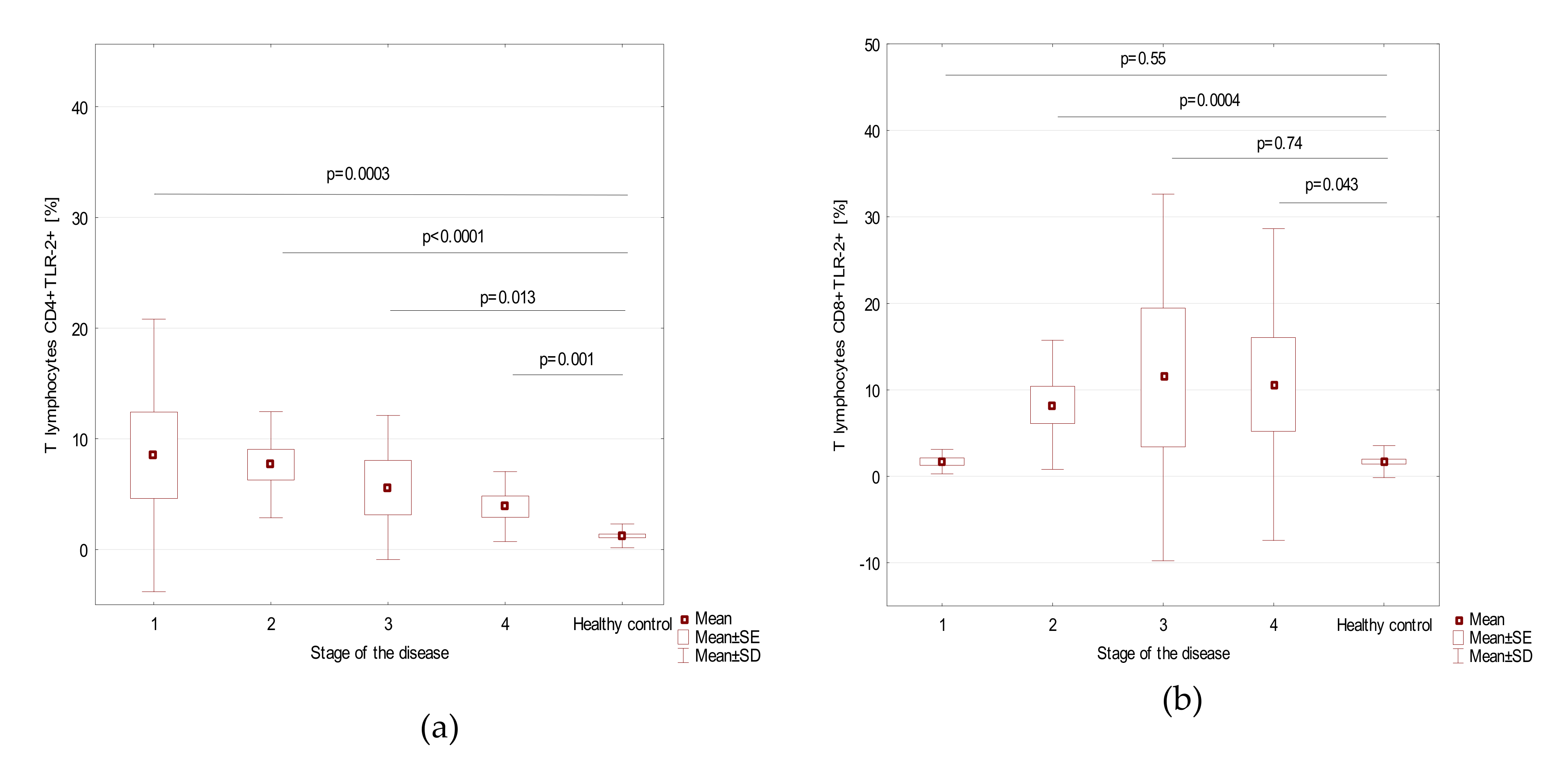
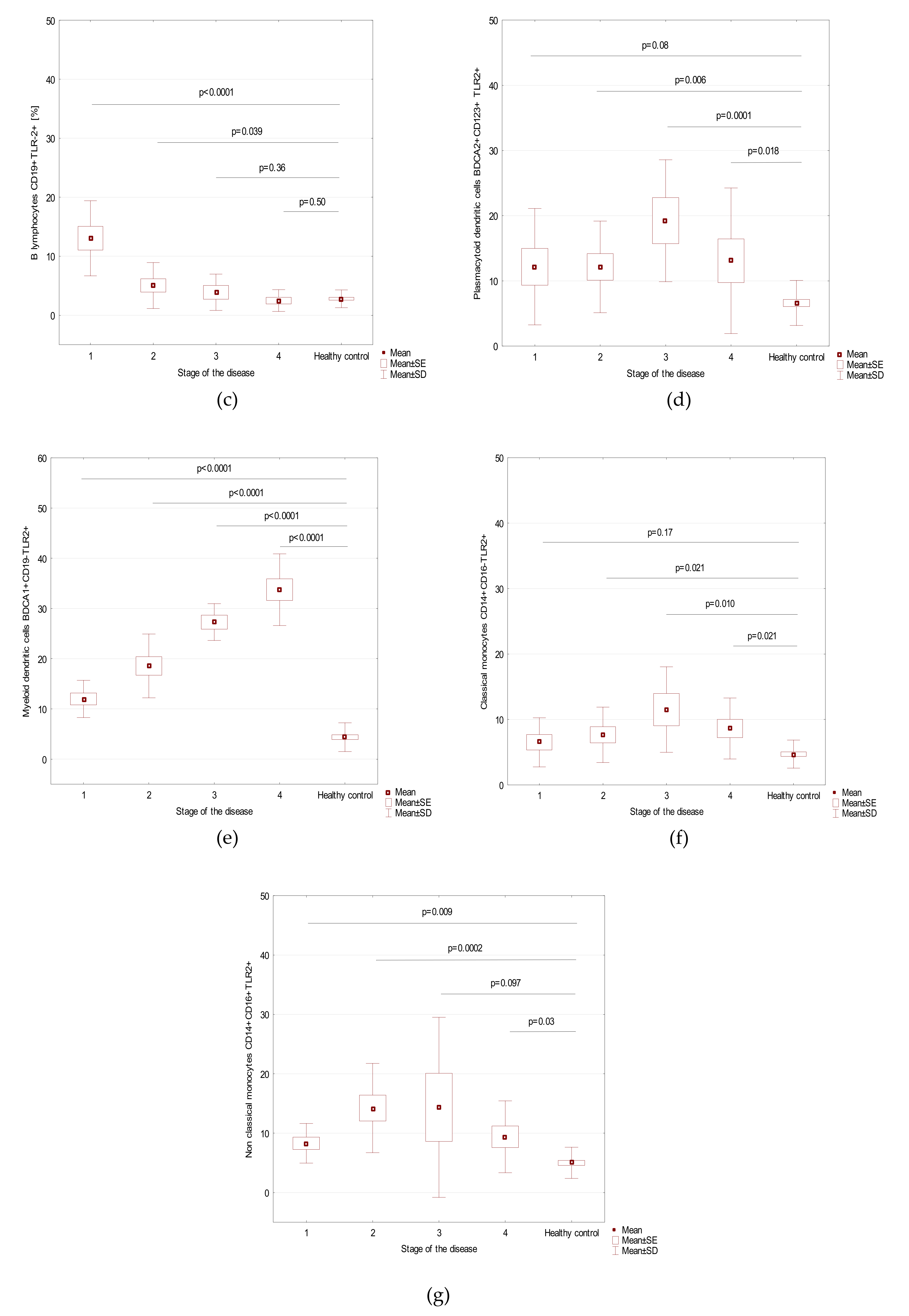
| Parameter | Stage of the Disease (Mean + SD) | p-Value | |||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ||
| Myeloid dendritic cells BDCA1+CD19- | 0.26 ± 0.21 | 0.34 ± 0.26 | 0.21 ± 0.11 | 0.29 ± 0.17 | 0.80 |
| Plasmacytoid dendritic cells BDCA2+CD123+ | 0.41 ± 0.21 | 0.21 ± 0.14 | 0.32 ± 0.2 | 0.36 ± 0.22 | 0.09 |
| Myeloid dendritic cells BDCA1+CD19−/Plasmacytoid dendritic cells BDCA2+CD123+ ratio | 0.80 ± 0.68 | 1.9 ± 1.5 | 0.73 ± 0.18 | 1.0 ± 0.5 | 0.13 |
| Classical monocytes CD14+CD16- | 87.4 ± 3.1 | 87.1 ± 5.1 | 83.6 ± 3.5 | 81.9 ± 7.0 | 0.064 |
| Non-classical monocytes CD14+CD16+ | 8.6 ± 4.2 | 9.5 ± 5.0 | 10.8 ± 2.7 | 13.7 ± 5.5 | 0.09 |
| T lymphocytes CD3+ [%] | 71.5 ± 4.6 | 73.1 ± 2.8 | 69.6 ± 6.1 | 71.8 ± 2.9 | 0.47 |
| B lymphocytes CD19+ [%] | 12.2 ± 3.2 | 11.1 ± 2.9 | 12.7 ± 3.3 | 10.5 ± 3.4 | 0.37 |
| NK cells CD3-CD16+CD56+ [%] | 11.2±5.8 | 9.9±3.8 | 10.5 ± 4.6 | 12.4 ± 2.9 | 0.54 |
| NKT-like cells CD3+CD16+CD56+ [%] | 2.4±2.1 | 3.1±3.0 | 2.7 ± 2.7 | 4.1 ± 3.5 | 0.56 |
| T lymphocytes CD3+CD4+ [%] | 41.7±6.7 | 39.8±3.9 | 36.3 ± 3.2 | 42.6 ± 7.2 | 0.076 |
| T lymphocytes CD3+CD8+ [%] | 28.0 ± 4.7 | 30.8 ± 5.2 | 31.6 ± 7.7 | 27.2 ± 4.4 | 0.28 |
| T lymphocyte ratio CD3+CD4+/T CD3+CD8+ | 1.6 ± 0.5 | 1.3 ± 0.37 | 1.2 ± 0.39 | 1.6 ± 0.5 | 0.21 |
| Myeloid dendritic cells BDCA1+CD19−TLR2+ | 12.0 ± 3.7 | 18.6 ± 6.4 | 27.3 ± 3.7 | 33.7 ± 7.1 | <0.0001 |
| Plasmacytoid dendritic cells BDCA2+CD123+ TLR2+ | 12.2 ± 8.9 | 12.1 ± 7.0 | 19.2 ± 9.4 | 13.1 ± 11.2 | 0.19 |
| Classical monocytes CD14+CD16-TLR2+ | 6.5 ± 3.7 | 7.7 ± 4.2 | 11.5 ± 6.5 | 8.6 ± 4.7 | 0.38 |
| Non-classical monocytes CD14+CD16+TLR2+ | 8.3 ± 3.3 | 14.2 ± 7.5 | 14.4 ± 15.2 | 9.4 ± 6.0 | 0.21 |
| T lymphocytes CD4+TLR-2+ [%] | 8.5 ± 12.3 | 7.7 ± 4.8 | 5.6 ± 6.5 | 3.9 ± 3.2 | 0.36 |
| T lymphocytes CD8+TLR-2+ [%] | 1.7 ± 1.4 | 8.3 ± 7.5 | 11.4 ± 21.2 | 10.6 ± 18.0 | 0.10 |
| B lymphocytes CD19+TLR-2+ [%] | 13.0 ± 6.4 | 5.0 ± 3.9 | 3.9 ± 3.1 | 2.5 ± 1.8 | 0.0007 |
| Factor | Parameter [%] | Prognostic Value | Youden Index | Area under the Curve (AUC) | 95% CI | p-Value |
|---|---|---|---|---|---|---|
| Stages 3–4 | Myeloid dendritic cells BDCA1+CD19-TLR2+ | 22.22 | 0.91 | 0.96 | 0.89–1.0 | <0.0001 |
| B lymphocytes CD19+TLR-2+ | 5.62 | 0.49 | 0.78 | 0.64–0.95 | 0.0001 | |
| Adhesion | Myeloid dendritic cells BDCA1+CD19-TLR2+ | 20.45 | 0.81 | 0.92 | 0.83–1.0 | <0.0001 |
| B lymphocytes CD19+TLR-2+ | 6.16 | 0.59 | 0.82 | 0.70–0.95 | <0.0001 | |
| Non classical monocytes CD14+CD16+TLR2+ | 10.0 | 0.39 | 0.70 | 0.53–0.87 | 0.018 | |
| Infertility | Myeloid dendritic cells BDCA1+CD19-TLR2+ | 30.51 | 0.60 | 0.83 | 0.68–0.98 | <0.0001 |
| Plasmacytoid dendritic cells BDCA2+CD123+ TLR2+ | 12.5 | 0.55 | 0.71 | 0.55–0.88 | 0.009 | |
| B lymphocytes CD19+TLR-2+ | 2.95 | 0.41 | 0.73 | 0.57–0.89 | 0.006 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobstyl, M.; Niedźwiedzka-Rystwej, P.; Grywalska, E.; Korona-Głowniak, I.; Sobstyl, A.; Bednarek, W.; Roliński, J. Toll-Like Receptor 2 Expression as a New Hallmark of Advanced Endometriosis. Cells 2020, 9, 1813. https://doi.org/10.3390/cells9081813
Sobstyl M, Niedźwiedzka-Rystwej P, Grywalska E, Korona-Głowniak I, Sobstyl A, Bednarek W, Roliński J. Toll-Like Receptor 2 Expression as a New Hallmark of Advanced Endometriosis. Cells. 2020; 9(8):1813. https://doi.org/10.3390/cells9081813
Chicago/Turabian StyleSobstyl, Małgorzata, Paulina Niedźwiedzka-Rystwej, Ewelina Grywalska, Izabela Korona-Głowniak, Anna Sobstyl, Wiesława Bednarek, and Jacek Roliński. 2020. "Toll-Like Receptor 2 Expression as a New Hallmark of Advanced Endometriosis" Cells 9, no. 8: 1813. https://doi.org/10.3390/cells9081813