Influence of Cathodic Water Invigoration on the Emergence and Subsequent Growth of Controlled Deteriorated Pea and Pumpkin Seeds

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Controlled Deterioration of Seeds
2.2. Preparation of Cathodic Water and Seed Priming
2.3. Plant Management/Experimental Design
2.4. Seedling Emergence
2.5. Physiological Measurements
2.6. Harvesting and Plant Tissue Analyses
2.7. Determination of MDA and 4-HNE
2.8. Quantification of Antioxidant Enzymes
2.9. Statistical Analyses
3. Results
3.1. Effect of Priming on Seedling Emergence and Growth
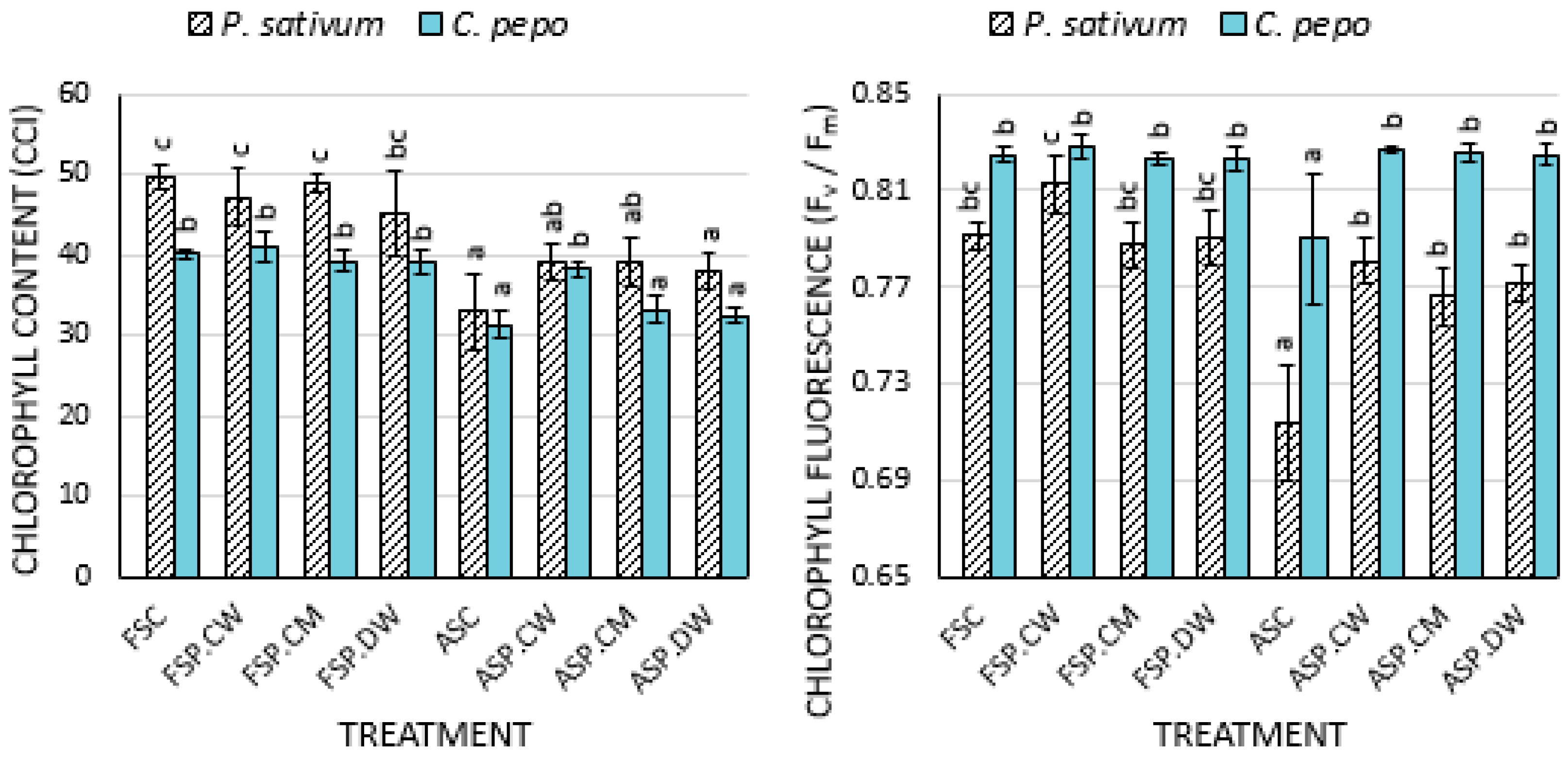
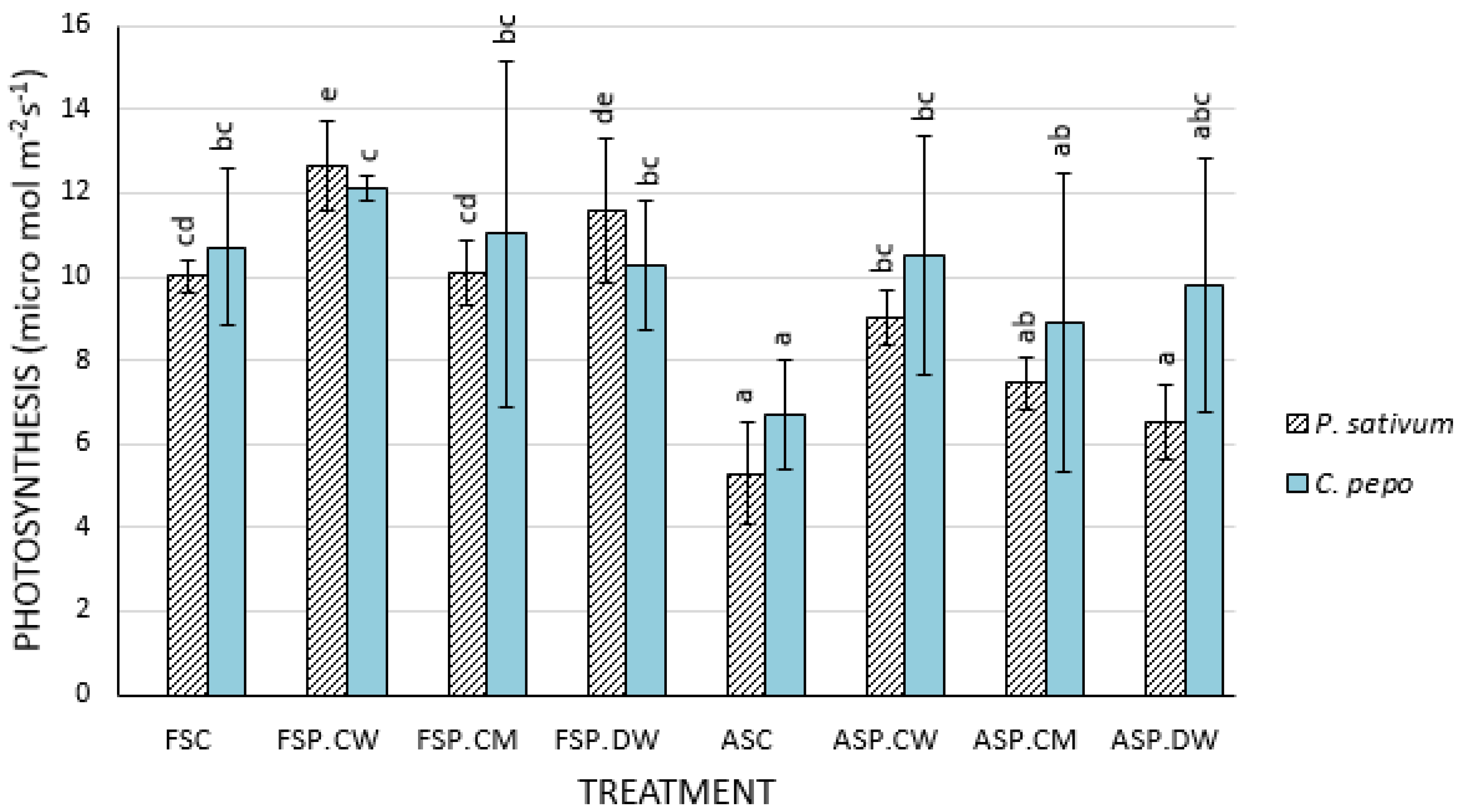
3.2. Effect of Priming on Physiological Parameters
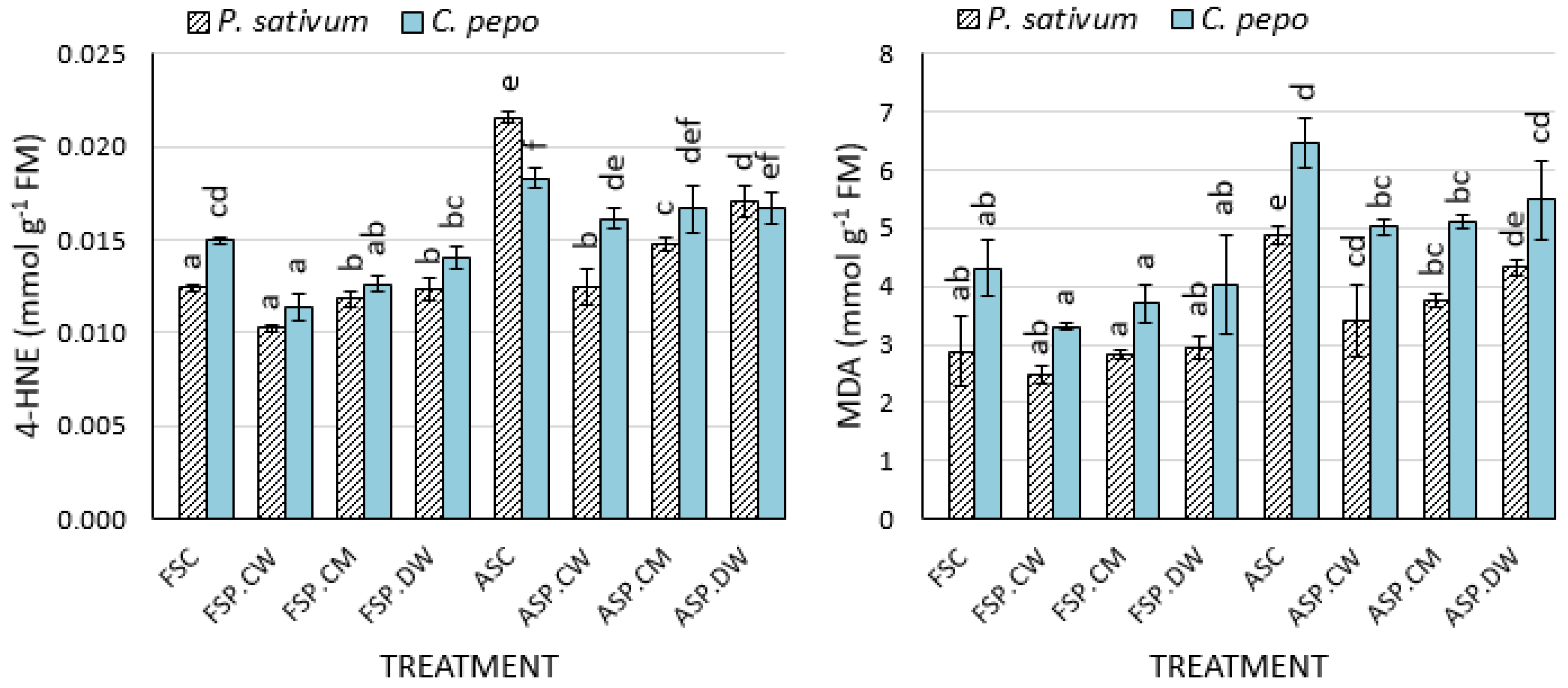
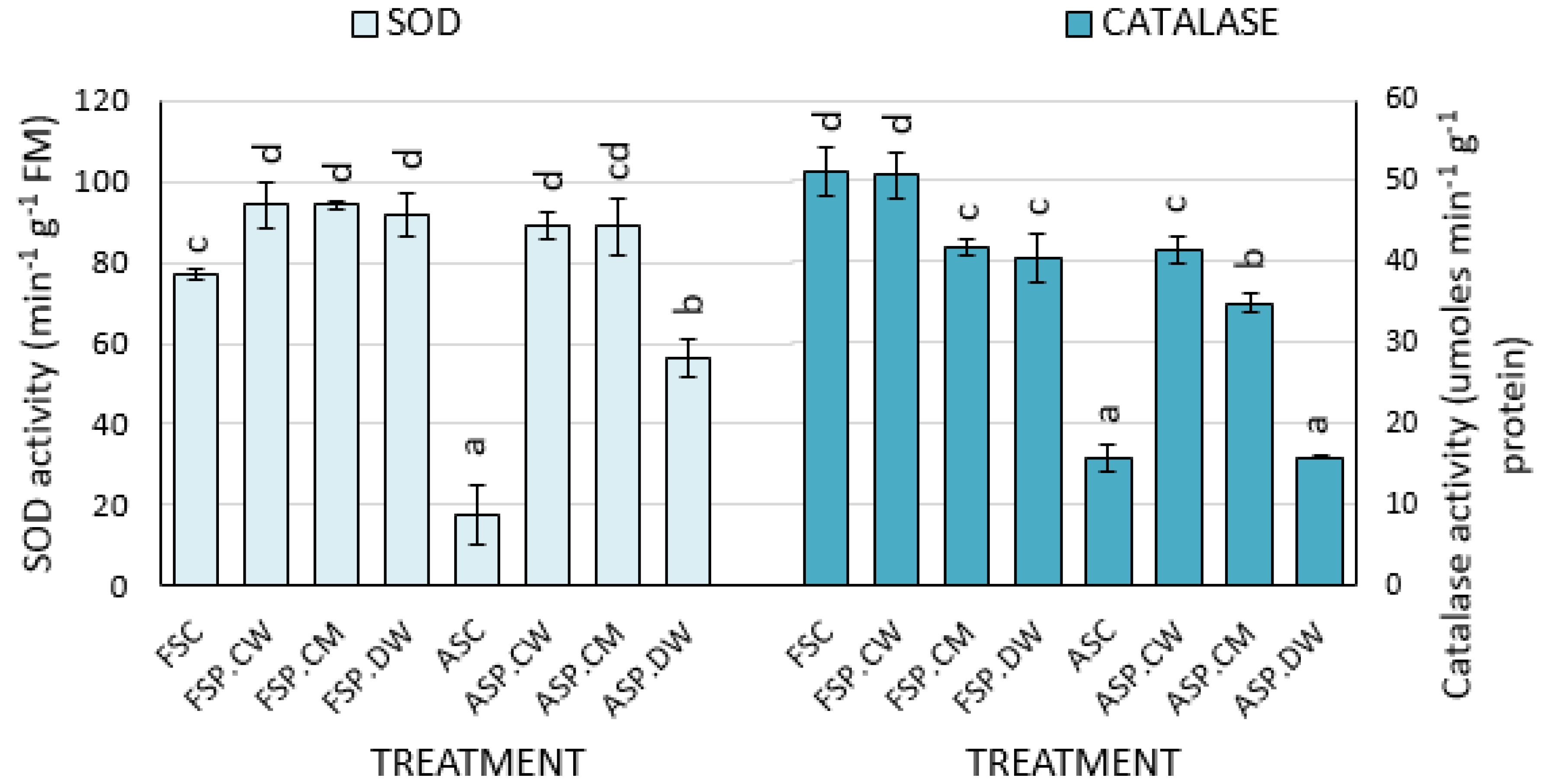
3.3. Effect of Priming on Lipid Peroxidation and Antioxidant Enzymes
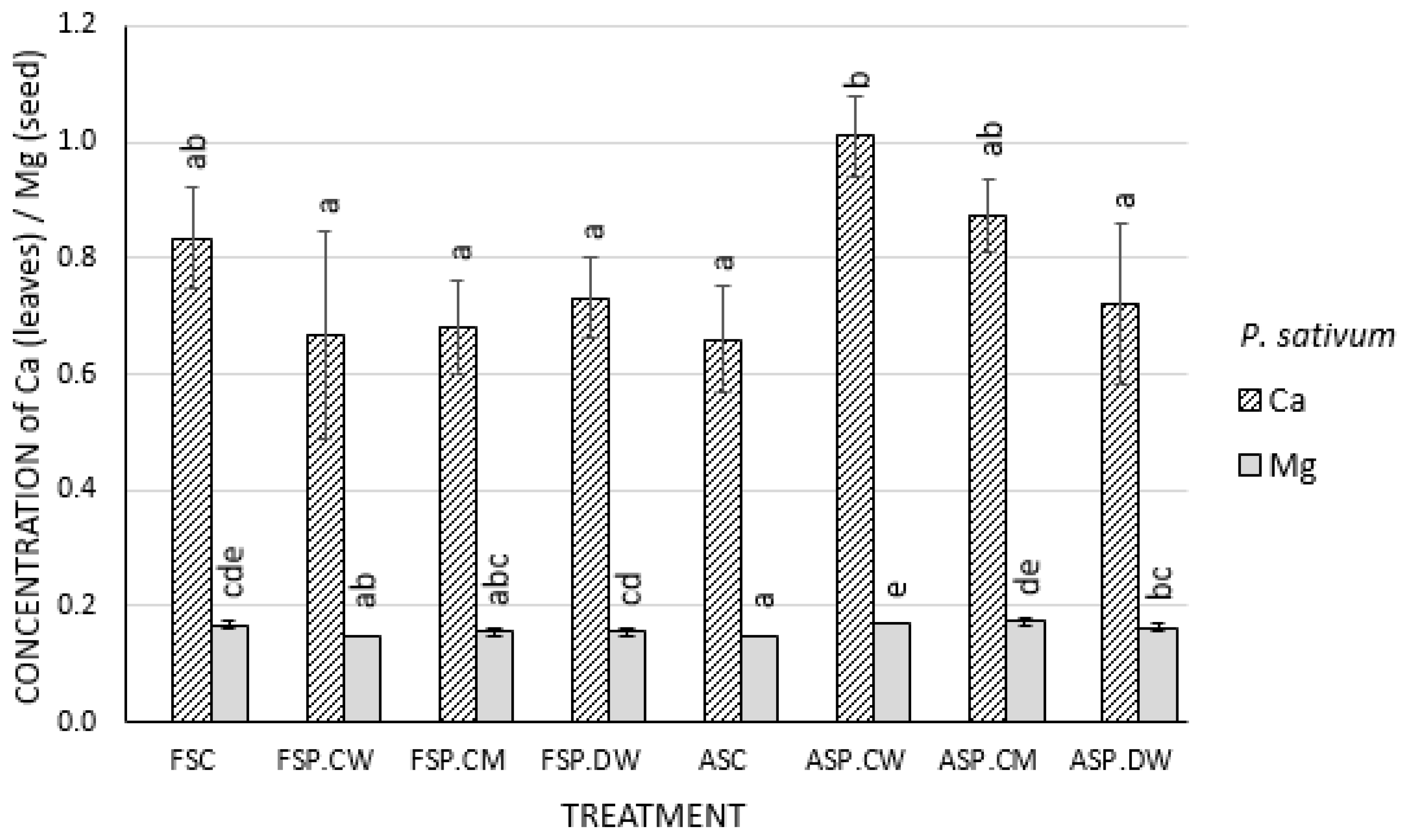
3.4. Effect of Cathodic Water, Calcium Magnesium Solution and Deionized Water on Mineral Content of Seeds and Shoot of Pisum sativum and Cucurbita pepo
4. Discussion
4.1. Effects of Priming on Emergence and Later Seedling Growth
4.2. Effects of Priming on Photosynthetic Parameters
4.3. Effects of Priming on Lipid Peroxidation Products, ROS Scavenging Enzymes and Composition
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Garza-Caligaris, L.E.; Avendaño-Vázquez, A.O.; Alvarado-López, S.; Zúñiga-Sánchez, E.; Orozco-Segovia, A.; Pérez-Ruíz, R.V.; Gamboa-Debuen, A. At3g08030 transcript: A molecular marker of seed ageing. Ann. Bot. 2012, 110, 1253–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desheva, G. The Longevity of Crop Seeds Stored Under Long-term Condition in the National Gene Bank of Bulgaria. Agriculture (Polnohospodárstvo) 2016, 62, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Andjelkovic, V.; Nikolic, A.; Kovacevic, D.; Mladenovic-Drinic, S.; Kravić, N.; Babić, V.; Srebric, M.; Jankulovska, M.; Ivanovska, S.; Bosev, D. Conserving maize in gene banks: Changes in genetic diversity revealed by morphological and SSR markers. Chil. J. Agric. Res. 2018, 78, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Skorupińska, A.; Kruczyńska, D.E.; Konopacka, D.; Kaczmarek, U.; Rutkowski, K.P.; Jóźwiak, Z.B. Fruit Quality and Storability of Some Asian Pear Cultivars Collected in the Gene Bank of the Research Institute of Horticulture, Central Poland. J. Hortic. Res. 2019, 27, 55–64. [Google Scholar] [CrossRef]
- Jorgenson, A.K.; Burns, T.J. Effects of Rural and Urban Population Dynamics and National Development on Deforestation in Less-Developed Countries, 1990–2000. Sociol. Inq. 2007, 77, 460–482. [Google Scholar] [CrossRef]
- FAO. The State of Food Security and Nutrition in the World; The Food and Agriculture Organization of the United Nations: Rome, Italy, 2018. [Google Scholar]
- Amanpour-Balaneji, B.; Sedghi, M. Effect of Aging and Priming on Physiological and Biochemical Traits of Common Bean (Phaseolus vulgaris L.). Not. Sci. Biol. 2012, 4, 95–100. [Google Scholar] [CrossRef]
- Sreepriya, S.; Girija, T. Seed invigoration- a technique for improving vigour and productivity of sesame (Sesamum indicum L.) variety Thilak. J. Trop. Agric. 2019, 57, 10. [Google Scholar]
- Ullah, A.; Farooq, M.; Hussain, M.; Ahmad, R.; Wakeel, A. Zinc Seed Priming Improves Stand Establishment, Tissue Zinc Concentration and Early Seedling Growth of Chickpea. JAPS J. Anim. Plant Sci. 2019, 29, 1046–1053. [Google Scholar]
- Berjak, P.; Pammenter, N.W. Effects of cryopreservation of recalcitrant Amaryllis belladonna zygotic embryos on vigour of recovered seedlings: A case of stress ‘hangover? Physiol. Plant. 2010, 139, 205–219. [Google Scholar]
- Wang, Y.; Jie, W.; Peng, X.; Hua, X.; Yan, X.; Zhou, Z.; Lin, J. Physiological Adaptive Strategies of Oil Seed Crop Ricinus communis Early Seedlings (Cotyledon vs. True Leaf) Under Salt and Alkali Stresses: From the Growth, Photosynthesis and Chlorophyll Fluorescence. Front. Plant Sci. 2019, 9, 9. [Google Scholar] [CrossRef]
- Chhabra, R.; Singh, T. Seed Aging, Storage and Deterioration: An Irresistible Physiological Phenomenon. Agric. Rev. 2019, 40, 234–238. [Google Scholar] [CrossRef] [Green Version]
- Sahu, B.; Sahu, A.; Thomas, V.; Naithani, S.C. Reactive oxygen species, lipid peroxidation, protein oxidation and antioxidative enzymes in dehydrating Karanj (Pongamia pinnata) seeds during storage. South Afr. J. Bot. 2017, 112, 383–390. [Google Scholar] [CrossRef]
- Da Silva, L.J.; Dias, D.C.F.D.S.; Sekita, M.C.; Finger, F.L. Lipid peroxidation and antioxidant enzymes of Jatropha curcas L. seeds stored at different maturity stages. Acta Sci. Agron. 2018, 40, 34978. [Google Scholar] [CrossRef] [Green Version]
- Chandrakar, V.; Naithani, S.C.; Keshavkant, S. Arsenic-induced metabolic disturbances and their mitigation mechanisms in crop plants: A review. Biologia 2016, 71, 367–377. [Google Scholar] [CrossRef]
- Karalija, E.; Selović, A. The effect of hydro and proline seed priming on growth, proline and sugar content, and antioxidant activity of maize under cadmium stress. Environ. Sci. Pollut. Res. 2018, 25, 33370–33380. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Guha, P.; Mandal, A.K. Deteriorative changes in enzyme activity of non-invigorated and invigorated soybean seeds (Glycine max [L.] Merrill, cv. Soyamax). Legum. Res. Int. J. 2018, 42, 633–639. [Google Scholar] [CrossRef] [Green Version]
- Selvi, D.T.; Saraswathy, S. Seed viability, seed deterioration and seed quality improvements in stored onion seeds: A review. J. Hortic. Sci. Biotechnol. 2017, 93, 1–7. [Google Scholar] [CrossRef]
- Heydecker, W.; Higgins, J.; Gulliver, R.L. Accelerated Germination by Osmotic Seed Treatment. Nature 1973, 246, 42–44. [Google Scholar] [CrossRef]
- Heydecker, W.; Coolbear, P. Seed treatments for improved performance-survey and attempted prognosis. Seed Sci. Technol. 1977, 5, 353–425. [Google Scholar]
- Bradford, K.J. Manipulation of Seed Water Relations via Osmotic Priming to Improve Germination under Stress Conditions; HortScience: Alexandria, VA, USA, 1986. [Google Scholar]
- Ashraf, M.; Foolad, M. Pre-Sowing Seed Treatment—A Shotgun Approach to Improve Germination, Plant Growth, and Crop Yield Under Saline and Non-Saline Conditions. Adv. Agron. 2005, 88, 223–271. [Google Scholar] [CrossRef]
- Fatokun, K.; Beckett, R.P.; Varghese, B.; Sershen, B.; Pammenter, N.W. Germination indices of orthodox seeds as influenced by controlled deterioration and cathodic water seed invigoration. J. Environ. Biol. 2020. accepted. [Google Scholar]
- Ella, E.S.; Dionisio-Sese, M.L.; Ismail, A.M. Seed pre-treatment in rice reduces damage, enhances carbohydrate mobilization and improves emergence and seedling establishment under flooded conditions. AoB PLANTS 2011, 2011, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umair, A.; Abid, M.; Ali, I.; Bashir, K.; Naseem, W.; Rauf, H.A. Nutrient priming in different maize cultivars and evaluation of vigour improvement under controlled conditions. Pak. J. Sci. Ind. Res. Ser. A Phys. Sci. 2017, 60, 69–73. [Google Scholar]
- Muhammad, I.; Volker, R.; Günter, N. Accumulation and Distribution of Zn and Mn in Soybean Seeds after Nutrient Seed Priming and its Contribution to Plant Growth under Zn and Mn Deficient Conditions. J. Plant Nutr. 2016, 40, 695–708. [Google Scholar] [CrossRef]
- Berjak, P.; Sershen, B.; Varghese, B.; Pammenter, N. Cathodic amelioration of the adverse effects of oxidative stress accompanying procedures necessary for cryopreservation of embryonic axes of recalcitrant-seeded species. Seed Sci. Res. 2011, 21, 187–203. [Google Scholar] [CrossRef]
- Mycock, D.J.; Pammenter, N.W.; Berjak, P. Improved germination performance of aged maize seed in the presence of cathodic water. In Proceedings of the 2013 International Seed Testing Association Congress, Bassersdorf, Switzerland, 8–10 September 2013. [Google Scholar]
- Gondwe, D.; Berjak, P.; Pammenter, N.; Sershen, B.; Varghese, B. Effect of priming with cathodic water and subsequent storage on invigoration of Pisum sativum, Cucurbita maxima and Lycopersicon esculentum seeds. Seed Sci. Technol. 2016, 44, 1–12. [Google Scholar] [CrossRef]
- Moura, M.L.D.S.; Chagas, E.A.; Smiderle, O.J.; Vilaça, R.; Farias, E.E. Biometric characterization, water absorption curve and vigor on araçá-boi seeds. Int. J. Plant Biol. 2016, 7, 22–25. [Google Scholar] [CrossRef] [Green Version]
- Mycock, D.J. Addition of calcium and magnesium to a glycerol and sucrose cryoprotectant solution improves the quality of plant embryo recovery from cryostorage. Cryo Lett. 1999, 20, 77–82. [Google Scholar]
- Smith, T. The Profitable culture of vegetables for market gardeners, small holders, and others. In The Profitable Culture of Vegetables for Market Gardeners, Small Holders, and Others; Smithsonian Institution: Washington, DC, USA, 1913; p. 9781429014755. [Google Scholar]
- Czabator, F.J. Germination value: An index combining speed and completeness of pine seed germination. Forest Sci. 1962, 8, 386–396. [Google Scholar]
- Ellis, R.A.; Roberts, E.R. The quantification of ageing and survival in orthodox seeds. Seed Sci. Technol. 1981, 9, 373–409. [Google Scholar]
- Abdollahi, M.; Eshghi, S.; Tafazzoli, E.; Moosavi, N. Effects of paclobutrazol, boric acid and zinc sulfate on vegetative and reproductive growth of strawberry cv. Selva. J. Agric. Sci. Technol. 2012, 14, 357–363. [Google Scholar]
- Hodges, D.M.; Delong, J.M.; Forney, C.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Ray, S.; Roy, K.; Sengupta, C. Evaluation of protective effects of water extract of Spirulina platensis on cisp;atin-induced lipid peroxidation. Indian J. Pharm. Sci. 2007, 69, 378–383. [Google Scholar]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the auto oxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Maehly, A.C.; Chance, B. The assay of catalases and peroxidases. Methods Biochem. Anal. 1954, 1, 764–817. [Google Scholar]
- Bewley, J.D.; Black, M. Seeds physiology of development and germination. In The Language of Science; Plenum Press: New York, NY, USA, 1994; p. 230. [Google Scholar]
- Ghassemi-Golezani, K.; Dalil, B. Effects of Seed Vigor on Growth And Grain Yield of Maize. Plant Breed. Seed Sci. 2014, 70, 81–90. [Google Scholar] [CrossRef]
- Anwar, P.; Juraimi, A.S.; Puteh, A.; Selamat, A.; Rahman, M.; Samedani, B. Seed priming influences weed competitiveness and productivity of aerobic rice. Acta Agric. Scand. Sect. B Plant Soil Sci. 2012, 62, 1–11. [Google Scholar] [CrossRef]
- Ahmed, N. Seed priming with glycine betaine improve seed germination characteristics and antioxidant capacity of wheat (triticum aestivum L.) Seedlings under water-stress conditions. Appl. Ecol. Environ. Res. 2019, 17, 8333–8350. [Google Scholar] [CrossRef]
- Wang, W.; He, A.; Peng, S.; Huang, J.; Cui, K.; Nie, L. The Effect of Storage Condition and Duration on the Deterioration of Primed Rice Seeds. Front. Plant Sci. 2018, 9, 172. [Google Scholar] [CrossRef]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- McDonald, M.B. Seed deterioration: Physiology, repair and assessment. Seed Sci. Technol. 1999, 27, 177–237. [Google Scholar]
- Balestrazzi, A.; Confalonieri, M.; Macovei, A.; Carbonera, D. Seed imbibition in Medicago truncatula Gaertn: Expression profiles of DNA repair genes in relation to PEG-mediated stress. J. Plant Physiol. 2011, 168, 706–713. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.B.; Gomes-Filho, E.; Enéas-Filho, J.; Prisco, J.T.; Alencar, N.L.M. Seed priming effects on growth, lipid peroxidation, and activity of ROS scavenging enzymes in NaCl-stressed sorghum seedlings from aged seeds. J. Plant Interact. 2012, 7, 151–159. [Google Scholar] [CrossRef]
- Goltsev, V.N.; Kalaji, H.M.; Paunov, M.; Bąba, W.; Horaczek, T.; Mojski, J.; Kociel, H.; Allakhverdiev, S.I. Variable chlorophyll fluorescence and its use for assessing physiological condition of plant photosynthetic apparatus. Russ. J. Plant Physiol. 2016, 63, 869–893. [Google Scholar] [CrossRef]
- Li, Z.; Li, Z.; Lu, K.; Liu, Z.M.; Wu, L.L. The effect of CaCl2 on calcium content, photosynthesis, and chlorophyll fluorescence of tung tree seedlings under drought conditions. Photosynthetica 2016, 55, 553–560. [Google Scholar] [CrossRef]
- He, L.; Yu, L.; Li, B.; Du, N.; Guo, S.-R. The effect of exogenous calcium on cucumber fruit quality, photosynthesis, chlorophyll fluorescence, and fast chlorophyll fluorescence during the fruiting period under hypoxic stress. BMC Plant Biol. 2018, 18, 180. [Google Scholar] [CrossRef]
- Fatokun, K.; Zharare, G.E. Influence of diesel contamination in soil on growth and dry matter partitioning of Lactuca sativa and Ipomoea batatas. J. Environ. Biol. 2015, 36, 1205–1213. [Google Scholar]
- Mouradi, M.; Bouizgaren, A.; Farissi, M.; Latrach, L.; Qaddoury, A.; Ghoulam, C. Seed osmopriming improves plant growth, nodulation, chlorophyll fluorescence and nutrient uptake in alfalfa (Medicago sativa L.)–rhizobia symbiosis under drought stress. Sci. Hortic. 2016, 213, 232–242. [Google Scholar] [CrossRef]
- Varghese, B.; Sershen, B.; Berjak, P.; Varghese, D.; Pammenter, N.W. Differential drying rate of recalcitrant Trichilia dregeana embryonic axes: A study of survival and oxidative stress mechanism. Physiol. Plant. 2011, 142, 326–338. [Google Scholar] [CrossRef]
- Hochmuth, G.; Maynard, D.; Vavrina, C.; Hanlon, E.; Simonne, E. Plant Tissue Analysis and Interpretation for Vegetable Crops in Florida; University of Florida: Gainesville, FL, USA, 2012. [Google Scholar]
- Demirkaya, M.; Dietz, K.; Sivritepe, H. Changes in Antioxidant Enzymes during ageing of onion seeds. Not. Bot. Horti Agrobot. Cluj-Napoca 2010, 38, 49–52. [Google Scholar]
- Ramamurthy, V.; Venugopalan, M.; Parhad, V.; Prasad, J. Effect of seed priming on emergence and yield of late sown wheat (Triticum aestivum L.) on Typic Haplusterts of Central India. Indian J. Agric. Res. 2015, 49, 245. [Google Scholar] [CrossRef]
- Hanaoka, K.; Sun, D.; Lawrence, R.; Kamitani, Y.; Fernandes, G. The mechanism of the enhanced antioxidant effects against superoxide anion radicals of reduced water produced by electrolysis. Biophys. Chem. 2004, 107, 71–82. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Emergence | FSC | FSP.CW | FSP.CM | FSP.DW | ASC | ASP.CW | ASP.CM | ASP.DW | LSD |
|---|---|---|---|---|---|---|---|---|---|
| Pisum sativum | |||||||||
| First day of emergence | 4.3 ± 0.3 ab | 4.0 ± 0 a | 4.3 ± 0.2 ab | 4.5 ± 0.3 abc | 6.3 ± 0.2 d | 4.5 ± 0.3 abc | 5.3 ± 0.2 bcd | 5.5 ± 0.3 abc | 0.7 ± 0.4 |
| Last day of emergence | 6.3 ± 0.3 bc | 4.5 ± 0.3 a | 5.8 ± 0.2 b | 6.3 ± 0.2 bc | 7.5 ± 0.3 d | 6.3 ± 0.2 bc | 7.5 ± 0.3 d | 7.3 ± 0.2 cd | 0.8 ± 0.4 |
| Emergence % | 100.0 ± 0 c | 100.0 ± 0 c | 100.0 ± 0 c | 100.0 ± 0 c | 60.0 ± 0 a | 80 ± 8.2 b | 75 ± 5.0 ab | 70 ± 5.8 ab | 11.5 ± 5.6 |
| Mean emergence time | 4.1 ± 0.2 cd | 4.9 ± 0.04 d | 4.3 ± 0.2 cd | 3.9 ± 0.2 cd | 1.7 ± 0.2 a | 3.3 ± 0.4 bc | 2.6 ± 0.2 ab | 2.1 ± 0.2 a | 0.7 ± 0.3 |
| Emergence index | 3.0 ± 0.3 d | 4.3 ± 0.1 e | 3.2 ± 0.3 de | 2.8 ± 0.3 cd | 1.0 ± 0.2 a | 2.4 ± 0.3 bcd | 1.7 ± 0.2 abc | 1.3 ± 0.2 ab | 0.7 ± 0.3 |
| Uniformity of emergence | 0.14 ± 0.03 bc | 0.31 ± 0.01 d | 0.18 ± 0.04 c | 0.12 ± 0.02 abc | 0.03 ± 0.01 a | 0.09 ± 0.02 abc | 0.05 ± 0.01 ab | 0.04 ± 0.01 a | 0.06 ± 0.03 |
| Cucurbita pepo | |||||||||
| First day of emergence | 6.5 ± 0.5 b | 4.0 ± 0 a | 6.0 ± 0 ab | 6.0 ± 0 ab | 10.0 ± 0.8 c | 6.0 ± 0 ab | 6.5 ± 0.5 b | 7.0 ± 0.6 b | 1.2 ± 0.6 |
| Last day of emergence | 9.5 ± 0.5 b | 6.5 ± 0.50 a | 10.0 ± 0 b | 10.0 ± 0 b | 14.5 ± 0.5 d | 10.5 ± 0.5 bc | 11.0 ± 0.6 bc | 12.5 ± 0.5 cd | 1.3 ± 0.6 |
| Emergence % | 100.0 ± 0 c | 100.0 ± 0 c | 100.0 ± 0 c | 100.0 ± 0 c | 60.0 ± 0.0 a | 80.0 ± 8.2 b | 70.0 ± 5.8 ab | 70.0 ± 5.8 ab | 11.9 ± 5.8 |
| Mean emergence time | 4.3 ± 0.10 d | 4.8 ± 0.03 d | 4.3 ± 0.04 d | 4.12 ± 0.04 cd | 1.6 ± 0.10 a | 3.4 ± 0.30 bc | 2.8 ± 0.21 b | 2.6 ± 0.21 b | 0.5 ± 0.20 |
| Emergence index | 2.4 ± 0.10 d | 3.6 ± 0.09 e | 2.4 ± 0.06 d | 2.2 ± 0.08 d | 0.6 ± 0.10 a | 1.9 ± 0.20 cd | 1.4 ± 0.12 bc | 1.3 ± 0.16 b | 0.35 ± 0.17 |
| Uniformity of emergence | 0.015 ± 0.001 de | 0.021 ± 0.0004 f | 0.016 ± 0.0004 e | 0.014 ± 0.001 de | 0.006 ± 0.001 a | 0.013 ± 0.001 cd | 0.010 ± 0.001 bc | 0.009 ± 0.001 cd | 0.002 ± 0.001 |
| Treatment | Pisum sativum | Cucurbita pepo | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Root Length (cm) | Stem Length (cm) | Number of Leaves | Number of Inflorescence | Leaves Area (cm2) | Root Length (cm) | Stem Length (cm) | Number of Leaves | Number of Inflorescence | Leaves Area (cm2) | |
| FSC | 39.8 ± 0.7 c | 36.2 ± 0.5 e | 79.5 ± 1.0 c | 5.5 ± 0.3 cd | 266.8 ± 1.3 d | 30.5 ± 1.7 abc | 28.6 ± 3.0 de | 9 ± 0.4 bcd | 9.3 ± 0.6 c | 681.2 ± 8.3 c |
| FSP.CW | 45.5 ± 1.0 d | 45.5 ± 0.6 f | 97 ± 3.1 d | 9.5 ± 0.6 e | 309.4 ± 3.3 d | 51.6 ± 4.9 d | 42.8 ± 2.6 f | 11.3 ± 0.7 d | 10.8 ± 1.0 c | 768.4 ± 12.2 d |
| FSP.CM | 44.8 ± 0.9 d | 37.0 ± 0.9 e | 87.3 ± 1.3 c | 6.8 ± 0.5 d | 271.6 ± 12.8 d | 38.5 ± 2.4 bc | 21.4 ± 1.3 bcd | 9.5 ± 1.2 bcd | 9.5 ± 0.3 c | 680.9 ± 10.1 c |
| FSP.DW | 40.8 ± 1.6 cd | 36.6 ± 1.0 e | 85.3 ± 2.3 c | 6.5 ± 0.6 d | 269.4 ± 15.4 d | 38.8 ± 1.1 bc | 32.3 ± 1.1 e | 10.5 ± 0.3 cd | 6.5 ± 0.3 b | 701.2 ± 4.9 cd |
| ASC | 15.7 ± 0.8 a | 18.8 ± 0.4 a | 36.2 ± 1.2 a | 2.3 ± 0.2 a | 67.5 ± 1.1 a | 26.0 ± 1.6 a | 11.0 ± 0 a | 5.5 ± 0.2 a | 0.0 ± 0 a | 350.1 ± 4.3 a |
| ASP.CW | 37.3 ± 1.3 c | 31.1 ± 0.4 d | 52.5 ± 0.3 b | 4.5 ± 0.3 bc | 205.6 ± 3.8 c | 40.8 ± 0.7 cd | 27.1 ± 1.1 cde | 8.5 ± 0.3 bc | 6.5 ± 0.3 b | 645.9 ± 9.5 c |
| ASP.CM | 26.8 ± 0.7 b | 27.7 ± 0.6 c | 38.7 ± 1.6 a | 3.3 ± 0.2 ab | 143.3 ± 6.1 b | 35.8 ± 1.6 abc | 20.1 ± 2.2 bc | 7.5 ± 0.3 ab | 2.5 ± 0.3 a | 635.6 ± 11.1 c |
| ASP.DW | 23.0 ± 0.7 b | 22.8 ± 0.7 b | 51.4 ± 1.1 b | 2.8 ± 0.2 ab | 132.4 ± 1.7 b | 27.8 ± 2.14 ab | 13.4 ± 1.07 ab | 7.0 ± 0.41 ab | 2.5 ± 0.7 a | 457.4 ± 9.5 b |
| LSD0.05 | 3.1 ± 1.5 | 2.0 ± 1.0 | 4.9 ± 2.3 | 1.2 ± 0.6 | 37.6 ± 2.2 | 6.7 ± 3.3 | 5.3 ± 2.5 | 1.7 ± 0.8 | 1.6 ± 0.8 | 48.2 ± 23.4 |
| Dry Mass (g plant−1) | FSC | FSP.CW | FSP.CM | FSP.DW | ASC | ASP.CW | ASP.CM | ASP.DW | LSD0.05 |
|---|---|---|---|---|---|---|---|---|---|
| Pisum sativum | |||||||||
| Root Mass | 1.2 ± 0.01 bc | 1.6 ± 0.16 c | 1.6 ± 0.2 c | 1.2 ± 0.14 bc | 0.1 ± 0.01 a | 0.9 ± 0.07 bc | 0.6 ± 0.27 ab | 0.8 ± 0.19 ab | 0.5 ± 0.24 |
| Stem Mass | 1.0 ± 0.08 c | 1.5 ± 0.09 d | 1.1 ± 0.1 c | 1.1 ± 0.02 c | 0.3 ± 0.03 a | 0.7 ± 0.09 b | 0.5 ± 0.02 ab | 0.7 ± 0.03 b | 2.0 ± 0.09 |
| Leaf Mass | 1.1 ± 0.05 cd | 1.2 ± 0.09 d | 0.9 ± 0.12 c | 1.0 ± 0.04 cd | 0.2 ± 0.02 a | 0.6 ± 0.03 b | 0.5 ± 0.03 b | 0.5 ± 0.01 b | 0.2 ± 0.08 |
| Shoot Mass | 2.9 ± 0.11 d | 5.3 ± 0.08 e | 2.8 ± 0.15 d | 2.9 ± 0.04 d | 0.7 ± 0.03 a | 2.1 ± 0.12 c | 1.4 ± 0.03 b | 1.9 ± 0.03 c | 0.3 ± 0.12 |
| Flower Mass | 0.8 ± 0.03 b | 2.6 ± 0.11 c | 0.9 ± 0.06 b | 0.8 ± 0.01 b | 0.2 ± 0.01 a | 0.8 ± 0.05 b | 0.4 ± 0.01 a | 0.7 ± 0.01 b | 0.1 ± 0.07 |
| Total Biomass | 4.1 ± 0.09 c | 6.9 ± 0.16 d | 4.4 ± 0.34 c | 4.0 ± 0.16 c | 0.8 ± 0.04 a | 2.9 ± 0.10 b | 2.1 ± 0.30 b | 2.7 ± 0.23 b | 0.6 ± 0.29 |
| Shoot/Root Ratio | 2.4 ± 0.10 a | 3.4 ± 0.34 a | 1.9 ± 0.37 a | 2.6 ± 0.29 a | 14.4 ± 0.66 b | 2.4 ± 0.28 a | 5.1 ± 2.39 a | 2.9 ± 0.66 a | 2.7 ± 1.32 |
| Cucurbita pepo | |||||||||
| Root Mass | 1.0 ± 0.1 c | 1.8 ± 0.14 d | 1.0 ± 0.09 c | 1.1 ± 0.14 c | 0.3 ± 0.002 a | 0.8 ± 0.05 bc | 0.5 ± 0.02 ab | 0.4 ± 0.08 a | 0.26 ± 0.13 |
| Stem Mass | 1.6 ± 0.1 de | 2.2 ± 0.18 f | 2.0 ± 0.17 def | 2.1 ± 0.09 ef | 0.3 ± 0.02 a | 1.5 ± 0.03 cd | 1.0 ± 0.11 bc | 0.6 ± 0.09 ab | 0.32 ± 0.16 |
| Leaf Mass | 1.8 ± 0.2 bc | 2.1 ± 0.29 c | 2.0 ± 0.15 bc | 2.1 ± 0.15 c | 0.9 ± 0.01 a | 2.0 ± 0.17 bc | 1.4 ± 0.1 abc | 1.3 ± 0.16 abc | 0.51 ± 0.24 |
| Shoot Mass | 3.7 ± 0.3 cd | 4.8 ± 0.45 d | 4.1 ± 0.23 cd | 4.4 ± 0.10 cd | 1.2 ± 0.01 a | 3.6 ± 0.18 c | 2.5 ± 0.19 b | 2.0 ± 0.21 ab | 0.70 ± 0.34 |
| Flower Mass | 0.3 ± 0.02 de | 0.4 ± 0.03 e | 0.2 ± 0.03 c | 0.2 ± 0.01 cd | 0.0 ± 0 a | 0.2 ± 0.01 bc | 0.1 ± 0.01 ab | 0.08 ± 0.03 a | 0.07 ± 0.03 |
| Total Biomass | 4.7 ± 0.2 c | 6.5 ± 0.53 d | 5.1 ± 0.17 c | 5.5 ± 0.14 cd | 1.5 ± 0.01 a | 4.5 ± 0.22 c | 3.0 ± 0.21 b | 2.3 ± 0.28 ab | 0.80 ± 0.38 |
| Shoot/Root Ratio | 4.0 ± 0.5 ab | 2.70 ± 0.26 a | 4.4 ± 0.66 ab | 4.4 ± 0.6 ab | 4.2 ± 0.07 ab | 4.4 ± 0.17 ab | 5.0 ± 0.27 b | 5.7 ± 0.78 b | 1.40 ± 0.68 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fatokun, K.; Beckett, R.P.; Varghese, B.; Cloete, J.; Pammenter, N.W. Influence of Cathodic Water Invigoration on the Emergence and Subsequent Growth of Controlled Deteriorated Pea and Pumpkin Seeds. Plants 2020, 9, 955. https://doi.org/10.3390/plants9080955
Fatokun K, Beckett RP, Varghese B, Cloete J, Pammenter NW. Influence of Cathodic Water Invigoration on the Emergence and Subsequent Growth of Controlled Deteriorated Pea and Pumpkin Seeds. Plants. 2020; 9(8):955. https://doi.org/10.3390/plants9080955
Chicago/Turabian StyleFatokun, Kayode, Richard P. Beckett, Boby Varghese, Jacques Cloete, and Norman W. Pammenter. 2020. "Influence of Cathodic Water Invigoration on the Emergence and Subsequent Growth of Controlled Deteriorated Pea and Pumpkin Seeds" Plants 9, no. 8: 955. https://doi.org/10.3390/plants9080955