
Abstract
Context
Intact tropical rainforests are considered robust to plant invasions. However, land-use change alters the structure and species composition of native forest, opening up tropical landscapes to invasion. Yet, the relative roles of key drivers on tropical forest invasions remain little investigated.
Objectives
We examine factors affecting plant invasion of rainforest remnants in oil-palm dominated landscapes in Sabah, Malaysian Borneo. We hypothesized that invasion is greater in highly fragmented landscapes, and in disturbed forests with lower native plant diversity (cf. old-growth rainforests).
Methods
Native and exotic plants were surveyed in 47 plots at 17 forest sites, spanning gradients in landscape-scale fragmentation and local forest disturbance. Using partial least squares path-modelling, we examined correlations between invasion, fragmentation, forest disturbance, propagule pressure, soil characteristics and native plant community.
Results
We recorded 6999 individuals from 329 genera in total, including eight exotic species (0–51% of individuals/plot, median = 1.4%) representing shrubs, forbs, graminoids and climbers. The best model (R2 = 0.343) revealed that invasion was correlated with disturbance and propagule pressure (high prevalence of exotic species in plantation matrix), the latter being driven by greater fragmentation of the landscape. Our models revealed a significant negative correlation between invasion and native tree seedlings and sapling community diversity.
Conclusions
Increasing landscape fragmentation promotes exotic plant invasion in remnant tropical forests, especially if local disturbance is high. The association between exotic species invasion and young native tree community may have impacts for regeneration given that fragmentation is predicted to increase and so plant invasion may become more prevalent.
Similar content being viewed by others

Introduction
Exotic species invasion is recognised as a major agent of environmental change across all ecosystems (Sala et al. 2005). Invasion is considered to be a function of four factors: propagule pressure, the abiotic characteristics of the invaded system, and the biotic characteristics of both the invaded system and invading species, all of which can be altered by human activity (Pyšek and Richardson 2006). In a given invasion scenario these factors contribute to varying degrees, and it is crucial to identify the relative role of each in driving invasion in a particular ecosystem (Catford et al. 2009). Many temperate forests are highly invaded by exotic plants due to historic land-use change causing substantial forest cover loss and resulting in highly fragmented and disturbed landscapes (McNeely 1995; Williams 2003; Haddad et al. 2015). Conversely, intact, lowland, humid tropical forests are usually considered to be robust to plant invasions due to low propagule pressure, low forest disturbance and because exotic species typically do not thrive in the strongly shaded conditions within undisturbed forests (Fine 2002; Denslow and DeWalt 2008). In addition, it has been hypothesised that the typically high native diversity in tropical rainforests means there are fewer empty niches, providing biotic resistance to invading exotic plants (Fine 2002; Denslow and DeWalt 2008). However, land-use change is impacting large tracts of forest across the tropics (Pütz et al. 2014; Haddad et al. 2015; Qie et al. 2017), opening up these landscapes to exotic propagules and further disturbance in the remaining patches of forest (Hulme 2009). Forest degradation alters the availability of light and other resources, changing the nature of inter-specific competition (Denslow and DeWalt 2008; Carson and Schnitzer 2011), and potentially exposes these forest patches to invasion of exotic plants. Yet, despite this, little is known about plant invasions within human-modified tropical landscapes (Kueffer et al. 2013), including remnant forests which are crucial for maintaining local and regional native biodiversity.
Large-scale deforestation in the tropics is predominantly driven by expansion of agriculture, and these agricultural areas have high levels of disturbance and abiotic conditions which differ from those of intact tropical forests (Malhi et al. 2014; Curtis et al. 2018). Abiotic conditions in tropical agricultural areas (e.g. high light levels, high temperature, low humidity, altered soil nutrients and pH) promote establishment of ruderal exotic plant species that are transported and introduced by humans via extensive networks of roads (With 2002; Von der Lippe and Kowarik 2007; Padmanaba and Sheil 2014; Fee et al. 2017), resulting in exotic-rich weed-covered banks along disturbed roadsides (Fee et al. 2017). These highly disturbed agricultural areas provide an on-going source of propagules and may promote invasion of natural forest remnants embedded within these landscapes. From the few studies of plant invasions within humid tropical forests, the presence of exotic species correlates with variables that are associated with propagule pressure, including distance from source population and to forest edge (Edward et al. 2009; Dawson et al. 2014). If propagule pressure is high enough, seed swamping of new environments can result in successful exotic establishment, regardless of whether the conditions of the invaded system are ideal for that species (see Propagule pressure hypothesis; Lockwood et al. 2005; Colautti et al. 2006). Nonetheless, abiotic and biotic factors also contribute to the invasion process.
Disturbance has been foundational in the formulation of several invasion hypotheses (e.g. Disturbance, Habitat filtering, Empty niche and Increased Resource Availiabilty hypotheses; see Catford et al. 2009 for invasion hypothesis overview), therefore invasion is expected to be higher in the most disturbed tropical rainforests. Commercial selective logging and the impact of fragmentation (e.g. via edge effects) alter the structure of remaining rainforest due to timber removal and disturbance-related tree mortality (Laurance et al. 2002; Malhi et al. 2014; Gibbs and Salmon 2015). The reduction in canopy cover increases light in the forest understorey, which affects the abiotic conditions of remnant forest patches, when compared with continuous, undisturbed forest (Laurance et al. 2002). Gradients from forest edge to interior have been recorded for temperature, humidity, wind intensity and soil erosion, with the most extreme differentiation seen at forest edges, known as edge effects (Laurance et al. 2002). In disturbed tropical forests, more exotic plants have been recorded at forest edges, driven by a combination of increased propagule pressure and increased canopy openness and soil disturbance associated with forest edge conditions and anthropogenic influence (Peters 2001; Harper et al. 2005; Edward et al. 2009; Dawson et al. 2014; Döbert et al. 2017). However, these studies only looked at a few forest sites, only one studied a gradient of disturbance (Döbert et al. 2017) and none have considered gradients in landscape fragmentation.
In addition to forest structural changes caused by logging and fragmentation, changes to species community composition also occur due to disruption of native species interactions (Laurance et al. 2011; Arellano-Rivas et al. 2016). For example, removal of large timber trees and isolation from larger tracts of forest reduce the regeneration of native old-growth tree species through reduced fruiting and dispersal events, shifting tree species composition to earlier successional states with lower community-average wood density (Laurance et al. 2006; Lôbo et al. 2011; Pütz et al. 2014; Gibbs and Salmon 2015; Qie et al. 2017). The negative impacts of fragmentation on native species communities are greater in small, isolated fragments and accumulate over time (Gonzalez 2000; Ferraz et al. 2003; Hill and Curran 2003; Haddad et al. 2015), with the understorey tree community strongly affected due to adult trees producing fewer viable seeds (i.e. extinction debt; Stride et al. 2018). Therefore, forest remnants may be particularly vulnerable to invasions due to reduced biotic resistance arising from the loss of native forest species. However, evidence for biotic resistance to exotic plant invasion in tropical rainforests is limited (Fine 2002; Levine et al. 2004; Denslow and DeWalt 2008), with no consensus currently (Teo et al. 2003; Brown et al. 2006). Thus it is still unclear whether biotic interactions between native and exotic species (i.e. biotic resistance from the native community as well as suppression of native growth by invading exotic species) play a significant role in mediating invasion of tropical forests.
South-East Asia has the highest rates of deforestation (Achard et al. 2002; Sodhi et al. 2004) and logging (primarily trees from Dipterocarpaceae; Hansen et al. 2008; Lewis et al. 2015; Brearley et al. 2016) in the world (Bryan et al. 2013; Gaveau et al. 2017), due in particular to the expansion of oil palm (Elaeis guineensis Jacq.) cultivation in Malaysia and Indonesia in recent decades. This expansion of agriculture results in a highly fragmented forest landscape made up of commercial oil palm plantations with uncultivated, often heavily logged forest remnants (Reynolds et al. 2011). These remnant forests are therefore usually highly degraded and contain reduced assemblages of both plants and animals, compared with old-growth continuous forest, particularly if very little core forest area remains within the remnants (Lucey et al. 2017). However, these forest remnants can be important for conserving biodiversity and ecosystem services and for connecting forest patches within oil palm dominated regions, even if they are highly degraded (Yeong et al. 2016; Lucey et al. 2017; Scriven et al. 2017; Fleiss et al. 2020).
In this study, we examine the relative influence of fragmentation, forest disturbance, propagule pressure, soil characteristics and native community composition on exotic plant invasions of forest remnants within oil palm-dominated landscapes in Sabah, Malaysian Borneo. Studies examining invasion of tropical forests are rare, and this study is the first to examine a range of environmental and ecological factors leading to invasion of forest remnants embedded within an oil palm landscape. We use partial least squares path modelling (PLS-PM) to test the expectation that invasion is positively correlated with landscape-scale fragmentation, local forest disturbance and propagule pressure, and negatively correlated with native community diversity, whilst accounting for soil characteristics. We also expect the native community to respond negatively to land-use change.
Material and methods
Study sites
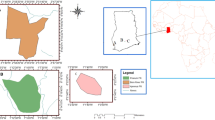
Between July and October 2017, we sampled 47 plots within 17 sites distributed widely across the state of Sabah, Malaysian Borneo (Fig. 1a). Sites were selected to span a wide gradient of fragmentation and disturbance, including isolated, heavily logged, small forest remnants (< 15 ha) that were fully surrounded by oil palm, as well as larger, heavily logged forest remnants (> 100 ha) that were connected at the plantation boundary to larger tracts of government protected forest. Forest sites in oil-palm dominated landscapes (n = 13 sites), which have between 1 and 64% natural forest cover remaining within a 2 km radius buffer surrounding study plots (see section below; characterising fragmentation), were compared with four sites within an extensive tract of intact ‘continuous’ logged and unlogged protected forest (> 1 million ha; Fig. 1a). These continuous forest sites provided baseline data for comparison of forest remnants within an oil palm landscape and also represented variation in levels of forest disturbance in the region, including both fully protected unlogged forest (n = 2) and commercially logged sites (n = 1 site in once-logged forest; n = 1 site in twice-logged forest). Thus, our choice of sites spanned a gradient in the degree to which the landscape surrounding our sites was fragmented as well as the intensity to which the forest areas were previously commercially selectively logged.

a Map of study locations, forest cover and industrial oil palm plantations within Sabah, on the island of Borneo (inset). b Schematic diagram of experimental plot design within fragmented and continuous forest sites. Distance of plots from forest edge and between plots are shown along with the radiuses of each plot and subplot, and location of quadrats. Measurements are as follows: C1 = trees ≥ 25 cm DBH; C2 = trees 10–25 cm DBH; C3 = tree saplings 2–10 cm DBH and Q = tree seedlings (< 2 cm DBH) and other ground vegetation (including shrubs, forbs and grasses). See text for full details on floristics surveys. CIFOR forest cover dataset available at Gaveau, Salim and Arjasakusuma (2017)
Sites within oil palm plantations were conservation set-asides, and the majority were managed as sites with ‘High Conservation Values’ (HCVs, Senior et al. 2015), which the plantations retain and manage as part of the Roundtable on Sustainable Palm Oil (RSPO) certification process for sustainable palm oil (RSPO 2018). These sites were selectively logged to varying degrees in the 1990s prior to oil palm cultivation (see details below on disturbance quantification) and were generally unsuitable for planting oil palm (i.e. too steep or too rocky). Study sites were at least 1 km apart to avoid spatial autocorrelation. The maximum distance between sites is c. 192 km with elevation range from 42–267 m above sea level, hence we do not expect the climate to vary greatly between sites.
Plot-level surveys of native and exotic plants
We surveyed plants in two or three circular plots (30 m radius; 0.28 ha) per site, depending on forest extent at each site, with only two in the smallest remnants (< 12.5 ha; n sites = 4). We used a four-level nested approach to sample plants within different size classes (Fig. 1b). The diameter-at-breast height (DBH) was recorded for all large trees ≥ 25 cm DBH within the 0.28 ha plot (Fig. 1b; C1), all small trees 10–25 cm DBH within a sub-plot of 20 m radius (Fig. 1b; C2), and all tree saplings 2–10 cm DBH within a sub-plot of 5 m radius (Fig. 1b; C3). Tree seedlings and other ground vegetation (forbs, shrubs, climbers, ferns and graminoids, from herein referred to simply as ground vegetation) were surveyed in eight 1 × 1 m quadrats (Fig. 1b; Q) located 25 m from the plot centre along eight randomly chosen bearings. Within each of these quadrats, we recorded all plants (tree seedlings, shrubs, climbers, forbs, graminoids and ferns) rooted inside the quadrat, and ≤ 2 cm DBH. We considered an individual as a plant with a distinct root system; for adult trees, tree saplings and tree seedlings individuals were straightforward to distinguish. All ground vegetation, including clonal species (e.g. graminoids and climbers) that can appear as distinct individuals above ground yet connected below ground, was manually removed from the soil to determine number of individuals for each morpho-genus, with unknown individuals taken as vouchers for identification. Adult trees and saplings were identified to genus in the field by an experienced local botanist (co-author AJ). Voucher specimens and photographs of plants in the quadrats were used to identify individuals by a botanist at Danum Valley Field Centre (co-author BBO) and the herbarium at the Forest Research Centre, Sepilok. Exotic species were identified based on lists and guides of Sabah’s known exotic plants prepared prior to fieldwork (see Pallewatta et al. 2003; Bakar, 2004; Peh, 2010; Döbert et al. 2017; Fee et al. 2017; CABI 2017) and confirmed by botanists from vouchers. The observed number of exotic species and number of exotic stems per plot were used as measurements of invasion within a plot. All analyses were carried out at plot-level, and at the level of genus, due to the challenges of reliably identifying native plants to species level in diverse tropical forests. Exotic plants were identified to the species level.
Quantifying the native community
We computed two measures of native community alpha diversity; observed genus richness and Faith’s phylogenetic diversity (PD; Faith 1992). Faith’s PD is the total sum of phylogenetic branch lengths between species—we chose this measure because it is intended to capture the evolutionary signal in functional traits and as such is associated with functional diversity and can be a valuable predictor of ecosystem-level responses (Flynn et al. 2011). These diversity metrics were calculated per plot (n = 47) for the total dataset and separately for trees, saplings, seedlings and the ground vegetation. Individuals that could not be identified to genus (n = 17; < 1% of all stems) were removed from the dataset prior to phylogenetic analyses. A phylogeny was created using Phylomatic v.3 (Webb and Donoghue 2005; available at https://phylodiversity.net/phylomatic/ [accessed 4 May 2018]) and a pruned version of the Angiosperm Phylogeny Groups APGIII maximally resolved supertree of angiosperms (R20120829). Two genera could not be included in the phylogeny, as they were not part of the original supertree, leaving 303 genera for inclusion. The phylogeny was rooted and branch lengths were based on node ages (most recent common ancestor) of Wikstrom et al. (2001) and estimated using the ‘BladJ’ algorithm in Phylocom (Webb et al. 2008). From the resulting phylogeny, phylogenetic diversity (PD; the sum of the total branch lengths measured in millions of years; Faith 1992) was calculated using the ‘picante’ package (Kembel et al. 2010) in R Studio 2.14 (R Core Team 2019).
Characterising fragmentation, disturbance, propagule pressure and soil
We measured forest fragmentation in the surrounding landscape by quantifying the area of forest and non-forest habitat (m2) within a 2 km radius buffer surrounding each plot. This buffer is a proxy for the dispersal potential of native forest species (i.e. more forest in the buffer would likely indicate a higher presence of mother trees as well as pollinators and animal dispersers). This buffer included forested areas not connected to forest remnants where plots were carried out, as well as connected forest within which the plots were embedded. Quantification of buffer habitat was carried out using drone images provided by plantation managers taken during May–November 2016, after which we assume no or negligible change to forested area within plantations had occurred before fieldwork took place (July–October 2017), due to protections in place. Plots in ‘continuous’ sites were almost entirely forest, but the tract of continuous forest nonetheless contains some small settlements (i.e. field research centre) and so there were small areas of non-forest (< 0.5%) in buffer zones around these plots. Within the 2 km buffer, the amount of forest edge in the landscape was also calculated using the ‘rgeos’ and ‘raster’ packages (Hijmans and van Etten 2012; Bivand and Rundel 2013), and was equal to the number of raster grid cells (5 × 5 m) the forest edge passed through. A unitless index of edge density was calculated for each plot as the area of edge cells divided by the area of forest cells. This edge density index was a proxy for the amount of edge habitat in the landscape that would likely be highly degraded based on edge effects. The time since fragmentation was calculated from the year of first planting of oil palm (between 1991 and 2009) as provided by estate managers.
At all sites, we derived two proxy variables for local forest community structure and recovery since disturbance (primarily the result of previous commercial selective logging, except at the two unlogged ‘continuous’ sites). These two variables were: (i) the number of large stems (> 25 cm DBH) belonging to the tree family Dipterocarpaceae, which comprises ~ 22% of tree species and 61% of biomass in the region (Slik et al. 2003; Saner et al. 2012), and provides a useful comparison between disturbed remnants and unlogged ‘continuous’ forest; and (ii) average plot-level wood density for trees > 10 cm DBH was calculated, because wood density can be an indicator of successional status in forest species (Slik et al. 2008; Feeley et al. 2011; Qie et al. 2017). Wood density was calculated for each genus using the ‘BIOMASS’ package in R (Réjou-Méchain et al. 2017), based on the Global Wood Density Database (Chave et al. 2009; Zanne et al. 2009).
As a direct proxy of propagule pressure, exotic plant species richness and abundance were measured along two 100 m transects located within the oil palm matrix at each site. The first transect was along the roadside of a major unpaved plantation road (lorries and car traffic) and the second at the oil palm-forest edge, either along an unpaved minor road (mostly motorbike traffic) or oil palm terrace track (pedestrians). Along both 100 m transects, the presence/absence of 18 exotic species were recorded in each 1 m section and 1 m either side of the transect line (i.e. 2 m2 sections). Exotic species richness was computed as the total number of exotic species (out of 18) recorded across these two transects. Exotic species abundance was the total number of 1 m transect sections (n = 100 per transect) that species were present in, and total abundance of all exotics was computed across the two transects (i.e. out of 18 species × 200 sections = 3600).
To measure soil available phosphorus and pH the topsoil was sampled in each plot (20 cm core; n = 5 per plot: one at the plot centre and four 15 m from centre in each of the cardinal directions). After drying (at 50 °C) soil samples were thoroughly mixed and sub-sampled for analyses at plot level (n = 47). Soil chemistry was analysed at the Forest Research Centre, Sepilok, Sabah. Available phosphorus was extracted using water and measured using the molybdenum-blue method and read at 880 nm on a spectrophotometer (following Anderson and Ingram 1994). Soil pH was measured with a combination glass-calomel electrode in a 1:2.5 ratio of soil to deionised water.
Data analysis
We analysed the relative importance of different factors in invasion using partial least squares path modelling (PLS-PM; see Table 1 for a summary of variables and Supporting Information Appendix S1 for full details on PLS-PM method and variable selection) using the ‘plspm’ R package (Sanchez and Trinchera 2012; see Waddell and Chapman 2020 for the R code). PLS-PM is a variance-based structural equation modelling method, which fits multiple regressions using measured and latent variables connected in a pathway reflecting ecological theory (Sanchez 2013). Latent variables are those which cannot be observed or directly measured but are of conceptual interest and are indicated by their associated measured variables, which comprise at least two highly correlated measured variables. Some measured variables were transformed (Table 1) to improve normality, in accordance with modelling assumptions (SI Appendix S1). All measured variables were centred and scaled (mean of 0 and variance of 1) prior to input into the PLS-PMs to assist with model convergence and interpretation.
All models were developed starting from the full specification in Fig. 2 and removing non-significant terms in a stepwise manner until all retained links were statistically significant (P < 0.05; Sanchez, 2013). Two-sided P-values for standardised path coefficients (i.e. the extent of standard deviation change of one latent variable attributed to one standard deviation change to another latent variable) were estimated by 10,000 bootstrap estimations. In ‘plspm’, site identity cannot be included as a random effect, so site effects were accommodated by taking bootstrap samples at the level of site, i.e. bootstrap samples were made by randomly selecting sites and all their constituent plots, with replacement.

Hypothetical partial least squares path model showing the relationships between fragmentation, disturbance, propagule pressure, soil characteristics and native plant diversity on invasion within of lowland tropical forests. Latent variables are shown in ovals and the measured variables in rectangles. Measurement model is represented by the dashed lines and the structural model is represented by the solid lines. Models were tested in both directions for the relationship between native community and invasion indicated by the double arrows. Full details on these variables are in Table 1 and within the text
We first fitted the model on the full dataset, i.e. where native community diversity comprised recorded native plants of all size classes. We then fitted separate PLS-PMs for native adult trees, tree saplings, tree seedlings and ground vegetation to test whether different subsets of the native community were more strongly correlated with invasion. As we do not know the directionality of the relationship between native community diversity and invasion, we tested all models in both directions for this relationship. In all cases, models were assessed for validity and reliability in both the measurement and structural model (see SI Appendix S1 for details on PLS-PM model evaluation).
Results
In the 47 plots we surveyed a total of 6999 individuals (SI Table S2) from 329 genera, including eight exotic species, ranging from 0 to 7 species (median = 1) and 0–51% of stems (median = 1.4%) per plot. Exotic species included shrubs, forbs, graminoids and climbers (there were no exotic trees). The most abundant exotic was a common tropical exotic shrub, Clidemia hirta (Melastomataceae), which made up 73.8% of all exotic stems (SI Table S3). The second most common exotic was also a shrub, Chromolaena odorata (11% of exotic stems), and the other six species were either forbs, climbers or a graminoid (1–4% of exotic stems).
Overall, native richness ranged from 18–69 genera/plot (median 45) with plots in the 13 oil palm remnant sites having lower richness (median of 42 genera/plot; range 18–69) compared with continuous sites (median of 46.5 genera/plot; range 39–57). In continuous sites the most common native genus amongst adult trees was Shorea (Dipterocarpaceae), for saplings Mallotus (Euphorbiaceae), for seedlings Shorea (Dipterocarpaceae) and for ground vegetation Selaginella (Selaginellaceae). By contrast in the oil palm remnant sites, the most common native genera were Ficus (Moraceae), Hopea (Dipterocarpaceae), Glochidion (Phyllanthaceae) and Selaginella (Selaginellaceae) for the same size classes, respectively.
The final PLS-PMs (Figs. 3, 4, S1 and S2) explained 34% of the variation in invasion, 74% of variation in propagule pressure, between 26 and 53% of native community diversity and 55% of soil characteristics (all of which are latent variables informed by 2–3 measured variables). There was strong evidence in support of our final PLS-PMs in terms of the reliability and validity of the measurement model and an adequate fit of the structural model (see Appendix S1 and Tables S4–6 for full details on model evaluation).

Fitted partial least squares path model showing the relationships between fragmentation, disturbance, propagule pressure, soil characteristics and native plant diversity on invasion within of lowland tropical forests. The model was simplified from the specification in Fig. 2 by removing non-significant effects. The goodness-of-fit index for the model and R2 values for the endogenous latent variables are displayed. Standardised path coefficients and P-values were estimated by 10,000 bootstrap estimations. Standardised path coefficients are shown next to arrows, red indicates a negative correlation and blue a positive correlation. They denote the extent of standard deviation change of one latent variable attributed to one standard deviation change to another latent variable. P-values: *P < 0.05; **P < 0.01 and ***P < 0.001

Partial least squares path model showing the relationships between fragmentation, disturbance, propagule pressure, soil characteristics and invasion on a native tree sapling and b native seedling diversity. The model was simplified from the specification in Fig. 2 by removing non-significant effects. The goodness-of-fit index (GoF) for each model and R2 values for the endogenous latent variables are displayed. Standardised path coefficients and P-values were estimated by 10,000 bootstrap estimations. Standardised path coefficients are shown next to arrows, red indicates a negative correlation and blue a positive correlation. They denote the extent of standard deviation change of one latent variable attributed to one standard deviation change to another latent variable. P-values based on 10,000 bootstrap estimations: *P < 0.05; **P < 0.01; ***P < 0.001. F fragmentation, D disturbance, P propagule pressure, S soil characteristics, N native community and I invasion
Drivers of invasion
After removal of non-significant terms, the best model with the full native community (i.e. combining data for native adult trees, saplings, seedlings and ground vegetation; Fig. 3), found a strong direct positive correlation between propagule pressure and invasion (P < 0.001), with a strong indirect positive correlation of fragmentation (P < 0.01) on invasion via increased propagule pressure. Forest disturbance also had a direct positive correlation with invasion (P < 0.05), but there was no significant effect of total native community diversity or soil characteristics (P > 0.05) on invasion.
Models accounting separately for native adult trees, saplings, seedlings and ground vegetation communities did not support a role of native community diversity on invasion (native → invasion; Fig. S1 and Table S7). However, when the direction of the biotic relationship in the model was reversed, i.e. invasion → native, we found significant negative effects of invasion on native tree sapling and tree seedling communities (Fig. 4; tree saplings: standardised path coefficients = − 0.348, P = 0.012, R2 = 0.4; tree seedlings: standardised path coefficients = -0.28, P < 0.01, R2 = 0.299). In these models, there was no significant effect of invasion on the total native community, adult trees or ground vegetation (Fig. S2).
Responses of the native community to land-use change
The exotic and native communities (both total and subsets) responded differently to disturbance and fragmentation (Figs. 3, 4, S1 and S2). The total native community was indirectly correlated negatively with both fragmentation and disturbance, via their positive influence on soil characteristics (i.e. higher native diversity and abundance when there was low soil pH and available soil phosphorus; Fig. 3 and Table S7). Instead, invasion was positively correlated with these land-use change variables, as we hypothesised. In the native subsets (Figs. 4, S1 and S2, and Table S7), adult tree and tree sapling communities were directly negatively correlated with disturbance, tree seedling community was directly negatively correlated with fragmentation, and the ground vegetation community (forbs, shrubs, climbers, ferns and graminoids) was indirectly negatively correlated with both fragmentation and disturbance, via interactions with soil characteristics. Hence, all the native community subsets (adult trees, tree saplings, tree seedlings and ground vegetation) were negatively associated with disturbance and fragmentation.
Discussion
Our study found that a higher degree of landscape fragmentation leads to higher invasion of exotic plant species in forest remnants, due to a greater source population of exotic plants found in the anthropogenic (i.e. plantation) areas of these landscapes. In contrast, we found little effect of soil characteristics and the native community on exotic invasion, but a negative influence of exotic invasion on the native tree sapling and seedling communities.
Fragmentation drives propagule pressure to increase invasion
Landscape fragmentation (i.e. lower forest cover, more edge habitat and a longer history of forest clearance) promotes higher invasion of the oil palm matrix (planted areas and roads), which increases the likelihood of remnant forests being invaded by exotics due to increased propagule pressure. We expected that greater numbers of exotic species would have established in the oil palm matrix because of enhanced transport of propagules (Ansong and Pickering 2013) and the higher availability of resources brought about through disturbance, which would promote colonisation by exotic species (Fee et al. 2017). Small, isolated forest remnants that are completely surrounded by oil palm, or other agricultural areas (c.f. those contiguous with larger tracts of forest) may therefore have more exotic propagules arriving from all directions, facilitating the invasion process. Although exotic invasions have not yet been thoroughly examined in fragmented tropical rainforests, similar patterns to those in this study have been reported in many natural habitats within fragmented landscapes, with invasion highest in small, isolated remnants (Vilà and Ibáñez 2011). We conclude that propagule pressure, driven by fragmentation, is the main factor influencing invasion of exotics in our study (based on total effect sizes in Table S7), however, invasion is generally low unless there is high local disturbance of the forest.
Increased invasion in disturbed forests
We found that invasion is more likely in forests with high levels of previous disturbance, represented by those with fewer large dipterocarp trees and lower average wood density, indicative of forest regressing to an early successional state. The strong positive relationship between disturbance and invasion is likely to indicate increased resource availability (e.g. light and space) associated with commercial selective logging, providing opportunities for invading exotics to colonize. The correlation between invasion and increased resource availability following disturbance is found across many habitats and disturbance regimes (natural and anthropogenic), and underpins several invasion hypotheses (Davis et al. 2000; Hood and Naiman 2000; Colautti et al. 2006). In tropical forests, increased light levels following selective logging can also lead to increases in native pioneer species and native weeds (Catford et al. 2009; Lockwood et al. 2013) and these environmental factors are likely to be instrumental in driving the invasion of exotic species observed in our data. Therefore, fragmentation and disturbance may operate synergistically; land-use change may bring exotic species propagules into the oil palm matrix, but these exotic species will only establish in the remnant forests if local conditions are suitable and disturbance promotes these conditions.
Little evidence for biotic interactions
We found no signal of biotic resistance to invasion from the native community in our results when the models specified an effect of the native community on invasion, which is in line with wider meta-analysis of terrestrial plant invasions (Jeschke et al. 2012). However, when the models specified an effect of invasion on the native community, we found significant negative correlations between invasion and native tree sapling and seedling communities. This may indicate either the impact of a dominant exotic inhibiting native recruitment or be driven by indirect effects of land-use change on these cohorts of native trees (i.e. indirect effect of the disturbance latent variable on saplings via invasion and similarly fragmentation on seedlings).
These findings may be explained by young native trees and exotic shrubs (~ 85% of exotic stems) having higher niche overlap than other components of the native community (i.e. adult trees and ground vegetation) because they have a similar woody habit and occupy the same lower understorey environment. Exotic species may have disrupted the recruitment of current tree saplings and seedlings, which would have been young saplings or seeds when exotic species first invaded these forests post-fragmentation (8–26 years before sampling), by outcompeting them for resources. The dominant exotic recorded in this study, Clidemia hirta (74% of exotic individuals), exhibits functional traits associated with high competitive ability (e.g. high seed number, fast growth rate, early age of reproduction, asexual reproduction, fruits produced year round and very large seed bank), allowing for rapid population growth (Rejmánek and Richardson 1996; Singhakumara et al. 2000; Daehler 2003), making it a highly invasive species and a pest throughout most of its introduced range (Wester and Wood 1977; Gerlach 1993). Similar patterns have been observed in temperate forests with dominant invasive species disrupting native seedling growth (Gorchov and Trisel 2003; Stinson et al. 2006), and in selectively logged Malaysian tropical rainforests, Döbert et al. (2017) found fewer Dipterocarpaceae seedlings where exotic biomass was higher. Therefore, high niche overlap may lead to strong competition between young trees and exotic shrubs for resources such as light. In addition, C. hirta and Chromolaena odorata (the second most common exotic invasive species recorded) both have allelopathic effects, which have been shown to significantly reduce native seed germination and growth during laboratory studies (Hu and Zhang 2013; Ismaini 2015).
Our results are correlational and do not track changes in the native community over time, so inferring biotic interactions is problematic. Nevertheless, our results indicate that invasion of exotic plants is more closely associated with the lack of young native trees than other components of the native plant community (e.g. ground vegetation and adult trees) in these forests, and patterns are likely driven by Clidemia hirta. However, in order to determine the mechanisms driving observed biotic relationships in our data, as well as the true directionality of observed relationships, long-term manipulative field experiments would be required.
Native and exotic plants respond differently to disturbance and fragmentation
Our results suggest that different components of human land-use change correlate, at varying strengths, with different subsets of the native community (adult trees, tree saplings, tree seedlings and ground vegetation). The strong direct correlation between disturbance and native adult tree diversity could potentially be driven by the disturbance variables included in the models being derived from the adult tree community (i.e. number of large dipterocarp trees remaining and mean adult tree wood density per plot). However, as this relationship is also found with sapling diversity, a true relationship between habitat disturbance and both tree communities seems plausible. This may be due to logging decreasing diversity by the physical removal of specific species via timber extraction (i.e. dipterocarps), affecting both timber trees (adult trees) and collateral damage to other trees surrounding the extracted tree, both over- and understorey (Malhi et al. 2014). In addition, altered abiotic conditions brought about by logging practices (e.g. increased light, temperature, soil erosion) result in tree mortality and reduced fruiting in adult trees which in turn reduce recruitment (Gibbs and Salmon 2015), decreasing the richness of old-growth forest species in fragments.
The association between fragmentation and tree seedling diversity (and to a lesser extent the marginally significant correlation with tree saplings) in our data means there are fewer seedlings in areas with less forest in the landscape, more edge habitat and in older plantations. This could reflect an extinction debt driven by pollination and dispersal limitations in highly fragmented landscapes (Ghazoul and McLeish 2001), which would have clear implications for the future regeneration of these forests (Haddad et al. 2015). Stride et al (2018) found similar results in other fragmented forest sites in our study region, with seedling richness 30% lower in forest remnants than in undisturbed old-growth forest.
The weak negative correlation between ground vegetation community composition and both fragmentation and disturbance (SI Table S7), may be due to this component of the plant community also containing light-loving native weedy species (e.g. grasses and forest edge shrubs), which act much the same as exotics in that they proliferate with disturbance (Catford et al. 2009; Lockwood et al. 2013). These native weeds could therefore dilute the relationship between native ground vegetation typical of old-growth forest and the effects of land-use change in our models. Changes in the native plant community associated with disturbance, as well as the invasion of exotic species, are likely to impact important ecosystem services in these forests, such as biodiversity, functional stability and carbon storage.
Conclusions and conservation implications
Exotic species are an understudied component of fragmented tropical landscapes, despite being an emerging threat to the natural regeneration of native tree communities within remnant forests. Indeed, three of the eight exotic species we found (Clidemia hirta, Chromolaena odorata and Mikania micrantha) are considered amongst the world’s worst 100 invasive species due to their ability to modify the ecosystems they invade (Lowe et al. 2000), with C. hirta in particular, considered a pest across much of its introduced range (Wester and Wood 1977; Gerlach 1993; Peters 2001; Teo et al. 2003). Our study shows for the first time that fragmentation and local disturbance increase the invasion of exotic species in rainforest remnants within anthropogenic landscapes. Therefore, we suggest that better management to control exotic populations within oil palm plantations (e.g. along roadsides and in oil palm planted areas), along with active regeneration of forest remnants to improve overall forest quality and canopy cover is important to reduce exotic invasions. In a natural resource management context, the way in which invasive species move through a landscape should be considered in the design of multi-functional landscapes, to ensure that these species do not undermine the conservation of forest remnants. This would involve consideration of maintaining large interior and closed canopy forests and reducing edge effects and propagule transport within the landscape. Vulnerable recovering forests in particular should be protected.
Our results highlight the consequences of fragmentation and logging on the invasion of remnant forests but also the impact on the native plant community. These findings are of critical importance given that around 80% of tropical forests are currently fragmented and/or anthropogenically modified (Potapov et al. 2017). Fragmentation is predicted to increase, with increases in the number of isolated forest remnants as well as decreases in their average size (Taubert et al. 2018) as a result of cyclical deforestation processes (i.e. deforestation and degradation making new areas of forest more accessible and susceptible to further degradation; Nowosad and Stepinski 2019). Therefore, invasion of forest remnants is likely to increase in future due to continuing deforestation, leading to changes in plant community composition and altered patterns of native regeneration, thereby potentially impacting local and regional biodiversity. Given, that maintaining regeneration of remnant forests is necessary for the sustainability of human dominated tropical landscapes, including RSPO-certified oil palm plantations (Senior et al. 2015), it is vital that the impacts of invading exotic species within these forests are properly evaluated and included in future studies.
Data availability
Data collected for this study are available to download (see Waddell et al. 2020).
References
Achard F, Eva HD, Stibig HJ et al (2002) Determination of deforestation rates of the world’s humid tropical forests. Science 297:999–1002
Anderson JM, Ingram JSI (1994) Tropical soil biology and fertility: a handbook of methods. Soil Sci 157:265
Ansong M, Pickering C (2013) Are weeds hitchhiking a ride on your car? A systematic review of seed dispersal on cars. PLoS ONE 8:e80275
Arellano-Rivas A, De-Nova JA, Munguía-Rosas MA (2016) Patch isolation and shape predict plant functional diversity in a naturally fragmented forest. J Plant Ecol 11:136–146
Bakar BH (2004) Invasive weed species in Malaysian agro-ecosystems: species, impacts and management. Malaysian J Sci 23:1–42
Bivand R, Rundel C (2013) rgeos: interface to geometry engine-open source (GEOS). R Packag version 03-2
Brearley FQ, Banin LF, Saner P (2016) The ecology of the Asian dipterocarps. Plant Ecol Divers 9:429–436
Brown KA, Scatena FN, Gurevitch J (2006) Effects of an invasive tree on community structure and diversity in a tropical forest in Puerto Rico. For Ecol Manage 226:145–152
Bryan JE, Shearman PL, Asner GP et al (2013) Extreme differences in forest degradation in Borneo: comparing practices in Sarawak, Sabah, and Brunei. PLoS ONE 8:e69679
CABI (2017) Invasive species compendium. CAB International, Wallingford
Carson W, Schnitzer S (2011) Tropical forest community ecology. Wiley, Hoboken
Catford JA, Jansson R, Nilsson C (2009) Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers Distrib 15:22–40
Chave J, Coomes D, Jansen S et al (2009) Towards a worldwide wood economics spectrum. Ecol Lett 12:351–366
Colautti RI, Grigorovich IA, MacIsaac HJ (2006) Propagule pressure: a null model for biological invasions. Biol Invasions 8:1023–1037
Curtis PG, Slay CM, Harris NL et al (2018) Classifying drivers of global forest loss. Science 361:1108–1111
Daehler CC (2003) Performance comparisons of co-occurring native and alien invasive plants: implications for conservation and restoration. Annu Rev Ecol Evol Syst 34:183–211
Davis MA, Grime JP, Thompson K et al (2000) In plant communities: fluctuating resources a general of invasibility theory. J Ecol 88:528–534
Dawson W, Burslem DFRP, Hulme PE (2014) Consistent effects of disturbance and forest edges on the invasion of a continental rain forest by alien plants. Biotropica 47:27–37
Denslow JS, DeWalt SJ (2008) Exotic plant invasions in tropical forests: patterns and hypotheses. In: Carson WP, Schnitzer SA (eds) Tropical forest community ecology. University of Chicago, Chicago, pp 409–426
Döbert TF, Webber BL, Sugau JB et al (2017) Logging, exotic plant invasions, and native plant reassembly in a lowland tropical rain forest. Biotropica. https://doi.org/10.1111/btp.12521
Edward E, Munishi PKT, Hulme PE (2009) Relative roles of disturbance and propagule pressure on the invasion of humid tropical forest by Cordia alliodora (Boraginaceae) in Tanzania. Biotropica 41:171–178
Faith DP (1992) Conservation evaluation and phylogentic diversity. Biol Conserv 61:1–10
Fee CG, Tui LC, Bin CS, Hoy CK (2017) Pictorial guide to common weeds of plantations and their control (No. L-0987), 2nd edn. Agricultural Crop Trust, ACT, Bonn
Feeley KJ, Davies SJ, Perez R et al (2011) Directional changes in the species composition of a tropical forest. Ecology 92:871–882
Ferraz G, Russell GJ, Stouffer PC et al (2003) Rates of species loss from Amazonian forest fragments. Proc Natl Acad Sci USA 100:14069–14073
Fine PVA (2002) The invasibility of tropical forests by exotic plants. Cambridge Univ Press Stable 18:687–705
Fleiss S, Waddell E, Ola BB, Banin LF, Benedick S, Sailim AB, Chapman DS, Jelling A, King H, McClean CJ, Yeong KL, Hill JK (2020) Conservation set-asides improve carbon storage and support associated plant diversity in certified sustainable oil palm plantations. Biol Conserv 248:108631
Flynn DFB, Mirotchnick N, Jain M et al (2011) Functional and phylogenetic diversity as predictors of biodiversity–ecosystem-function relationships. Ecology 92:1573–1581
Gaveau DLA, Salim M, Arjasakusuma S (2017) Deforestation and industrial plantations development in Borneo. Center for International Forestry Research, Bogor
Gerlach J (1993) Invasive melastomataceae in seychelles. Oryx 27:22–26
Ghazoul J, McLeish M (2001) Reproductive ecology of tropical forest trees in logged and fragmented habitats in Thailand and Costa Rica. Plant Ecol 153:335–345
Gibbs HK, Salmon JM (2015) Mapping the world’s degraded lands. Appl Geogr 57:12–21
Gonzalez A (2000) Community relaxation in fragmented landscapes: the relation between species richness, area and age. Ecol Lett 3:441–448
Gorchov DL, Trisel DE (2003) Competitive effects of the invasive shrub, Lonicera maackii (Rupr.) Herder (Caprifoliaceae), on the growth and survival of native tree seedlings. Plant Ecol 166:13–24
Haddad NM, Brudvig LA, Clobert J, Davies KF, Gonzalez A, Holt RD, Lovejoy TE, Sexton JO, Austin MP, Collins CD, Cook WM (2015) Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci Adv 1(2):e1500052
Hansen MC, Stehman SV, Potapov PV, Loveland TR, Townshend JR, DeFries RS, Pittman KW, Arunarwati B, Stolle F, Steininger MK, Carroll M (2008) Humid tropical forest clearing from 2000 to 2005 quantified by using multitemporal and multiresolution remotely sensed data. Proc Natl Acad Sci USA 105(27):9439–9444
Harper KA, Macdonald SE, Burton PJ, Chen J, Brosofske KD, Saunders SC, Euskirchen ES, Roberts DAR, Jaiteh MS, Esseen PA (2005) Edge influence on forest structure and composition in fragmented landscapes. Conserv Biol 19:768–782
Hijmans RJ, van Etten J (2012) raster: Geographic analysis and modeling with raster data. R package version 2.0-12
Hill JL, Curran PJ (2003) Area, shape and isolation of tropical forest fragments: effects on tree species diversity and implications for conservation. J Biogeogr 30:1391–1403
Hood WG, Naiman RJ (2000) Vulnerability of riparian zones to invasion by exotic vascular plants. Plant Ecol 148:105–114
Hu G, Zhang Z (2013) Allelopathic effects of Chromolaena odorata on native and non-native invasive herbs. J Food Agric Environ 11:878–882
Hulme PE (2009) Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J Appl Ecol 46:10–18
Ismaini L (2015) Pengaruh alelopati tumbuhan invasif (Clidemia hirta) terhadap germinasi biji tumbuhan asli (Impatiens platypetala). Jurnal Lemb Ilmu Pengetah Indones (LIPI) Jawa Barat
Jeschke J, Gómez Aparicio L, Haider S et al (2012) Support for major hypotheses in invasion biology is uneven and declining. NeoBiota 14:1–20
Kembel SW, Cowan PD, Helmus MR, Cornwell WK, Morlon H, Ackerly DD, Blomberg SP, Webb CO (2010) Picante: r tools for integrating phylogenies and ecology. Bioinformatics 26(11):1463–1464. https://doi.org/10.1093/bioinformatics/btq166
Kueffer C, Pysek P, Richardson DM (2013) Integrative invasion science: model systems, multi-site studies, focused meta-analysis and invasion syndromes. New Phytol 200:615–633
Laurance WF, Camargo JL, Luizão RC, Laurance SG, Pimm SL, Bruna EM, Stouffer PC, Williamson GB, Benítez-Malvido J, Vasconcelos HL, Van Houtan KS (2011) The fate of Amazonian forest fragments: a 32-year investigation. Biol Conserv 144:56–67
Laurance WF, Vasconcelos HL, Stouffer PC, Laurance SG (2002) Ecosystem decay of Amazonian forest fragments: a 22-year investigation. Conserv Biol 16:605–618
Laurance WF, Nascimento HEM, Laurance SG, Andrade AC, Fearnside PM, Ribeiro JEL, Capretz RL (2006) Rain forest fragmentation and the proliferation of successional trees. Ecology 87(2):469–482. https://doi.org/10.1890/05-0064
Levine JM, Adler PB, Yelenik SG (2004) A meta-analysis of biotic resistance to exotic plant invasions. Ecol Lett 7:975–989
Lewis SL, Edwards DP, Galbraith D (2015) Increasing human dominancee of tropical forests. Science 349:827–832
Lôbo D, Leão T, Melo FPL et al (2011) Forest fragmentation drives Atlantic forest of northeastern Brazil to biotic homogenization. Divers Distrib 17:287–296
Lockwood JL, Cassey P, Blackburn T (2005) The role of propagule pressure in explaining species invasions. Trends Ecol Evol 20:223–228
Lockwood JL, Hoopes MF, Marchetti MP (2013) Invasion ecology. Wiley, West Sussex
Lowe S, Browne M, Boudjelas S, De Poorter M (2000) 100 of the world’s worst invasive alien species: a selection from the global invasive species database. Invasive Species Specialist Group Auckland
Lucey JM, Palmer G, Yeong KL, Edwards DP, Senior MJ, Scriven SA, Reynolds G, Hill JK (2017) Reframing the evidence base for policy-relevance to increase impact: a case study on forest fragmentation in the oil palm sector. J Appl Ecol 54:731–736
Malhi Y, Gardner TA, Goldsmith GR et al (2014) Tropical forests in the Anthropocene. Annu Rev Environ Resour 39:125–159
McNeely JA (1995) The impact of human activity on biodiversity. Global biodiversity assessment. Cambridge University Press, Cambridge, pp 733–783
Nowosad J, Stepinski TF (2019) Stochastic, empirically-informed model of landscape dynamics and its application to deforestation scenarios. Geophys Res Lett 46(23):13845–13852
Padmanaba M, Sheil D (2014) Spread of the invasive alien species Piper aduncum via logging roads in Borneo. Trop Conserv Sci 7:35–44
Pallewatta N, Reaser JK, Gutierrez AT (2003) Invasive alien species in South-Southeast Asia: national reports and directory of resources.
Peh KSH (2010) Invasive species in Southeast Asia: the knowledge so far. Biodivers Conserv 19:1083–1099
Peters H (2001) Clidemia hirta invasion at the Pasoh Forest Reserve: an unexpected plant invasion in an undisturbed tropical forest. Biotropica 33:60–68
Potapov P, Hansen MC, Laestadius L, Turubanova S, Yaroshenko A, Thies C, Smith W, Zhuravleva I, Komarova A, Minnemeyer S, Esipova E (2017) The last frontiers of wilderness: tracking loss of intact forest landscapes from 2000 to 2013. Sci Adv 3:e1600821
Pütz S, Groeneveld J, Alves LF, Metzger JP, Huth A (2011) Fragmentation drives tropical forest fragments to early successional states: a modelling study for Brazilian Atlantic forests. Ecol Model 222(12):1986–1997. https://doi.org/10.1016/j.ecolmodel.2011.03.038
Pütz S, Groeneveld J, Henle K, Knogge C, Martensen AC, Metz M, Metzger JP, Ribeiro MC, De Paula MD, Huth A (2014) Long-term carbon loss in fragmented Neotropical forests. Nat Commun 5:5037
Pyšek P, Richardson DM (2006) The biogeography of naturalization in alien plants. J Biogeogr 33:2040–2050
Qie L, Lewis SL, Sullivan MJ, Lopez-Gonzalez G, Pickavance GC, Sunderland T, Ashton P, Hubau W, Salim KA, Aiba SI, Banin LF (2017) Long-term carbon sink in Borneo’s forests halted by drought and vulnerable to edge effects. Nat Commun. https://doi.org/10.1038/s41467-017-01997-0
R Core Team. (2019). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Retrieved from https://www.r-project.org/
Rejmánek M, Richardson DM (1996) What attributes make some plant species more invasive? Ecology 77:1655–1661
Réjou-Méchain M, Tanguy A, Piponiot C et al (2017) Biomass: an R Package for estimating above-ground biomass and its uncertainty in tropical forests. Methods Ecol Evol 8:1163–1167
Reynolds G, Payne J, Sinun W et al (2011) Changes in forest land use and management in Sabah, Malaysian Borneo, 1990–2010, with a focus on the Danum Valley region. Philos Trans R Soc B Biol Sci 366:3168–3176
RSPO (2018) Roundtable on sustainable palm oil principles & criteria for sustainable palm oil. Retrieved from https://rspo.org/resources/certification/rspo-principles-criteria-certification
Sala OE, Chapin FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leemans R (2005) Biodiversity: global biodiversity scenarios for the year 2100. Science 287:1770–1775
Sanchez G (2013) PLS path modeling with R. Trowchez Editions, Berkeley
Sanchez G, Trinchera L (2012) plspm: Partial Least Squares data analysis methods. R Packag version 02-2. http://www.CRANR-projectorg/package=plspm
Saner P, Loh YY, Ong RC, Hector A (2012) Carbon stocks and fluxes in tropical lowland dipterocarp rainforests in Sabah, Malaysian Borneo. PLoS ONE 7:e29642
Scriven SA, Carlson KM, Hodgson JA, et al (2017) The impact of RSPO membership on avoiding biodiversity losses in oil palm landscapes. Sci Brief Sens Program. https://www.sensorprojectnet/wp-content/uploads/2017/09/SEnSOR-HCV-Project-Two-Page-Report-FINAL.pdf
Senior MJM, Brown E, Villalpando P, Hill JK (2015) Increasing the scientific evidence base in the “high conservation value”(HCV) approach for biodiversity conservation in managed tropical landscapes. Conserv Lett 8:361–367
Singhakumara BMP, Uduporuwa RSJP, Ashton PMS (2000) Soil seed banks in relation to light and topographic position of a hill dipterocarp forest in Sri Lanka. Biotropica 32:190–197
Slik JWF, Bernard CS, Breman FC et al (2008) Wood density as a conservation tool: quantification of disturbance and identification of conservation-priority areas in tropical forests. Conserv Biol 22:1299–1308
Slik JWF, Poulsen AD, Ashton PS, Cannon CH, Eichhorn KAO, Kartawinata K, Lanniari I, Nagamasu H, Nakagawa M, Van Nieuwstadt MGL, Payne J (2003) A floristic analysis of the lowland dipterocarp forests of Borneo. J Biogeogr 30:1517–1531
Sodhi NS, Koh LP, Brook BW, Ng PKL (2004) Southeast Asian biodiversity: an impending disaster. Trends Ecol Evol. https://doi.org/10.1016/j.tree.2004.09.006
Stinson KA, Campbell SA, Powell JR, Wolfe BE, Callaway RM, Thelen GC, Hallett SG, Prati D, Klironomos JN (2006) Invasive plant suppresses the growth of native tree seedlings by disrupting belowground mutualisms. PLoS Biol 4:727–731
Stride G, Thomas CD, Benedick S et al (2018) Contrasting patterns of local richness of seedlings, saplings, and trees may have implications for regeneration in rainforest remnants. Biotropica 50:1–9
Taubert F, Fischer R, Groeneveld J, Lehmann S, Müller MS, Rödig E, Wiegand T, Huth A (2018) Global patterns of tropical forest fragmentation. Nature 554(7693):519–522. https://doi.org/10.1038/nature25508
Teo DHL, Tan HTW, Corlett RT et al (2003) Continental rain forest fragments in Singapore resist invasion by exotic plants. J Biogeogr 30:305–310
Vilà M, Ibáñez I (2011) Plant invasions in the landscape. Landsc Ecol 26:461–472
Von der Lippe M, Kowarik I (2007) Long-distance dispersal of plants by vehicles as a driver of plant invasions. Conserv Biol 21:986–996
Waddell EH, Chapman DS (2020) R code to reproduce analyses of exotic plant invasion in fragmented and continuous forest sites in Sabah, Malaysian Borneo, 2017. NERC Environ Inf Data Cent. https://doi.org/10.5285/adbf6d29-ee7b-4dd1-9730-11d2308d526c
Waddell EH, Fleiss S, Bala Ola B, Sailim AB, Jelling A, Yeong KL, Tangah J, Benedick S, King H, Hughes M, McClean CJ, Chapman DS, Hill JK, Banin LF (2020) Vegetation and habitat data for fragmented and continuous forest sites in Sabah, Malaysian Borneo, 2017. NERC Environ Inf Data Cent. https://doi.org/10.5285/c67b06b7-c3f6-49a3-baf2-9fc3a72cb98a
Webb CO, Donoghue MJ (2005) Phylomatic: tree assembly for applied phylogenetics. Mol Ecol Notes 5(1):181–183. https://doi.org/10.1111/j.1471-8286.2004.00829.x
Webb CO, Ackerly DD, Kembel SW (2008) Phylocom: software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics 24(18):2098–2100. https://doi.org/10.1093/bioinformatics/btn358
Wester LL, Wood HB (1977) Koster’s curse (Clidemia hirta), a weed pest in Hawaiian forests. Environ Conserv 4:35–41
Wikstrom N, Savolainen V, Chase MW (2001) Evolution of the angiosperms: calibrating the family tree. Proceedings Royal Soc B: Biol Sci 268:2211–2220. https://doi.org/10.1098/rspb.2001.1782
Williams M (2003) Deforesting the earth: from prehistory to global crisis. University of Chicago Press
With KA (2002) The landscape ecology of invasive species. Conserv Biol 16:1192–1203
Yeong KL, Reynolds G, Hill JK (2016) Enrichment planting to improve habitat quality and conservation value of tropical rainforest fragments. Biodivers Conserv 25:957–973
Zanne AE, Lopez-Gonzalez G, Coomes DA, Ilic J, Jansen S, Lewis SL, Miller RB, Swenson NG, Wiemann MC, Chave J (2009) Global wood density database
Acknowledgements
We thank Sabah Biodiversity Council, Danum Valley Management Committee, Sabah Forestry Department and the South East Asia Rainforest Research Partnership (SEARRP) for permissions to carry out research. We thank Noreen Majalap, John Baptist Sugau and Suzana Sabran at the Forest Research Centre for help analysing soil samples and plant identification. We thank Wilmar Int. Ltd for allowing us access to sites and for their support throughout fieldwork, especially Ginny Ng and Marcie Elene. We thank Robin Hayward for assisting during fieldwork. We thank two anonymous reviewers for comments improving the manuscript.
Funding
EHW was funded by the Natural Environment Research Council ACCE doctoral training programme NE/L002450/1.
Author information
Authors and Affiliations
Contributions
EHW, LFB, MH, JKH and DSC conceived the ideas. EHW, LFB, MH, SF, JT, JKH, CB, YKL, ABS, AJ and DSC designed methodology; EHW, SF, ABS and AJ collected the data; BBO and AJ led plant identification; EHW and DSC analysed the data; EHW led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Authors report no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Waddell, E.H., Banin, L.F., Fleiss, S. et al. Land-use change and propagule pressure promote plant invasions in tropical rainforest remnants. Landscape Ecol 35, 1891–1906 (2020). https://doi.org/10.1007/s10980-020-01067-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-020-01067-9