Frog Skin-Derived Peptides Against Corynebacterium jeikeium: Correlation between Antibacterial and Cytotoxic Activities

 ,
, 

Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Activity of Frog Skin-Derived AMPs
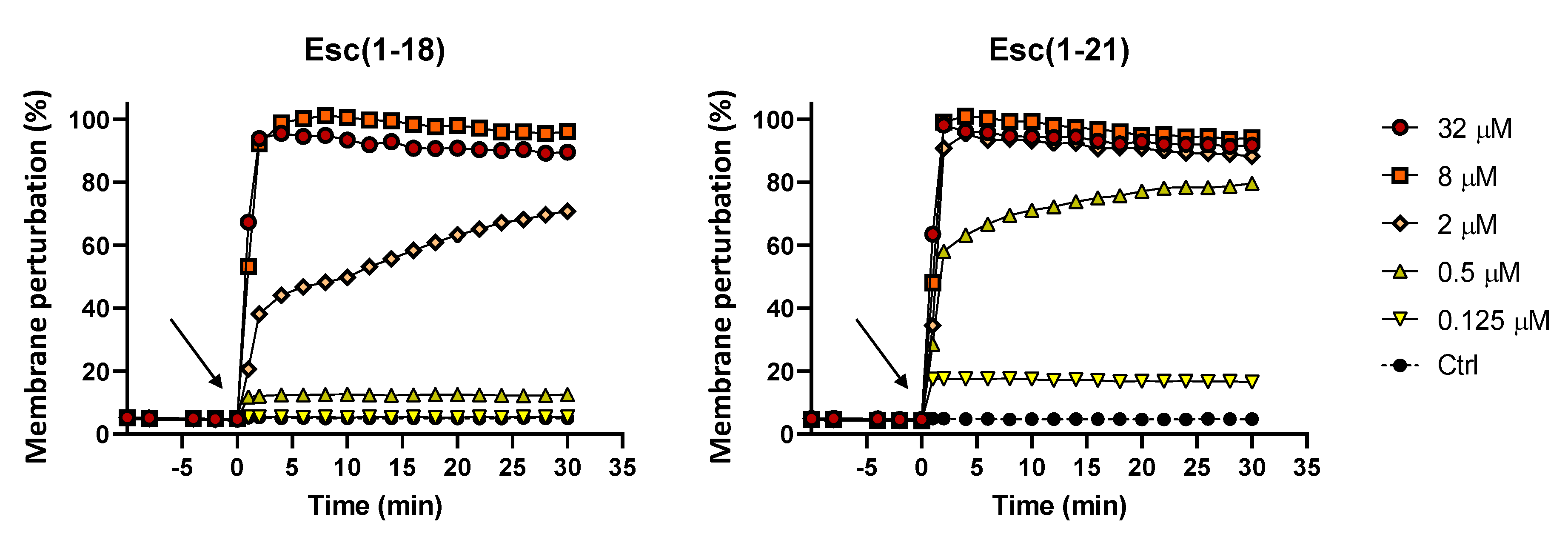
2.2. Membrane Permeabilization
2.3. Cytotoxicity
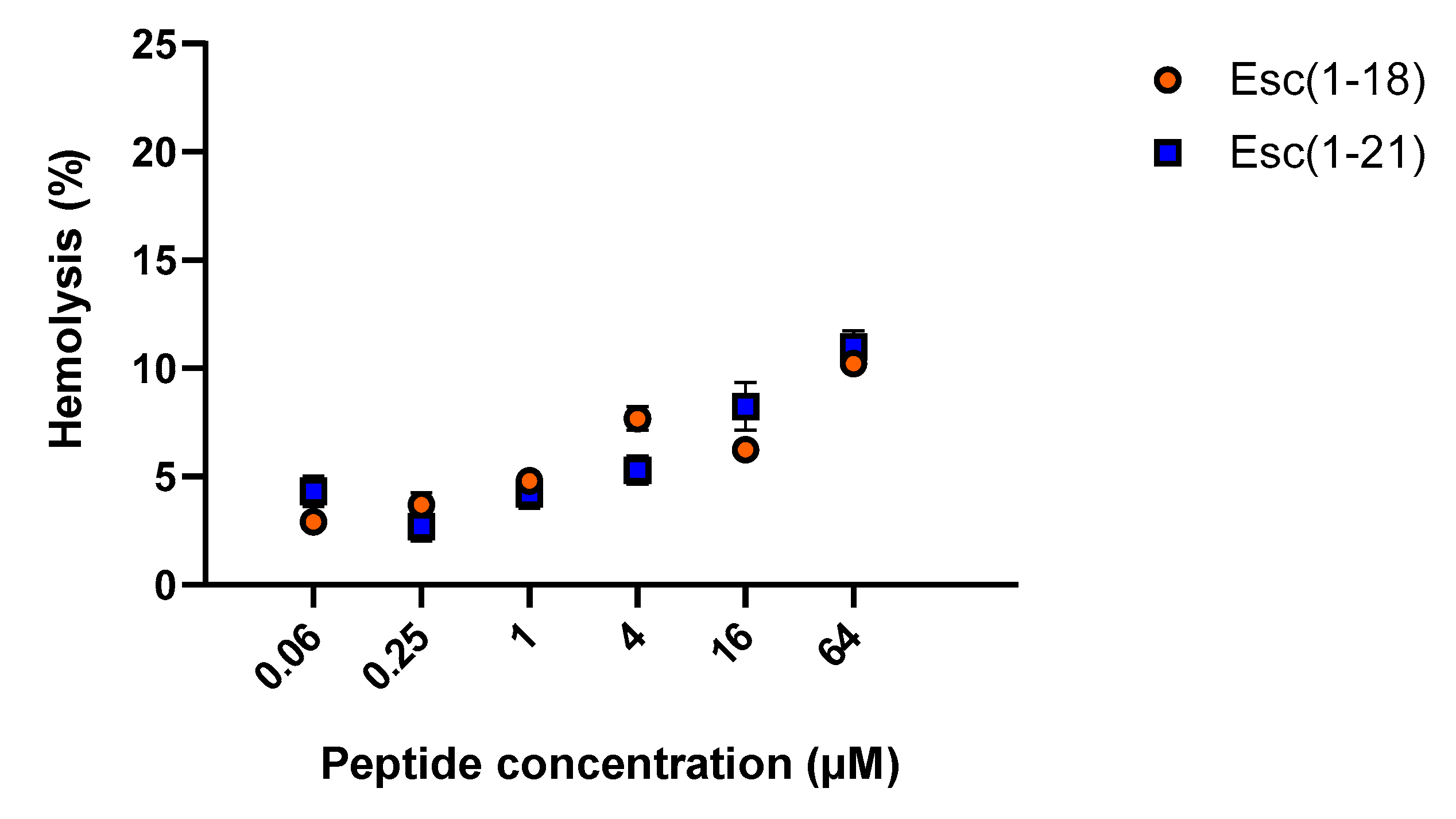
2.3.1. Hemolytic Activity
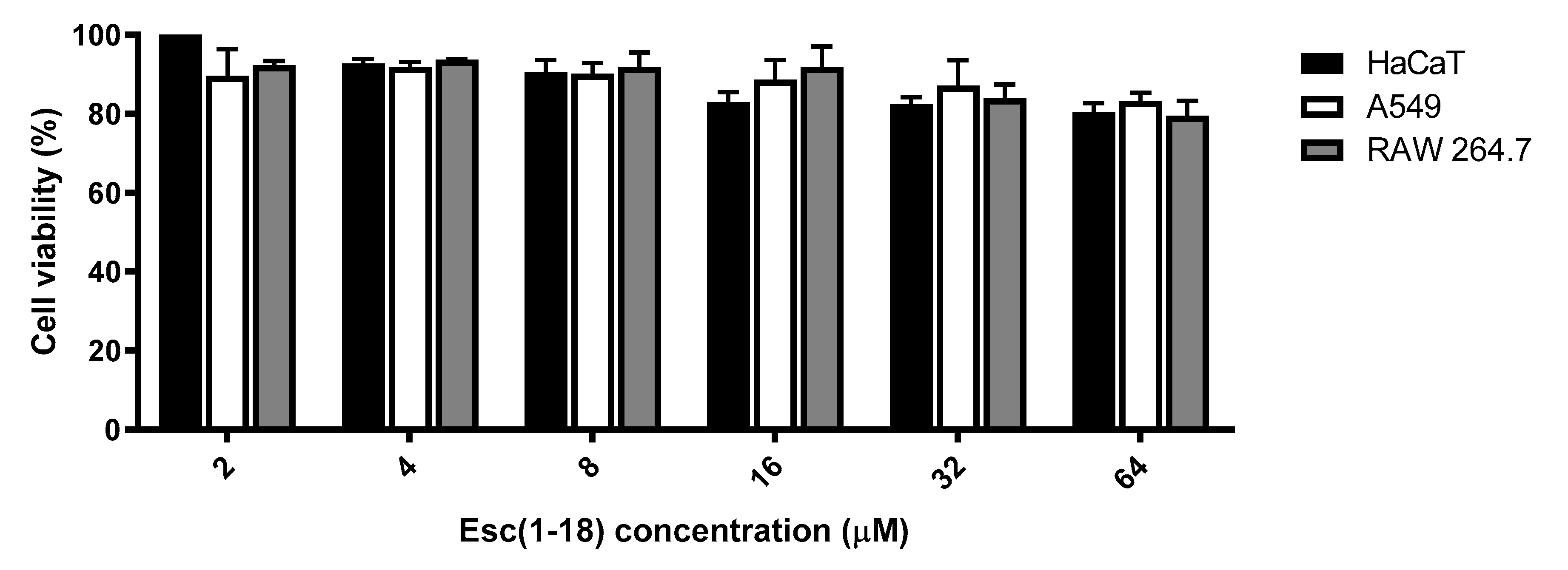
2.3.2. Peptides’ Effect on the Metabolic Activity of Mammalian Cells
3. Discussion
4. Materials and Methods
4.1. Microorganism and Cell Lines
4.2. Peptides
4.3. Antimicrobial Assays
4.4. Membrane Permeabilization: Sytox Green Assay
4.5. Hemolytic Assay
4.6. Cytotoxicity Test on Mammalian Cell Lines
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bernard, K. The genus corynebacterium and other medically relevant coryneform-like bacteria. J. Clin. Microbiol. 2012, 50, 3152–3158. [Google Scholar] [CrossRef] [Green Version]
- Cogen, A.L.; Nizet, V.; Gallo, R.L. Skin microbiota: a source of disease or defence? Br. J. Dermatol. 2008, 158, 442–455. [Google Scholar] [CrossRef] [Green Version]
- Coyle, M.B.; Lipsky, B.A. Coryneform bacteria in infectious diseases: clinical and laboratory aspects. Clin. Microbiol. Rev. 1990, 3, 227–246. [Google Scholar] [CrossRef] [PubMed]
- Larson, E.L.; McGinley, K.J.; Leyden, J.J.; Cooley, M.E.; Talbot, G.H. Skin colonization with antibiotic-resistant (JK group) and antibiotic-sensitive lipophilic diphtheroids in hospitalized and normal adults. J. Infect. Dis. 1986, 153, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Altonsy, M.O.; Kurwa, H.A.; Lauzon, G.J.; Amrein, M.; Gerber, A.N.; Almishri, W.; Mydlarski, P.R. Corynebacterium tuberculostearicum, a human skin colonizer, induces the canonical nuclear factor-kappaB inflammatory signaling pathway in human skin cells. Immun. Inflamm. Dis. 2020, 8, 62–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barzantny, H.; Brune, I.; Tauch, A. Molecular basis of human body odour formation: insights deduced from corynebacterial genome sequences. Int. J. Cosmet. Sci. 2012, 34, 2–11. [Google Scholar] [CrossRef] [PubMed]
- James, A.G.; Austin, C.J.; Cox, D.S.; Taylor, D.; Calvert, R. Microbiological and biochemical origins of human axillary odour. FEMS Microbiol. Ecol. 2013, 83, 527–540. [Google Scholar] [CrossRef]
- Noble, W.C. Short Communications: Activity of Corynebacterium jeikeium Bacteriocin in Vivo. Microb. Ecol. Health Disease 1988, 1, 201–203. [Google Scholar] [CrossRef]
- Proenca, J.T.; Barral, D.C.; Gordo, I. Commensal-to-pathogen transition: One-single transposon insertion results in two pathoadaptive traits in Escherichia coli -macrophage interaction. Sci. Rep. 2017, 7, 4504. [Google Scholar] [CrossRef]
- Littman, D.R.; Pamer, E.G. Role of the commensal microbiota in normal and pathogenic host immune responses. Cell Host Microbe 2011, 10, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Gallo, R.L. Commensal skin bacteria as the probiotic of the cutaneous immune response. Expert Rev. Dermatol. 2010, 5, 251–253. [Google Scholar] [CrossRef] [Green Version]
- Rath, S.; Rud, T.; Karch, A.; Pieper, D.H.; Vital, M. Pathogenic functions of host microbiota. Microbiome 2018, 6, 174. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Cancio, M.R. Infections and the compromised immune status in the chronically critically ill patient: prevention strategies. Respir. Care 2012, 57, 979–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardo, S.M.M.; Patel, R.H.; Ramsakal, A.; Greene, J. Disseminated Corynebacterium jeikeium Infection in Cancer Patients. Cureus 2020, 12. [Google Scholar] [CrossRef]
- Yang, K.; Kruse, R.L.; Lin, W.J.V.; Musher, D.M. Corynebacteria as a cause of pulmonary infection: a case series and literature review. Pneumonia 2018, 10. [Google Scholar] [CrossRef]
- Letek, M.; Ordonez, E.; Fernandez-Natal, I.; Gil, J.A.; Mateos, L.M. Identification of the emerging skin pathogen Corynebacterium amycolatum using PCR-amplification of the essential divIVA gene as a target. FEMS Microbiol. Lett. 2006, 265, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Haney, E.F.; Straus, S.K.; Hancock, R.E.W. Reassessing the Host Defense Peptide Landscape. Front. Chem. 2019, 7, 43. [Google Scholar] [CrossRef] [Green Version]
- Di Somma, A.; Moretta, A.; Cane, C.; Cirillo, A.; Duilio, A. Antimicrobial and Antibiofilm Peptides. Biomolecules 2020, 10, 652. [Google Scholar] [CrossRef] [Green Version]
- Marcocci, M.E.; Amatore, D.; Villa, S.; Casciaro, B.; Aimola, P.; Franci, G.; Grieco, P.; Galdiero, M.; Palamara, A.T.; Mangoni, M.L.; et al. The Amphibian Antimicrobial Peptide Temporin B Inhibits In Vitro Herpes Simplex Virus 1 Infection. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Lupetti, A.; Danesi, R.; van ’t Wout, J.W.; van Dissel, J.T.; Senesi, S.; Nibbering, P.H. Antimicrobial peptides: therapeutic potential for the treatment of Candida infections. Expert Opin. Investig. Drugs 2002, 11, 309–318. [Google Scholar] [CrossRef]
- Lazzaro, B.P.; Zasloff, M.; Rolff, J. Antimicrobial peptides: Application informed by evolution. Science 2020, 368. [Google Scholar] [CrossRef] [PubMed]
- Bondaryk, M.; Staniszewska, M.; Zielinska, P.; Urbanczyk-Lipkowska, Z. Natural Antimicrobial Peptides as Inspiration for Design of a New Generation Antifungal Compounds. J. Fungi (Basel) 2017, 3, 46. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Siman-Tov, G.; Hall, G.; Bhalla, N.; Narayanan, A. Human Antimicrobial Peptides as Therapeutics for Viral Infections. Viruses 2019, 11, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musale, V.; Casciaro, B.; Mangoni, M.L.; Abdel-Wahab, Y.H.A.; Flatt, P.R.; Conlon, J.M. Assessment of the potential of temporin peptides from the frog Rana temporaria (Ranidae) as anti-diabetic agents. J. Pept. Sci. 2018, 24. [Google Scholar] [CrossRef] [PubMed]
- Thapa, R.K.; Diep, D.B.; Tonnesen, H.H. Topical antimicrobial peptide formulations for wound healing: Current developments and future prospects. Acta Biomater. 2020, 103, 52–67. [Google Scholar] [CrossRef]
- Brunetti, J.; Roscia, G.; Lampronti, I.; Gambari, R.; Quercini, L.; Falciani, C.; Bracci, L.; Pini, A. Immunomodulatory and Anti-inflammatory Activity in Vitro and in Vivo of a Novel Antimicrobial Candidate. J. Biol. Chem. 2016, 291, 25742–25748. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Lee, J.; Vu, T.H.; Lee, S.; Lillehoj, H.S.; Hong, Y.H. Immunomodulatory effects of avian beta-defensin 5 in chicken macrophage cell line. Res. Vet. Sci. 2020, 132, 81–87. [Google Scholar] [CrossRef]
- Ting, D.S.J.; Beuerman, R.W.; Dua, H.S.; Lakshminarayanan, R.; Mohammed, I. Strategies in Translating the Therapeutic Potentials of Host Defense Peptides. Front. Immunol. 2020, 11, 983. [Google Scholar] [CrossRef]
- Yang, B.; Good, D.; Mosaiab, T.; Liu, W.; Ni, G.; Kaur, J.; Liu, X.; Jessop, C.; Yang, L.; Fadhil, R.; et al. Significance of LL-37 on Immunomodulation and Disease Outcome. Biomed. Res. Int. 2020, 2020, 8349712. [Google Scholar] [CrossRef]
- Rauf, A.; Ali Shariati, M.; Ahmed Khalil, A.; Bawazeer, S.; Heydari, M.; Plygun, S.; Laishevtcev, A.; Bilal Hussain, M.; Alhumaydhi, F.A.; Aljohani, A.S.M. Hepcidin, an Overview of Biochemical and Clinical Properties. Steroids 2020, 108661. [Google Scholar] [CrossRef]
- Barik, A.; Pandiyan, R.; Manthiram, M.; Vellaisamy, B. Potential Antimicrobial Peptides Elucidation From The Marine Bacteria. Cardiovasc. Hematol. Agents Med. Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Sonnevend, A. Antimicrobial peptides in frog skin secretions. Methods Mol. Biol. 2010, 618, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Ladram, A.; Nicolas, P. Antimicrobial peptides from frog skin: biodiversity and therapeutic promises. Front. Biosci. (Landmark Ed) 2016, 21, 1341–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Mangoni, M.L.; Di, Y.P. In vivo therapeutic efficacy of frog skin-derived peptides against Pseudomonas aeruginosa-induced pulmonary infection. Sci. Rep. 2017, 7, 8548. [Google Scholar] [CrossRef] [Green Version]
- Luca, V.; Olivi, M.; Di Grazia, A.; Palleschi, C.; Uccelletti, D.; Mangoni, M.L. Anti-Candida activity of 1-18 fragment of the frog skin peptide esculentin-1b: in vitro and in vivo studies in a Caenorhabditis elegans infection model. Cell Mol. Life Sci. 2014, 71, 2535–2546. [Google Scholar] [CrossRef]
- Islas-Rodriguez, A.E.; Marcellini, L.; Orioni, B.; Barra, D.; Stella, L.; Mangoni, M.L. Esculentin 1-21: a linear antimicrobial peptide from frog skin with inhibitory effect on bovine mastitis-causing bacteria. J. Pept. Sci. 2009, 15, 607–614. [Google Scholar] [CrossRef]
- Kolar, S.S.N.; Luca, V.; Baidouri, H.; Mannino, G.; McDermott, A.M.; Mangoni, M.L. Esculentin-1a(1-21)NH2: a frog skin-derived peptide for microbial keratitis. Cell Mol. Life Sci. 2015, 72, 617–627. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.Y.; Yang, J.R. Analysis and prediction of highly effective antiviral peptides based on random forests. PLoS ONE 2013, 8, e70166. [Google Scholar] [CrossRef] [Green Version]
- ExPasy: Bioinformatics Resourch Portal. Available online: https://web.expasy.org/protparam (accessed on 17 July 2020).
- Di Grazia, A.; Cappiello, F.; Imanishi, A.; Mastrofrancesco, A.; Picardo, M.; Paus, R.; Mangoni, M.L. The Frog Skin-Derived Antimicrobial Peptide Esculentin-1a(1-21)NH2 Promotes the Migration of Human HaCaT Keratinocytes in an EGF Receptor-Dependent Manner: A Novel Promoter of Human Skin Wound Healing? PLoS ONE 2015, 10, e0128663. [Google Scholar] [CrossRef] [Green Version]
- Di Grazia, A.; Cappiello, F.; Cohen, H.; Casciaro, B.; Luca, V.; Pini, A.; Di, Y.P.; Shai, Y.; Mangoni, M.L. D-Amino acids incorporation in the frog skin-derived peptide esculentin-1a(1-21)NH2 is beneficial for its multiple functions. Amino Acids 2015, 47, 2505–2519. [Google Scholar] [CrossRef]
- Ordonez-Palau, S.; Boquet, D.; Gil-Garcia, M.; Pardina-Solano, M. Chronic osteomyelitis of the metatarsal sesamoid due to Corynebacterium jeikeium in a patient with rheumatoid arthritis. Joint Bone Spine 2007, 74, 516–517. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.T.; Huang, J.W.; Yen, C.J. A rare and under-recognized pathogen in peritoneal dialysis peritonitis: Corynebacterium jeikeium. Perit. Dial. Int. 2013, 33, 580–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, B.L. Pathology of culture-proven JK Corynebacterium pneumonia. An autopsy case report. Am. J. Clin. Pathol. 1989, 91, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Turett, G.S.; Fazal, B.A.; Johnston, B.E.; Telzak, E.E. Liver abscess due to Corynebacterium jeikeium in a patient with AIDS. Clin. Infect. Dis. 1993, 17, 514–515. [Google Scholar] [CrossRef] [PubMed]
- Greene, K.A.; Clark, R.J.; Zabramski, J.M. Ventricular CSF shunt infections associated with Corynebacterium jeikeium: report of three cases and review. Clin. Infect. Dis. 1993, 16, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Yoshitomi, Y.; Kohno, S.; Koga, H.; Maesaki, S.; Higashiyama, Y.; Matsuda, H.; Mitsutake, K.; Miyazaki, Y.; Yamada, H.; Hara, K.; et al. Fatal pneumonia caused by Corynebacterium group JK after treatment of Staphylococcus aureus pneumonia. Intern. Med. 1992, 31, 930–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.C.; Mattson, D.; Wald, A. Corynebacterium jeikeium bacteremia in bone marrow transplant patients with Hickman catheters. Bone Marrow Transpl. 2001, 27, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Tleyjeh, I.M.; Qutub, M.O.; Bakleh, M.; Sohail, M.R.; Virk, A. Corynebacterium jeikeium prosthetic joint infection: case report and literature review. Scand. J. Infect. Dis. 2005, 37, 151–153. [Google Scholar]
- Schiffl, H.; Mucke, C.; Lang, S.M. Exit-site infections by non-diphtheria corynebacteria in CAPD. Perit. Dial. Int. 2004, 24, 454–459. [Google Scholar] [CrossRef]
- Olender, A. Antibiotic resistance and detection of the most common mechanism of resistance (MLSB) of opportunistic Corynebacterium. Chemotherapy 2013, 59, 294–306. [Google Scholar] [CrossRef]
- Balci, I.; Eksi, F.; Bayram, A. Coryneform bacteria isolated from blood cultures and their antibiotic susceptibilities. J. Int. Med. Res. 2002, 30, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Philippon, A.; Bimet, F. In vitro susceptibility of Corynebacterium group D2 and Corynebacterium jeikeium to twelve antibiotics. Eur. J. Clin. Microbiol. Infect. Dis. 1990, 9, 892–895. [Google Scholar] [CrossRef] [PubMed]
- Neemuchwala, A.; Soares, D.; Ravirajan, V.; Marchand-Austin, A.; Kus, J.V.; Patel, S.N. In Vitro Antibiotic Susceptibility Pattern of Non-diphtheriae Corynebacterium Isolates in Ontario, Canada, from 2011 to 2016. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Mondal, J.; Ghosh, P.; Zhu, X. An Atomistic view of Short-chain Antimicrobial Biomimetic peptides in Action. bioRxiv 2018, 323592. [Google Scholar] [CrossRef]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casciaro, B.; Loffredo, M.R.; Luca, V.; Verrusio, W.; Cacciafesta, M.; Mangoni, M.L. Esculentin-1a Derived Antipseudomonal Peptides: Limited Induction of Resistance and Synergy with Aztreonam. Protein Pept. Lett. 2018, 25, 1155–1162. [Google Scholar] [CrossRef]
- Huang, H.W. DAPTOMYCIN, its membrane-active mechanism vs. that of other antimicrobial peptides. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183395. [Google Scholar] [CrossRef]
- Raheem, N.; Straus, S.K. Mechanisms of Action for Antimicrobial Peptides With Antibacterial and Antibiofilm Functions. Front. Microbiol. 2019, 10, 2866. [Google Scholar] [CrossRef] [Green Version]
- Bechinger, B.; Gorr, S.U. Antimicrobial Peptides: Mechanisms of Action and Resistance. J. Dent. Res. 2017, 96, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D.; et al. HaCaT Cells as a Reliable In Vitro Differentiation Model to Dissect the Inflammatory/Repair Response of Human Keratinocytes. Mediat. Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef]
- Petushkova, N.A.; Rusanov, A.L.; Pyatnitskiy, M.A.; Larina, O.V.; Zgoda, V.G.; Lisitsa, A.V.; Luzgina, N.G. Proteomic characterization of HaCaT keratinocytes provides new insights into changes associated with SDS exposure. Biomed. Dermatol. 2020, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V. Macrophages: The Potent Immunoregulatory Innate Immune Cells. In Macrophage Activation-Biology and Disease; Bhat, K.H., Ed.; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef] [Green Version]
- Hawdon, N.A.; Aval, P.S.; Barnes, R.J.; Gravelle, S.K.; Rosengren, J.; Khan, S.; Ciofu, O.; Johansen, H.K.; Høiby, N.; Ulanova, M. Cellular responses of A549 alveolar epithelial cells to serially collected Pseudomonas aeruginosa from cystic fibrosis patients at different stages of pulmonary infection. FEMS Immunol. Med. Microbiol. 2010, 59, 207–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, K.A.; Oster, C.G.; Mayer, M.M.; Avery, M.L.; Audus, K.L. Characterization of the A549 cell line as a type II pulmonary epithelial cell model for drug metabolism. Exp. Cell Res. 1998, 243, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Barzantny, H.; Schroder, J.; Strotmeier, J.; Fredrich, E.; Brune, I.; Tauch, A. The transcriptional regulatory network of Corynebacterium jeikeium K411 and its interaction with metabolic routes contributing to human body odor formation. J. Biotechnol. 2012, 159, 235–248. [Google Scholar] [CrossRef]
- Kursun, S.; Acar, B.; Atakan, C.; Oztas, B.; Paksoy, C.S. Relationship between genuine and pseudohalitosis and social anxiety disorder. J. Oral Rehabil. 2014, 41, 822–828. [Google Scholar] [CrossRef]
- Mogilnicka, I.; Bogucki, P.; Ufnal, M. Microbiota and Malodor-Etiology and Management. Int. J. Mol. Sci. 2020, 21, 2886. [Google Scholar] [CrossRef]
- AlgPred: Prediction of Allergenic Proteins and Mapping of IgE Epitopes. Available online: https://webs.iiitd.edu.in/raghava/algpred/submission.html (accessed on 17 July 2020).
- Casciaro, B.; d’Angelo, I.; Zhang, X.; Loffredo, M.R.; Conte, G.; Cappiello, F.; Quaglia, F.; Di, Y.P.; Ungaro, F.; Mangoni, M.L. Poly(lactide- co-glycolide) Nanoparticles for Prolonged Therapeutic Efficacy of Esculentin-1a-Derived Antimicrobial Peptides against Pseudomonas aeruginosa Lung Infection: in Vitro and in Vivo Studies. Biomacromolecules 2019, 20, 1876–1888. [Google Scholar] [CrossRef]
- Cappiello, F.; Ranieri, D.; Carnicelli, V.; Casciaro, B.; Chen, H.T.; Ferrera, L.; Di, Y.P.; Mangoni, M.L. Bronchial epithelium repair by Esculentin-1a-derived antimicrobial peptides: involvement of metalloproteinase-9 and interleukin-8, and evaluation of peptides’ immunogenicity. Sci. Rep. 2019, 9, 18988. [Google Scholar] [CrossRef] [Green Version]
- Rahnamaeian, M.; Vilcinskas, A. Short antimicrobial peptides as cosmetic ingredients to deter dermatological pathogens. Appl. Microbiol. Biotechnol. 2015, 99, 8847–8855. [Google Scholar] [CrossRef] [Green Version]
- Haisma, E.M.; Goblyos, A.; Ravensbergen, B.; Adriaans, A.E.; Cordfunke, R.A.; Schrumpf, J.; Limpens, R.W.; Schimmel, K.J.; den Hartigh, J.; Hiemstra, P.S.; et al. Antimicrobial Peptide P60.4Ac-Containing Creams and Gel for Eradication of Methicillin-Resistant Staphylococcus aureus from Cultured Skin and Airway Epithelial Surfaces. Antimicrob. Agents Chemother. 2016, 60, 4063–4072. [Google Scholar] [CrossRef] [Green Version]
- Casciaro, B.; Calcaterra, A.; Cappiello, F.; Mori, M.; Loffredo, M.R.; Ghirga, F.; Mangoni, M.L.; Botta, B.; Quaglio, D. Nigritanine as a New Potential Antimicrobial Alkaloid for the Treatment of Staphylococcus aureus-Induced Infections. Toxins (Basel) 2019, 11, 511. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | MIC (µM) | Net Charge | GRAVY |
|---|---|---|---|---|
| Esc(1-18) | GIFSKLAGKKLKNLLISG-NH2 | 4 | +5 | 0.383 |
| Esc(1-21) | GIFSKLAGKKIKNLLISGLKG-NH2 | 0.125 | +6 | 0.338 |
| Temporin A | FLPLIGRVLSGIL-NH2 | 8 | +2 | 1.808 |
| Temporin B | LLPIVGNLLKSLL-NH2 | 8 | +2 | 1.638 |
| Temporin G | FFPVIGRILNGIL-NH2 | 16 | +2 | 1.577 |
| Bombinin H2 | IIGPVLGLVGSALGGLLKKI-NH2 | 8 | +3 | 1.525 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casciaro, B.; Loffredo, M.R.; Cappiello, F.; Verrusio, W.; Corleto, V.D.; Mangoni, M.L. Frog Skin-Derived Peptides Against Corynebacterium jeikeium: Correlation between Antibacterial and Cytotoxic Activities. Antibiotics 2020, 9, 448. https://doi.org/10.3390/antibiotics9080448
Casciaro B, Loffredo MR, Cappiello F, Verrusio W, Corleto VD, Mangoni ML. Frog Skin-Derived Peptides Against Corynebacterium jeikeium: Correlation between Antibacterial and Cytotoxic Activities. Antibiotics. 2020; 9(8):448. https://doi.org/10.3390/antibiotics9080448
Chicago/Turabian StyleCasciaro, Bruno, Maria Rosa Loffredo, Floriana Cappiello, Walter Verrusio, Vito Domenico Corleto, and Maria Luisa Mangoni. 2020. "Frog Skin-Derived Peptides Against Corynebacterium jeikeium: Correlation between Antibacterial and Cytotoxic Activities" Antibiotics 9, no. 8: 448. https://doi.org/10.3390/antibiotics9080448