
Abstract
T-type, low-voltage activated, calcium channels, now designated Cav3 channels, are involved in a wide variety of physiological functions, especially in nervous systems. Their unique electrophysiological properties allow them to finely regulate neuronal excitability and to contribute to sensory processing, sleep, and hormone and neurotransmitter release. In the last two decades, genetic studies, including exploration of knock-out mouse models, have greatly contributed to elucidate the role of Cav3 channels in normal physiology, their regulation, and their implication in diseases. Mutations in genes encoding Cav3 channels (CACNA1G, CACNA1H, and CACNA1I) have been linked to a variety of neurodevelopmental, neurological, and psychiatric diseases designated here as neuronal Cav3 channelopathies. In this review, we describe and discuss the clinical findings and supporting in vitro and in vivo studies of the mutant channels, with a focus on de novo, gain-of-function missense mutations recently discovered in CACNA1G and CACNA1H. Overall, the studies of the Cav3 channelopathies help deciphering the pathogenic mechanisms of corresponding diseases and better delineate the properties and physiological roles Cav3 channels.
Similar content being viewed by others


Introduction
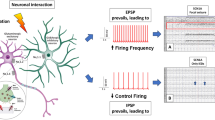
In the early 1980s, Llinas and Yarom [91] reported that hyperpolarization of inferior olivary neurons of the cerebellum could reveal a low-threshold Ca2+ conductance, which was inactivated at their resting membrane potential. The concept of low-voltage activated (LVA) Ca2+ current then arose in the neuroscience community as this conductance was described in many different types of neurons, including thalamic [90], sensory [19, 152], and hippocampal [165] neurons. This LVA Ca2+ current, also typical for its fast inactivation (Transient) and small unitary conductance (Tiny), was soon after named “T-type” [110, 112]. The unique voltage sensitivity of T-type Ca2+ channels is particularly well suited to regulate neuronal excitability and their oscillatory behavior near the resting membrane potential. A transient membrane hyperpolarization arising from inhibitory post-synaptic potentials (IPSPs) or activation of potassium (K+) channels deinactivates T-type channels. A subsequent rebound in the membrane potential triggers opening of T-type channels and favors a low-threshold spike (LTS) that initiates rebound burst firing (Fig. 1a). The role of T-type channels in bursting behavior is physiologically relevant, especially in sleep [5, 84] with the generation of sleep spindles. In the last two decades, following the molecular cloning of the Cav3 (T-type) channels in the 2000s, genetic studies have greatly contributed to elucidate the role of T-type channels in normal physiology, as well as to identify their implication in diseases. Notably, mutations in the genes encoding the Cav3 channels have been linked essentially to neurodevelopmental, neurological, and psychiatric diseases designated here as neuronal Cav3 channelopathies.

Electrophysiological properties of T-type/Ca3 channels. a Illustration of the implication of Cav3.1 channels in rebound burst firing in thalamocortical relay neurons, as reported in [81]. Hyperpolarization deinactivates T-type channels, which promotes low-threshold spike and rebound burst firing. This firing activity in completely lost in thalamocortical relay neurons from Cav3.1−/− mice (for details, see [81]). b Current traces for Cav3.1, Cav3.2, and Cav3.3 channels obtained in HEK-293 cells, illustrating their differences in inactivation kinetics (see [13, 28]). c Illustration of the Cav3 window current that occurs in the range of the resting membrane potential
Cav3 channels: from molecular to physiological diversity
Cav3 molecular diversity
Before the cloning era, a diversity within T-type channels was already proposed, based on observed differences in inactivation properties and sensitivity to blockers, such as nickel (Ni2+) ions [74]. The first cDNA coding for the pore-forming subunit of a T-type channel was cloned in 1998 by Perez-Reyes and colleagues [119]. This was followed by extensive molecular cloning of several paralogs (isoforms) and orthologs, mainly in vetebrates, leading to the actual landscape of three genes (CACNA1G, CACNA1H, and CACNA1I) encoding the α1 subunit of T-type channels, Cav3.1 (α1G), Cav3.2 (α1H), and Cav3.3 (α1I), respectively (Figs. 1, 2, and 3) (for representative reviews, see [117, 118, 158]). The distinctive features of T-type channels making them well suited to regulate excitability (low voltage range for activation, ion selectivity, fast kinetics for activation and inactivation) are conserved in the most early-diverging animals, such as in Trichoplax adhaerens, which expresses a single Cav3 channel [135]. Hence, genome survey in Salpingoeca rosetta indicates that Cav3 channels have emerged more than a billion years ago in an eukaryotic ancestor of choanoflagellates and metazoans [103].

Schematic representation of the main Cav3 regulations (for previous reviews, see [29, 73, 75, 170]). The yellow asterisks point to the Cav3.2-selective regulations, including the metal/redox (His191, yellow circle) and glycosylation (Asn192, red circle) sites in S3–S4 extracellular linker of the domain I

Cladogram representation of the Cav channel family including the gene names and the corresponding Cav subunits. HVA stands for high-voltage activated channels (L-, P/Q-, N-, and R-types) and LVA stands for low-voltage activated channels (T-type). The channelopathies column refers to the overall so-called Ca2+ channelopathies, with the detailed properties of the Cav3 channelopathies presented and discussed in the text. The diseases caused by mutations in the S6 segments of the corresponding Cav channels are indicated (#)
In mammals, the functional diversity in T-type channels arises not only from the three genes expressing Cav3 isoforms with distinct electrophysiological properties [13, 28] but also from several alternative splicing events [56, 98, 99, 118]. Alternative splicing can generate multiple variants from a single Cav3 isoform with significantly distinct electrophysiological properties and drug sensitivity [25, 26, 54, 83, 101, 105, 132, 172]. Also, alternative splicing can regulate the Cav3 channel expression at the plasma membrane [133]. Alternative splicing could contribute to the clinical severity of Cav3 channelopathies, as documented by in vitro studies showing that disease-associated mutations exhibit distinct electrophysiological properties when reproduced in different splice variants [66, 122].
The tissue-specific expression of the Cav3 channels is clearly important to consider when investigating their physiological roles, as well as their implication in disease phenotypes [131]. In mammals, all Cav3 channels are expressed early during development. In adult, the three Cav3 isoforms are expressed mainly in the central and peripheral nervous systems and also in neuroendocrine and cardiac tissues [101, 102]. Within the brain, in situ hybridization studies have shown that the three Cav3 isoforms display both specific and distinct patterns of expression [12, 144]. In addition, Cav3 splice variants can be expressed in a tissue/cell-specific manner and be developmentally regulated [118]. Until now, the lack of highly specific antibodies for any of the Cav3 isoforms/variants has hampered precise analysis of their tissue and cellular and subcellular distribution at the protein level [1, 100, 166], which was partly circumvented by the generation of knock-in (KI) animals carrying epitope-tagged Cav3 channels [8, 58].
Cav3 physiology
A hallmark of Cav3 channels is their unique ability to control neuronal excitability, requiring small membrane depolarizations to open (LVA), which distinguishes them from the high-voltage activated (HVA) channels [108, 168]. Their low threshold of voltage activation, coupled with their tonic inactivation near resting membrane potential, allows Cav3 channels to deinactivate and to underly the low-threshold spike/rebound bursting phenomenon seen in many types of neurons (Fig. 1a). The three Cav3 isoforms, which exhibit distinct electrophysiological properties [13, 28] (Fig. 1b), regulate differentially neuronal excitability [12, 39, 100]. In addition, the Ca2+ influx through Cav3 channels can also directly regulate intracellular Ca2+ concentrations [24, 51]. Indeed, all three Cav3 channels display an overlap of their steady-state inactivation and activation properties giving rise to a window current (Fig. 1c) that ressembles a “background” Ca2+ current [153]. It results from the activity of a small fraction of Cav3 channels remaining open in the voltage range near the resting membrane potential [34, 40]. The physiological role of this Cav3 window current is still poorly understood. It was shown to contribute to the slow oscillation in non-REM sleep [46].
Genetic manipulation of Cav3 expression in the mouse has provided significant information regarding the physiological roles of neuronal Cav3 channels and a quick overview of the findings obtained with Cav3 knock-out (KO) mouse models is provided here. In KO mice for Cacna1g (Cav3.1−/−), no LVA T-type current could be recorded in thalamocortical relay neurons and these neurons showed no burst firing activity [81] (Fig. 1a). In these animals, spike-and-wave discharges that occur in absence epilepsy models were prevented. The loss of thalamocortical oscillations was also observed in central medial nucleus, which reflects the overall importance of Cav3.1 channels in thalamic neurons [146]. Cav3.1−/− mice were less prone to develop tonic seizures in the maximal electroshock seizure test, compared with wt littermates and Cav3.2−/− mice, suggesting a prominent role of the Cav3.1 isoform in mediating tonic seizure [127]. Interestingly, overexpression of the Cav3.1 channel in a Cacna1g transgenic mouse line results in a pure absence epilepsy phenotype with no ataxia or other neurological disturbances [57], suggesting that an increase in Cav3.1 current is sufficient to the pathogenesis of spike-wave seizures. Cav3.1−/− animals display a deficit in motor performance and in cerebellar learning [23, 94] and are resistant to harmaline-induced tremor [115]. In these animals, the T-type current was also significantly reduced in the subiculum, which is involved in hippocampal-dependent cognitive processes [79].
The KO mice for Cacna1h (Cav3.2−/−) exhibit a variety of phenotypes including neurological deficits [36]. The nociceptive role of Cav3.2 channels, which are highly expressed in the dorsal root sensory neurons [10, 11, 136], was validated using these Cav3.2 KO animals [41, 147] and further established when Cav3.2 channels were selectively deleted in low-threshold mechanoreceptor primary afferent neurons [58]. In the brain, Cav3.2 is predominantly expressed in the dentate gyrus of the hippocampus [1, 8]. In Cav3.2−/− animals subjected to pilocarpine-induced status epilepticus, which models temporal lobe epilepsy, the appearance of epileptic seizures was strongly attenuated, validating the pro-epileptogenesis role of upregulated hippocampal Cav3.2 channels [7, 151]. In addition, Cav3.2−/− mice show elevated anxiety and impaired hippocampus-dependent contextual memory and learning [38, 59].
Inactivation of Cacna1i in the mouse (Cav3.3−/−) resulted in the loss of LVA Ca2+ currents in the thalamic reticular nucleus (nRT) neurons and revealed a role of Cav3.3 channels in sleep. Cav3.3 channels dominate nRT rhythmogenesis and play a role in sleep spindles, the electroencephalographic hallmark of non-rapid eye movement (NREM) sleep [5, 85]. Of note, experiments performed in double Cav3.3 and Cav3.2 KO mice revealed that the lack of Cav3.2 channels further aggravates neuronal, synaptic, and EEG deficits in the Cav3.3−/− background, indicating a role of Cav3.2 channels in regulating nRT excitability and rhythmogenesis [116, 145].
Cav3 modulation
There is no evidence for a requirement of protein-protein association to obtain “native-like” properties of T-type channels when the Cav3 proteins are expressed in heterologous systems, as the HEK-293 cell line. This is contrasting with HVA Ca2+ channels, the L-type Cav1.1 to Cav1.4, and neuronal Cav2.1 P/Q-type, Cav2.2 N-type and Cav2.3 R-type, which require the auxiliary α2/δ, β, and γ subunits for their proper expression and function (reviewed in [108, 168]). However, a regulatory role of several proteins was identified for Cav3 channels, including the HVA auxiliary subunits [52], Kelch-like 1 [3], Stac1 [126], or the putative “Ca2+ channel and chemotaxis receptor domain containing 1,” CACHD1 [43] (Fig. 2). Regulation of Cav3 channels by such endogenous proteins would more likely reflect the numerous signalling pathways targeting Cav3 channels, as reported for the G protein βγ-dimer [50, 162], calmodulin [33, 86], syntaxin-1A [159], and spectrin α/β and ankyrin B [61] (Fig. 2).
The fine tuning of the functional properties of Cav3 channels by a large variety of endogenous pathways and ligands is now well established [29, 73, 75, 170] (Fig. 2). One of the first endogenous modulations described for Cav3 channels was the inhibitory effect of the endocannabinoid anandamide [27]. Other bioactive lipids, including arachidonic acid [143], N-acyl ethanolamides and polyunsaturated fatty acids [31], or 5,6-EET [21], were shown to also inhibit Cav3 channels in the micromolar range through a direct interaction [32]. Cav3 channels are also modulated by phosphorylation pathways, including the serine/threonine kinases, PKA and PKC [30, 71], Ca2+/CaM-dependent protein kinase II (CaMKII) [4, 161], rho-associated kinase (ROCK) [76], CDK5 [63], exchange factor activated by cAMP (Epac) [111], and hypoxia-inducible factor (HIF) [18]. As demonstrated for Cav3.2 channels, the phosphorylation status greatly influences the gating properties [9]. Phosphorylation also regulates an activity-dependent Ca2+ inhibition recently discovered for Cav3 channels, especially Cav3.3 [22, 35]. Other post-translational modifications regulating Cav3 channels, here Cav3.2 channels, including ubiquitination [60] and glycosylation [114, 160] have also been described. These latter studies point out that Cav3 isoform-specific modulations exist and are important to investigate further as they represent physiologically relevant selective regulations (Fig. 2). Of interest, the metal/redox modulation of T-type channels is also Cav3.2-specific. Cav3.2 channels are selectively upregulated by reducing agents such as l-cysteine, while the oxidizing agent ascorbate produces Cav3.2 channel inhibition [148]. This redox regulation occurs through the metal-catalyzed oxidation of a histidine residue (His191 in the human isoform) localized in the extracellular S3–S4 linker of domain I of Cav3.2 channel [72, 80]. Cav3.2 channels are also preferentially inhibited by the trace metal zinc (Zn2+) with an IC50 in the submicromolar range (∼ 0.8 μM), which is 100- and 200-fold lower than Cav3.1 and Cav3.3 channels, respectively [149]. Further studies, using a KI mouse model, have demonstrated that His191 is important for fine tuning of neuronal excitability in dorsal root sensory neurons [156].
Cav3 channelopathies
CACNA1G/Cav3.1 in late-onset cerebellar ataxia ADCA/SCA42
The Cav3.1 channel is highly expressed in the cerebellum, especially in Purkinje neurons [144], and was therefore a likely candidate for cerebellar disorders, especially ataxia. Hereditary cerebellar ataxias are rare neurodegenerative disorders, characterized by a cerebellar syndrome (gait alteration, limb incoordination, dysarthria, eye movement anomalies) with or without other neurological symptoms [45]. Using linkage analysis and whole-exome sequencing, CACNA1G was linked to an autosomal dominant cerebellar ataxia (ADCA) phenotype in three families, supporting its implication in spinocerebellar ataxia SCA42 [44] (Fig. 3). A recurrent missense mutation causing the p.Arg1715His substitution in the voltage sensor S4 segment of domain IV (IVS4, Fig. 4) of Cav3.1 was identified in these three unrelated pedigrees. This p.Arg1715His mutation affects the gating properties of the Cav3.1 channel with the steady-state activation properties shifted positively when expressed in HEK-293 cells. The expected reduction in channel activity was confirmed using computer modeling in deep cerebellar nuclei neurons that showed a decreased neuronal excitability. SCA42 is characterized by a slowly progressive ataxia with a variable onset but mainly in young adulthood. Athough the prevalence of SCA42 is very low, the association of this p.Arg1715His-Cav3.1 mutation with SCA42 was subsequently confirmed in Japanese and Chinese families [82, 88, 104, 109]. Additional CACNA1G missense mutations have been identified in other SCA42 patients, including p.Arg1068Cys, p.His1611Gln, and p.Pro2273His variants. However, they have shown no statistically significant electrophysiological effect in heterologous expression systems [44], while the variant p.Met1574Lys [88] has not been yet electrophysiologically explored. These additional CACNA1G variants clearly require further functional analysis to validate them as SCA42-causative mutations. Recently a KI mouse model of SCA42 was generated [65], harboring the above described mutation (p.Arg1723His in the mouse). Both heterozygous and homozygous KI mice demonstrated an adult-onset mild ataxia phenotype with comparable levels of motor impairment using rotarod and footprint tests, confirming the dominant inheritance of SCA42. Significant Purkinje neuron loss and degeneration of the molecular layer were also observed. This mouse model of SCA42 recapitulates well the observations made in SCA42 patients, as well as the electrophysiological analyses showing a positive shift of the voltage dependence of Cav3.1 channels [44]. Overall, this study confirms that SCA42 is caused by the p.Arg1715His mutation in Cav3.1 [65].

Schematic localization of the main Cav3 mutations described in the text, including (i) the Cav3.1 mutations: p.Arg1715His in SCA42 [44, 65], p.Arg961Thr and p.Met1531Val in ChCA [6, 34]; (ii) the Cav3.2 mutations: p.Arg1549Val and p.Arg1549Ile in PA/FH4 [47, 130], and p.Cys456Ser in CAE [53, 155]; and (iii) the Cav3.3 mutation: p.Arg1346His in SCZ [2, 62]
CACNA1G/Cav3.1 in childhood cerebellar atrophy
Another set of de novo CACNA1G mutations was identified in a cohort of infants with childhood cerebellar atrophy (ChCA), using a combination of candidate gene panel and whole-exome sequencing [34]. ChCA is a devastating infantile neurodevelopmental disorder, with severe motor and cognitive impairments, cerebellar atrophy, and variable features including facial dysmorphism, digital anomalies, microcephaly, hirsutism, and epilepsy. Two mutations were identified in four individuals, three patients carrying a p.Ala961Thr mutation (in the Cav3.1 IIS6 segment) and one patient carrying a p.Met1531Val mutation (in the Cav3.1 IIIS6 segment) [34] (Fig. 4). Both mutations drastically altered the channel gating properties, especially the inactivation properties, with a significant slowing of the inactivation kinetics (5 times) and a negative shift (− 10 mV) of the potential for half-inactivation. In addition, these two mutations promoted a much larger window current that was fully inhibited by TTA-P2, a selective Cav3 channel blocker [34]. Overall, this study has demonstrated that p.Ala961Thr and p.Met1531Val are gain-of-function mutations. Importantly, this first description of de novo dominant CACNA1G mutations causing ChCA was confirmed by a recent study reporting on the same mutations (p.Ala961Thr and p.Met1531Val), identified in three patients and in one patient, respectively [6], strengthening the recurrence of these mutations in ChCA. The devastating consequence of ChCA gain-of-function mutations in humans reveals that Cav3.1 channel has a critical role in setting up cerebellar physiology during development. Further studies are necessary to uncover the pathogenic mechanism underlying the ChCA condition and to establish more precisely the developmental and functional roles of Cav3.1 channels in the cerebellum.
CACNA1G/Cav3.1 in epilepsy
Because of their presence in cortical and thalamic structures and their role in modulating neuronal firing, T-type channels have always been considered candidates for idiopathic generalized epilepsies (IGEs). Interestingly, it was shown that Cacna1g is a genetic modifier of epilepsy in a mouse model of Dravet syndrome caused by mutations in the voltage-gated Na+ channel gene Scn1a [16], as well as a modifier in a Scn2a mouse model of focal epilepsy [15]. In humans, mutations in CACNA1G have been reported in juvenile myoclonic epilepsy patients [134]. However, the two reported missense mutations coding for p.Ala570Val and p.Ala1089Ser substitutions had no electrophysiological effect when explored in heterologous expression system, questioning their pathogenic status on a monogenic basis. A recent study has documented that IGEs have complex (oligogenic or multigenic) inheritance patterns with a likely combination of both common and rare genetic risk variants required to cause the disease. Among them, CACNA1G, carrying a high number of missense variants in IGEs samples, still represents a susceptibility gene [55].
CACNA1G/Cav3.1 in other neurological diseases
Potential disease-causing variants in CACNA1G have also been identified in intellectual disability/cognitive disorders [106] and monoallelic deletions of the CACNA1G gene have been associated with mild intellectual disability without cerebellum atrophy [123]. Also, CACNA1G was identified as a candidate gene for autism spectrum disorder (ASD) in a subset of cases [142] but the CACNA1G association with ASD has yet to be replicated in a larger study [48]. CACNA1G also appeared to be a candidate gene in essential tremor, one of the most common movement disorders, with CACNA1G variants identified in three families [113]. It is therefore likely that the clinical spectrum of diseases associated with CACNA1G mutations will increase in a near future.
CACNA1H/Cav3.2 in epilepsy
The CACNA1H gene, as CACNA1G, has received much attention regarding its potential implication in inherited epilepsy phenotypes. In some spontaneous mouse models of generalized epilepsy, the tottering (tg), lethargic (lh), and stargazer (stg) mouse strains, an increase in T-type current density was observed [169]. Strikingly, in the GAERS rat (Genetic Absence Epilepsy Rats from Strasbourg), a missense mutation, p.Arg1584Pro in Cacna1h, was found to co-segregate with the slow-wave discharge phenotype [150]. In heterologous expression system, this missense substitution in the intracellular loop linking the domains III to IV (LIII–IV) could induce a gain of channel activity when introduced in a Cav3.2 splice variant containing exon 25 [14, 122].
In humans, several studies have reported associations between CACNA1H single nucleotide polymorphisms (SNPs) and epileptic phenotypes, especially in idiopathic generalized epilepsy (IGE) [42, 69, 87, 138], reviewed in [158, 167], since the first report by Chen et al. [37] describing CACNA1H SNPs in childhood absence epilepsy (CAE) patients. Functional studies of several of these CACNA1H missense variants revealed that they could modify biophysical properties or protein trafficking of Cav3.2 in heterologous expression systems [69, 154, 155], in a loss- or gain-of-function manner. Many of these missense variants were found in the intracellular loop linking the domains I and II (LI–II) of Cav3.2 and one of them, p.Cys456Ser (Fig. 4), significantly increased spontaneous firing and reduced the threshold for rebound burst firing, when overexpressed in hippocampal neurons [53]. Yet, to date, none of these CACNA1H variants has been undoubtedly identified as causing seizure phenotypes. In other words, CACNA1H variants are not causing monogenic epilepsy [17]. As for CACNA1G, CACNA1H variants should be considered a risk factor for developing epilepsy, most likely implicating other genetic and/or environmental factors [67].
CACNA1H/Cav3.2 in other neurological diseases
Four missense variants in the CACNA1H gene were identified in six individuals with ASD and the corresponding Cav3.2 variants showed altered electrophysiological properties in heterologous expression [140]. Whether these CACNA1H variants segregate with the ASD phenotype remains to be validated as these variations have low penetrance and some of them were also found in unaffected individuals. Additional CACNA1H variants were reported in a patient with persistent pain [137] and in patients with amyotrophy lateral sclerosis [125, 141]. Again, further studies are needed to validate association of CACNA1H variation with these conditions.
CACNA1H/Cav3.2 in primary aldosteronism
Using whole-exome sequencing, Scholl et al. [130] identified a recurrent missense mutation, p.Met1549Val, in the CACNA1H gene in five unrelated patients from a cohort of patients diagnosed with primary aldosteronism (PA) in early childhood (Fig. 3). This point mutation resulted in a significant gain of Cav3.2 channel activity. Soon after, Daniil et al. [47] also performing whole-exome sequencing in PA patients reported another substitution at this residue, p.Met1549Val, with similar gain-of-function properties. Additional gain-of-function mutations, p.Ser196Leu, p.Pro2083Leu, and p.Val1951Glu, were also identified in this study [47]. If one patient was diagnosed with minor mental retardation and multiplex developmental disorder, other patients showed no apparent signs of seizures, cardiac arrhythmia, or muscular or neurological alterations. CACNA1H-related PA is now defined as familial hyperaldosteronism type 4 (FH4; [120]).
The Met1549 amino acid is located in highly conserved sequence of the IIIS6 segment of Cav3.2 that lines the inner part of the channel pore and is involved in channel inactivation [97] (Fig. 4), indeed at the same position as Met1531 in Cav3.1 (Fig. 3). The two pathogenic substitutions of Met1549 (Val and Ile) confer ultraslow inactivation kinetics, significant negative shift in the steady-state inactivation properties, and an increased window current. These data support an increase in channel activity and a rise in intracellular Ca2+ [47, 130]. Notably, all these mutants led to increased aldosterone production and increased expression of the genes coding for steroidogenic enzymes in the adrenocortical H295R cell line after K+ stimulation [47, 124]. Additional CACNA1H mutations causing PA/FH4 will likely be identified, as exemplified by the recent description of a p.Ile1430Thr substitution (IIIS5 segment), in an aldosterone-producing adenoma [107].
CACNA1I/Cav3.3 in neurological/psychiatric diseases
Genome-wide association studies (GWAS), as well as the identification of de novo variants in the CACNA1I gene, have contributed to implicate CACNA1I as a genetic risk factor in schizophrenia (SCZ) [64, 77, 129]. When expressed in the HEK-293 cell line, one of the two Cav3.3 missense variations identified in [64], p.Arg1346His (Fig. 4), resulted in a lower expression level of the Cav3.3 protein, a reduced N-glycosylation, and a reduced expression at the plasma membrane, reducing the Cav3.3 current but with no change in the electrophysiological properties [2]. A KI mouse model was generated using the CRISPR/Cas9 editing approach to introduce the p.Arg1305His orthologous mutation [62]. The homozygous animals show altered excitability in the nRT and deficits in sleep spindle occurrence and at NREM/REM transitions. This animal model will facilitate further investigations of the role of Cav3.3 channels in impaired sleep spindle and nRT function in SCZ. Additional CACNA1I variants have been identified in SCZ patients, confirming CACNA1I as a genetic risk factor in SCZ [95, 163, 164]. CACNA1I is also considered a risk gene in autism [93] and other complex neuropsychiatric disorders [128].
Conclusions and perspectives
De novo gain-of-function mutations in Cav3 channels: a wider group of S6-pathies?
These last years, many novel disease-related Cav3 channel variants have been reported and some of them are causing severe disorders. This is the case for the de novo gain-of-function mutations in Cav3.1 and Cav3.2 channels in childhood cerebellar atrophy (ChCA) and primary aldosteronism (PA/FH4), respectively. These deleterious missense mutations involve residues, Ala961 and Met1531 in Cav3.1 and Met1549 in Cav3.2, in the highly conserved S6 segments lining the inner part of the pore channel (Fig. 4). These residues were implicated in Cav3 channel inactivation in earlier structure-function studies [49, 97]. Notably, these “S6 mutations” in Cav3.1 and Cav3.2 are reminiscent to several de novo gain-of-function mutations recently described in other genes encoding Cav channels (Fig. 3). These other Cav “S6 mutations” also cause severe, mainly neurodevelopmental, clinical phenotypes. S6 mutations in CACNA1C, which encodes the Cav1.2 L-type channel, cause Timothy syndrome (TS), a congenital long-QT cardiac arrhythmia with or without severe neurological phenotypes, including autism and mental retardation [89, 139]. There are S6 mutations in CACNA1D, which encodes Cav1.3, another L-type channel, that causes a neurodevelopmental disorder including ASD, intellectual disability with or without neurological (hypotonia, epilepsy) and endocrine (primary aldosteronism or hyperinsulinemic hypoglycaemia) features (PASNA) [70, 121]. There are also S6 mutations in Cav2.3, the neuronal R-type channel encoded by CACNA1E, that causes developmental and epileptic encephalopathies (DEE) [68]. There is also recent evidence for de novo S6 mutations in CACNA1A, encoding the neuronal P/Q-type Cav2.1 channel, linked to severe DEE with intellectual disability and variable motor symptoms [78]. All these S6 missense mutations share functional features: they significantly impair the inactivation properties of the affected Cav channels, likely promoting increase in intracellular Ca2+ concentration and the subsequent cellular damages caused by abnormal Ca2+ homeostasis [96]. Considering the similarity in their pathogenic mechanism, we tentatively propose here to define this group of Ca2+ channelopathies as “S6-pathies.” Further studies, exploiting animal models of the corresponding channelopathies, will help to identify the pathogenic mechanisms underlying the diseases and better delineate the precise implication(s) of the corresponding Cav channels. Deciphering the role of Cav3.1 and Cav3.2 in ChCA and PA/FH4, respectively, should benefit from a combined effort of the “calcium channelopathy” community.
Structural studies of Cav3 channels: further deciphering of the disease mechanisms
High-resolution structural studies can provide atomic-level views of disease mechanisms [20]. Notably, the Cryo-EM structure of the Cav3.1 channel was recently reported [171], opening new opportunities to better understand the molecular and functional consequences of disease mutations in Cav3.1 channel, as well as in Cav3.2 and Cav3.3 channels by homology modeling. It is also anticipated that the pharmacology of Cav3 channels will benefit from the development of novel therapeutic approaches using structure-guided drug discovery. Cav3/T-type channels have always been considered promising pharmacological targets considering their implication in a wide variety of neurological conditions, including epilepsy and pain. However, until now, the clinical development of drugs targeting Cav3 channels has not been as successful as expected [92, 157], likely because of the wide tissue expression of the Cav3 channels, the lack of selective Cav3 channel blockers, and, beyond that, the lack of Cav3 isoform-specific blockers. Further studies should establish whether there is a therapeutic potential of Cav3 blockers in the treatment of ChCA and PA/FH4 diseases that are directly caused by increased activity of the Cav3.1 and Cav3.2 channels.
References
Aguado C, Garcia-Madrona S, Gil-Minguez M, Lujan R (2016) Ontogenic changes and differential localization of T-type Ca(2+) channel subunits Cav3.1 and Cav3.2 in mouse hippocampus and cerebellum. Front Neuroanat 10:83. https://doi.org/10.3389/fnana.2016.00083
Andrade A, Hope J, Allen A, Yorgan V, Lipscombe D, Pan JQ (2016) A rare schizophrenia risk variant of CACNA1I disrupts CaV3.3 channel activity. Sci Rep 6:34233. https://doi.org/10.1038/srep34233
Aromolaran KA, Benzow KA, Cribbs LL, Koob MD, Piedras-Renteria ES (2010) T-type current modulation by the actin-binding protein Kelch-like 1. Am J Physiol Cell Physiol 298:C1353–C1362. https://doi.org/10.1152/ajpcell.00235.2009
Asmara H, Micu I, Rizwan AP, Sahu G, Simms BA, Zhang FX, Engbers JDT, Stys PK, Zamponi GW, Turner RW (2017) A T-type channel-calmodulin complex triggers alphaCaMKII activation. Mol Brain 10:37. https://doi.org/10.1186/s13041-017-0317-8
Astori S, Wimmer RD, Prosser HM, Corti C, Corsi M, Liaudet N, Volterra A, Franken P, Adelman JP, Luthi A (2011) The Ca(V)3.3 calcium channel is the major sleep spindle pacemaker in thalamus. Proc Natl Acad Sci U S A 108:13823–13828. https://doi.org/10.1073/pnas.1105115108
Barresi S, Dentici ML, Manzoni F, Bellacchio E, Agolini E, Pizzi S, Ciolfi A, Tarnopolsky M, Brady L, Garone G, Novelli A, Mei D, Guerrini R, Capuano A, Pantaleoni C, Tartaglia M (2020) Infantile-onset syndromic cerebellar ataxia and CACNA1G mutations. Pediatr Neurol 104:40–45. https://doi.org/10.1016/j.pediatrneurol.2019.09.005
Becker AJ, Pitsch J, Sochivko D, Opitz T, Staniek M, Chen CC, Campbell KP, Schoch S, Yaari Y, Beck H (2008) Transcriptional upregulation of Cav3.2 mediates epileptogenesis in the pilocarpine model of epilepsy. J Neurosci 28:13341–13353. https://doi.org/10.1523/JNEUROSCI.1421-08.2008
Bernal Sierra YA, Haseleu J, Kozlenkov A, Begay V, Lewin GR (2017) Genetic tracing of Cav3.2 T-type calcium channel expression in the peripheral nervous system. Front Mol Neurosci 10:70. https://doi.org/10.3389/fnmol.2017.00070
Blesneac I, Chemin J, Bidaud I, Huc-Brandt S, Vandermoere F, Lory P (2015) Phosphorylation of the Cav3.2 T-type calcium channel directly regulates its gating properties. Proc Natl Acad Sci U S A 112:13705–13710. https://doi.org/10.1073/pnas.1511740112
Bourinet E, Alloui A, Monteil A, Barrere C, Couette B, Poirot O, Pages A, McRory J, Snutch TP, Eschalier A, Nargeot J (2005) Silencing of the Cav3.2 T-type calcium channel gene in sensory neurons demonstrates its major role in nociception. EMBO J 24:315–324. https://doi.org/10.1038/sj.emboj.7600515
Bourinet E, Francois A, Laffray S (2016) T-type calcium channels in neuropathic pain. Pain 157(Suppl 1):S15–S22. https://doi.org/10.1097/j.pain.0000000000000469
Broicher T, Kanyshkova T, Meuth P, Pape HC, Budde T (2008) Correlation of T-channel coding gene expression, IT, and the low threshold Ca2+ spike in the thalamus of a rat model of absence epilepsy. Mol Cell Neurosci 39:384–399. https://doi.org/10.1016/j.mcn.2008.07.012
Cain SM, Snutch TP (2010) Contributions of T-type calcium channel isoforms to neuronal firing. Channels (Austin) 4:475–482. https://doi.org/10.4161/chan.4.6.14106
Cain SM, Tyson JR, Choi HB, Ko R, Lin PJC, LeDue JM, Powell KL, Bernier LP, Rungta RL, Yang Y, Cullis PR, O’Brien TJ, MacVicar BA, Snutch TP (2018) CaV 3.2 drives sustained burst-firing, which is critical for absence seizure propagation in reticular thalamic neurons. Epilepsia 59:778–791. https://doi.org/10.1111/epi.14018
Calhoun JD, Hawkins NA, Zachwieja NJ, Kearney JA (2016) Cacna1g is a genetic modifier of epilepsy caused by mutation of voltage-gated sodium channel Scn2a. Epilepsia 57:e103–e107. https://doi.org/10.1111/epi.13390
Calhoun JD, Hawkins NA, Zachwieja NJ, Kearney JA (2017) Cacna1g is a genetic modifier of epilepsy in a mouse model of Dravet syndrome. Epilepsia 58:e111–e115. https://doi.org/10.1111/epi.13811
Calhoun JD, Huffman AM, Bellinski I, Kinsley L, Bachman E, Gerard E, Kearney JA, Carvill GL (2020) CACNA1H variants are not a cause of monogenic epilepsy. Hum Mutat 41:1138–1144. https://doi.org/10.1002/humu.24017
Carabelli V, Marcantoni A, Comunanza V, de Luca A, Diaz J, Borges R, Carbone E (2007) Chronic hypoxia up-regulates alpha1H T-type channels and low-threshold catecholamine secretion in rat chromaffin cells. J Physiol 584:149–165. https://doi.org/10.1113/jphysiol.2007.132274
Carbone E, Lux HD (1984) A low voltage-activated, fully inactivating Ca channel in vertebrate sensory neurones. Nature 310:501–502. https://doi.org/10.1038/310501a0
Catterall WA, Lenaeus MJ, Gamal El-Din TM (2020) Structure and pharmacology of voltage-gated sodium and calcium channels. Annu Rev Pharmacol Toxicol 60:133–154. https://doi.org/10.1146/annurev-pharmtox-010818-021757
Cazade M, Bidaud I, Hansen PB, Lory P, Chemin J (2014) 5,6-EET potently inhibits T-type calcium channels: implication in the regulation of the vascular tone. Pflugers Arch 466:1759–1768. https://doi.org/10.1007/s00424-013-1411-0
Cazade M, Bidaud I, Lory P, Chemin J (2017) Activity-dependent regulation of T-type calcium channels by submembrane calcium ions. Elife 6. https://doi.org/10.7554/eLife.22331
Chang KY, Park YG, Park HY, Homanics GE, Kim J, Kim D (2011) Lack of CaV3.1 channels causes severe motor coordination defects and an age-dependent cerebellar atrophy in a genetic model of essential tremor. Biochem Biophys Res Commun 410:19–23. https://doi.org/10.1016/j.bbrc.2011.05.082
Chemin J, Monteil A, Briquaire C, Richard S, Perez-Reyes E, Nargeot J, Lory P (2000) Overexpression of T-type calcium channels in HEK-293 cells increases intracellular calcium without affecting cellular proliferation. FEBS Lett 478:166–172. https://doi.org/10.1016/s0014-5793(00)01832-9
Chemin J, Monteil A, Bourinet E, Nargeot J, Lory P (2001) Alternatively spliced alpha(1G) (Ca(V)3.1) intracellular loops promote specific T-type Ca(2+) channel gating properties. Biophys J 80:1238–1250. https://doi.org/10.1016/S0006-3495(01)76100-0
Chemin J, Monteil A, Dubel S, Nargeot J, Lory P (2001) The alpha1I T-type calcium channel exhibits faster gating properties when overexpressed in neuroblastoma/glioma NG 108-15 cells. Eur J Neurosci 14:1678–1686. https://doi.org/10.1046/j.0953-816x.2001.01796.x
Chemin J, Monteil A, Perez-Reyes E, Nargeot J, Lory P (2001) Direct inhibition of T-type calcium channels by the endogenous cannabinoid anandamide. EMBO J 20:7033–7040. https://doi.org/10.1093/emboj/20.24.7033
Chemin J, Monteil A, Perez-Reyes E, Bourinet E, Nargeot J, Lory P (2002) Specific contribution of human T-type calcium channel isotypes (alpha(1G), alpha(1H) and alpha(1I)) to neuronal excitability. J Physiol 540:3–14. https://doi.org/10.1113/jphysiol.2001.013269
Chemin J, Traboulsie A, Lory P (2006) Molecular pathways underlying the modulation of T-type calcium channels by neurotransmitters and hormones. Cell Calcium 40:121–134. https://doi.org/10.1016/j.ceca.2006.04.015
Chemin J, Mezghrani A, Bidaud I, Dupasquier S, Marger F, Barrere C, Nargeot J, Lory P (2007) Temperature-dependent modulation of CaV3 T-type calcium channels by protein kinases C and A in mammalian cells. J Biol Chem 282:32710–32718. https://doi.org/10.1074/jbc.M702746200
Chemin J, Nargeot J, Lory P (2007) Chemical determinants involved in anandamide-induced inhibition of T-type calcium channels. J Biol Chem 282:2314–2323. https://doi.org/10.1074/jbc.M610033200
Chemin J, Cazade M, Lory P (2014) Modulation of T-type calcium channels by bioactive lipids. Pflugers Arch 466:689–700. https://doi.org/10.1007/s00424-014-1467-5
Chemin J, Taiakina V, Monteil A, Piazza M, Guan W, Stephens RF, Kitmitto A, Pang ZP, Dolphin AC, Perez-Reyes E, Dieckmann T, Guillemette JG, Spafford JD (2017) Calmodulin regulates Cav3 T-type channels at their gating brake. J Biol Chem 292:20010–20031. https://doi.org/10.1074/jbc.M117.807925
Chemin J, Siquier-Pernet K, Nicouleau M, Barcia G, Ahmad A, Medina-Cano D, Hanein S, Altin N, Hubert L, Bole-Feysot C, Fourage C, Nitschke P, Thevenon J, Rio M, Blanc P, Vidal C, Bahi-Buisson N, Desguerre I, Munnich A, Lyonnet S, Boddaert N, Fassi E, Shinawi M, Zimmerman H, Amiel J, Faivre L, Colleaux L, Lory P, Cantagrel V (2018) De novo mutation screening in childhood-onset cerebellar atrophy identifies gain-of-function mutations in the CACNA1G calcium channel gene. Brain 141:1998–2013. https://doi.org/10.1093/brain/awy145
Chemin J, Stamenic TT, Cazade M, Llinares J, Blesneac I, Todorovic SM, Lory P (2019) A novel phospho-modulatory mechanism contributes to the calcium-dependent regulation of T-type Ca(2+) channels. Sci Rep 9:15642. https://doi.org/10.1038/s41598-019-52194-6
Chen CC, Lamping KG, Nuno DW, Barresi R, Prouty SJ, Lavoie JL, Cribbs LL, England SK, Sigmund CD, Weiss RM, Williamson RA, Hill JA, Campbell KP (2003) Abnormal coronary function in mice deficient in alpha1H T-type Ca2+ channels. Science 302:1416–1418. https://doi.org/10.1126/science.1089268
Chen Y, Lu J, Pan H, Zhang Y, Wu H, Xu K, Liu X, Jiang Y, Bao X, Yao Z, Ding K, Lo WH, Qiang B, Chan P, Shen Y, Wu X (2003) Association between genetic variation of CACNA1H and childhood absence epilepsy. Ann Neurol 54:239–243. https://doi.org/10.1002/ana.10607
Chen CC, Shen JW, Chung NC, Min MY, Cheng SJ, Liu IY (2012) Retrieval of context-associated memory is dependent on the Ca(v)3.2 T-type calcium channel. PLoS One 7:e29384. https://doi.org/10.1371/journal.pone.0029384
Cheong E, Shin HS (2014) T-type Ca2+ channels in absence epilepsy. Pflugers Arch 466:719–734. https://doi.org/10.1007/s00424-014-1461-y
Chevalier M, Lory P, Mironneau C, Macrez N, Quignard JF (2006) T-type CaV3.3 calcium channels produce spontaneous low-threshold action potentials and intracellular calcium oscillations. Eur J Neurosci 23:2321–2329. https://doi.org/10.1111/j.1460-9568.2006.04761.x
Choi S, Na HS, Kim J, Lee J, Lee S, Kim D, Park J, Chen CC, Campbell KP, Shin HS (2007) Attenuated pain responses in mice lacking Ca(V)3.2 T-type channels. Genes Brain Behav 6:425–431. https://doi.org/10.1111/j.1601-183X.2006.00268.x
Chourasia N, Osso-Rivera H, Ghosh A, Von Allmen G, Koenig MK (2019) Expanding the phenotypic spectrum of CACNA1H mutations. Pediatr Neurol 93:50–55. https://doi.org/10.1016/j.pediatrneurol.2018.11.017
Cottrell GS, Soubrane CH, Hounshell JA, Lin H, Owenson V, Rigby M, Cox PJ, Barker BS, Ottolini M, Ince S, Bauer CC, Perez-Reyes E, Patel MK, Stevens EB, Stephens GJ (2018) CACHD1 is an alpha2delta-like protein that modulates CaV3 voltage-gated calcium channel activity. J Neurosci 38:9186–9201. https://doi.org/10.1523/JNEUROSCI.3572-15.2018
Coutelier M, Blesneac I, Monteil A, Monin ML, Ando K, Mundwiller E, Brusco A, Le Ber I, Anheim M, Castrioto A, Duyckaerts C, Brice A, Durr A, Lory P, Stevanin G (2015) A recurrent mutation in CACNA1G alters Cav3.1 T-type calcium-channel conduction and causes autosomal-dominant cerebellar ataxia. Am J Hum Genet 97:726–737. https://doi.org/10.1016/j.ajhg.2015.09.007
Coutelier M, Coarelli G, Monin ML, Konop J, Davoine CS, Tesson C, Valter R, Anheim M, Behin A, Castelnovo G, Charles P, David A, Ewenczyk C, Fradin M, Goizet C, Hannequin D, Labauge P, Riant F, Sarda P, Sznajer Y, Tison F, Ullmann U, Van Maldergem L, Mochel F, Brice A, Stevanin G, Durr A, network S (2017) A panel study on patients with dominant cerebellar ataxia highlights the frequency of channelopathies. Brain 140:1579–1594. https://doi.org/10.1093/brain/awx081
Crunelli V, Lorincz ML, Connelly WM, David F, Hughes SW, Lambert RC, Leresche N, Errington AC (2018) Dual function of thalamic low-vigilance state oscillations: rhythm-regulation and plasticity. Nat Rev Neurosci 19:107–118. https://doi.org/10.1038/nrn.2017.151
Daniil G, Fernandes-Rosa FL, Chemin J, Blesneac I, Beltrand J, Polak M, Jeunemaitre X, Boulkroun S, Amar L, Strom TM, Lory P, Zennaro MC (2016) CACNA1H mutations are associated with different forms of primary aldosteronism. EBioMedicine 13:225–236. https://doi.org/10.1016/j.ebiom.2016.10.002
Deciphering Developmental Disorders S (2015) Large-scale discovery of novel genetic causes of developmental disorders. Nature 519:223–228. https://doi.org/10.1038/nature14135
Demers-Giroux PO, Bourdin B, Sauve R, Parent L (2013) Cooperative activation of the T-type CaV3.2 channel: interaction between domains II and III. J Biol Chem 288:29281–29293. https://doi.org/10.1074/jbc.M113.500975
DePuy SD, Yao J, Hu C, McIntire W, Bidaud I, Lory P, Rastinejad F, Gonzalez C, Garrison JC, Barrett PQ (2006) The molecular basis for T-type Ca2+ channel inhibition by G protein beta2gamma2 subunits. Proc Natl Acad Sci U S A 103:14590–14595. https://doi.org/10.1073/pnas.0603945103
Dreyfus FM, Tscherter A, Errington AC, Renger JJ, Shin HS, Uebele VN, Crunelli V, Lambert RC, Leresche N (2010) Selective T-type calcium channel block in thalamic neurons reveals channel redundancy and physiological impact of I(T)window. J Neurosci 30:99–109. https://doi.org/10.1523/JNEUROSCI.4305-09.2010
Dubel SJ, Altier C, Chaumont S, Lory P, Bourinet E, Nargeot J (2004) Plasma membrane expression of T-type calcium channel alpha(1) subunits is modulated by high voltage-activated auxiliary subunits. J Biol Chem 279:29263–29269. https://doi.org/10.1074/jbc.M313450200
Eckle VS, Shcheglovitov A, Vitko I, Dey D, Yap CC, Winckler B, Perez-Reyes E (2014) Mechanisms by which a CACNA1H mutation in epilepsy patients increases seizure susceptibility. J Physiol 592:795–809. https://doi.org/10.1113/jphysiol.2013.264176
Emerick MC, Stein R, Kunze R, McNulty MM, Regan MR, Hanck DA, Agnew WS (2006) Profiling the array of Ca(v)3.1 variants from the human T-type calcium channel gene CACNA1G: alternative structures, developmental expression, and biophysical variations. Proteins 64:320–342. https://doi.org/10.1002/prot.20877
Epi25 Collaborative, Electronic address sbuea, Epi C (2019) Ultra-rare genetic variation in the epilepsies: a whole-exome sequencing study of 17,606 individuals. Am J Hum Genet 105:267–282. https://doi.org/10.1016/j.ajhg.2019.05.020
Ernst WL, Noebels JL (2009) Expanded alternative splice isoform profiling of the mouse Cav3.1/alpha1G T-type calcium channel. BMC Mol Biol 10:53. https://doi.org/10.1186/1471-2199-10-53
Ernst WL, Zhang Y, Yoo JW, Ernst SJ, Noebels JL (2009) Genetic enhancement of thalamocortical network activity by elevating alpha 1g-mediated low-voltage-activated calcium current induces pure absence epilepsy. J Neurosci 29:1615–1625. https://doi.org/10.1523/JNEUROSCI.2081-08.2009
Francois A, Schuetter N, Laffray S, Sanguesa J, Pizzoccaro A, Dubel S, Mantilleri A, Nargeot J, Noel J, Wood JN, Moqrich A, Pongs O, Bourinet E (2015) The low-threshold calcium channel Cav3.2 determines low-threshold mechanoreceptor function. Cell Rep 10:370–382. https://doi.org/10.1016/j.celrep.2014.12.042
Gangarossa G, Laffray S, Bourinet E, Valjent E (2014) T-type calcium channel Cav3.2 deficient mice show elevated anxiety, impaired memory and reduced sensitivity to psychostimulants. Front Behav Neurosci 8:92. https://doi.org/10.3389/fnbeh.2014.00092
Garcia-Caballero A, Gadotti VM, Stemkowski P, Weiss N, Souza IA, Hodgkinson V, Bladen C, Chen L, Hamid J, Pizzoccaro A, Deage M, Francois A, Bourinet E, Zamponi GW (2014) The deubiquitinating enzyme USP5 modulates neuropathic and inflammatory pain by enhancing Cav3.2 channel activity. Neuron 83:1144–1158. https://doi.org/10.1016/j.neuron.2014.07.036
Garcia-Caballero A, Zhang FX, Hodgkinson V, Huang J, Chen L, Souza IA, Cain S, Kass J, Alles S, Snutch TP, Zamponi GW (2018) T-type calcium channels functionally interact with spectrin (alpha/beta) and ankyrin B. Mol Brain 11:24. https://doi.org/10.1186/s13041-018-0368-5
Ghoshal A, Uygun DS, Yang L, McNally JM, Lopez-Huerta VG, Arias-Garcia MA, Baez-Nieto D, Allen A, Fitzgerald M, Choi S, Zhang Q, Hope JM, Yan K, Mao X, Nicholson TB, Imaizumi K, Fu Z, Feng G, Brown RE, Strecker RE, Purcell SM, Pan JQ (2020) Effects of a patient-derived de novo coding alteration of CACNA1I in mice connect a schizophrenia risk gene with sleep spindle deficits. Transl Psychiatry 10:29. https://doi.org/10.1038/s41398-020-0685-1
Gomez K, Calderon-Rivera A, Sandoval A, Gonzalez-Ramirez R, Vargas-Parada A, Ojeda-Alonso J, Granados-Soto V, Delgado-Lezama R, Felix R (2020) Cdk5-dependent phosphorylation of CaV3.2 T-type channels: possible role in nerve ligation-induced neuropathic allodynia and the compound action potential in primary afferent C fibers. J Neurosci 40:283–296. https://doi.org/10.1523/JNEUROSCI.0181-19.2019
Gulsuner S, Walsh T, Watts AC, Lee MK, Thornton AM, Casadei S, Rippey C, Shahin H, Consortium on the Genetics of S, Group PS, Nimgaonkar VL, Go RC, Savage RM, Swerdlow NR, Gur RE, Braff DL, King MC, McClellan JM (2013) Spatial and temporal mapping of de novo mutations in schizophrenia to a fetal prefrontal cortical network. Cell 154:518–529. https://doi.org/10.1016/j.cell.2013.06.049
Hashiguchi S, Doi H, Kunii M, Nakamura Y, Shimuta M, Suzuki E, Koyano S, Okubo M, Kishida H, Shiina M, Ogata K, Hirashima F, Inoue Y, Kubota S, Hayashi N, Nakamura H, Takahashi K, Katsumoto A, Tada M, Tanaka K, Sasaoka T, Miyatake S, Miyake N, Saitsu H, Sato N, Ozaki K, Ohta K, Yokota T, Mizusawa H, Mitsui J, Ishiura H, Yoshimura J, Morishita S, Tsuji S, Takeuchi H, Ishikawa K, Matsumoto N, Ishikawa T, Tanaka F (2019) Ataxic phenotype with altered CaV3.1 channel property in a mouse model for spinocerebellar ataxia 42. Neurobiol Dis 130:104516. https://doi.org/10.1016/j.nbd.2019.104516
He M, Xu Z, Zhang Y, Hu C (2019) Splice-variant-specific effects of primary aldosteronism point mutations on human CaV3.2 calcium channels. Cell Calcium 84:102104. https://doi.org/10.1016/j.ceca.2019.102104
Helbig I, Riggs ER, Barry CA, Klein KM, Dyment D, Thaxton C, Sadikovic B, Sands TT, Wagnon JL, Liaquat K, Cilio MR, Mirzaa G, Park K, Axeen E, Butler E, Bardakjian TM, Striano P, Poduri A, Siegert RK, Grant AR, Helbig KL, Mefford HC (2018) The ClinGen Epilepsy Gene Curation Expert Panel-bridging the divide between clinical domain knowledge and formal gene curation criteria. Hum Mutat 39:1476–1484. https://doi.org/10.1002/humu.23632
Helbig KL, Lauerer RJ, Bahr JC, Souza IA, Myers CT, Uysal B, Schwarz N, Gandini MA, Huang S, Keren B, Mignot C, Afenjar A, Billette de Villemeur T, Heron D, Nava C, Valence S, Buratti J, Fagerberg CR, Soerensen KP, Kibaek M, Kamsteeg EJ, Koolen DA, Gunning B, Schelhaas HJ, Kruer MC, Fox J, Bakhtiari S, Jarrar R, Padilla-Lopez S, Lindstrom K, Jin SC, Zeng X, Bilguvar K, Papavasileiou A, Xing Q, Zhu C, Boysen K, Vairo F, Lanpher BC, Klee EW, Tillema JM, Payne ET, Cousin MA, Kruisselbrink TM, Wick MJ, Baker J, Haan E, Smith N, Sadeghpour A, Davis EE, Katsanis N, Task Force for Neonatal G, Corbett MA, MacLennan AH, Gecz J, Biskup S, Goldmann E, Rodan LH, Kichula E, Segal E, Jackson KE, Asamoah A, Dimmock D, McCarrier J, Botto LD, Filloux F, Tvrdik T, Cascino GD, Klingerman S, Neumann C, Wang R, Jacobsen JC, Nolan MA, Snell RG, Lehnert K, Sadleir LG, Anderlid BM, Kvarnung M, Guerrini R, Friez MJ, Lyons MJ, Leonhard J, Kringlen G, Casas K, El Achkar CM, Smith LA, Rotenberg A, Poduri A, Sanchis-Juan A, Carss KJ, Rankin J, Zeman A, Raymond FL, Blyth M, Kerr B, Ruiz K, Urquhart J, Hughes I, Banka S, Deciphering Developmental Disorders S, UBS H, Scheffer IE, Helbig I, Zamponi GW, Lerche H, Mefford HC (2018) De novo pathogenic variants in CACNA1E cause developmental and epileptic encephalopathy with contractures, macrocephaly, and dyskinesias. Am J Hum Genet 103:666–678. https://doi.org/10.1016/j.ajhg.2018.09.006
Heron SE, Khosravani H, Varela D, Bladen C, Williams TC, Newman MR, Scheffer IE, Berkovic SF, Mulley JC, Zamponi GW (2007) Extended spectrum of idiopathic generalized epilepsies associated with CACNA1H functional variants. Ann Neurol 62:560–568. https://doi.org/10.1002/ana.21169
Hofer NT, Tuluc P, Ortner NJ, Nikonishyna YV, Fernandes-Quintero ML, Liedl KR, Flucher BE, Cox H, Striessnig J (2020) Biophysical classification of a CACNA1D de novo mutation as a high-risk mutation for a severe neurodevelopmental disorder. Mol Autism 11:4. https://doi.org/10.1186/s13229-019-0310-4
Hu C, Depuy SD, Yao J, McIntire WE, Barrett PQ (2009) Protein kinase A activity controls the regulation of T-type CaV3.2 channels by Gbetagamma dimers. J Biol Chem 284:7465–7473. https://doi.org/10.1074/jbc.M808049200
Huang D, Shi S, Liang C, Zhang X, Du X, An H, Peers C, Zhang H, Gamper N (2020) Delineating an extracellular redox-sensitive module in T-type Ca(2+) channels. J Biol Chem 295:6177–6186. https://doi.org/10.1074/jbc.RA120.012668
Huc S, Monteil A, Bidaud I, Barbara G, Chemin J, Lory P (2009) Regulation of T-type calcium channels: signalling pathways and functional implications. Biochim Biophys Acta 1793:947–952. https://doi.org/10.1016/j.bbamcr.2008.11.003
Huguenard JR (1996) Low-threshold calcium currents in central nervous system neurons. Annu Rev Physiol 58:329–348. https://doi.org/10.1146/annurev.ph.58.030196.001553
Iftinca MC, Zamponi GW (2009) Regulation of neuronal T-type calcium channels. Trends Pharmacol Sci 30:32–40. https://doi.org/10.1016/j.tips.2008.10.004
Iftinca M, Hamid J, Chen L, Varela D, Tadayonnejad R, Altier C, Turner RW, Zamponi GW (2007) Regulation of T-type calcium channels by rho-associated kinase. Nat Neurosci 10:854–860. https://doi.org/10.1038/nn1921
Irish Schizophrenia Genomics C, the Wellcome Trust Case Control C (2012) Genome-wide association study implicates HLA-C*01:02 as a risk factor at the major histocompatibility complex locus in schizophrenia. Biol Psychiatry 72:620–628. https://doi.org/10.1016/j.biopsych.2012.05.035
Jiang X, Raju PK, D’Avanzo N, Lachance M, Pepin J, Dubeau F, Mitchell WG, Bello-Espinosa LE, Pierson TM, Minassian BA, Lacaille JC, Rossignol E (2019) Both gain-of-function and loss-of-function de novo CACNA1A mutations cause severe developmental epileptic encephalopathies in the spectrum of Lennox-Gastaut syndrome. Epilepsia 60:1881–1894. https://doi.org/10.1111/epi.16316
Joksimovic SM, Eggan P, Izumi Y, Joksimovic SL, Tesic V, Dietz RM, Orfila JE, DiGruccio MR, Herson PS, Jevtovic-Todorovic V, Zorumski CF, Todorovic SM (2017) The role of T-type calcium channels in the subiculum: to burst or not to burst? J Physiol 595:6327–6348. https://doi.org/10.1113/JP274565
Kang HW, Vitko I, Lee SS, Perez-Reyes E, Lee JH (2010) Structural determinants of the high affinity extracellular zinc binding site on Cav3.2 T-type calcium channels. J Biol Chem 285:3271–3281. https://doi.org/10.1074/jbc.M109.067660
Kim D, Song I, Keum S, Lee T, Jeong MJ, Kim SS, McEnery MW, Shin HS (2001) Lack of the burst firing of thalamocortical relay neurons and resistance to absence seizures in mice lacking alpha(1G) T-type Ca(2+) channels. Neuron 31:35–45. https://doi.org/10.1016/s0896-6273(01)00343-9
Kimura M, Yabe I, Hama Y, Eguchi K, Ura S, Tsuzaka K, Tsuji S, Sasaki H (2017) SCA42 mutation analysis in a case series of Japanese patients with spinocerebellar ataxia. J Hum Genet 62:857–859. https://doi.org/10.1038/jhg.2017.51
Latour I, Louw DF, Beedle AM, Hamid J, Sutherland GR, Zamponi GW (2004) Expression of T-type calcium channel splice variants in human glioma. Glia 48:112–119. https://doi.org/10.1002/glia.20063
Lee J, Kim D, Shin HS (2004) Lack of delta waves and sleep disturbances during non-rapid eye movement sleep in mice lacking alpha1G-subunit of T-type calcium channels. Proc Natl Acad Sci U S A 101:18195–18199. https://doi.org/10.1073/pnas.0408089101
Lee SE, Lee J, Latchoumane C, Lee B, Oh SJ, Saud ZA, Park C, Sun N, Cheong E, Chen CC, Choi EJ, Lee CJ, Shin HS (2014) Rebound burst firing in the reticular thalamus is not essential for pharmacological absence seizures in mice. Proc Natl Acad Sci U S A 111:11828–11833. https://doi.org/10.1073/pnas.1408609111
Lee N, Jeong S, Kim KC, Kim JA, Park JY, Kang HW, Perez-Reyes E, Lee JH (2017) Ca(2+) regulation of Cav3.3 T-type Ca(2+) channel is mediated by calmodulin. Mol Pharmacol 92:347–357. https://doi.org/10.1124/mol.117.108530
Lee CG, Lee J, Lee M (2018) Multi-gene panel testing in Korean patients with common genetic generalized epilepsy syndromes. PLoS One 13:e0199321. https://doi.org/10.1371/journal.pone.0199321
Li X, Zhou C, Cui L, Zhu L, Du H, Liu J, Wang C, Fang S (2018) A case of a novel CACNA1G mutation from a Chinese family with SCA42: a case report and literature review. Medicine (Baltimore) 97:e12148. https://doi.org/10.1097/MD.0000000000012148
Liao P, Soong TW (2010) CaV1.2 channelopathies: from arrhythmias to autism, bipolar disorder, and immunodeficiency. Pflugers Arch 460:353–359. https://doi.org/10.1007/s00424-009-0753-0
Llinas R, Jahnsen H (1982) Electrophysiology of mammalian thalamic neurones in vitro. Nature 297:406–408. https://doi.org/10.1038/297406a0
Llinas R, Yarom Y (1981) Electrophysiology of mammalian inferior olivary neurones in vitro. Different types of voltage-dependent ionic conductances. J Physiol 315:549–567. https://doi.org/10.1113/jphysiol.1981.sp013763
Lory P, Chemin J (2007) Towards the discovery of novel T-type calcium channel blockers. Expert Opin Ther Targets 11:717–722. https://doi.org/10.1517/14728222.11.5.717
Lu AT, Dai X, Martinez-Agosto JA, Cantor RM (2012) Support for calcium channel gene defects in autism spectrum disorders. Mol Autism 3:18. https://doi.org/10.1186/2040-2392-3-18
Ly R, Bouvier G, Schonewille M, Arabo A, Rondi-Reig L, Lena C, Casado M, De Zeeuw CI, Feltz A (2013) T-type channel blockade impairs long-term potentiation at the parallel fiber-Purkinje cell synapse and cerebellar learning. Proc Natl Acad Sci U S A 110:20302–20307. https://doi.org/10.1073/pnas.1311686110
Manoach DS, Pan JQ, Purcell SM, Stickgold R (2016) Reduced sleep spindles in schizophrenia: a treatable endophenotype that links risk genes to impaired cognition? Biol Psychiatry 80:599–608. https://doi.org/10.1016/j.biopsych.2015.10.003
Mark MD, Schwitalla JC, Groemmke M, Herlitze S (2017) Keeping our calcium in balance to maintain our balance. Biochem Biophys Res Commun 483:1040–1050. https://doi.org/10.1016/j.bbrc.2016.07.020
Marksteiner R, Schurr P, Berjukow S, Margreiter E, Perez-Reyes E, Hering S (2001) Inactivation determinants in segment IIIS6 of Ca(v)3.1. J Physiol 537:27–34. https://doi.org/10.1111/j.1469-7793.2001.0027k.x
Mittman S, Guo J, Agnew WS (1999) Structure and alternative splicing of the gene encoding alpha1G, a human brain T calcium channel alpha1 subunit. Neurosci Lett 274:143–146. https://doi.org/10.1016/s0304-3940(99)00716-8
Mittman S, Guo J, Emerick MC, Agnew WS (1999) Structure and alternative splicing of the gene encoding alpha1I, a human brain T calcium channel alpha1 subunit. Neurosci Lett 269:121–124. https://doi.org/10.1016/s0304-3940(99)00319-5
Molineux ML, McRory JE, McKay BE, Hamid J, Mehaffey WH, Rehak R, Snutch TP, Zamponi GW, Turner RW (2006) Specific T-type calcium channel isoforms are associated with distinct burst phenotypes in deep cerebellar nuclear neurons. Proc Natl Acad Sci U S A 103:5555–5560. https://doi.org/10.1073/pnas.0601261103
Monteil A, Chemin J, Bourinet E, Mennessier G, Lory P, Nargeot J (2000) Molecular and functional properties of the human alpha(1G) subunit that forms T-type calcium channels. J Biol Chem 275:6090–6100. https://doi.org/10.1074/jbc.275.9.6090
Monteil A, Chemin J, Leuranguer V, Altier C, Mennessier G, Bourinet E, Lory P, Nargeot J (2000) Specific properties of T-type calcium channels generated by the human alpha 1I subunit. J Biol Chem 275:16530–16535. https://doi.org/10.1074/jbc.C000090200
Moran Y, Zakon HH (2014) The evolution of the four subunits of voltage-gated calcium channels: ancient roots, increasing complexity, and multiple losses. Genome Biol Evol 6:2210–2217. https://doi.org/10.1093/gbe/evu177
Morino H, Matsuda Y, Muguruma K, Miyamoto R, Ohsawa R, Ohtake T, Otobe R, Watanabe M, Maruyama H, Hashimoto K, Kawakami H (2015) A mutation in the low voltage-gated calcium channel CACNA1G alters the physiological properties of the channel, causing spinocerebellar ataxia. Mol Brain 8:89. https://doi.org/10.1186/s13041-015-0180-4
Murbartian J, Arias JM, Lee JH, Gomora JC, Perez-Reyes E (2002) Alternative splicing of the rat Ca(v)3.3 T-type calcium channel gene produces variants with distinct functional properties(1). FEBS Lett 528:272–278. https://doi.org/10.1016/s0014-5793(02)03341-0
Najmabadi H, Hu H, Garshasbi M, Zemojtel T, Abedini SS, Chen W, Hosseini M, Behjati F, Haas S, Jamali P, Zecha A, Mohseni M, Puttmann L, Vahid LN, Jensen C, Moheb LA, Bienek M, Larti F, Mueller I, Weissmann R, Darvish H, Wrogemann K, Hadavi V, Lipkowitz B, Esmaeeli-Nieh S, Wieczorek D, Kariminejad R, Firouzabadi SG, Cohen M, Fattahi Z, Rost I, Mojahedi F, Hertzberg C, Dehghan A, Rajab A, Banavandi MJ, Hoffer J, Falah M, Musante L, Kalscheuer V, Ullmann R, Kuss AW, Tzschach A, Kahrizi K, Ropers HH (2011) Deep sequencing reveals 50 novel genes for recessive cognitive disorders. Nature 478:57–63. https://doi.org/10.1038/nature10423
Nanba K, Blinder AR, Rege J, Hattangady NG, Else T, Liu CJ, Tomlins SA, Vats P, Kumar-Sinha C, Giordano TJ, Rainey WE (2020) Somatic CACNA1H mutation as a cause of aldosterone-producing adenoma. Hypertension 75:645–649. https://doi.org/10.1161/HYPERTENSIONAHA.119.14349
Nanou E, Catterall WA (2018) Calcium channels, synaptic plasticity, and neuropsychiatric disease. Neuron 98:466–481. https://doi.org/10.1016/j.neuron.2018.03.017
Ngo K, Aker M, Petty LE, Chen J, Cavalcanti F, Nelson AB, Hassin-Baer S, Geschwind MD, Perlman S, Italiano D, Lagana A, Cavallaro S, Coppola G, Below JE, Fogel BL (2018) Expanding the global prevalence of spinocerebellar ataxia type 42. Neurol Genet 4:e232. https://doi.org/10.1212/NXG.0000000000000232
Nilius B, Hess P, Lansman JB, Tsien RW (1985) A novel type of cardiac calcium channel in ventricular cells. Nature 316:443–446. https://doi.org/10.1038/316443a0
Novara M, Baldelli P, Cavallari D, Carabelli V, Giancippoli A, Carbone E (2004) Exposure to cAMP and beta-adrenergic stimulation recruits Ca(V)3 T-type channels in rat chromaffin cells through Epac cAMP-receptor proteins. J Physiol 558:433–449. https://doi.org/10.1113/jphysiol.2004.061184
Nowycky MC, Fox AP, Tsien RW (1985) Three types of neuronal calcium channel with different calcium agonist sensitivity. Nature 316:440–443. https://doi.org/10.1038/316440a0
Odgerel Z, Sonti S, Hernandez N, Park J, Ottman R, Louis ED, Clark LN (2019) Whole genome sequencing and rare variant analysis in essential tremor families. PLoS One 14:e0220512. https://doi.org/10.1371/journal.pone.0220512
Orestes P, Osuru HP, McIntire WE, Jacus MO, Salajegheh R, Jagodic MM, Choe W, Lee J, Lee SS, Rose KE, Poiro N, Digruccio MR, Krishnan K, Covey DF, Lee JH, Barrett PQ, Jevtovic-Todorovic V, Todorovic SM (2013) Reversal of neuropathic pain in diabetes by targeting glycosylation of Ca(V)3.2 T-type calcium channels. Diabetes 62:3828–3838. https://doi.org/10.2337/db13-0813
Park YG, Park HY, Lee CJ, Choi S, Jo S, Choi H, Kim YH, Shin HS, Llinas RR, Kim D (2010) Ca(V)3.1 is a tremor rhythm pacemaker in the inferior olive. Proc Natl Acad Sci U S A 107:10731–10736. https://doi.org/10.1073/pnas.1002995107
Pellegrini C, Lecci S, Luthi A, Astori S (2016) Suppression of sleep spindle rhythmogenesis in mice with deletion of CaV3.2 and CaV3.3 T-type Ca(2+) channels. Sleep 39:875–885. https://doi.org/10.5665/sleep.5646
Perez-Reyes E (2003) Molecular physiology of low-voltage-activated t-type calcium channels. Physiol Rev 83:117–161. https://doi.org/10.1152/physrev.00018.2002
Perez-Reyes E, Lory P (2006) Molecular biology of T-type calcium channels. CNS Neurol Disord Drug Targets 5:605–609. https://doi.org/10.2174/187152706779025508
Perez-Reyes E, Cribbs LL, Daud A, Lacerda AE, Barclay J, Williamson MP, Fox M, Rees M, Lee JH (1998) Molecular characterization of a neuronal low-voltage-activated T-type calcium channel. Nature 391:896–900. https://doi.org/10.1038/36110
Perez-Rivas LG, Williams TA, Reincke M (2019) Inherited forms of primary hyperaldosteronism: new genes, new phenotypes and proposition of a new classification. Exp Clin Endocrinol Diabetes 127:93–99. https://doi.org/10.1055/a-0713-0629
Pinggera A, Lieb A, Benedetti B, Lampert M, Monteleone S, Liedl KR, Tuluc P, Striessnig J (2015) CACNA1D de novo mutations in autism spectrum disorders activate Cav1.3 L-type calcium channels. Biol Psychiatry 77:816–822. https://doi.org/10.1016/j.biopsych.2014.11.020
Powell KL, Cain SM, Ng C, Sirdesai S, David LS, Kyi M, Garcia E, Tyson JR, Reid CA, Bahlo M, Foote SJ, Snutch TP, O’Brien TJ (2009) A Cav3.2 T-type calcium channel point mutation has splice-variant-specific effects on function and segregates with seizure expression in a polygenic rat model of absence epilepsy. J Neurosci 29:371–380. https://doi.org/10.1523/JNEUROSCI.5295-08.2009
Preiksaitiene E, Mannik K, Dirse V, Utkus A, Ciuladaite Z, Kasnauskiene J, Kurg A, Kucinskas V (2012) A novel de novo 1.8 Mb microdeletion of 17q21.33 associated with intellectual disability and dysmorphic features. Eur J Med Genet 55:656–659. https://doi.org/10.1016/j.ejmg.2012.07.008
Reimer EN, Walenda G, Seidel E, Scholl UI (2016) CACNA1H(M1549V) mutant calcium channel causes autonomous aldosterone production in HAC15 cells and is inhibited by mibefradil. Endocrinology 157:3016–3022. https://doi.org/10.1210/en.2016-1170
Rzhepetskyy Y, Lazniewska J, Blesneac I, Pamphlett R, Weiss N (2016) CACNA1H missense mutations associated with amyotrophic lateral sclerosis alter Cav3.2 T-type calcium channel activity and reticular thalamic neuron firing. Channels (Austin) 10:466–477. https://doi.org/10.1080/19336950.2016.1204497
Rzhepetskyy Y, Lazniewska J, Proft J, Campiglio M, Flucher BE, Weiss N (2016) A Cav3.2/Stac1 molecular complex controls T-type channel expression at the plasma membrane. Channels (Austin) 10:346–354. https://doi.org/10.1080/19336950.2016.1186318
Sakkaki S, Gangarossa G, Lerat B, Francon D, Forichon L, Chemin J, Valjent E, Lerner-Natoli M, Lory P (2016) Blockade of T-type calcium channels prevents tonic-clonic seizures in a maximal electroshock seizure model. Neuropharmacology 101:320–329. https://doi.org/10.1016/j.neuropharm.2015.09.032
Sanchez-Roige S, Fontanillas P, Elson SL, Gray JC, de Wit H, MacKillop J, Palmer AA (2019) Genome-wide association studies of impulsive personality traits (BIS-11 and UPPS-P) and drug experimentation in up to 22,861 adult research participants identify loci in the CACNA1I and CADM2 genes. J Neurosci 39:2562–2572. https://doi.org/10.1523/JNEUROSCI.2662-18.2019
Schizophrenia Working Group of the Psychiatric Genomics C (2014) Biological insights from 108 schizophrenia-associated genetic loci. Nature 511:421–427. https://doi.org/10.1038/nature13595
Scholl UI, Stolting G, Nelson-Williams C, Vichot AA, Choi M, Loring E, Prasad ML, Goh G, Carling T, Juhlin CC, Quack I, Rump LC, Thiel A, Lande M, Frazier BG, Rasoulpour M, Bowlin DL, Sethna CB, Trachtman H, Fahlke C, Lifton RP (2015) Recurrent gain of function mutation in calcium channel CACNA1H causes early-onset hypertension with primary aldosteronism. Elife 4:e06315. https://doi.org/10.7554/eLife.06315
Seitter H, Koschak A (2018) Relevance of tissue specific subunit expression in channelopathies. Neuropharmacology 132:58–70. https://doi.org/10.1016/j.neuropharm.2017.06.029
Senatore A, Spafford JD (2012) Gene transcription and splicing of T-type channels are evolutionarily-conserved strategies for regulating channel expression and gating. PLoS One 7:e37409. https://doi.org/10.1371/journal.pone.0037409
Shcheglovitov A, Vitko I, Bidaud I, Baumgart JP, Navarro-Gonzalez MF, Grayson TH, Lory P, Hill CE, Perez-Reyes E (2008) Alternative splicing within the I-II loop controls surface expression of T-type Ca(v)3.1 calcium channels. FEBS Lett 582:3765–3770. https://doi.org/10.1016/j.febslet.2008.10.013
Singh B, Monteil A, Bidaud I, Sugimoto Y, Suzuki T, Hamano S, Oguni H, Osawa M, Alonso ME, Delgado-Escueta AV, Inoue Y, Yasui-Furukori N, Kaneko S, Lory P, Yamakawa K (2007) Mutational analysis of CACNA1G in idiopathic generalized epilepsy. Mutation in brief #962. Online. Hum Mutat 28:524–525. doi:https://doi.org/10.1002/humu.9491
Smith CL, Abdallah S, Wong YY, Le P, Harracksingh AN, Artinian L, Tamvacakis AN, Rehder V, Reese TS, Senatore A (2017) Evolutionary insights into T-type Ca(2+) channel structure, function, and ion selectivity from the Trichoplax adhaerens homologue. J Gen Physiol 149:483–510. https://doi.org/10.1085/jgp.201611683
Snutch TP, Zamponi GW (2018) Recent advances in the development of T-type calcium channel blockers for pain intervention. Br J Pharmacol 175:2375–2383. https://doi.org/10.1111/bph.13906
Souza IA, Gandini MA, Wan MM, Zamponi GW (2016) Two heterozygous Cav3.2 channel mutations in a pediatric chronic pain patient: recording condition-dependent biophysical effects. Pflugers Arch 468:635–642. https://doi.org/10.1007/s00424-015-1776-3
Souza IA, Gandini MA, Zhang FX, Mitchell WG, Matsumoto J, Lerner J, Pierson TM, Zamponi GW (2019) Pathogenic Cav3.2 channel mutation in a child with primary generalized epilepsy. Mol Brain 12:86. https://doi.org/10.1186/s13041-019-0509-5
Splawski I, Timothy KW, Sharpe LM, Decher N, Kumar P, Bloise R, Napolitano C, Schwartz PJ, Joseph RM, Condouris K, Tager-Flusberg H, Priori SG, Sanguinetti MC, Keating MT (2004) Ca(V)1.2 calcium channel dysfunction causes a multisystem disorder including arrhythmia and autism. Cell 119:19–31. https://doi.org/10.1016/j.cell.2004.09.011
Splawski I, Yoo DS, Stotz SC, Cherry A, Clapham DE, Keating MT (2006) CACNA1H mutations in autism spectrum disorders. J Biol Chem 281:22085–22091. https://doi.org/10.1074/jbc.M603316200
Stringer RN, Jurkovicova-Tarabova B, Huang S, Haji-Ghassemi O, Idoux R, Liashenko A, Souza IA, Rzhepetskyy Y, Lacinova L, Van Petegem F, Zamponi GW, Pamphlett R, Weiss N (2020) A rare CACNA1H variant associated with amyotrophic lateral sclerosis causes complete loss of Cav3.2 T-type channel activity. Mol Brain 13:33. https://doi.org/10.1186/s13041-020-00577-6
Strom SP, Stone JL, Ten Bosch JR, Merriman B, Cantor RM, Geschwind DH, Nelson SF (2010) High-density SNP association study of the 17q21 chromosomal region linked to autism identifies CACNA1G as a novel candidate gene. Mol Psychiatry 15:996–1005. https://doi.org/10.1038/mp.2009.41
Talavera K, Staes M, Janssens A, Droogmans G, Nilius B (2004) Mechanism of arachidonic acid modulation of the T-type Ca2+ channel alpha1G. J Gen Physiol 124:225–238. https://doi.org/10.1085/jgp.200409050
Talley EM, Cribbs LL, Lee JH, Daud A, Perez-Reyes E, Bayliss DA (1999) Differential distribution of three members of a gene family encoding low voltage-activated (T-type) calcium channels. J Neurosci 19:1895–1911
Tatsuki F, Sunagawa GA, Shi S, Susaki EA, Yukinaga H, Perrin D, Sumiyama K, Ukai-Tadenuma M, Fujishima H, Ohno R, Tone D, Ode KL, Matsumoto K, Ueda HR (2016) Involvement of Ca(2+)-dependent hyperpolarization in sleep duration in mammals. Neuron 90:70–85. https://doi.org/10.1016/j.neuron.2016.02.032
Timic Stamenic T, Feseha S, Valdez R, Zhao W, Klawitter J, Todorovic SM (2019) Alterations in oscillatory behavior of central medial thalamic neurons demonstrate a key role of CaV3.1 isoform of T-channels during isoflurane-induced anesthesia. Cereb Cortex 29:4679–4696. https://doi.org/10.1093/cercor/bhz002
Todorovic SM, Jevtovic-Todorovic V (2013) Neuropathic pain: role for presynaptic T-type channels in nociceptive signaling. Pflugers Arch 465:921–927. https://doi.org/10.1007/s00424-012-1211-y
Todorovic SM, Jevtovic-Todorovic V (2014) Redox regulation of neuronal voltage-gated calcium channels. Antioxid Redox Signal 21:880–891. https://doi.org/10.1089/ars.2013.5610
Traboulsie A, Chemin J, Chevalier M, Quignard JF, Nargeot J, Lory P (2007) Subunit-specific modulation of T-type calcium channels by zinc. J Physiol 578:159–171. https://doi.org/10.1113/jphysiol.2006.114496
Tsakiridou E, Bertollini L, de Curtis M, Avanzini G, Pape HC (1995) Selective increase in T-type calcium conductance of reticular thalamic neurons in a rat model of absence epilepsy. J Neurosci 15:3110–3117
van Loo KM, Schaub C, Pitsch J, Kulbida R, Opitz T, Ekstein D, Dalal A, Urbach H, Beck H, Yaari Y, Schoch S, Becker AJ (2015) Zinc regulates a key transcriptional pathway for epileptogenesis via metal-regulatory transcription factor 1. Nat Commun 6:8688. https://doi.org/10.1038/ncomms9688
Veselovskii NS, Fedulova SA (1983) 2 types of calcium channels in the somatic membrane of spinal ganglion neurons in the rat. Dokl Akad Nauk SSSR 268:747–750
Visa A, Shaikh S, Alza L, Herreros J, Canti C (2019) The hard-to-close window of T-type calcium channels. Trends Mol Med 25:571–584. https://doi.org/10.1016/j.molmed.2019.03.001
Vitko I, Chen Y, Arias JM, Shen Y, Wu XR, Perez-Reyes E (2005) Functional characterization and neuronal modeling of the effects of childhood absence epilepsy variants of CACNA1H, a T-type calcium channel. J Neurosci 25:4844–4855. https://doi.org/10.1523/JNEUROSCI.0847-05.2005
Vitko I, Bidaud I, Arias JM, Mezghrani A, Lory P, Perez-Reyes E (2007) The I-II loop controls plasma membrane expression and gating of Ca(v)3.2 T-type Ca2+ channels: a paradigm for childhood absence epilepsy mutations. J Neurosci 27:322–330. https://doi.org/10.1523/JNEUROSCI.1817-06.2007
Voisin T, Bourinet E, Lory P (2016) Genetic alteration of the metal/redox modulation of Cav3.2 T-type calcium channel reveals its role in neuronal excitability. J Physiol 594:3561–3574. https://doi.org/10.1113/JP271925
Weiss N, Zamponi GW (2019) T-type channel druggability at a crossroads. ACS Chem Neurosci 10:1124–1126. https://doi.org/10.1021/acschemneuro.9b00031
Weiss N, Zamponi GW (2020) Genetic T-type calcium channelopathies. J Med Genet 57:1–10. https://doi.org/10.1136/jmedgenet-2019-106163
Weiss N, Hameed S, Fernandez-Fernandez JM, Fablet K, Karmazinova M, Poillot C, Proft J, Chen L, Bidaud I, Monteil A, Huc-Brandt S, Lacinova L, Lory P, Zamponi GW, De Waard M (2012) A Ca(v)3.2/syntaxin-1A signaling complex controls T-type channel activity and low-threshold exocytosis. J Biol Chem 287:2810–2818. https://doi.org/10.1074/jbc.M111.290882
Weiss N, Black SA, Bladen C, Chen L, Zamponi GW (2013) Surface expression and function of Cav3.2 T-type calcium channels are controlled by asparagine-linked glycosylation. Pflugers Arch 465:1159–1170. https://doi.org/10.1007/s00424-013-1259-3
Welsby PJ, Wang H, Wolfe JT, Colbran RJ, Johnson ML, Barrett PQ (2003) A mechanism for the direct regulation of T-type calcium channels by Ca2+/calmodulin-dependent kinase II. J Neurosci 23:10116–10121
Wolfe JT, Wang H, Howard J, Garrison JC, Barrett PQ (2003) T-type calcium channel regulation by specific G-protein betagamma subunits. Nature 424:209–213. https://doi.org/10.1038/nature01772
Xie Y, Huang D, Wei L, Luo XJ (2018) Further evidence for the genetic association between CACNA1I and schizophrenia. Hereditas 155:16. https://doi.org/10.1186/s41065-017-0054-0
Xu W, Liu Y, Chen J, Guo Q, Liu K, Wen Z, Zhou Z, Song Z, Zhou J, He L, Yi Q, Shi Y (2018) Genetic risk between the CACNA1I gene and schizophrenia in Chinese Uygur population. Hereditas 155:5. https://doi.org/10.1186/s41065-017-0037-1
Yaari Y, Hamon B, Lux HD (1987) Development of two types of calcium channels in cultured mammalian hippocampal neurons. Science 235:680–682. https://doi.org/10.1126/science.2433765
Yunker AM, Sharp AH, Sundarraj S, Ranganathan V, Copeland TD, McEnery MW (2003) Immunological characterization of T-type voltage-dependent calcium channel CaV3.1 (alpha 1G) and CaV3.3 (alpha 1I) isoforms reveal differences in their localization, expression, and neural development. Neuroscience 117:321–335. https://doi.org/10.1016/s0306-4522(02)00936-3
Zamponi GW, Lory P, Perez-Reyes E (2010) Role of voltage-gated calcium channels in epilepsy. Pflugers Arch 460:395–403. https://doi.org/10.1007/s00424-009-0772-x
Zamponi GW, Striessnig J, Koschak A, Dolphin AC (2015) The physiology, pathology, and pharmacology of voltage-gated calcium channels and their future therapeutic potential. Pharmacol Rev 67:821–870. https://doi.org/10.1124/pr.114.009654
Zhang Y, Mori M, Burgess DL, Noebels JL (2002) Mutations in high-voltage-activated calcium channel genes stimulate low-voltage-activated currents in mouse thalamic relay neurons. J Neurosci 22:6362–6371. https://doi.org/10.1523/JNEUROSCI.22-15-06362.2002
Zhang Y, Jiang X, Snutch TP, Tao J (2013) Modulation of low-voltage-activated T-type Ca(2)(+) channels. Biochim Biophys Acta 1828:1550–1559. https://doi.org/10.1016/j.bbamem.2012.08.032
Zhao Y, Huang G, Wu Q, Wu K, Li R, Lei J, Pan X, Yan N (2019) Cryo-EM structures of apo and antagonist-bound human Cav3.1. Nature 576:492–497. https://doi.org/10.1038/s41586-019-1801-3
Zhong X, Liu JR, Kyle JW, Hanck DA, Agnew WS (2006) A profile of alternative RNA splicing and transcript variation of CACNA1H, a human T-channel gene candidate for idiopathic generalized epilepsies. Hum Mol Genet 15:1497–1512. https://doi.org/10.1093/hmg/ddl068
Acknowledgments
This work is supported by the Labex “Ion Channel Science and Therapeutics” (ICST), Fondation Maladies Rares (FMR), Association Française contre les Myopathies (AFM), and Fondation pour la Recherche Médicale (FRM).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the special issue on Channelopathies: from mutation to diseases in Pflügers Archiv—European Journal of Physiology
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lory, P., Nicole, S. & Monteil, A. Neuronal Cav3 channelopathies: recent progress and perspectives. Pflugers Arch - Eur J Physiol 472, 831–844 (2020). https://doi.org/10.1007/s00424-020-02429-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00424-020-02429-7