Effects of PEG-Coated Silver and Gold Nanoparticles on Spirulina platensis Biomass during Its Growth in a Closed System

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nanoparticles
2.2. Nanoparticles Characterization
2.3. Object of Study
2.4. Experiment
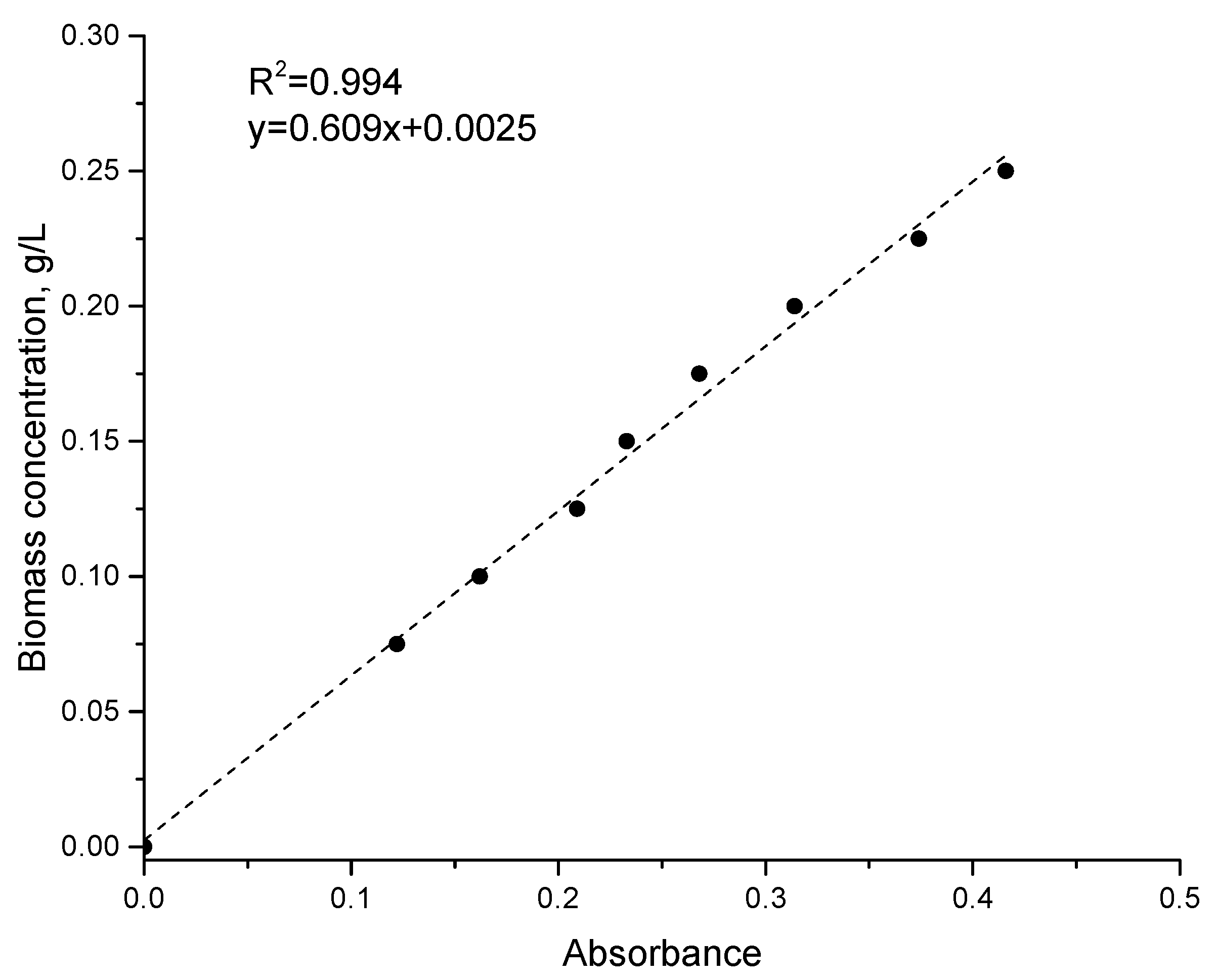
2.5. Biochemical Analysis
2.6. Antioxidant Activity by the ABTS+ Radical Cation Assay (ABTS Method)
2.7. TEM Analysis
3. Results
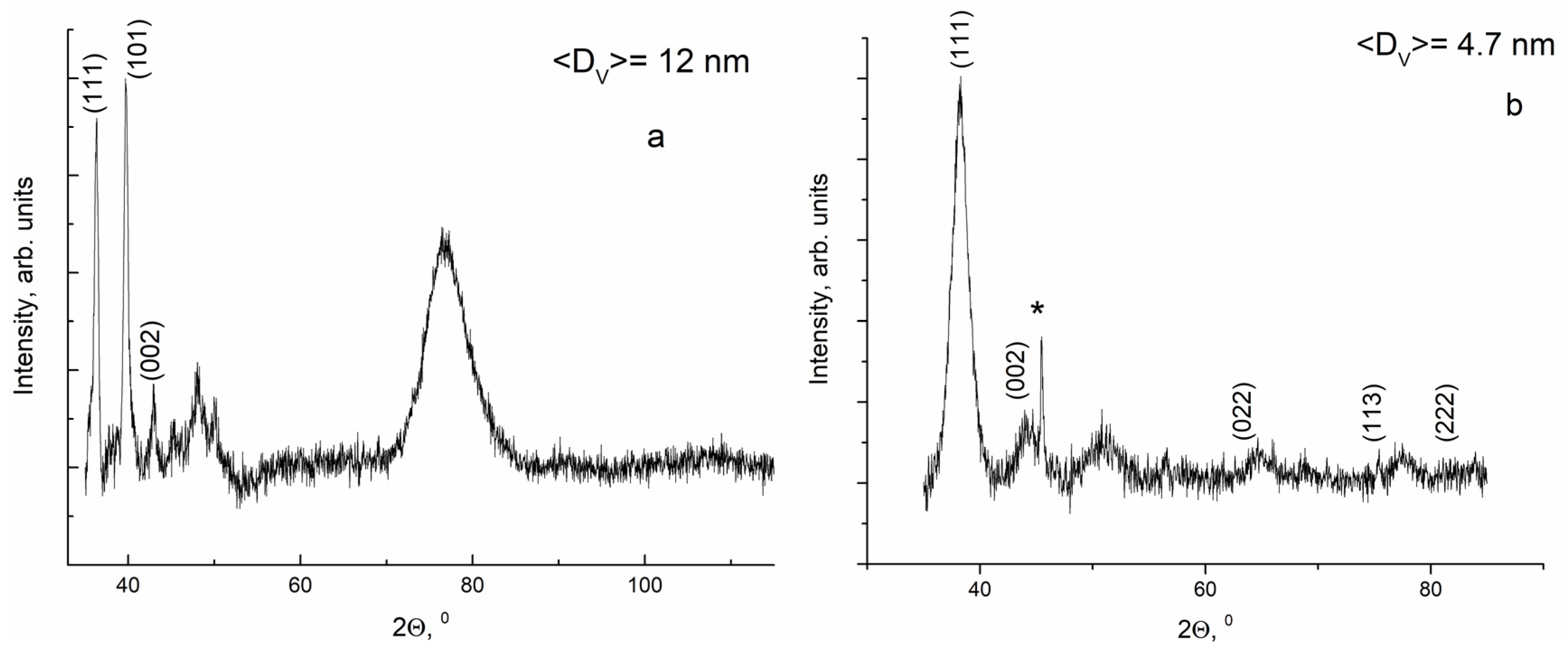
3.1. Nanoparticles Characterization
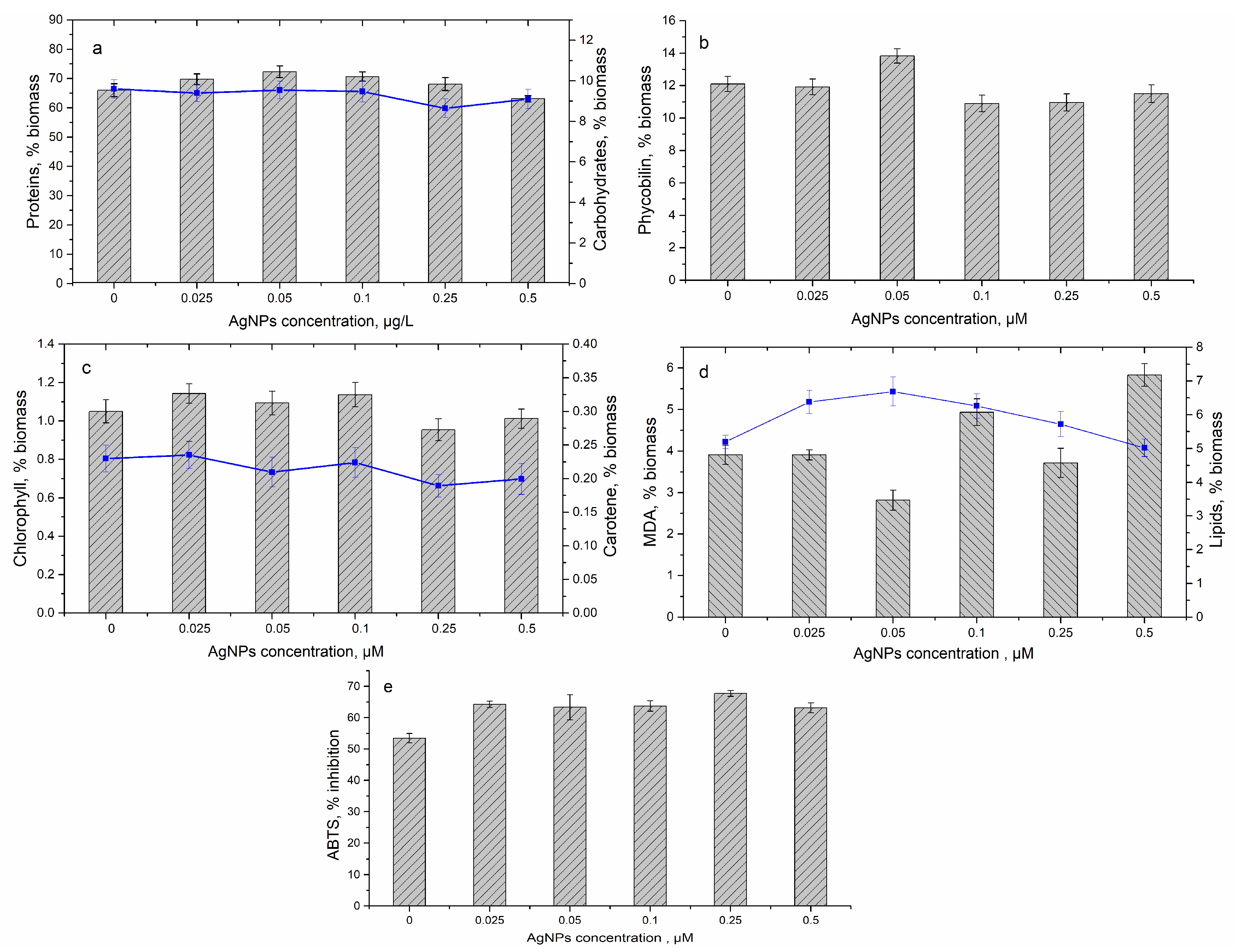
3.2. Effect of Silver Nanoparticles on Spirulina platensis Growth
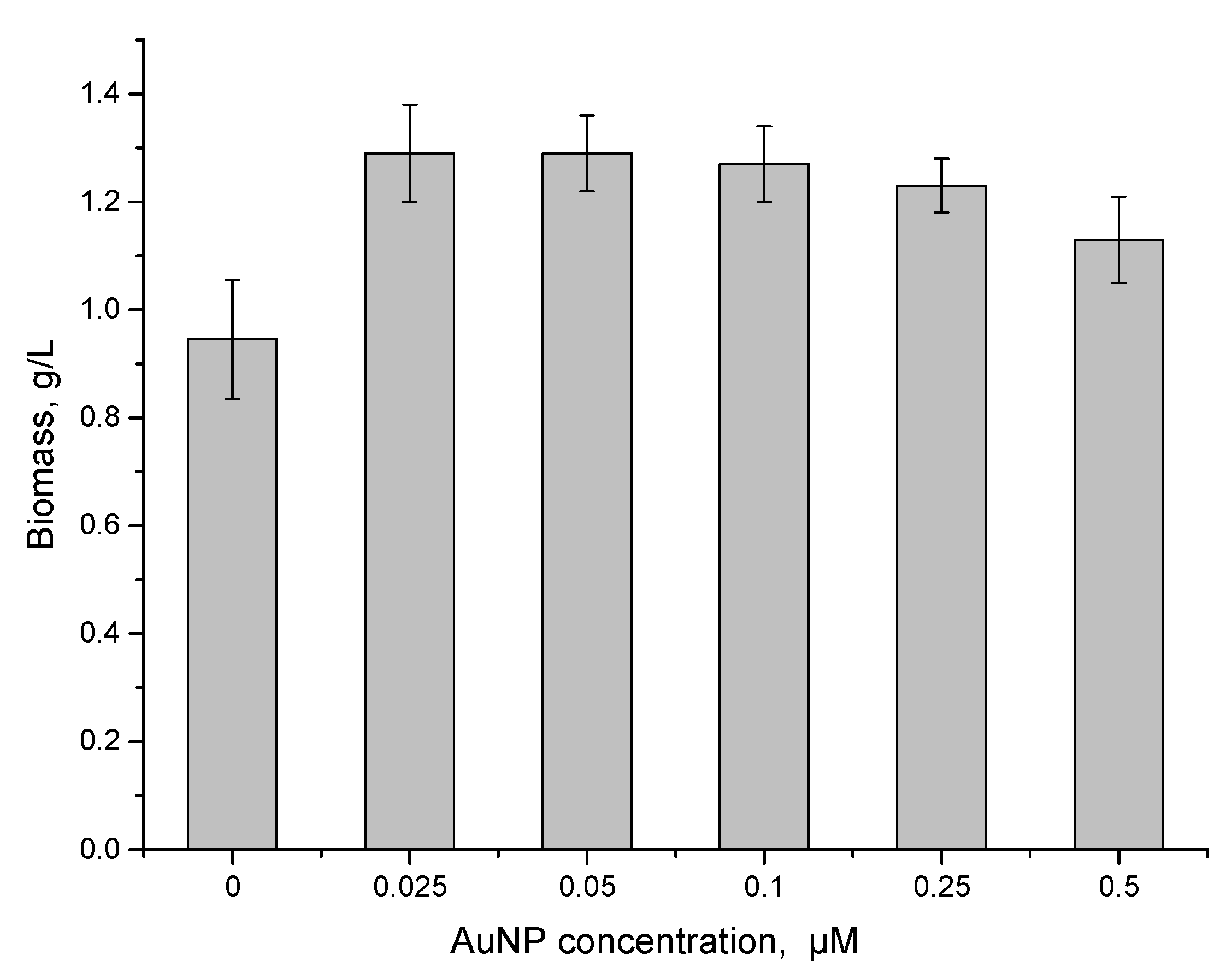
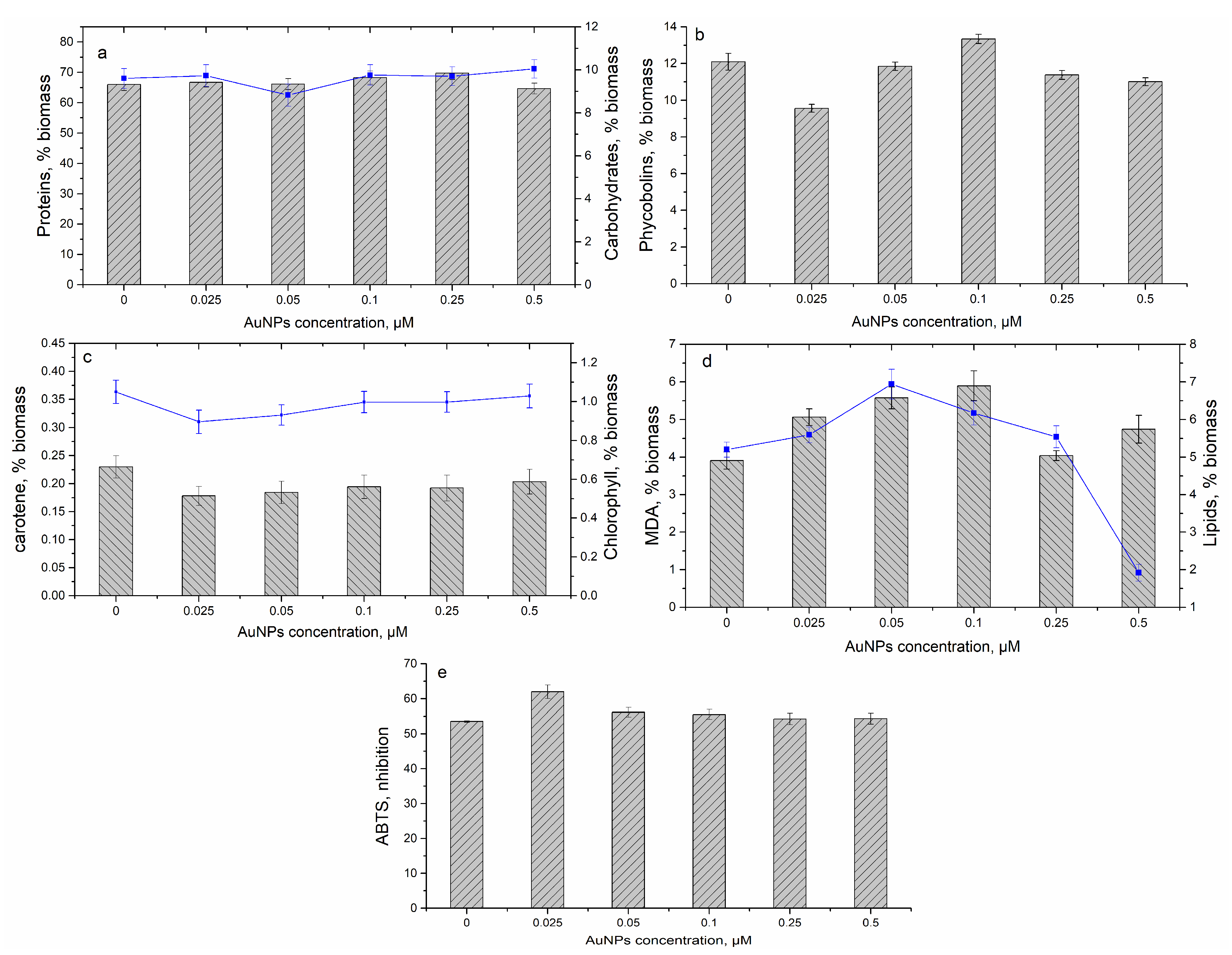
3.3. Effect of Gold Nanoparticles on Spirulina platensis Growth
3.4. Transmission Electron Microscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Planchon, M.; Jittawuttipoka, T.; Cassier-Chauvat, C.; Guyot, F.; Gelabert, A.; Benedetti, M.F.; Chauvat, F.; Spalla, O. Exopolysaccharides protect Synechocystis against the deleterious effects of titanium dioxide nanoparticles in natural and artificial waters. J. Colloid Interface Sci. 2013, 405, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Pulit-Prociak, J.; Banach, M. Silver nanoparticles—A material of the future…? Open Chem. 2016, 14, 76–91. [Google Scholar] [CrossRef]
- Elahi, N.; Kamali, M.; Baghersad, M.H. Recent biomedical applications of gold nanoparticles: A review. Talanta 2018, 184, 537–556. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Jiménez, B.; Johnson, M.E.; Montoro Bustos, A.R.; Murphy, K.E.; Winchester, M.R.; Vega Baudrit, J.R. Silver nanoparticles: Technological advances, societal impacts, and metrological challenges. Front. Chem. 2017, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Chaloupka, K.; Malam, Y.; Seifalian, A.M. Nanosilver as a new generation of nanoproduct in biomedical applications. Trends Biotechnol. 2010, 28, 580–588. [Google Scholar] [CrossRef]
- Chen, H.; Gao, F.; He, R.; Cui, D. Chemiluminescence of luminol catalyzed by silver nanoparticles. J. Colloid Interface Sci. 2007, 315, 158–163. [Google Scholar] [CrossRef]
- Nair, L.S.; Laurencin, C.T. Silver nanoparticles: Synthesis and therapeutic applications. J. Biomed. Nanotechnol. 2007, 3, 301–316. [Google Scholar] [CrossRef]
- Prabhu, S.; Poulose, E.K. Silver nanoparticles: Mechanism of antimicrobial action, synthesis, medical applications, and toxicity effects. Int. Nano Lett. 2012, 2, 32. [Google Scholar] [CrossRef] [Green Version]
- Savithramma, N.; Linga Rao, M.; Rukmini, K.; Suvarnalatha Devi, P. Antimicrobial activity of silver nanoparticles synthesized by using medicinal plants. Int. J. Chemtech. Res. 2011, 3, 1394–1402. Available online: https://pdfs.semanticscholar.org/a482/f5f6f29aff65a1c82f692cde97c0884ba997.pdf (accessed on 7 June 2020).
- Marques, L.; Martinez, G.; Guidelli, É.; Tamashiro, J.; Segato, R.; Payão, S.L.M.; Baffa, O.; Kinoshita, A. Performance on bone regeneration of a silver nanoparticle delivery system based on natural rubber membrane NRL-AgNP. Coatings 2020, 10, 323. [Google Scholar] [CrossRef] [Green Version]
- Parashar, V.; Parashar, R.; Sharma, B.; Pandey, A.C. Parthenium leafe extract mediated synthesis of silver nanoparticles: A novel approach towards weed utilization. Dig. J. Nanomater. Biostruct. 2009, 4, 45–50. [Google Scholar]
- Krishnaraj, C.; Jagan, E.G.; Ramachandran, R.; Abirami, S.M.; Mohan, N.; Kalaichelvan, P.T. Effect of biologically synthesized silver nanoparticles on Bacopa monnieri (Linn.) Wettst. plant growth metabolism. Process. Biochem. 2012, 47, 651–658. [Google Scholar] [CrossRef]
- Hassanen, E.I.; Morsy, E.A.; Hussien, A.M.; Ibrahim, M.A.; Farroh, K.Y. The effect of different concentrations of gold nanoparticles on growth performance, toxicopathological and immunological parameters of broiler chickens. Biosci. Rep. 2020, 40, BSR20194296. [Google Scholar] [CrossRef] [Green Version]
- Sengani, M.; Grumezescu, A.M.; Rajeswaria, V.D. Recent trends and methodologies in gold nanoparticle synthesis—A prospective review on drug delivery aspect. Open Nano 2017, 2, 37–46. [Google Scholar] [CrossRef]
- Cabuzu, D.; Cirja, A.; Puiu, R.; Grumezescu, A.M. Biomedical applications of gold nanoparticles. Curr. Top. Med. Chem. 2015, 15, 1605–1613. [Google Scholar] [CrossRef]
- Cai, W.; Gao, T.; Hong, H.; Sun, J. Applications of gold nanoparticles in cancer nanotechnology. Nanotechnol. Sci. Appl. 2008, 1, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Rahme, K.; He, Y.; Li, L.L.; Holmes, J.; O’Driscoll, C. Gold nanoparticles enlighten the future of cancer theranostics. Int. J. Nanomed. 2017, 12, 6131–6152. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.; Pinnaka, A.K.; Raje, M.; Fnu, A.; Bhattacharyya, M.S.; Choudhury, A.R. Exploitation of marine bacteria for production of gold nanoparticles. Microb. Cell Factories 2012, 11, 86. [Google Scholar] [CrossRef] [Green Version]
- Ahuja, K.; Mamtani, K. Gold Nanoparticles Market Size by Application (Electronics, Medical & Dentistry, Catalysis), Industry Analysis Report, Regional Outlook, Application Potential, Price Trend, Competitive Market Share & Forecast, 2015–2022; Global Market Insights, Inc.: Selbyville, DE, USA, 2019. [Google Scholar]
- Niazi, J.H.; Gu, M.B. Toxicity of metallic nanoparticles in microorganisms—A review. In Atmospheric and Biological Environmental Monitoring; Kim, Y.J., Platt, U., Gu, M.B., Iwahashi, H., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 193–206. [Google Scholar] [CrossRef]
- Zinicovscaia, I.; Pavlov, S.S.; Frontasyeva, M.V.; Ivlieva, A.L.; Petritskaya, E.N.; Rogatkin, D.A.; Demin, V.A. Accumulation of silver nanoparticles in mice tissues studied by neutron activation analysis. J. Radioanal. Nucl. Chem. 2018, 318, 985–989. [Google Scholar] [CrossRef]
- Marsalek, B.; Jancula, D.; Marsalkova, E.; Mashlan, M.; Safarova, K.; Tucek, J.; Zboril, R. Multimodal action and selective toxicity of zerovalent iron nanoparticles against cyanobacteria. Environ. Sci. Technol. 2012, 46, 2316–2323. [Google Scholar] [CrossRef]
- Abed, R.M.M.; Dobretsov, S.; Sudesh, K. Applications of cyanobacteria in biotechnology. J. Appl. Microbiol. 2009, 106, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Campanella, L.; Crescentini, G.; Avino, P. Chemical composition and nutritional evaluation of some natural and commercial food products based on Spirulina. Analusis 1999, 27, 533–540. [Google Scholar] [CrossRef]
- Thajuddin, N.; Subramanian, G. Cyanobacterial biodiversity and potential applications in biotechnology. Curr. Sci. 2005, 89, 47–57. Available online: https://pdfs.semanticscholar.org/98e1/4c455aa390fa156350be3b4433421785fec1.pdf (accessed on 12 May 2020).
- Burchardt, A.D.; Carvalho, R.N.; Valente, A.; Nativo, P.; Gilliland, D.; Garcìa, C.P.; Passarella, R.; Pedroni, V.; Rossi, F.; Lettieri, T. Effects of silver nanoparticles in diatom Thalassiosira pseudonana and cyanobacterium synechococcus sp. Environ. Sci. Technol. 2012, 46, 11336–11344. [Google Scholar] [CrossRef]
- Cherchi, C.; Gu, A.Z. Impact of titanium dioxide nanomaterials on nitrogen fixation rate and intracellular nitrogen storage in Anabaena variabilis. Environ. Sci. Technol. 2010, 44, 8302–8307. [Google Scholar] [CrossRef] [PubMed]
- Shuguang, W.; Lawson, R.; Ray, P.C.; Hongtao, Y. Toxic effects of gold nanoparticles on Salmonella typhimurium bacteria. Toxicol. Ind. Health 2011, 27, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Sukumaran, P.; Nulit, R.; Zulkifly, S.; Halimoon, N.; Omar, H.; Ismail, A. Potential of fresh POME as a growth medium in mass production of Arthrospira platensis. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 235–250. [Google Scholar]
- Nor, N.M.; Naqqiuddin, M.A.; Mashor, N.; Zulkifly, S.; Oar, H.; Ismail, A. The effect of different nitrogen sources on continuous growth of Arthrospira platensis in simple floating photobiorector design in outdoor conditions. J. Algal Biomass Util. 2015, 6, 1–11. [Google Scholar]
- Boussiba, S.; Richmond, A.E. C-phycocyanin as a storage protein in the blue-green alga Spirulina platensis. Arch. Microbiol. 1980, 123, 143–147. [Google Scholar] [CrossRef]
- Park, J.Y.; Jeong, H.J.; Yoon, E.Y.; Moon, S.J. Easy and rapid quantification of lipid cantent of marine dinoflagellates using the sulpho-phospho-vanillin method. Algae 2016, 31, 391–401. [Google Scholar] [CrossRef] [Green Version]
- Lejaeghere, K.; Van Speybroeck, V.; Van Oost, G.; Cottenier, S. Error estimates for solid-state density-functional theory predictions: An overview by means of the ground-state elemental crystals. Crit. Rev. Solid State Mater. Sci. 2014, 39, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Novgorodovo, M.I.; Gorshkov, A.I.; Mokhov, A.V. Native silver and its new structural modifications. Int. Geol. Rev. 1981, 23, 485–494. [Google Scholar] [CrossRef]
- Upadhya, S.; Parekh, K.; Pandey, B. Influence of crystallite size on the magnetic properties of Fe3O4 nanoparticles. J. Alloy. Compd. 2016, 678, 478–485. [Google Scholar] [CrossRef]
- Choi, O.; Hu, Z. Size dependent and reactive oxygen species related nanosilver toxicity to nitrifying bacteria. Environ. Sci. Technol. 2008, 42, 4583–4588. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Garrido, I.; Pérez, S.; Blasco, J. Toxicity of silver and gold nanoparticles on marine microalgae. Mar. Environ. Res. 2015, 111, 60–73. [Google Scholar] [CrossRef]
- Yang, X.; Gondikas, A.P.; Marinakos, S.M.; Auffan, M.; Liu, J.; Hsu-Kim, H.; Meyer, J.N. Mechanism of silver nanoparticle toxicity is dependent on dissolved silver and surface coating in caenorhabditis elegans. Environ. Sci. Technol. 2012, 46, 1119–1127. [Google Scholar] [CrossRef]
- Pinzaru, I.; Coricovac, D.; Dehelean, C.; Moacă, E.A.; Mioc, M.; Baderca, F.; Sizemore, I.; Brittle, S.; Marti, D.; Calina, C.D.; et al. Stable PEG-coated silver nanoparticles—A comprehensive toxicological profile. Food Chem. Toxicol. 2018, 111, 546–556. [Google Scholar] [CrossRef]
- Bastos, V.; Ferreira de Oliveira, J.M.P.; Brown, D.; Jonhston, H.; Malheiro, E.; Daniel-da-Silva, A.L.; Duarte, I.F.; Santos, C.; Oliveira, H. The influence of Citrate or PEG coating on silver nanoparticle toxicity to a human keratinocyte cell line. Toxicol. Lett. 2016, 249, 29–41. [Google Scholar] [CrossRef]
- Hazeem, L.J.; Kuku, G.; Dewailly, E.; Slomianny, C.; Barras, A.; Hamdi, A.; Boukherroub, R.; Culha, M.; Bououdina, M. Toxicity effect of silver nanoparticles on photosynthetic pigment content, growth, ROS production and ultrastructural changes of microalgae Chlorella vulgaris. Nanomaterials (Basel) 2019, 9, 914. [Google Scholar] [CrossRef] [Green Version]
- Nam, S.H.; Kwak, J.I.; An, Y.J. Quantification of silver nanoparticle toxicity to algae in soil via photosynthetic and flow-cytometric analyses. Sci. Rep. 2018, 8, 292. [Google Scholar] [CrossRef] [Green Version]
- Johari, S.A.; Sarkheil, M.; Tayemeh, M.B.; Veisi, S. Influence of salinity on the toxicity of silver nanoparticles (AgNPs) and silver nitrate (AgNO3) in halophilic microalgae, Dunaliella salina. Chemosphere 2018, 209, 156–162. [Google Scholar] [CrossRef]
- Pascual García, C.; Burchardt, A.D.; Carvalho, R.N.; Gilliland, D.; António, D.C.; Rossi, F.; Lettieri, T. Detection of silver nanoparticles inside marine diatom Thalassiosira pseudonana by electron microscopy and focused ion beam. PLoS ONE 2014, 9, e96078. [Google Scholar] [CrossRef] [Green Version]
- Pletikapić, G.; Žutić, V.; Vinković Vrček, I.; Svetličić, V. Atomic force microscopy characterization of silver nanoparticles interactions with marine diatom cells and extracellular polymeric substance. J. Mol. Recognit. 2012, 25, 309–317. [Google Scholar] [CrossRef]
- Fratoddi, I.; Venditti, I.; Cametti, C.; Russo, M.V. How toxic are gold nanoparticles? The state-of-the-art. Nano Res. 2015, 8, 1771–1799. [Google Scholar] [CrossRef]
- Buchman, J.T.; Rahnamoun, A.; Landy, K.M.; Zhang, X.; Vartanian, A.M.; Jacob, L.M.; Murphy, C.J.; Hernandez, R.; Haynes, C.L. Using an environmentally-relevant panel of Gram-negative bacteria to assess the toxicity of polyallylamine hydrochloride-wrapped gold nanoparticles. Environ. Sci. Nano 2018, 5, 279–288. [Google Scholar] [CrossRef]
- Liu, K.; He, Z.; Byrne, H.J.; Curtin, J.F.; Tian, F. Investigating the role of gold nanoparticle shape and size in their toxicities to fungi. Int. J. Environ. Res. Public Health 2018, 15, 998. [Google Scholar] [CrossRef] [Green Version]
- Boda, S.K.; Broda., J.; Schiefer, F.; Weber-Heynemann, J.; Hoss, M.; Simon., U.; Basu, B.; Jahnen-Dechent, W. Cytotoxicity of ultrasmall gold nanoparticles on planktonic and biofilm encapsulated gram-positive staphylococci. Small 2015, 11, 3183–3193. [Google Scholar] [CrossRef]
- Van Hoecke, K.; De Schamphelaere, K.A.C.; Ali, Z.; Zhang, F.; Elsaesser, A.; Rivera-Gil, P.; Parak, W.J.; Smagghe, G.; Hivard, C.V.; Janssen, C.R. Ecotoxicity and uptake of polymer coated gold nanoparticles. Nanotoxicology 2013, 7, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, A.D.; Parak, W.J.; Zhang, F.; Ali, Z.; Röcker, C.; Nienhaus, G.U.; Gehr, P.; Rothen-Rutishauser, B. Fluorescent-magnetic hybrid nanoparticles induce a dose-dependent increase in proinflammatory response in lung cells in vitro correlated with intracellular localization. Small 2010, 6, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Tatur, S.; MacCarini, M.; Barker, R.; Nelson, A.; Fragneto, G. Effect of functionalized gold nanoparticles on floating lipid bilayers. Langmuir 2013, 29, 6606–6614. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.M.; Fouad, S.A.; Elshoky, H.A.; Mohammed, G.M.; Salaheldin, T.A. Antibacterial effect of gold nanoparticles against Corynebacterium pseudotuberculosis. Int. J. Vet. Sci. Med. 2017, 5, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pajerski, W.; Ochonska, D.; Brzychczy-Wloch, M.; Indyka, P.; Jarosz, M.; Golda-Cepa, M.; Sojka, Z.; Kotarba, A. Attachment efficiency of gold nanoparticles by Gram-positive and Gram-negative bacterial strains governed by surface charges. J. Nanoparticle Res. 2019, 21, 186. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Nanoparticles | Chl/Carotene Ratio | |||||
|---|---|---|---|---|---|---|
| Depending on the Nanoparticles Concentration, µM | ||||||
| 0 (C) | 0.025 | 0.05 | 0.10 | 0.25 | 0.50 | |
| AgNPs | 4.57 | 4.86 | 5.21 | 5.07 | 5.04 | 4.17 |
| AuNPs | 4.57 | 5.03 | 5.05 | 5.13 | 5.18 | 5.05 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cepoi, L.; Zinicovscaia, I.; Rudi, L.; Chiriac, T.; Rotari, I.; Turchenko, V.; Djur, S. Effects of PEG-Coated Silver and Gold Nanoparticles on Spirulina platensis Biomass during Its Growth in a Closed System. Coatings 2020, 10, 717. https://doi.org/10.3390/coatings10080717
Cepoi L, Zinicovscaia I, Rudi L, Chiriac T, Rotari I, Turchenko V, Djur S. Effects of PEG-Coated Silver and Gold Nanoparticles on Spirulina platensis Biomass during Its Growth in a Closed System. Coatings. 2020; 10(8):717. https://doi.org/10.3390/coatings10080717
Chicago/Turabian StyleCepoi, Liliana, Inga Zinicovscaia, Ludmila Rudi, Tatiana Chiriac, Ion Rotari, Vitalii Turchenko, and Svetlana Djur. 2020. "Effects of PEG-Coated Silver and Gold Nanoparticles on Spirulina platensis Biomass during Its Growth in a Closed System" Coatings 10, no. 8: 717. https://doi.org/10.3390/coatings10080717