Gap Analysis for DNA Barcode Reference Libraries for Aquatic Macroinvertebrate Species in the Apulia Region (Southeast of Italy)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Macroinvertebrates Database and Gap-Analysis
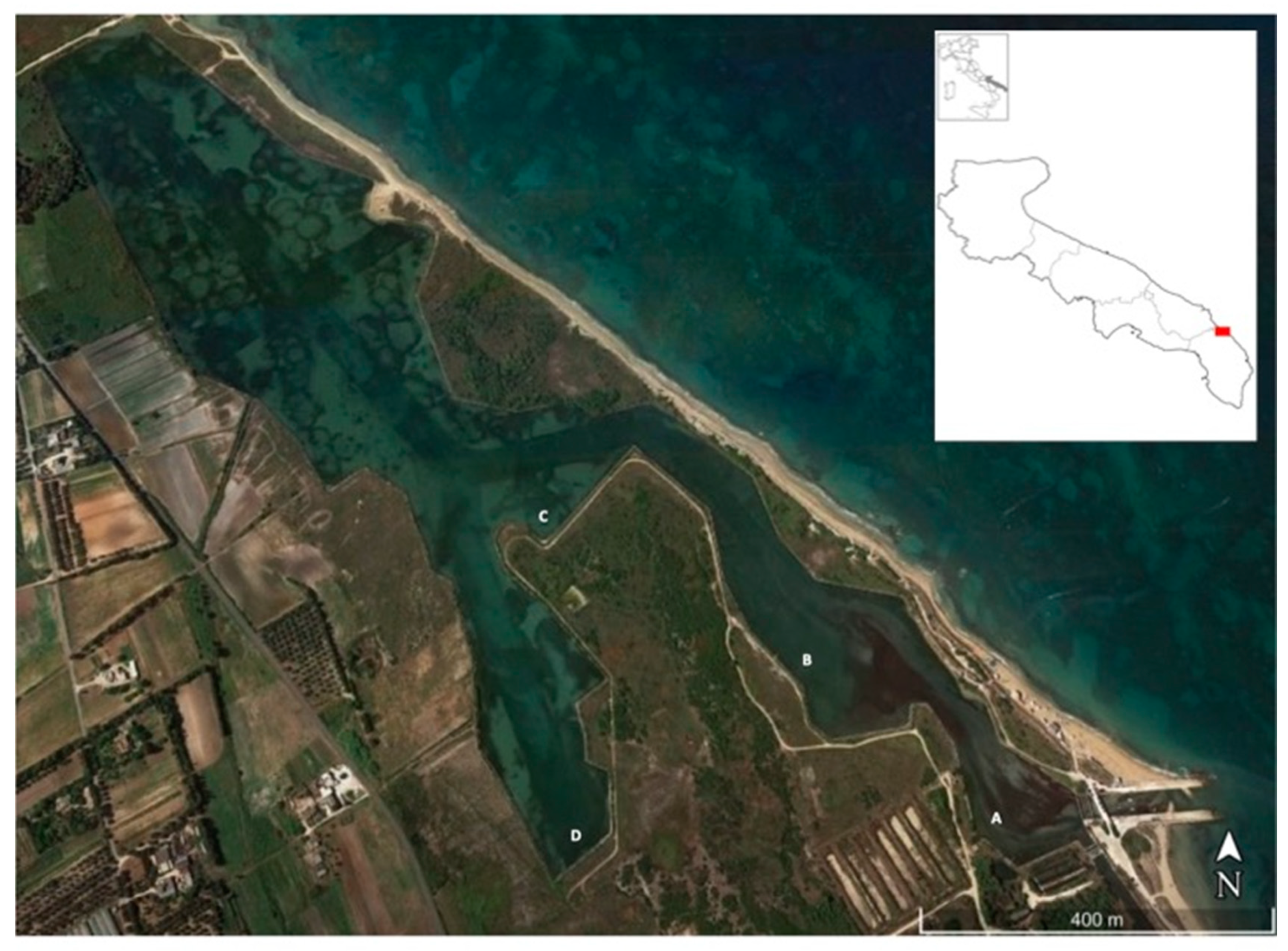
2.2. Field Sampling of Macroinvertebrates
2.3. Single Species DNA-Barcoding
3. Results
3.1. Gap Analysis of the Barcoded Species in the Reference Libraries
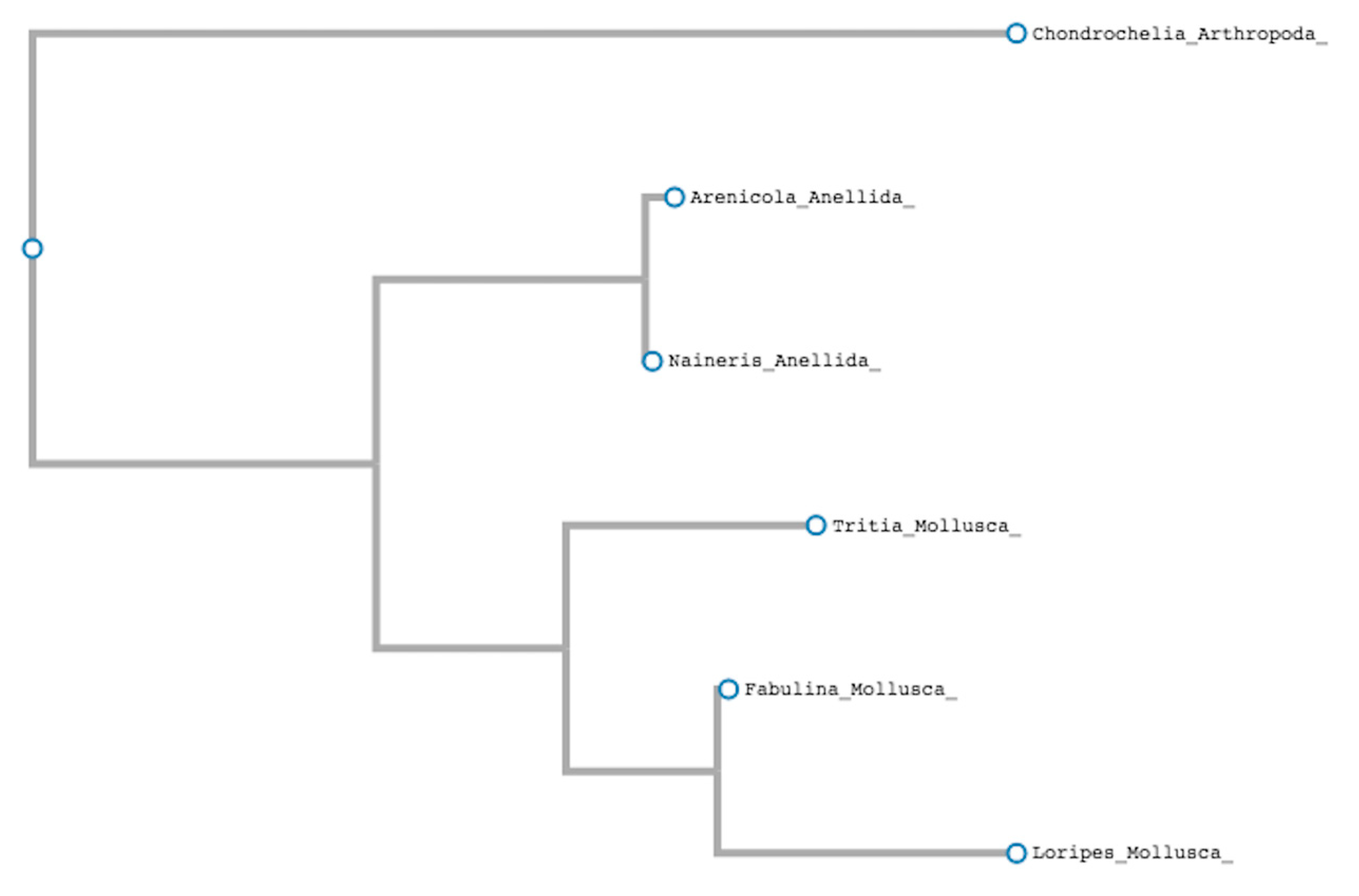
3.2. Application of Single Species DNA Barcoding to a Lagoon Ecosystem and First Identification of the DNA Barcode Sequence in Two Macroinvertabrate Species
3.3. The Identification of Species for Macroinvertabrates Early Developmental Stages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Pinna, M.; Janzen, S.; Franco, A.; Specchia, V.; Marini, G. Role of habitats and sampling techniques on macroinvertebrate descriptors and ecological indicators: An experiment in a protected Mediterranean lagoon. Ecol. Indic. 2017, 83, 495–503. [Google Scholar] [CrossRef]
- Dauer, D.M. Biological criteria environmental health and estuarine macrobenthic community structure. Mar. Pollut. Bull. 1993, 26, 249–257. [Google Scholar] [CrossRef]
- Pinna, M.; Marini, G.; Rosati, I.; Neto, J.M.; Patrício, J.; Marques, J.C.; Basset, A. The usefulness of large body-size macroinvertebrates in the rapid ecological assessment of Mediterranean lagoons. Ecol. Indic. 2013, 29, 48–61. [Google Scholar] [CrossRef]
- Pinna, M.; Marini, G.; Mancinelli, G.; Basset, A. Influence of sampling effort on ecological descriptors and indicators in perturbed and unperturbed conditions: A study case using benthic macroinvertebrates in Mediterranean. Ecol. Indic. 2014, 37, 27–39. [Google Scholar] [CrossRef]
- Thomsen, P.F.; Willerslev, E. Environmental DNA—An emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 2015, 183, 4–18. [Google Scholar] [CrossRef]
- Baird, D.J.; Hajibabaei, M. Biomonitoring 2.0: A new paradigm in ecosystem assessment made possible by next-generation DNA sequencing. Mol. Ecol. 2012, 21, 2039–2044. [Google Scholar] [CrossRef]
- Besansky, N.J.; Severson, D.W.; Ferdig, M.T. DNA barcoding of parasites and invertebrate disease vectors: What you don’t know can hurt you. Trends Parasitol. 2003, 19, 545–546. [Google Scholar] [CrossRef]
- Buckwalter, J.; Angermeier, P.L.; Argentina, J.; Wolf, S.; Floyd, S.; Hallerman, E.M. Drift of larval darters (Family Percidae) in the upper Roanoke River basin, USA, characterized using phenotypic and DNA barcoding markers. Fishes 2019, 4, 59. [Google Scholar] [CrossRef] [Green Version]
- Caesar, R.M.; Sörensson, M.; Cognato, A.I. Integrating DNA data and traditional taxonomy to streamline biodiversity assessment: An example from edaphic beetles in the Klamath ecoregion, California, USA. Divers. Distrib. 2006, 12, 483–489. [Google Scholar] [CrossRef]
- Beermann, A.J.; Zizka, V.M.; Elbrecht, V.; Baranov, V.; Leese, F. DNA metabarcoding reveals the complex and hidden responses of chironomids to multiple stressors. Environ. Sci. Eur. 2018, 30, 26. [Google Scholar] [CrossRef] [Green Version]
- Lobo, J.; Shokralla, S.; Costa, M.H.; Hajibabaei, M.; Costa, F.O. DNA metabarcoding for high-throughput monitoring of estuarine macrobenthic communities. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaxter, M.L. The promise of a DNA taxonomy. Philos. Trans. R. Soc. B 2004, 359, 669–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratnasingham, S.; Hebert, P.D. BOLD: The Barcode of Life Data System. Mol. Ecol. Notes 2007, 7, 355–364. Available online: http://www.barcodinglife.org (accessed on 16 June 2020). [CrossRef] [PubMed] [Green Version]
- Taberlet, P.; Coissac, E.; Pompanon, F.; Brochmann, C.; Willerslev, E. Towards next-generation biodiversity assessment using DNA metabarcoding. Mol. Ecol. 2012, 21, 2045–2050. [Google Scholar] [CrossRef]
- Aylagas, E.; Borja, Á.; Irigoien, X.; Rodríguez-Ezpeleta, N. Benchmarking DNA metabarcoding for biodiversity-based monitoring and assessment. Front. Mar. Sci. 2016, 3, 96. [Google Scholar] [CrossRef] [Green Version]
- Deiner, K.; Bik, H.M.; Mächler, E.; Seymour, M.; Lacoursière-Roussel, A.; Altermatt, F.; Creer, S.; Bista, I.; Lodge, D.M.; de Vere, N.; et al. Environmental DNA metabarcoding: Transforming how we survey animal and plant communities. Mol. Ecol. 2017, 26, 5872–5895. [Google Scholar] [CrossRef]
- Ji, Y.; Ashton, L.; Pedley, S.M.; Edwards, D.P.; Tang, Y.; Nakamura, A.; Kitching, R.; Dolman, P.M.; Woodcock, P.; Edwards, F.A.; et al. Reliable, verifiable and efficient monitoring of biodiversity via metabarcoding. Ecol. Lett. 2013, 16, 1245–1257. [Google Scholar] [CrossRef]
- Rees, H.C.; Maddison, B.C.; Middleditch, D.J.; Patmore, J.R.M.; Gough, K.C. The detection of aquatic animal species using environmental DNA – a review of eDNA as a survey tool in ecology. J. Appl. Ecol. 2014, 51, 1450–1459. [Google Scholar] [CrossRef]
- Carew, M.E.; Pettigrove, V.J.; Metzeling, L.; Hoffmann, A.A. Environmental monitoring using next generation sequencing: Rapid identification of macroinvertebrate bioindicator species. Front. Zool. 2013, 10, 45. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, B.W.; Battle, J.M.; Jackson, J.K.; Dapkey, T. Can DNA barcodes of stream macroinvertebrates improve descriptions of community structure and water quality? J. North. Am. Benthol. Soc. 2011, 30, 195–216. [Google Scholar] [CrossRef]
- Borja, A.; Miles, A.; Occhipinti-Ambrogi, A.; Berg, T. Current status of macroinvertebrate methods used for assessing the quality of European marine waters: Implementing the Water Framework Directive. Hydrobiologia 2009, 633, 181–196. [Google Scholar] [CrossRef]
- Borja, A.; Elliott, M.; Andersen, J.H.; Cardoso, A.C.; Carstensen, J.; Ferreira, J.G.; Heiskanen, A.; Marques, J.C.; Neto, J.M.; Teixeira, H.; et al. Good environmental status of marine ecosystems: What is it and how do we know when we have attained it? Mar. Pollut. Bull. 2013, 76, 16–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leese, F.; Altermatt, F.; Bouchez, A.; Ekrem, T.; Hering, D.; Meissner, K.; Mergen, P.; Pawlowski, J.; Piggott, J.; Rimet, F.; et al. DNAqua-Net: Developing new genetic tools for bioassessment and monitoring of aquatic ecosystems in Europe. Res. Ideas Outcomes 2016, 2, e11321. [Google Scholar] [CrossRef] [Green Version]
- Macher, J.N.; Macher, T.H.; Leese, F. Combining NCBI and BOLD databases for OTU assignment in metabarcoding and metagenomic data: The BOLD_NCBI _Merger. PeerJ 2017, 5, e3133v1. [Google Scholar]
- Weigand, H.; Beermann, A.J.; Čiampor, F.; Costa, F.O.; Csabai, Z.; Duarte, S.; Geigerg, M.F.; Grabowski, M.; Rimet, F.; Rulik, B.; et al. DNA barcode reference libraries for the monitoring of aquatic biota in Europe: Gap-analysis and recommendations for future work. Sci. Total Environ. 2019, 678, 499–524. [Google Scholar] [CrossRef]
- Elbrecht, V.; Leese, F. Validation and development of COI metabarcoding primers for freshwater macroinvertebrate bioassessment. Front. Environ. Sci. 2017, 5, 11. [Google Scholar]
- Agenzia Regionale per la Protezione dell’Ambiente della Puglia, ARPA-Puglia. Monitoraggio di sorveglianza, Relazione finale 2010–2011; ARPA-Puglia: Bari, Italy, 2011. [Google Scholar]
- Marrocco, V.; Zangaro, F.; Sicuro, A.; Pinna, M. A scaling down mapping of Pinna nobilis (Linnaeus, 1758) through the combination of scientific literature, NATURA 2000, grey literature and citizen science data. Nat. Conserv. 2018, 33, 21. [Google Scholar] [CrossRef]
- Marrocco, V.; Sicuro, A.; Zangaro, F.; Pinna, M. First record of the protected species Pinna nobilis (Linnaeus, 1758) in the Aquatina Lagoon (NATURA 2000 site IT9150003, South-East Italian coastline). Nat. Conserv. 2019, 28, 51. [Google Scholar] [CrossRef]
- Bellan-Santini, D.; Karaman, G.; Krapp-Schickel, G.; Ledoyer, M.; Ruffo, S. The amphipoda of the mediterranean. Part 1: Gammaridea (acanthonotozomatidae to gammaridae). In Mémoires de l’Institut Océanographique, Monaco, 13; Institut Océanographique: Monaco, Monaco, 1982; p. 364. ISBN 2-7260-0160-2. [Google Scholar]
- Bellan-Santini, D.; Karaman, G.; Krapp-Schickel, G.; Ledoyer, M.; Ruffo, S. The amphipoda of the mediterranean. Part 2: Gammaridea (haustoriidae to lysianassidae). In Mémoires de l’Institut Océanographique, Monaco, 13; Institut Océanographique: Monaco, Monaco, 1989; p. 576. ISBN 2-7260-0160-2. [Google Scholar]
- Bellan-Santini, D.; Karaman, G.; Krapp-Schickel, G.; Ledoyer, M.; Ruffo, S. The amphipoda of the mediterranean. Part 3: Gammaridea (melphidippidae to talitridae), ingolfiellidea, caprellidea. In Mémoires de l’Institut océanographique, Monaco, 13; Institut Océanographique: Monaco, Monaco, 1993; p. 813. ISBN 2-7260-0160-2. [Google Scholar]
- Campaioli, S.; Ghetto, P.F.; Mineli, A.; Ruffo, S. Manuale Per il Riconoscimento del Macroinvertebrati Delle Acque Dolci Italiane; Provincia Autonoma di Trento: Trento, Italia, 1999; Volume I and Volume II, p. 484. (In Italian) [Google Scholar]
- Campani, E. La cerniera dei Bivalvi per scopi diagnostici: Un esempio. In Proceedings of the Atti del Secondo Convegno Malacologico Pontino, Sabaudia, Italy, 20 September 2008. [Google Scholar]
- POLiKEY. Version 2. Available online: http://www.environment.gov.au/biodiversity/abrs/online-resources/polikey/index.html (accessed on 12 June 2019).
- Karamagn, S. Anfipodi delle acque dolci italiane. In Fauna d’Italia, XXXI; Calderini: Bologna, Italy, 1993; p. 333. [Google Scholar]
- Sansoni, G. Atlante Per il Riconoscimento dei Macroinvertebrati Bentonici dei Corsi d’acqua Italiani; Provincia Autonoma di Trento: Trento, Italy, 1988; p. 191. (In Italian) [Google Scholar]
- Sconfinetti, R. Chiave di riconoscimento visuale dei più comuni peracaridi (Crustacea, Peracarida) lagunari italiani. Studi Trent. Sci. Nat. Acta Biol. 2004, 81, 79–89. [Google Scholar]
- Leray, M.; Yang, J.Y.; Meyer, C.P.; Mills, S.C.; Agudelo, N.; Ranwez, V.; Bohem, J.T.; Machida, R.J. A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity: Application for characterizing coral reef fish gut contents. Front. Zool. 2013, 10, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawlowski, J.; Kelly-Quinn, M.; Altermatt, F.; Apothéloz-Perret-Gentil, L.; Beja, P.; Boggero, A.; Borja, A.; Bouchez, A.; Cordier, T.; Domaizon, I.; et al. The future of biotic indices in the ecogenomic era: Integrating (e)DNA metabarcoding in biological assessment of aquatic ecosystems. Sci. Total Environ. 2018, 637–638, 1295–1310. [Google Scholar] [CrossRef] [PubMed]
- Hering, D.; Borja, A.; Jones, J.I.; Pont, D.; Boets, P.; Bouchez, A.; Bruce, K.; Drakare, S.; Hänfling, B.; Kahlert, M.; et al. Implementation options for DNA-based identification into ecological status assessment under the European Water Framework Directive. Water Res. 2018, 138, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Di Sabatino, A.; Cristiano, G.; Pinna, M.; Lombardo, P.; Miccoli, F.P.; Marini, G.; Vignini, P.; Cicolani, B. Structure, functional organization and biological traits of macroinvertebrate assemblages from leaf-bags and benthic samples in a third-order stream of Central Apennines (Italy). Ecol. Indic. 2014, 46, 84–91. [Google Scholar] [CrossRef]
- Basset, A.; Barbone, E.; Rosati, I.; Vignes, F.; Breber, P.; Specchiulli, A.; D’Adamo, R.; Renzi, M.; Focardi, S.; Ungaro, N.; et al. Resistance and resilience of ecosystem descriptors and properties to dystrophic events: A study case in a Mediterranean lagoon. Transit. Waters Bull. 2013, 7, 1–22. [Google Scholar]
- Barbone, E.; Rosati, I.; Pinna, M.; Basset, A. Taxonomic and dimensional structure of benthic macroinvertebrate guilds in the Margherita di Savoia Salt Pans (Italy). Transit. Waters Bull. 2007, 1, 21–31. [Google Scholar]
- Ponti, M.; Vadrucci, M.R.; Orfanidis, S.; Pinna, M. Biotic indices for ecological status of transitional water ecosystems. Transit. Waters Bull. 2009, 3, 32–90. [Google Scholar]
- Elbrecht, V.; Vamos, E.E.; Meissner, K.; Ariviita, J.; Leese, F. Assessing strengths and weaknesses of DNA metabarcoding-based macroinvertebrate identification for routine stream monitoring. Methods Ecol. Evol. 2017, 8, 1265–1275. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Phylum | Number of Species | Number of Species without Barcode | Percentage of the Gap |
|---|---|---|---|
| Mollusca | 438 | 174 | 39.7 |
| Annelida | 423 | 196 | 46.3 |
| Arthropoda | 383 | 135 | 35.2 |
| Bryozoa | 87 | 47 | 54.0 |
| Echinodermata | 56 | 22 | 39.3 |
| Cnidaria | 54 | 23 | 42.6 |
| Porifera | 37 | 21 | 56.7 |
| Chordata | 24 | 3 | 12.5 |
| Nematoda | 9 | 6 | 66.7 |
| Platyhelminthes | 7 | 3 | 42.8 |
| Sipuncula | 7 | 3 | 42.8 |
| Entoprocta | 5 | 3 | 60 |
| Nemertea | 3 | 0 | 0 |
| Phoronida | 3 | 0 | 0 |
| Hemichordata | 1 | 0 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Specchia, V.; Tzafesta, E.; Marini, G.; Scarcella, S.; D’Attis, S.; Pinna, M. Gap Analysis for DNA Barcode Reference Libraries for Aquatic Macroinvertebrate Species in the Apulia Region (Southeast of Italy). J. Mar. Sci. Eng. 2020, 8, 538. https://doi.org/10.3390/jmse8070538
Specchia V, Tzafesta E, Marini G, Scarcella S, D’Attis S, Pinna M. Gap Analysis for DNA Barcode Reference Libraries for Aquatic Macroinvertebrate Species in the Apulia Region (Southeast of Italy). Journal of Marine Science and Engineering. 2020; 8(7):538. https://doi.org/10.3390/jmse8070538
Chicago/Turabian StyleSpecchia, Valeria, Eftychia Tzafesta, Gabriele Marini, Salvatore Scarcella, Simona D’Attis, and Maurizio Pinna. 2020. "Gap Analysis for DNA Barcode Reference Libraries for Aquatic Macroinvertebrate Species in the Apulia Region (Southeast of Italy)" Journal of Marine Science and Engineering 8, no. 7: 538. https://doi.org/10.3390/jmse8070538