
Abstract
The vertical distribution of phytoplankton primary production (PP) and chlorophyll a (Chl) was studied based on the data carried out in August–September 2015, 2017, and 2018. The PP maximum was located at the surface or within the 0–5 m subsurface layer. The subsurface chlorophyll maximum (SCM) was recorded at 39% stations on the outer shelf and in the vicinity of the continental slope. The SCM was not detected along the northward transect (130° E) from the Lena River delta. As in other areas of the World Ocean, the SCM was located below the upper mixed layer (UML), in the nitracline, near the boundary of the euphotic zone (1% of photosynthetically active radiation). Generally, the SCM was not accompanied by an additional PP maximum. The Chl concentration at the SCM did not exceed 1 mg m–3. PP produced within the UML and SCM contributed 72 and 23%, respectively, to the integrated primary production (IPP) of the water column. Our results suggest that the influence of the SCM on IPP was insufficient due to low Chl concentration and PP colimitation by the low light and temperature at these depths.
Similar content being viewed by others
INTRODUCTION
Optical remote-sensing data obtained from satellite-based color scanners pertain only to a comparatively narrow near-surface layer in the ocean. The value of the water-leaving reflectance formed in this layer does not take into account the features of the vertical distribution of chlorophyll a (Chl a), in particular, the subsurface Chl maximum (SCM). At the same time, to calculate the values of the integrated primary production (IPP) of the water column using algorithms based on satellite data, it is necessary to establish relationships between the surface and integrated values of production parameters that can be affected by the SCM.
The vertical Chl a distribution is applied in vertical-resolution primary production (PP) models. The shape of this distribution curve depends on the trophic state of water if this parameter is accepted as the Chl a concentration at the surface (Chl0) [45, 64]. The role of the SCM in IPP and regulation of upward nutrient fluxes is considered in scenarios of further climatic change in the Arctic that may cause increasing of the pycnocline gradients and decreasing of nutrient flux to the euphotic zone [59].
The presence of the SCM in Arctic seas is generally considered as typically of summer–autumn feature [4, 11, 12, 15–17, 22, 23, 28, 39–43, 62]. The formation of the SCM is a consequence of elevated phytoplankton biomass at these depths and/or an increasing cell-specific Chl a concentration as adaptation to low light conditions [18]. At present, the contribution of the SCM to IPP when computed using production models is a matter of debate. In addressing this problem, the effect of the SCM on the annual IPP estimates in the Arctic Ocean is discussed [5, 6, 29, 49, 54].
It should be noted that the SCM studies in the Arctic Ocean were mainly conducted in the so-called Case I waters, where the optical properties are formed by autochthonous organic matter, mainly, phytoplankton [25, 32]. Far fewer studies were conducted in the Siberian Arctic seas, subjected to the influence of river runoff, which governs the high content of dissolved and particulate organic and mineral matter and, low water transparency, and the shallow depth of the euphotic zone. The first studies on this problem demonstrated that the SCM in the Kara Sea is weakly pronounced in autumn and its contribution to IPP is from 1 to 27% [21]. In summer, the contribution of PP formed in the SCM was higher, averaging 53%. Nevertheless, it should be noted that its effect was local and restricted to the southwestern part of the sea [19].
A few studies on the vertical distribution of PP and Chl a in the Laptev Sea were mainly conducted in the eastern part of the area [27, 34, 56, 63] or in a deep-water region beyond the continental slope [4, 10]. Gaps in knowledge of the vertical distribution of PP parameters in the Laptev Sea are also indicated by the fact that the work [7] presents averaged Chl a vertical profiles for calculation of the annual IPP only for two stations. Moreover, no stations in the Laptev Sea were reported in the work [5] in which the SCM was studied in detail in regional and seasonal aspects and an estimate was made of its contribution to the annual IPP of the Arctic Ocean.
Thus, studies of the vertical variability of phytoplankton production parameters in the Laptev Sea conducted during three years are actually important. In view of the above problems the main aims of this study are as follows: (1) determination of the types of the vertical distribution of Chl a and PP in regions subjected to the effect of the river runoff and in the continental slope of the Laptev Sea and (2) estimation of the SCM contribution to the water column PP.
MATERIALS AND METHODS
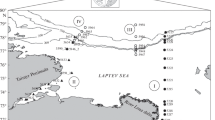
Study area and sampling procedure. The studies were conducted on a transect along 130° E northward from the Lena River delta (hereinafter, Lena transect, study region I in Fig. 1) in September 2015, along a transect from the Khatanga River estuary toward the open sea (hereinafter, Khatanga transect, study region II in Fig. 1) in September 2017 and along transects across the continental slope in the Laptev Sea (hereinafter, Eastern slope and Western slope transects, study regions III and IV in Fig. 1) in August–September 2018. Studies were conducted during cruises 63, 69 and 72 of the R/V Akademik Mstislav Keldysh, respectively.

Location of stations in the Laptev Sea. Circles denote cruise 63 of R/V Akademik Mstislav Keldysh (September 2015). Triangles denote cruise 69 of R/V Akademik Mstislav Keldysh (September 2017). Squares denote cruise 72 of R/V Akademik Mstislav Keldysh (August–September 2018). Dark symbols mark stations where SCM was observed. Roman numerals indicate: I, Lena transect; II, Khatanga transect; III, Eastern slope transect; IV, Western slope transect.
Water samples were taken using a Carousel Water Sampler equipped with Niskin bottles from six to nine depths in the upper 100-m layer after preliminary temperature, conductivity, and chlorophyll fluorescence measurements with a CTD probe (SBE-19 and SBE-32, Seabird Electronics). Samples from the surface layer were collected with a plastic bucket as the bottles were being closed near the surface.
PP and Chlaconcentration measurements. PP was estimated using a radiocarbon modification of the light and dark bottle technique [57]. Experiments were conducted according to a simulated in situ method [37, 58] or according to the Ryther and Yentsch approach [51] with modifications [1, 19]. Chl a concentrations was measured with a fluorometric technique [30]. Chl a and pheophytin a concentrations were calculated according to [31]. The methods of PP and Chl a determination were described in detail in -the previous work based on the data of the above-mentioned Laptev Sea expeditions [2].
Determination of incident and underwater irradiance and nutrients. The intensity of incident and underwater photosynthetically active radiation (PAR) was measured with LI-190 and LI-192 sensors (LI-COR, USA). Measurements of incident PAR were made during light day; underwater PAR was measured by probing to depths of ∼60–80 m and to the bottom at shallow stations.
Samples for determining forms of nitrogen were treated immediately after sampling. The nitrite (NO2) and nitrate (NO3) contents were determined according to [26]. Dissolved carbon dioxide and various forms of dissolved inorganic carbon were calculated by the pH-Alk method using thermodynamic equations for the carbon balance with Roy’s constants for carbonic acid dissociation [26, 44] with corrections [38].
Determination of SCM and boundaries of the upper mixed layer and nitracline. We define the SCM as a layer below the pycnocline where the CTD-mounted fluorometer registered increased values compared with overlying and underlying layers [12]. The upper and lower SCM boundaries were determined based on the assumption that the vertical Chl a distribution is described by a Gaussian curve [48]. Then, the SCM thickness (σ) was calculated according to the formula
where Chlint is the integrated Chl a value within a water column and Chlmax is the maximum Chl a concentration. Note that we considered the Chlmax value as significantly exceeding the value on the surface (Chl0), when Chlmax /Chl0 ≥ 1.15 [64].
The depth where the water density (σt) first exceeded the surface chlorophyll value by 0.3 kg/m3 was considered the lower boundary of the UML [60]. The upper boundary of the nitracline consisted of the depths below which the sum of (NO2 + NO3) started to increase.
Stability of the water column was estimated according to the Brent–Väisälä frequency
where g is gravitational acceleration, taken equal to 9.81 m/s2 at the latitude of the Laptev Sea, and ρ is the water density at depth z.
RESULTS
Vertical Chladistribution along transects. Along the Lena transect, the Chl a concentration in different layers varied by more than two orders of magnitude, from 0.01 to 1.39 mg/m3 (Fig. 2а). The maximum values were recorded in the surface water. The Chl a concentration varied from 50 to 100%, on average 81%, in the euphotic layer with the lower boundary at depths from 10 to 31 m (Fig. 2a).

Vertical Chl a distribution along Lena (a) and Khatanga (b) transects. Нeu is euphotic layer.
A similar scale of Chl variability was recorded along the Khatanga transect. The Chl concentration varied by more than two orders of magnitude and ranged from 0.02 to 2.78 mg/m3 (Fig. 2b). The near-bottom Chl a maximum was recorded at stations in the southern part of the Khatanga River estuary. On the adjacent shelf (<50 m), the chlorophyll maximum was recorded on the surface or within the UML. The SCM was formed on the open shelf (50–200 m) and in the vicinity of the continental slope (Fig. 1) at depths of 15–30 m. On average, 69% of Chl a (from 42 to 100%) was recorded within the euphotic layer, the lower boundary of which was at depths of 14–32 m.
Along the Eastern slope transect, the range of Chl a variability was lower compared to the first two transects and the maximum values did not exceed 1 mg/m3. The Chl a concentration varied by a factor of 70, from 0.01 to 0.70 mg/m3. The SCM was recorded at all stations; except for station 5954, 67–93% (on average 80%) of Chl a was observed in the euphotic layer with a depth of 39–65 m (Fig. 3а).

Vertical Chl a distribution along Eastern (a) and Western slope (b) transects. Нeu is euphotic layer.
Even smaller range of Chl a variability was recorded along the Western slope transect. The ratio of maximum to minimum values was equal to 59; Chl a concentrations in different layers ranged from 0.01 to 0.59 mg/m3. The SCM was a specific feature of the vertical Chl a distribution along the Eastern slope transect; it was absent only at two stations (stations 5945_2 and 5965) (Fig. 1). From 77 to 92% (on average 83%) of Chl a was recorded in the euphotic layer (Fig. 3b).
Vertical distribution of PP, typical vertical profiles of Chla, and characteristics of the SCM. Along the Lena transect, the maximum PP values were recorded on the surface or in the subsurface layer (0–5 m) (Fig. 4). PP gradually decreased with depth and did not form additional maxima. The boundary of the euphotic layer was, as a rule, between 1 and 0.1% levels of the subsurface PAR. The vertical Chl a profiles can be subdivided into four types. The first type was characterized by a uniform Chl distribution in the UML and a sharp decrease below this layer (Fig. 4a). The second type was characterized by an exponential decrease in Chl a values from the surface (Fig. 4b). The third type had the maximum within the UML (Fig. 4c). The fourth type was distinguished by a gradual decrease in Chl a values in the UML and in the layer of maximum pycnocline gradients, and by their sharp decrease below these layers (Fig. 4d). The absence of the SCM was a specific feature to all types.

Types of vertical profiles of PP (PP, mgС m–3 day–1), Chl а (Chl, mg/m3), sum of nitrites and nitrates (NO2 + NO3, µМ), and water density (σt, kg/m3) along the Lena transect. Horizontal lines mark depths with 10, 1 and 0.1% PAR.
As in the above-mentioned region, PP along the Khatanga transect formed the maximum on the surface or within UML (Fig. 5). PP values decreased with depth. The boundary of the euphotic layer was either close to the horizon with a 1% PAR level or was deeper than 0.1% subsurface PAR (station 5635). Three types of the vertical Chl a distribution were distinguished along this transect. The first was characterized by an increase in Chl a to the bottom (Fig. 5a). The inherent property of the second type was a maximum within well-lit UML (>10% of subsurface PAR) (Fig. 5b), which cannot be classified as the SCM based on the above criteria. Finally, the third type of vertical Chl a distribution was characterized by the SCM in which the layer with Chlmax was located below the UML, close to nitracline and 1% subsurface PAR level (Fig. 5c).

Types of vertical profiles of PP (PP, mgС m–3 day–1), Chl а (Chl, mg/m3), sum of nitrites and nitrates (NO2 + NO3, µМ), and water density (σt, kg/m3) along the Khatanga transect. Horizontal lines mark depths with 10, 1, and 0.1% PAR.
Two PP peaks were formed along the Eastern slope transect. One of them was observed close to the surface and the second one was recorded in the SCM (Fig. 6a). In other cases PP values decreased with depth without the formation of secondary maxima (Figs. 6b, 6c). The boundary of the euphotic layer was located between 1 and 0.1% of subsurface PAR or was close to 0.1% level. The SCM was observed at most stations of the Eastern slope transect. It was registered in the nitracline, in the well-lit zone (close to 10% of PAR) (Fig. 6a) or located deeper close to the boundary of the euphotic zone (1% PAR). The SCM was not recorded only at one station of the transect (station 5954) (Fig. 6c). At this station, the vertical Chl a distribution within the UML and pycnocline layer was close to uniform; its values decreased below these layers.

Types of vertical profiles of PP (PP, mgС m–3 day–1), Chl а (Chl, mg/m3), sum of nitrites and nitrates (NO2 + NO3, µМ), and water density (σt, kg/m3) along the Eastern slope transect. Horizontal lines mark depths with 10, 1, and 0.1% PAR.
PP profiles had one maximum on the surface at most stations of the Western slope transect (Fig. 7). An increase in PP in the SCM was recorded only at station 5964 (Fig. 7c). The boundary of the euphotic layer was at depths between the 1 and 0.1% subsurface PAR. The Chl a concentrations at stations without a pronounced SCM exponentially decreased with depth (Fig. 7a) or formed more a complex profile that can be described by a curve looks like sine wave (Fig. 7d). At stations with a well-pronounced SCM, it was located like in other regions in the nitracline between the 1 and 0.1% PAR level (Figs. 7b, 7c).

Types of vertical profiles of PP (PP, mgС m–3 day–1), Chl а (Chl, mg/m3), sum of nitrites and nitrates (NO2 + NO3, µМ), and water density (σt, kg/m3) along the Eastern slope transect. Horizontal lines mark depths with 10, 1, and 0.1% PAR.
The SCM was recorded at 39% of stations in the Laptev Sea in August–September; its thickness was 9–19 m. Chlmax was recorded at depths from 10 to 38 m (Table 1). The maximum Chl a values in the SCM varied by a factor of 2.9, from 0.34 to 0.98 mg/m3. The degree of SCM manifestation can be determined by the Chlmax/Chl0 ratio. This ratio varied from 1.19 to 4.38, its average value was 2.1 (Table 1). PP values in the Chlmax layer (PPSCM) never exceeded surface and subsurface values (PPmax). The PPmax/PPscm ratio varied from 1.5 to 100 and its average value was 15.8. The pheophytin a content in the Chlmax layer varied from 7 to 66% of the sum of Chl a and pheophytin a (41% on average).
In August–September, the SCM in the Laptev Sea was located, as a rule, in the layer of maximum pycnocline gradients with negative or close-to-negative water temperature (Table 2). The NO2 + NO3 content in the Chlmax layer was usually above values limiting the growth and photosynthesis of phytoplankton [61]. The exceptions were stations 5590_2 and 5634 along the Khatanga transect and stations 5949 and 5952 along the Eastern slope transect (Table 2), where the SCM was recorded immediately above the upper boundary of the nitracline. Absolute radiation values (Iz) at the depth of Chlmax varied from 0.03 to 3.22 Ein m–2 day–1, which consistent with range from 0.88 to 21.02% of the subsurface PAR.
DISCUSSION
In the Arctic Ocean the SCM is usually formed in summer–autumn after the bloom under conditions of low nutrients in the UML. As in the entire World Ocean, the SCM is formed in the nitracline near the boundary of the euphotic zone (1% subsurface PAR) [18]. Thus, phytoplankton develops within water layers under favorable nutrient supply and light conditions for photosynthesis [41].
The Chl a profile is used in vertical-resolved PP and biogeochemical models [33, 50, 54]. One approach assumes that the Chl a concentration is uniformly distributed in the UML and decreases exponentially with depth below this layer [46]. Another approach assumes seasonal variations in the vertical Chl a distribution. In spring, its distribution is uniform, and for summer and autumn, the above assumption is accepted [29]. The point of view that the SCM is widespread throughout the Arctic Ocean leads to the need to take into account its contribution to IPP in biogeochemical models [33, 50, 54].
In the Laptev Sea, the SCM was recorded at the northernmost stations of the transect from the Lena River delta to the open sea in autumn 1991 [27] and beyond the continental slope in deep-water areas [4, 10]. In the present study, we analyzed for the first time the results of a study on the vertical Chl a distribution in nearly all Laptev Sea areas. The SCM was not recorded on the transect along 130° E northward from the Lena River delta, surveyed in September 2015. The NO2 + NO3 concentration in the UML exceeded the limiting values at nearly all stations of that transect [2, 3]. In September 2017, along the Khatanga transect, the SCM was not pronounced in the river estuary and on the adjacent shelf, in the zone desalinated by river water (S < 28 PSU). Unfavorable SCM formation conditions were created in this region due to low water transparency caused by high concentrations of dissolved and particulate organic matter. On the contrary, the conditions were favorable for SCM formation along the transects across the continental slope (study regions III and IV) (Fig. 1). NO2 + NO3 values within the Chlmax layer did not, as a rule, exceed the limiting values (Table 2). The SCM was formed in layers where the stability of water column estimated by the Brent–Väisälä frequency was, on average, 2.7 times higher than its average value in the UML (\(\bar {N}\)2) (Table 2). At three stations of the Eastern slope transect (stations 5950, 5952, and 5956), this value was from one to two orders of magnitude higher. The exceptions were stations 5958, 5962, and 5963, where N2 was lower than \(\bar {N}\)2. The absolute PAR values at the Chlmax depth were higher than the compensation light intensity for photosynthesis [61] (Table 2). In addition, it should be noted that the phytoplankton population in the SCM contains cells well-adapted to low irradiance [41]. It should also be mentioned that algae at the Chlmax depth were in a good physiological state, which can be estimated according to the pheophytin a content (on average 41% of the sum of Chl a and pheophytin a).
It is accepted to divide IPP models to depth-integrated and depth-resolved [8, 13, 14, 24, 35, 52, 53]. It was demonstrated earlier that using of Chl a vertical distribution in production models insignificantly improves the results of IPP calculations [9, 20, 52], and omission of the SCM is a minor error on a large spatiotemporal scale [5, 6]. Other researchers believe that accounting of the SCM increase nearly twofold the annual IPP value on the scale of the Arctic Ocean [29]. In any case, the SCM is an important feature of the vertical Chl a distribution in seasons when the water column is sharply stratified, nutrients limit PP in UML, and the contribution of the SCM to IPP may be significant [41].
It is interesting to estimate the contribution of PP in different layers to IPP in the Laptev Sea where such studies were not conducted. The data presented in Table 3 demonstrate that in the water layer forming the signal recorded by ocean color scanners (1/Kd) defined as penetration depth, (Hpd) where Kd is the diffuse attenuation coefficient of the downwelling PAR, IPP ranges from 21 to 79% (on average 45%), the contribution of the UML to IPP is 38–100% (on average 72%), and the contribution of the SCM to IPP varies from 8 to 56% (on average 23%). Therefore, near-surface layers in the Laptev Sea were the main contributors to IPP at the end of summer and at the beginning of autumn. The same results were obtained in the Kara Sea in autumn [21], where environmental conditions are similar to the Laptev Sea. Our results are also similar to estimates of the SCM contribution to IPP obtained in September for the Baffin Bay (5.1–15.8%), the Beaufort Sea (20.4%) and Greenland Sea (16.6%) [6]. It should be mentioned that the contribution of the SCM to IPP in the Kara Sea in midsummer may be considerable in some regions, reaching 95% [19]. High PP values in the SCM (from 43 to 76%, on average 62%) were also recorded in the Canadian Arctic [41].
Our findings suggest that the contribution of SCM to IPP was on average insignificant in the Laptev Sea in August–September. The maximum PP was observed at the surface or within near-surface layers, and the SCM, as a rule, was not accompanied by a secondary PPmaximum. These results somewhat contradict the notions based on the data obtained in other regions of the Arctic Ocean where the SCM was accompanied by a PP maximum [17, 28, 40]. It should be also noted that the contribution of the SCM to IPP exceeded 20% only in cases when it was located rather high in the water column (horizons with Chlmax ≤ 20 m), as a rule, within well-lit water layers (Table 3). The NO2 + NO3 content at these depths could be both higher and lower than the limiting values (Table 2). T values (from –1.52 to 2.28°C) were also different. In earlier studies, it was concluded that the concentration of nitrates in the SCM is not a factor limiting PP [41]. This conclusion was based on the fact that, first, the level of light saturation for nitrate uptake in the SCM is lower than for carbon assimilation and, second, there is no relationship between the carbon assimilation rate and NO3 concentration. The light level in the SCM was lower than light saturation. Thus, light may be the major factor limiting PP in the SCM [41]. Another reason for PP limitation at the depths of SCM is apparently low water temperature. It is known that, on one hand, the temperature optimum for phytoplankton growth is above 10°C [36], and on the other hand, adaptation of photosynthetic parameters to low temperatures in the Arctic has not been found [47, 55].
CONCLUSIONS
Studies on the vertical variability of the PP and Chl a conducted in the Laptev Sea at the end of August and at the beginning of September have shown various shapes of its curves. The PP distribution with respect to depth was more uniform with the maximum on the surface or close to it. A wide occurrence of the SCM in the vertical Chl a distribution was not detected. SCM was observed only on the open shelf (>50 m) and in vicinity of the continental slope. The PP was formed mainly in the UML. The contribution of the SCM to IPP did not exceed 20% at most stations. A more considerable contribution (to 56%) to IPP was observed when the SCM was located within the upper well-lit layers of the euphotic zone.
REFERENCES
V. I. Vedernikov, “Specific distribution of primary production and chlorophyll in the Black Sea in spring and summer,” in Variability of the Black Sea Ecosystem: Natural and Anthropogenic Factors, Ed. by M. E. Vinogradov (Nauka, Moscow, 1991), pp. 128–147.
A. B. Demidov, V. I. Gagarin, E. G. Arashkevich, et al., “Spatial variability of primary production and chlorophyll in the Laptev Sea in August–September,” Oceanology (Engl. Transl.) 59, 678–691 (2019).
I. N. Sukhanova, M. V. Flint, E. Ju. Georgieva, et al., “The structure of phytoplankton communities in the eastern part of the Laptev Sea,” Oceanology (Engl. Transl.) 57, 75–90 (2017).
S. H. Ahn, T. Whitledge, D. A. Stockwell, et al., “The biochemical composition of phytoplankton in the Laptev and East Siberian seas during the summer of 2013,” Polar Biol. 42 (1), 133–148 (2019).
M. Ardyna, M. Babin, M. Gosselin, et al., “Parameterization of vertical chlorophyll a in the Arctic Ocean: impact of the subsurface chlorophyll maximum on regional, seasonal and annual primary production estimates,” Biogeosciences 10 (3), 1345–1399 (2013).
K. R. Arrigo, P. A. Matrai, and G. L. van Dijken, “Primary productivity in the Arctic Ocean: Impacts of complex optical properties and subsurface chlorophyll maxima on large-scale estimates,” J. Geophys. Res.: Oceans 116, C11022 (2011). https://doi.org/10.1029/2011JC007273
K. R. Arrigo and G. L. van Dijken, “Secular trends in Arctic Ocean net primary production,” J. Geophys. Res.: Oceans 116, C09011 (2011). https://doi.org/10.1029/2011JC007151
M. Babin, S. Bélanger, I. Ellingsen, et al., “Estimating of primary production in the Arctic Ocean using ocean color remote sensing and coupled physical-biological models: strengths, limitations and how they compare,” Progr. Oceanogr. 139, 197–220 (2015).
M. J. Behrenfeld and P. G. Falkowski, “A consumer’s guide to phytoplankton primary productivity models,” Limnol. Oceanogr. 42, 1479–1491 (1997).
P. S. Bhavya, J. H. Lee, H. W. Lee, et al., “First in situ estimations of small phytoplankton carbon and nitrogen uptake rates in the Kara, Laptev and East Siberian seas,” Biogeosciences 15 (18), 5503–5517 (2018).
B. C. Booth, P. Larouche, S. Bélanger, et al., “Dynamics of Chaetoceros socialis in the north water,” Deep Sea Res., Part II 49 (22–23), 5003–5025 (2002).
Z. W. Brown, K. E. Lowry, M. A. Palmer, G. L. van Dijken, M. M. Mills, R. S. Pickart, and K. R. Arrigo, “Characterizing the subsurface chlorophyll a maximum in the Chukchi Sea and Canada Basin,” Deep Sea Res., Part II 118, 88–104 (2015).
J. Campbell, D. Antoine, R. Armstrong, et al., “Comparison of algorithms for estimating ocean primary production from surface chlorophyll, temperature, and irradiance,” Global Biogeochem. Cycles 16, (2002). https://doi.org/10.1029/2001GB001444
M.-E. Carr, M. A. M. Friedrichs, M. Schmeltz, et al., “A comparison of global estimates of marine primary production from ocean color,” Deep Sea Res., Part II 53, 741–770 (2006).
E. C. Carmack, R. W. Macdonald, and S. Jasper, “Phytoplankton productivity on the Canadian Shelf of the Beaufort Sea,” Mar. Ecol.: Progr. Ser. 277, 37–50 (2004).
A. Cherkasheva, E. M. Nöthig, E. Bauerfeind, C. Melsheimer, and A. Bracher, “From the chlorophyll-a in the surface layer to its vertical profile: a Greenland Sea relationship for satellite applications,” Ocean Sci. 9, 431–445 (2013).
G. F. Cota, L. R. Pomeroy, W. G. Harrison, et al., “Nutrients, primary production and microbial heterotrophy in the southeastern Chukchi Sea: Arctic summer nutrient depletion and heterotrophy,” Mar. Ecol.: Progr. Ser. 135, 247–258 (1996).
J. J. Cullen, “Subsurface chlorophyll maximum layers: enduring enigma or mystery solved?” Annu. Rev. Mar. Sci. 7, 207–239 (2015).
A. B. Demidov, V. I. Gagarin, O. V. Vorobieva, et al., “Spatial and vertical variability of primary production in the Kara Sea in July and August 2016: the influence of the river plume and subsurface chlorophyll maxima,” Polar Biol. 41 (3), 563–578 (2018).
A. B. Demidov, O. V. Kopelevich, S. A. Mosharov, et al., “Modeling Kara Sea phytoplankton primary production: development and skill assessment of regional algorithms,” J. Sea Res. 125, 1–17 (2017).
A. B. Demidov, S. A. Mosharov, and P. N. Makkaveev, “Patterns of the Kara Sea primary production in autumn: biotic and abiotic forcing of subsurface layer,” J. Mar. Sys. 132, 130–149 (2014).
S. R. Erga, N. Ssebionga, B. Hamre, et al., “Environmental control of phytoplankton distribution and photosynthetic performance at the Jan Mayen Front in the Norwegian Sea,” J. Mar. Sys. 130, 193–205 (2014).
J. Ferland, M. Gosselin, and M. Starr, “Environmental control of summer primary production in the Hudson Bay system: the role of stratification,” J. Mar. Sys. 88, 385–400 (2011).
M. A. M. Friedrichs, M.-E. Carr, R. Barber, et al., “Assessing the uncertainties of model estimates of primary productivity in the tropical Pacific Ocean,” J. Mar. Sys. 76, 113–133 (2009).
H. G. Gordon and A. Morel, Remote Assessment of Ocean Color for Interpretation of Satellite Visible Imagery: A Review (Springer-Verlag, New York, 1983).
H. P. Hansen and F. Koroleff, “Determination of nutrients, in Methods of Seawater Analysis, Ed. by K. Grashoff, (Wiley, Chichester, 1999), pp. 149–228.
A.-S. Heiskanen and A. Keck, “Distribution and sinking rates of phytoplankton, detritus and particulate biogenic silica in the Laptev Sea and Lena River (Arctic Siberia),” Mar. Chem. 53, 229–245 (1996).
V. Hill and G. Cota, “Spatial patterns of primary production on the shelf, slope and basin of the Western Arctic in 2002,” Deep Sea Res., Part II 57 (24–26), 3344–3354 (2005).
V. J. Hill, P. A. Matrai, E. Olson, et al., “Synthesis of integrated primary production in the Arctic Ocean: II. In situ and remotely sensed estimates,” Progr. Oceanogr. 110, 107–125 (2013).
O. Holm-Hansen, C. J. Lorenzen, R. W. Holmes, and J. D. H. Strickland, “Fluorometric determination of chlorophyll,” J. Cons. Perm. Int. Explor. Mer. 30, 3–15 (1965).
O. Holm-Hansen and B. Riemann, “Chlorophyll a determination: improvements in methodology,” Oikos 30, 438–447 (1978).
H. G. Jerlov, Optical Oceanography (Elsevier, New York, 1968).
M. Jin, C. Deal, S. H. Lee, et al., “Investigation of Arctic sea and ocean primary production for the period 1992–2007 using a 3-D global ice-ocean ecosystem model,” Deep Sea Res., Part II 81–84, 28–35 (2012).
K. V. Juterzenka and K. Knickmeier, “Chlorophyll a distribution in water column and sea ice during the Laptev Sea freeze-up study in autumn 1995,” in Land-Ocean Systems in the Siberian Arctic: Dynamics and History, Ed. by H. Kassens, (Springer, Berlin, 1999), pp. 153–160.
Y. J. Lee, P. A. Matrai, M. A. M. Friedrichs, et al., “An assessment of phytoplankton primary productivity in the Arctic Ocean from satellite ocean color/in situ chlorophyll-a based models,” J. Geophys. Res.: Oceans 120, (2015). https://doi.org/10.1002/2015/JC11018
W. W. Li, “Photosynthetic response to temperature of marine phytoplankton along a latitudinal gradient (16°N to 74°N),” Deep-Sea Res., Part A 32 (11), 1381–1391 (1985).
S. E. Lohrenz, “Estimation of primary production by the simulated in situ method,” ICES Mar. Sci. Symp. 197, 159–171 (1993).
P. N. Makkaveev, “The total alkalinity in the anoxic waters of the Black sea and in sea-river mixture zones,” in Proceedings of the Joint IOC-JGOFS CO2Advisory Panel Meeting (Intergovernmental Oceanographic Commission, UNESCO, Paris, 1998).
J. Martin, D. Dumont, and J.-E. Tremblay, “Contribution of subsurface chlorophyll maxima to primary production in the coastal Beaufort Sea (Canadian Arctic): a model assessment,” J. Geophys. Res. 118 (11), 5873–6318 (2013).
J. Martin, J.-E. Tremblay, J. Gagnon, et al., “Prevalence, structure and properties of subsurface chlorophyll maxima in Canadian Arctic waters,” Mar. Ecol.: Progr. Ser. 412, 69–84 (2010).
J. Martin, J.-E. Tremblay, and N. M. Price, “Nutritive and photosynthetic ecology of subsurface chlorophyll maxima in Canadian Arctic waters,” Biogeosciences 9 (12), 5353–5371 (2012).
K. I. Martini, P. J. Stabeno, and C. Ladd, “Dependence of subsurface chlorophyll on seasonal water masses in the Chukchi Sea,” J. Geophys. Res.: Oceans 121, 1755–1770 (2016). https://doi.org/10.1002/2015JC011359
F. A. McLaughlin and E. C. Carmack, “Deepening of the nutricline and chlorophyll maximum in the Canada Basin interior, 2003–2009,” Geophys. Res. Lett. 37, L24602 (2010). https://doi.org/10.1029/2010GL045459
F. J. Millero, “Thermodynamics of the carbon dioxide system in oceans,” Geochim. Cosmochim. Acta 59 (4), 661–677 (1995).
A. Morel and J.-F. Berthon, “Surface pigments, algal biomass profiles, and potential production of the euphotic layer: relationships reinvestigated in view of remote-sensing applications,” Limnol. Oceanogr. 34 (1), 1545–1562 (1989).
S. Pabi, G. L. van Dijken, and K. R. Arrigo, “Primary production in the Arctic Ocean, 1998–2006,” J. Geophys. Res.: Oceans 113, C08005 (2008). https://doi.org/10.1029/2007/JC004578
T. Platt, W. G. Harrison, B. Irwin, et al., “Photosynthesis and photoadaptation of marine phytoplankton in the Arctic,” Deep-Sea Res. 29 (10), 1159–1170 (1982).
T. Platt, S. Sathyendranath, O. Ulloa, et al., “Ocean primary production and available light: further algorithms for remote sensing,” Deep Sea Res., Part I 35, 855–879 (1988).
E. E. Popova, A. Yool, A. C. Coward, et al., “Control of primary production in the Arctic by nutrients and light: insights from a high resolution ocean general circulation model,” Biogeosciences 7 (11), 3569–3591 (2010).
E. E. Popova, A. Yool, A. C. Coward, et al., “What controls primary production in the Arctic Ocean? Results from an intercomparison of five general circulation models with biogeochemistry,” J. Geophys. Res.: Oceans 117, C00D12 (2012). https://doi.org/10.1029/2011JC007112
J. H. Ryther and C. S. Yentsch, “The estimation of phytoplankton production in the ocean from chlorophyll and light data,” Limnol. Oceanogr. 2, 281–286 (1957).
V. Saba, M. A. M. Friedrichs, D. Antoine, et al., “An evaluation of ocean color model estimates of marine primary productivity in coastal and pelagic regions across the globe,” Biogeosciences 8, 489–503 (2011).
V. Saba, S. Marjorie, M. A. M. Friedrichs, et al., “Challenges of modeling depth-integrated marine primary productivity over multiple decades: a case study at BATS and HOT,” Global Biogeochem. Cycles 24, GB3020 (2010). https://doi.org/10.1029/2009GB003655
V. Schourup-Kristensen, C. Wekerle, D. A. Wolf-Gladrow, and C. Völker, “Arctic Ocean biogeochemistry in the high resolution FESOM 1.4-REcoM2 model,” Progr. Oceanogr. 168, 65–81 (2018).
W. O. Smith and W. G. Harrison, “New production in polar regions: the role of environmental controls,” Deep-Sea Res. 38 (12), 1463–1479 (1991).
Yu. I. Sorokin and P. Yu. Sorokin, “Plankton and primary production in the Lena river estuary and in the south-eastern Laptev Sea,” Estuarine, Coastal Shelf Sci. 43, 399–418 (1996).
E. Steemann Nielsen, “The use of radioactive carbon (C14) for measuring organic production in the sea,” J. Cons. Perm. Ins. Explor. Mer. 18, 117–140 (1952).
E. Steemann Nielsen, “Experimental methods for measuring organic production in the sea,” Rapp. P.-v. Réun. Cons. perm. int. Explor. Mer. 144, 38–46 (1958).
N. S. Steiner, T. Sou, C. Deal, et al., “The future of the subsurface chlorophyll-a maximum in the Canada Basin—A model intercomparison,” J. Geophys. Res.: Oceans 121, 387–409 (2015). https://doi.org/10.1002/2015JC011232
M. L. Timmermans, S. Cole, and J. Toole, “Horizontal density structure and restratification of the Arctic Ocean surface layer,” J. Phys. Oceanogr. 42 (4), 659–668 (2012).
J.-É. Tremblay, C. Michel, and K. A. Hobson, “Bloom dynamics in early opening waters of the Arctic Ocean,” Limnol. Oceanogr. 51 (2), 900–912 (2006).
J.-É. Tremblay, K. Simpson, J. Martin, et al., “Vertical stability and the annual dynamics of nutrients and chlorophyll fluorescence in the coastal, southeast Beaufort Sea,” J. Geophys. Res.: Oceans 113, C07S90 (2008). https://doi.org/10.1029/2007JC004547
K. Tuschling, “Phytoplankton ecology in the arctic Laptev Sea—a comparison of three seasons,” Ber. Polarforschung. 347, (2000).
J. Uitz, H. Claustre, A. Morel, and S. B. Hooker, “Vertical distribution of phytoplankton communities in open ocean: an assessment on surface chlorophyll,” J. Geophys. Res.: Oceans 111, C08005 (2006). https://doi.org/10.1029/2005JC003207
Funding
This work was performed according to the State assignment of Ministry of Science and Higher Education of Russian Federation (no. 0149-2019-0008). Collection of phytoplankton data was supported by the Russian Foundation of Basic Research, project no. 18-05-60069. Collection of hydrophysical data was supported by the Russian Foundation for Basic Research, project no. 18-05-60302.
Author information
Authors and Affiliations
Corresponding author
Additional information
Translated by N. Ruban
Rights and permissions
About this article

Cite this article
Demidov, A.B., Gagarin, V.I., Artemiev, V.A. et al. Vertical Variability of Primary Production and Features of the Subsurface Chlorophyll Maximum in the Laptev Sea in August–September, 2015, 2017, and 2018. Oceanology 60, 189–204 (2020). https://doi.org/10.1134/S0001437020010063
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0001437020010063