Unveiling the Properties of Thai Stingless Bee Propolis via Diminishing Cell Wall-Associated Cryptococcal Melanin and Enhancing the Fungicidal Activity of Macrophages



Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Cells and Culture Conditions
2.2. Propolis Preparation
2.2.1. Extraction of Ethanolic Extract of SLB Propolis (EEP)
2.2.2. Total Phenolic Content
2.2.3. Total Flavonoid Content
2.2.4. High-Performance Liquid Chromatography (HPLC) Analysis
2.3. Effect of EEP on Chitin/Chitosan Production of C. neoformans
2.3.1. Chitin and Chitosan Synthesis
2.3.2. Detection of CDA1 mRNA by Real-Time Reverse Transcription-Polymerase Chain Reaction (rRT-PCR)
2.4. Effect of EEP on C. neoformans Melanization
2.4.1. Melanin Production
2.4.2. Laccase Activity
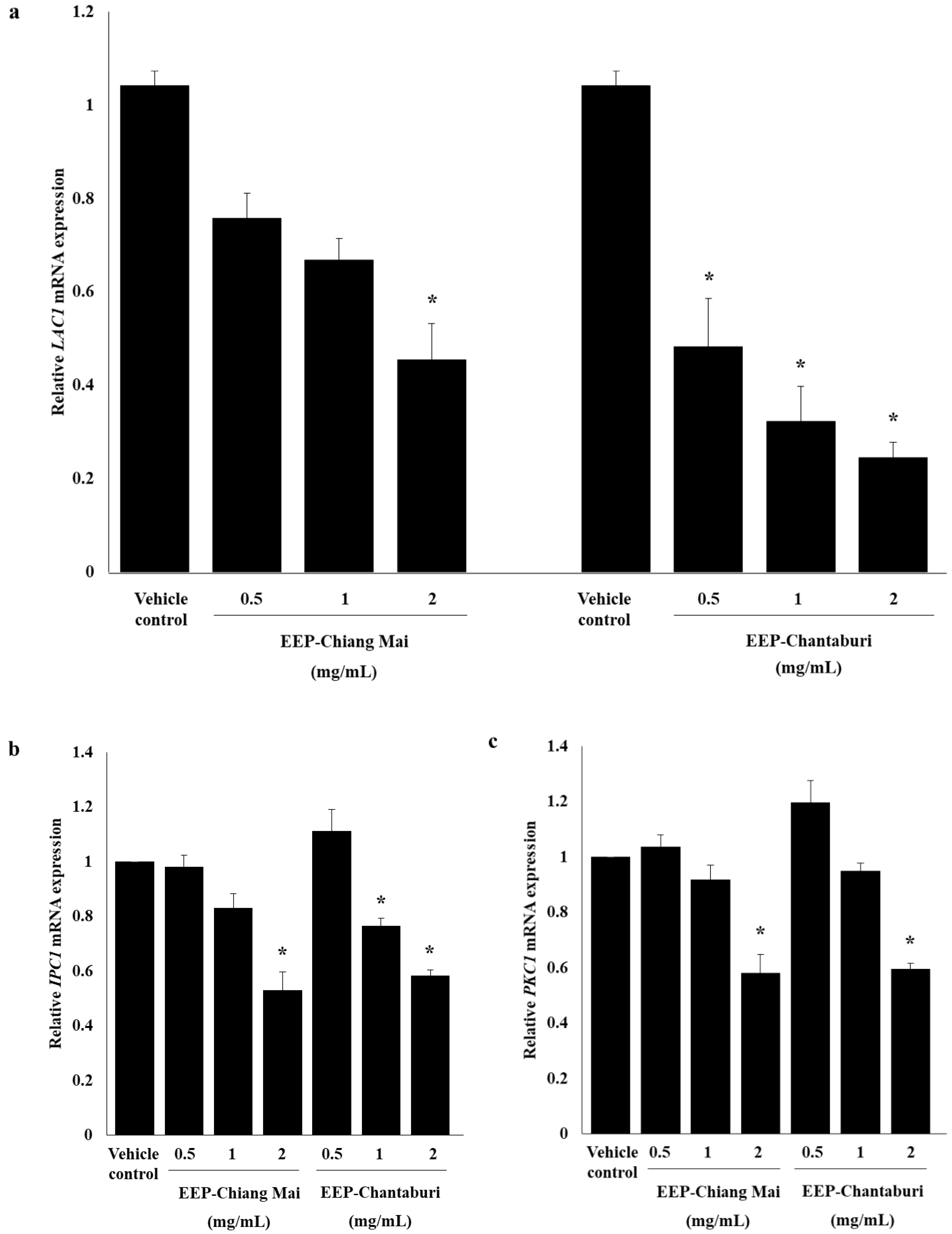
2.4.3. Detection of Melanin-Related Gene Expression by rRT-PCR
2.5. Macrophages Cultures
2.6. Immune Response with EEP-Treated C. neoformans
2.6.1. Macrophages Infected with EEP-Treated C. neoformans
2.6.2. Intracellular Cryptococcal Cell Replication
2.6.3. LysoTracker Staining
2.7. Statistical Analysis
3. Results and Discussion
3.1. Quantitation of Essential Compounds in SLB Propolis
3.2. Effect of EEP on Chitin/Chitosan Synthesis of C. neoformans
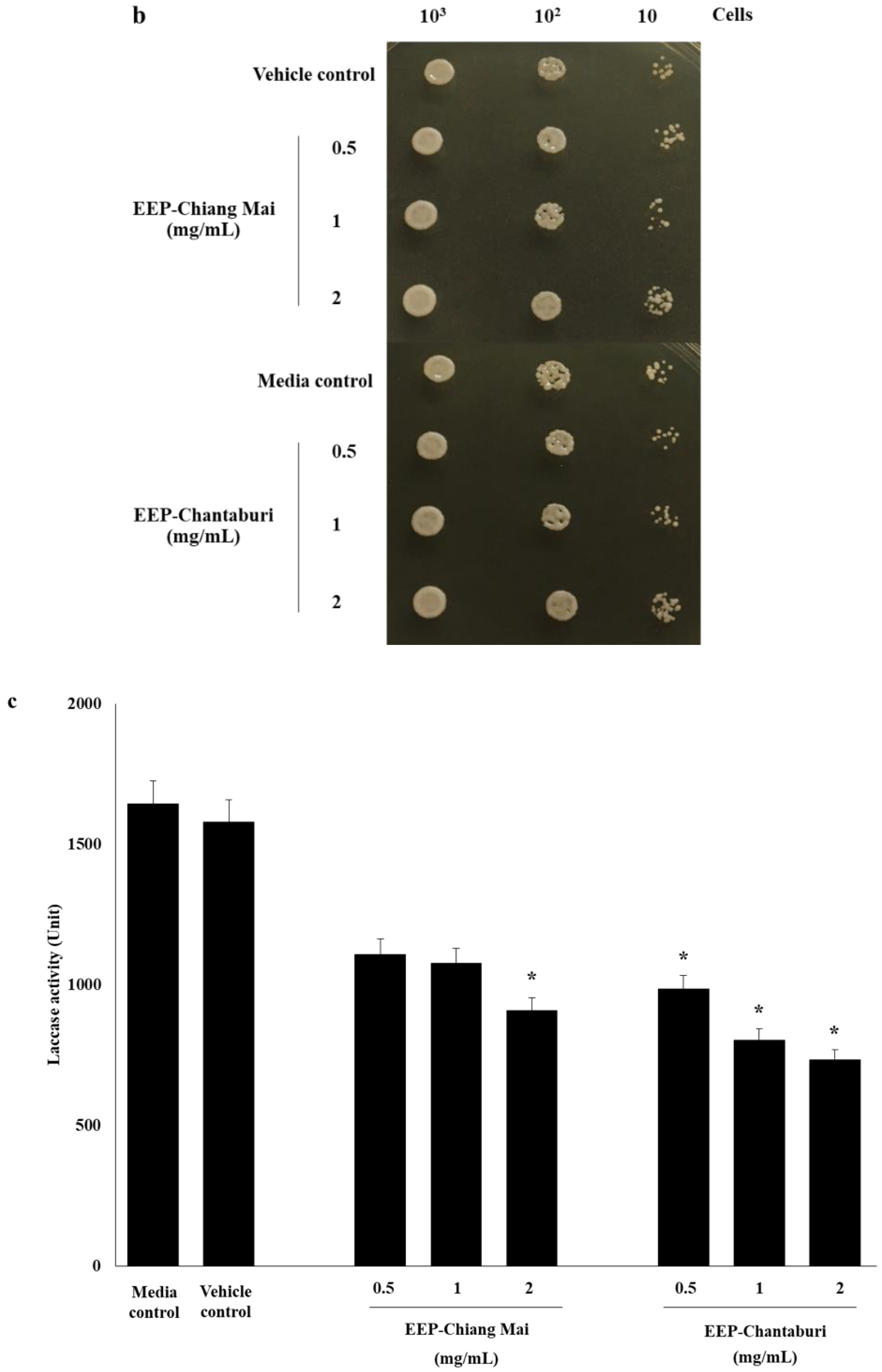
3.3. Anti-Melanization of EEP-Treated C. neoformans
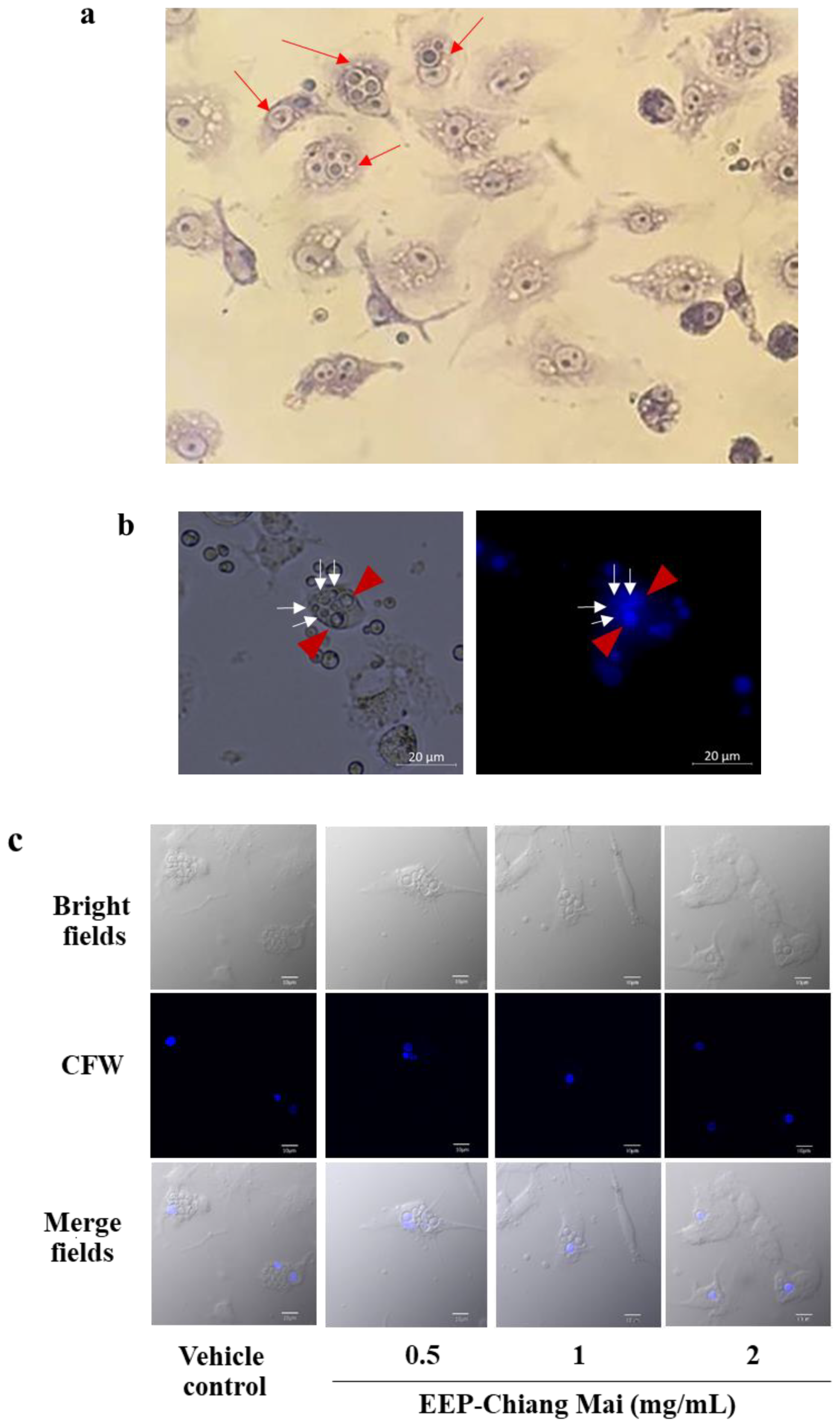
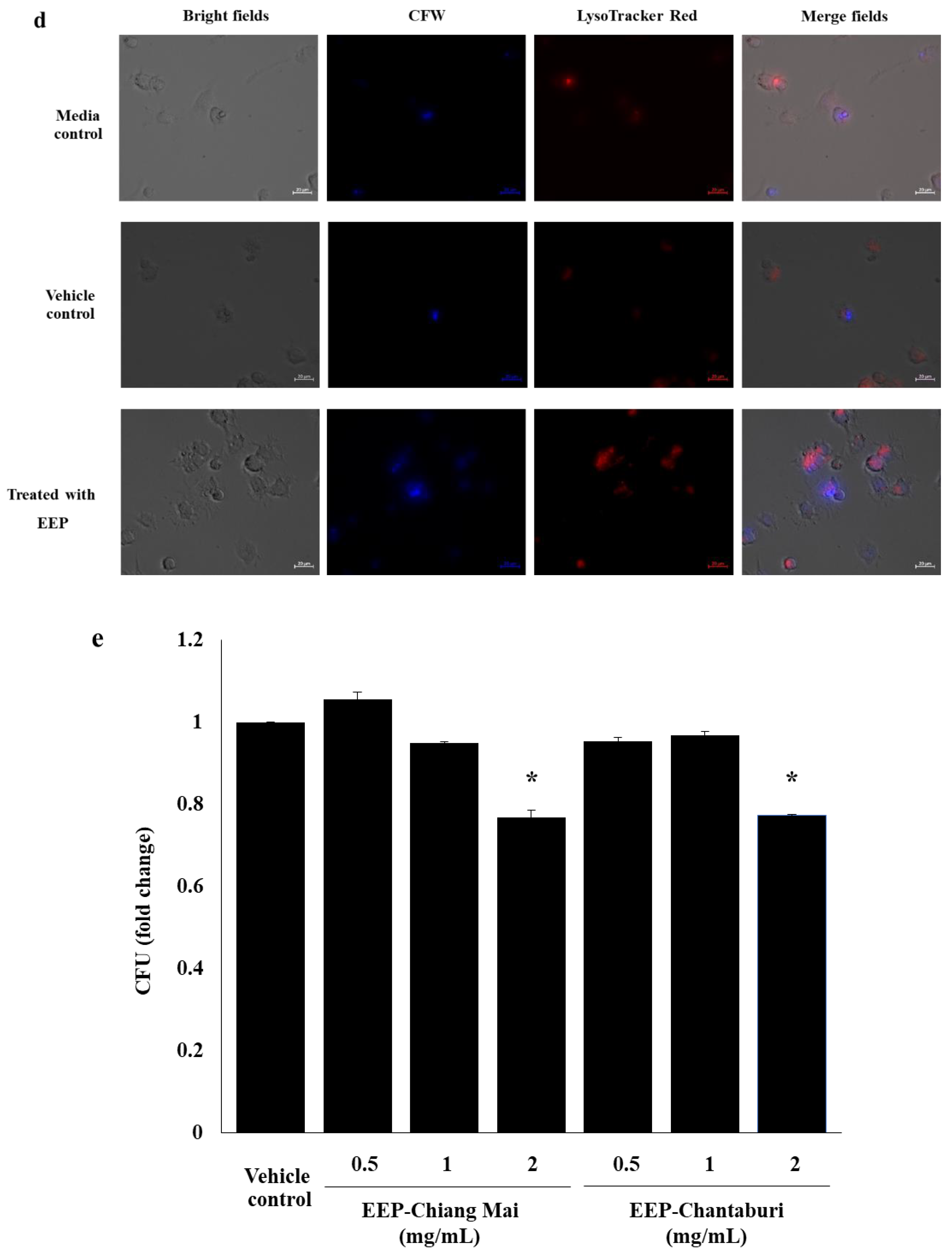
3.4. Intracellular Killing of EEP-Treated C. neoformans in Phagolysosome in Macrophages
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reese, A.J.; Yoneda, A.; Breger, J.A.; Beauvais, A.; Liu, H.; Griffith, C.L.; Bose, I.; Kim, M.J.; Skau, C.; Yang, S.; et al. Loss of cell wall alpha (1–3) glucan affects Cryptococcus neoformans from ultrastructure to virulence. Mol. Microbiol. 2007, 63, 1385–1398. [Google Scholar] [CrossRef] [Green Version]
- Perez-Dulzaides, R.; Camacho, E.; Cordero, R.B.; Casadevall, A. Cell-wall dyes interfere with Cryptococcus neoformans melanin deposition. Microbiology 2018, 164, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Baker, L.G.; Specht, C.A.; Lodge, J.K. Cell wall chitosan is necessary for virulence in the opportunistic pathogen Cryptococcus neoformans. Eukaryot Cell 2011, 10, 1264–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenman, H.C.; Mues, M.; Weber, S.E.; Frases, S.; Chaskes, S.; Gerfen, G.; Casadevall, A. Cryptococcus neoformans laccase catalyses melanin synthesis from both D- and L-DOPA. Microbiology 2007, 153, 3954–3962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heung, L.J.; Kaiser, A.E.; Luberto, C.; Del Poeta, M. The role and mechanism of diacylglycerol-protein kinase C1 signaling in melanogenesis by Cryptococcus neoformans. J. Biol. Chem. 2005, 280, 28547–28555. [Google Scholar] [CrossRef] [Green Version]
- Rosas, A.L.; Casadevall, A. Melanization decreases the susceptibility of Cryptococcus neoformans to enzymatic degradation. Mycopathologia 2001, 151, 53–56. [Google Scholar] [CrossRef]
- Alvarez, M.; Casadevall, A. Phagosome extrusion and host-cell survival after Cryptococcus neoformans phagocytosis by macrophages. Curr. Biol. 2006, 16, 2161–2165. [Google Scholar] [CrossRef] [Green Version]
- Kondo, C.; Aoki, M.; Yamamoto, E.; Tonomura, Y.; Ikeda, M.; Kaneto, M.; Yamate, J.; Torii, M.; Uehara, T. Predictive genomic biomarkers for drug-induced nephrotoxicity in mice. J. Toxicol. Sci. 2012, 37, 723–737. [Google Scholar] [CrossRef] [Green Version]
- Duin, D.; Casadevall, A.; Nosanchuk, J.D. Melanization of Cryptococcus neoformans and Histoplasma capsulatum reduces their susceptibilities to amphotericin B and caspofungin. Antimicrob. Agents Chemother. 2002, 46, 3394–3400. [Google Scholar] [CrossRef] [Green Version]
- Burdock, G.A. Review of the biological properties and toxicity of bee propolis (propolis). Food Chem. Toxicol. 1998, 36, 347–363. [Google Scholar] [CrossRef]
- Sforcin, J.M. Biological properties and therapeutic applications of propolis. Phytother. Res. 2016, 30, 894–905. [Google Scholar] [CrossRef] [PubMed]
- Sanpa, S.; Popova, M.; Bankova, V.; Tunkasiri, T.; Eitssayeam, S.; Chantawannakul, P. Antibacterial compounds from propolis of Tetragonula laeviceps and Tetrigona melanoleuca (Hymenoptera: Apidae) from Thailand. PLoS ONE 2015, 10, 1–11. [Google Scholar] [CrossRef]
- Thammasit, P.; Iadnut, A.; Mamoon, K.; Khacha-Ananda, S.; Chupradit, K.; Tayapiwatana, C.; Kasinrerk, W.; Tragoolpua, Y.; Tragoolpua, K. A potential of propolis on major virulence factors of Cryptococcus neoformans. Microb. Pathog. 2018, 123, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Khacha-ananda, S.; Tragoolpua, K.; Chantawannakul, P.; Tragoolpua, Y. Antioxidant and anti-cancer cell proliferation activity of propolis extracts from two extraction methods. Asian Pac. J. Cancer Prev. 2013, 14, 6991–6995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iadnut, A.; Mamoon, K.; Thammasit, P.; Pawichai, S.; Tima, S.; Preechasuth, K.; Kaewkod, T.; Tragoolpua, Y.; Tragoolpua, K. In vitro antifungal and antivirulence activities of biologically synthesized ethanolic extract of propolis-loaded PLGA nanoparticles against Candida albicans. Evid. Based Complement. Alternat. Med. 2019, 2019, 3715481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerik, K.J.; Bhimireddy, S.R.; Ryerse, J.S.; Specht, C.A.; Lodge, J.K. PKC1 is essential for protection against both oxidative and nitrosative stresses, cell integrity, and normal manifestation of virulence factors in the pathogenic fungus Cryptococcus neoformans. Eukaryot Cell 2008, 7, 1685–1698. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, F.L.; Guimaraes, A.J.; Kmetzsch, L.; Dutra, F.F.; Silva, F.D.; Taborda, C.P.; Araujo Gde, S.; Frases, S.; Staats, C.C.; Bozza, M.T.; et al. Binding of the wheat germ lectin to Cryptococcus neoformans chitooligomers affects multiple mechanisms required for fungal pathogenesis. Fungal. Genet. Biol. 2013, 60, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Sun, N.; Xiao, D.; Pan, J.; Wang, Y.; Zhu, X. A copper-responsive factor gene CUF1 is required for copper induction of laccase in Cryptococcus neoformans. FEMS Microbiol. Lett. 2009, 296, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Araujo Gde, S.; Fonseca, F.L.; Pontes, B.; Torres, A.; Cordero, R.J.; Zancope-Oliveira, R.M.; Casadevall, A.; Viana, N.B.; Nimrichter, L.; Rodrigues, M.L.; et al. Capsules from pathogenic and non-pathogenic Cryptococcus spp. manifest significant differences in structure and ability to protect against phagocytic cells. PLoS ONE 2012, 7, 1–11. [Google Scholar] [CrossRef]
- Nicola, A.M.; Casadevall, A. In vitro measurement of phagocytosis and killing of Cryptococcus neoformans by macrophages. Methods. Mol. Biol. 2012, 844, 189–197. [Google Scholar] [CrossRef]
- Surawut, S.; Ondee, T.; Taratummarat, S.; Palaga, T.; Pisitkun, P.; Chindamporn, A.; Leelahavanichkul, A. The role of macrophages in the susceptibility of Fc gamma receptor IIb deficient mice to Cryptococcus neoformans. Sci. Rep. 2017, 7, 40006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Przybylek, I.; Karpinski, T.M. Antibacterial properties of propolis. Molecules 2019, 24, 2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trusheva, B.; Trunkova, D.; Bankova, V. Different extraction methods of biologically active components from propolis: A preliminary study. Chem. Cent J. 2007, 1, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasote, D.M.; Pawar, M.V.; Gundu, S.S.; Bhatia, R.; Nandre, V.S.; Jagtap, S.D.; Mahajan, S.G.; Kulkarni, M.V. Chemical profiling, antioxidant, and antimicrobial activities of Indian stingless bees propolis samples. J. Apic. Res. 2019, 58, 617–625. [Google Scholar] [CrossRef]
- Massaro, F.C.; Brooks, P.R.; Wallace, H.M.; Nsengiyumva, V.; Narokai, L.; Russell, F.D. Effect of Australian propolis from stingless bees (Tetragonula carbonaria) on pre-contracted human and porcine isolated arteries. PLoS ONE 2013, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banks, I.R.; Specht, C.A.; Donlin, M.J.; Gerik, K.J.; Levitz, S.M.; Lodge, J.K. A chitin synthase and its regulator protein are critical for chitosan production and growth of the fungal pathogen Cryptococcus neoformans. Eukaryot Cell 2005, 4, 1902–1912. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Guo, M. Studies on transition metal-quercetin complexes using electrospray ionization tandem mass spectrometry. Molecules 2015, 20, 8583–8594. [Google Scholar] [CrossRef]
- Andreini, C.; Bertini, I.; Cavallaro, G.; Holliday, G.L.; Thornton, J.M. Metal ions in biological catalysis: From enzyme databases to general principles. J. Biol. Inorg. Chem. 2008, 13, 1205–1218. [Google Scholar] [CrossRef]
- Kim, Y.J.; Zhao, Y.; Oh, K.T.; Nguyen, V.N.; Park, R.D. Enzymatic deacetylation of chitin by extracellular chitin deacetylase from a newly screened Mortierella sp. DY-52. J. Microbiol. Biotechnol. 2008, 18, 759–766. [Google Scholar] [PubMed]
- Upadhya, R.; Baker, L.G.; Lam, W.C.; Specht, C.A.; Donlin, M.J.; Lodge, J.K. Cryptococcus neoformans Cda1 and its chitin deacetylase activity are required for fungal pathogenesis. MBio 2018, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Helsel, M.E.; White, E.J.; Razvi, S.Z.; Alies, B.; Franz, K.J. Chemical and functional properties of metal chelators that mobilize copper to elicit fungal killing of Cryptococcus neoformans. Metallomics 2017, 9, 69–81; [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, F.D.; Rossi, D.C.; Martinez, L.R.; Frases, S.; Fonseca, F.L.; Campos, C.B.; Rodrigues, M.L.; Nosanchuk, J.D.; Daffre, S. Effects of microplusin, a copper-chelating antimicrobial peptide, against Cryptococcus neoformans. FEMS Microbiol. Lett. 2011, 324, 64–72; [Google Scholar] [CrossRef] [Green Version]
- Gu, C.; Stashko, M.A.; Puhl-Rubio, A.C.; Chakraborty, M.; Chakraborty, A.; Frye, S.V.; Pearce, K.H.; Wang, X.; Shears, S.B.; Wang, H. Inhibition of inositol polyphosphate kinases by quercetin and related flavonoids: A structure-activity analysis. J. Med. Chem. 2019, 62, 1443–1454; [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Casadevall, A. Susceptibility of melanized and nonmelanized Cryptococcus neoformans to nitrogen- and oxygen-derived oxidants. Infect. Immun. 1994, 62, 3004–3007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steenbergen, J.N.; Shuman, H.A.; Casadevall, A. Cryptococcus neoformans interactions with amoebae suggest an explanation for its virulence and intracellular pathogenic strategy in macrophages. Proc. Natl. Acad. Sci. USA 2001, 98, 15245–15250; [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, T.; Kawai, C. Experimental cryptococcal myocarditis. Res. Exp. Med (Berl). 1980, 178, 11–19. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Rudolph, J.; Rosas, A.L.; Casadevall, A. Evidence that Cryptococcus neoformans is melanized in pigeon excreta: Implications for pathogenesis. Infect. Immun. 1999, 67, 5477–5479. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EEP Source | Total Phenolic (mg GAE 1/g extract) | Total Flavonoid (mg QE 2/g extract) |
|---|---|---|
| Chiang Mai | 4.31 ± 0.11 | 6.53 ± 1.07 |
| Chantaburi | 3.84 ± 0.03 | 4.75 ± 1.56 |
| EEP Source | Chemical Compound | Concentration (µg/mL) |
|---|---|---|
| Chiang Mai | Gallic acid | 0.34 |
| Quercetin | 1.13 | |
| Pinocembrin | 2.19 | |
| Chantaburi | Gallic acid | 1.03 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mamoon, K.; Thammasit, P.; Iadnut, A.; Kitidee, K.; Anukool, U.; Tragoolpua, Y.; Tragoolpua, K. Unveiling the Properties of Thai Stingless Bee Propolis via Diminishing Cell Wall-Associated Cryptococcal Melanin and Enhancing the Fungicidal Activity of Macrophages. Antibiotics 2020, 9, 420. https://doi.org/10.3390/antibiotics9070420
Mamoon K, Thammasit P, Iadnut A, Kitidee K, Anukool U, Tragoolpua Y, Tragoolpua K. Unveiling the Properties of Thai Stingless Bee Propolis via Diminishing Cell Wall-Associated Cryptococcal Melanin and Enhancing the Fungicidal Activity of Macrophages. Antibiotics. 2020; 9(7):420. https://doi.org/10.3390/antibiotics9070420
Chicago/Turabian StyleMamoon, Ketsaya, Patcharin Thammasit, Anupon Iadnut, Kuntida Kitidee, Usanee Anukool, Yingmanee Tragoolpua, and Khajornsak Tragoolpua. 2020. "Unveiling the Properties of Thai Stingless Bee Propolis via Diminishing Cell Wall-Associated Cryptococcal Melanin and Enhancing the Fungicidal Activity of Macrophages" Antibiotics 9, no. 7: 420. https://doi.org/10.3390/antibiotics9070420