Antimicrobial Effects of Selected, Cultivated Red Seaweeds and Their Components in Combination with Tetracycline, against Poultry Pathogen Salmonella Enteritidis

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain, Chemicals and Antibiotics
2.2. Antibiotic Sensitivity Assay
2.3. Determination of MIC of Antibiotics
2.4. Combined Effect of Seaweed Extracts (SWE) and Antibiotics on Salmonella Enteritidis
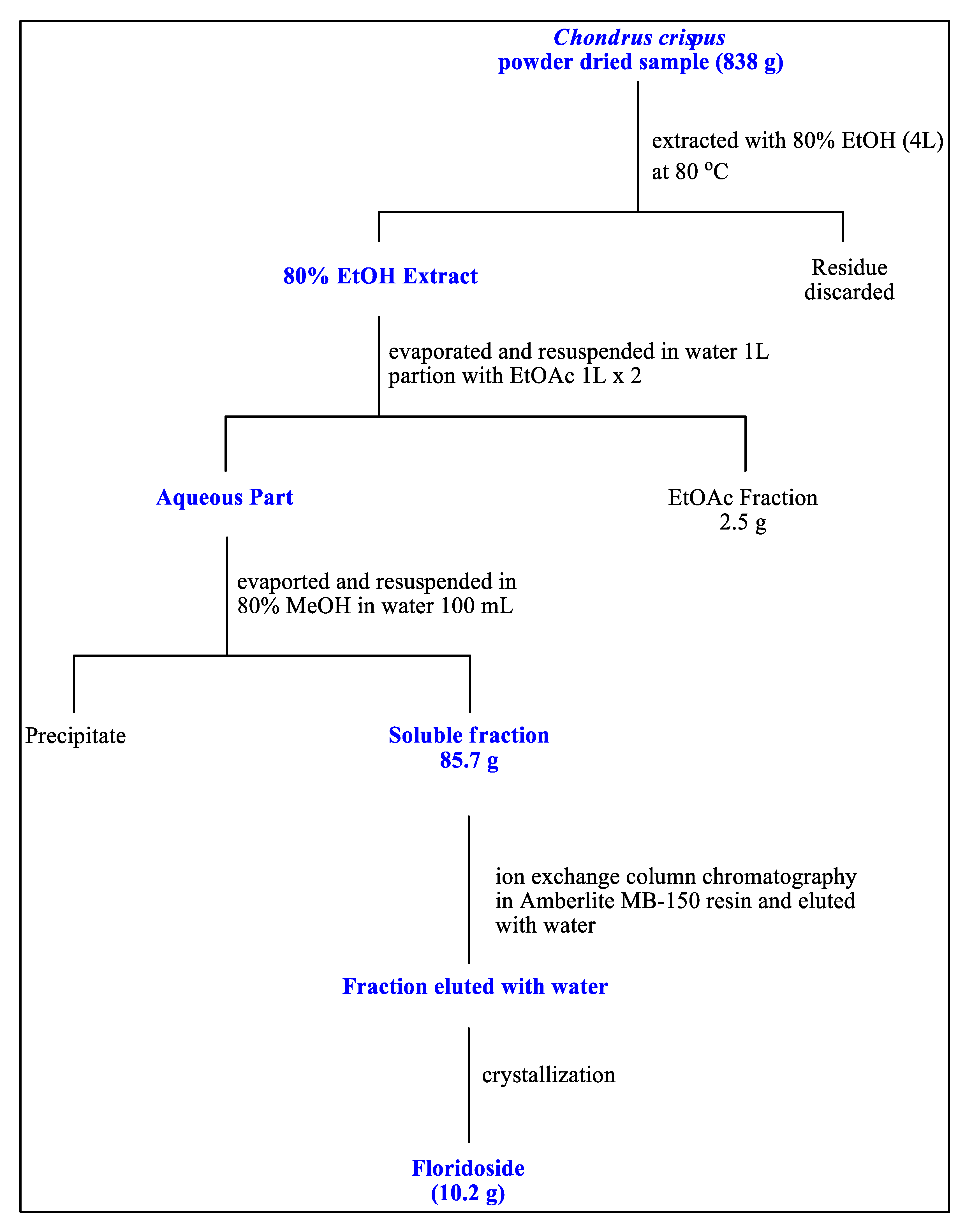
2.5. Extraction of Seaweed and Isolation of Floridoside
2.6. Antimicrobial Effects of Seaweed Components on Salmonella Enteritidis
2.7. Combined Effects of Floridoside and Tetracycline on Salmonella Enteritidis
2.8. Effects of Floridoside and Tetracycline on the Expression of Efflux-Pump-Related Genes
2.9. Statistical Analyses
3. Results
3.1. Screening of Antibiotics against Salmonella Enteritidis
3.2. Determination of Minimum Inhibitory Concentrations (MIC25 and MIC50)
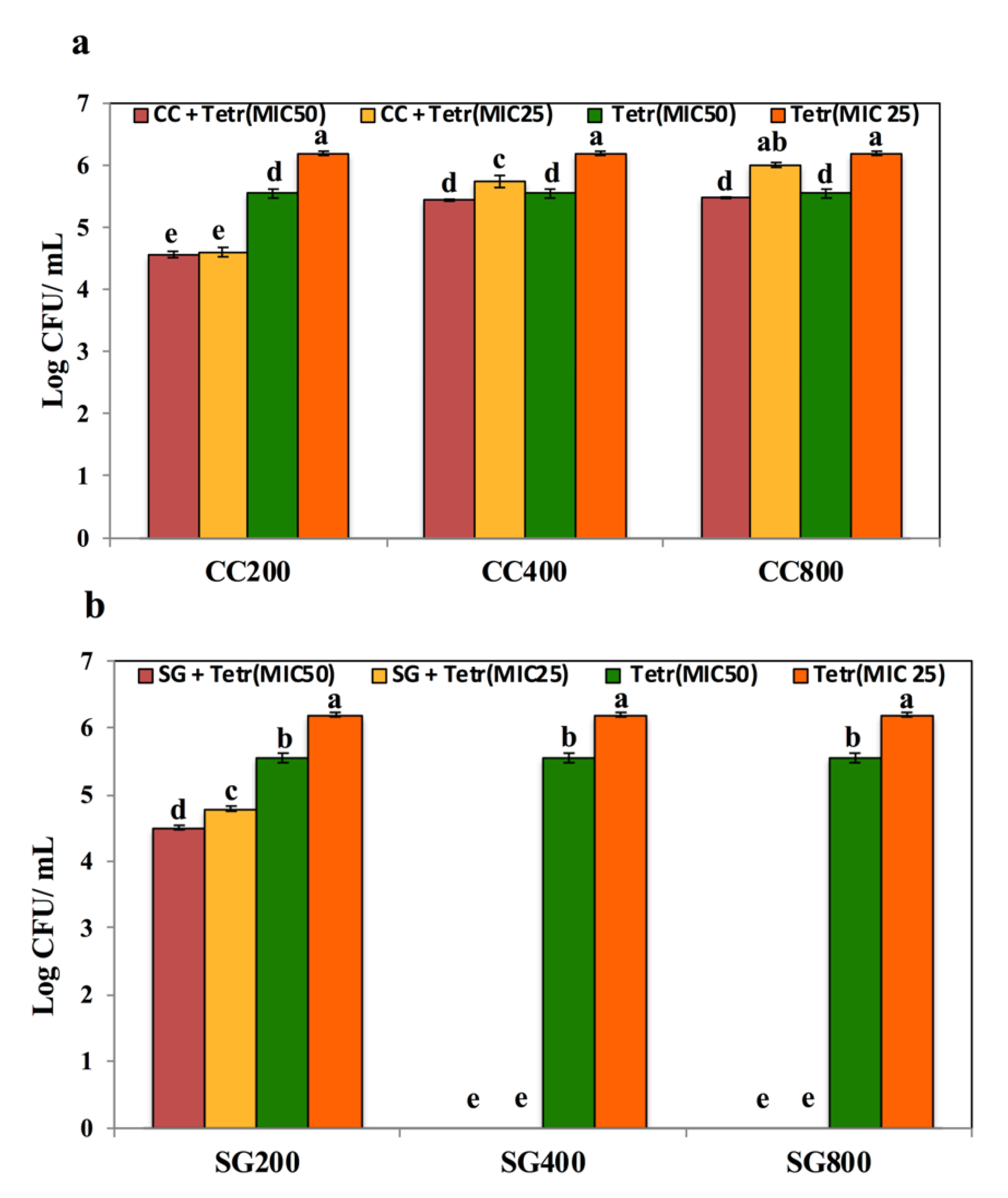
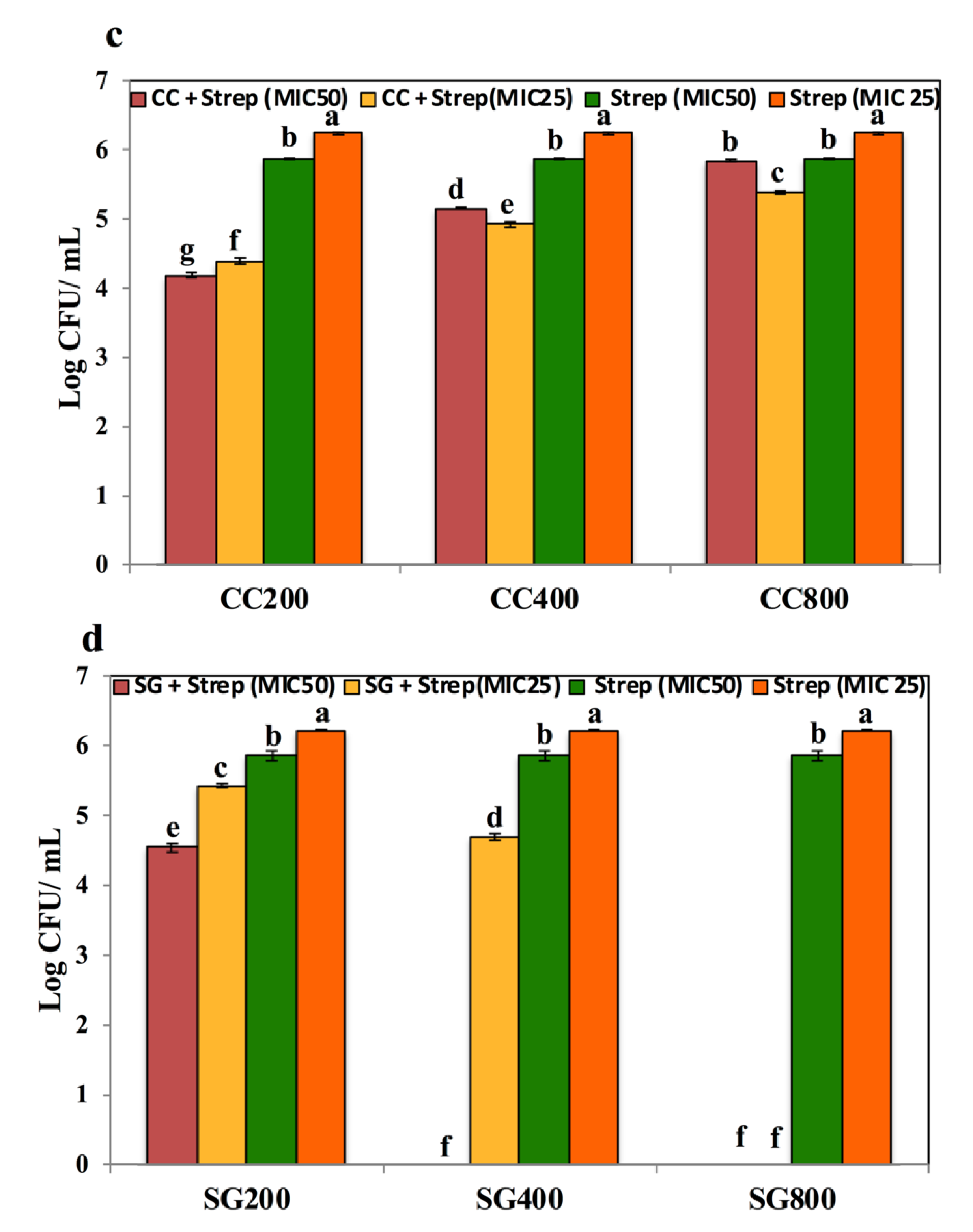
3.3. SWE Potentiated the Effect of Antibiotics on Salmonella Enteritidis
3.4. 1H Nuclear Magnetic Resonance Spectroscopy of Seaweed Water Extracts
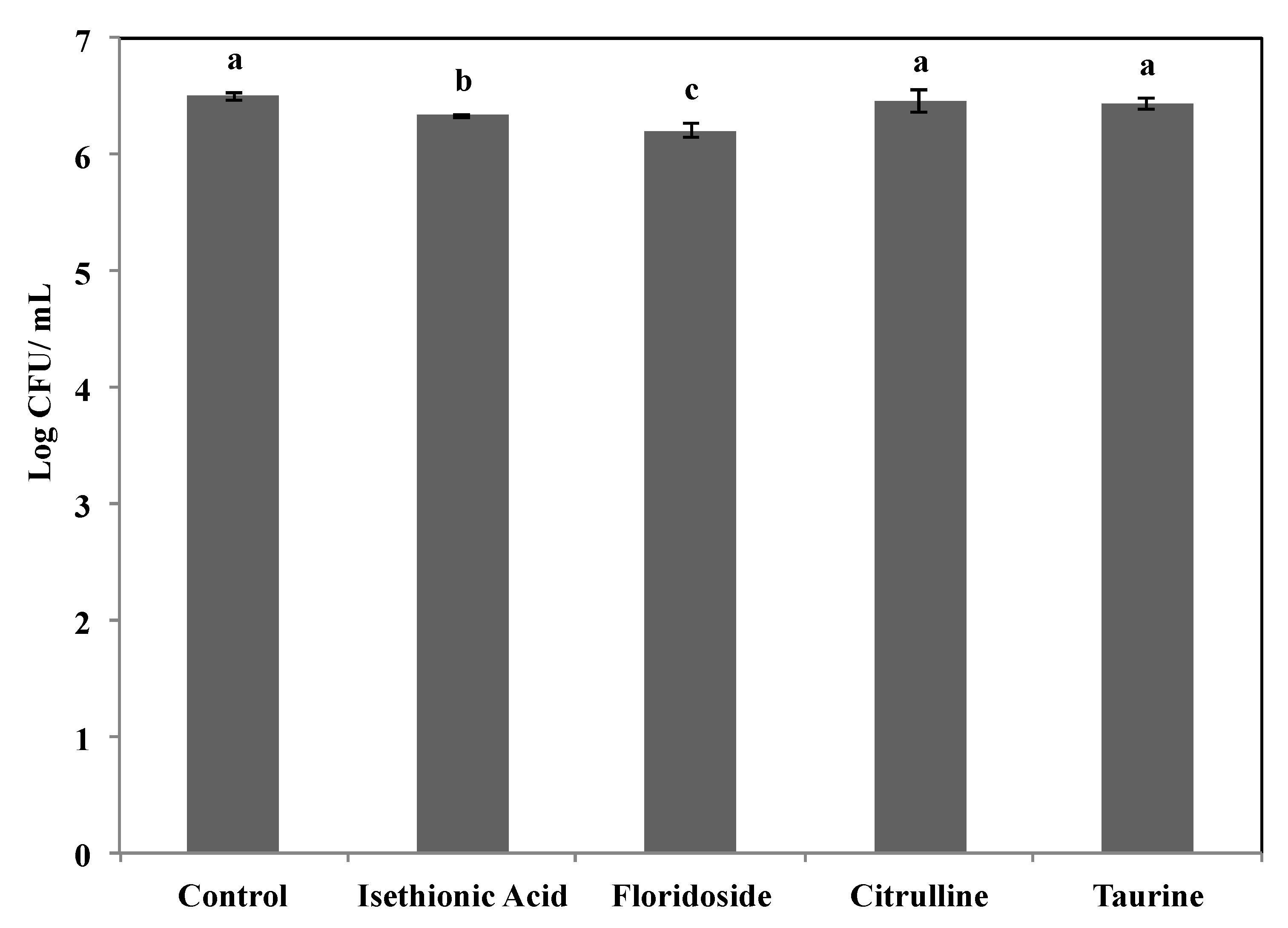
3.5. Floridoside Affected the Growth of S. Enteritidis
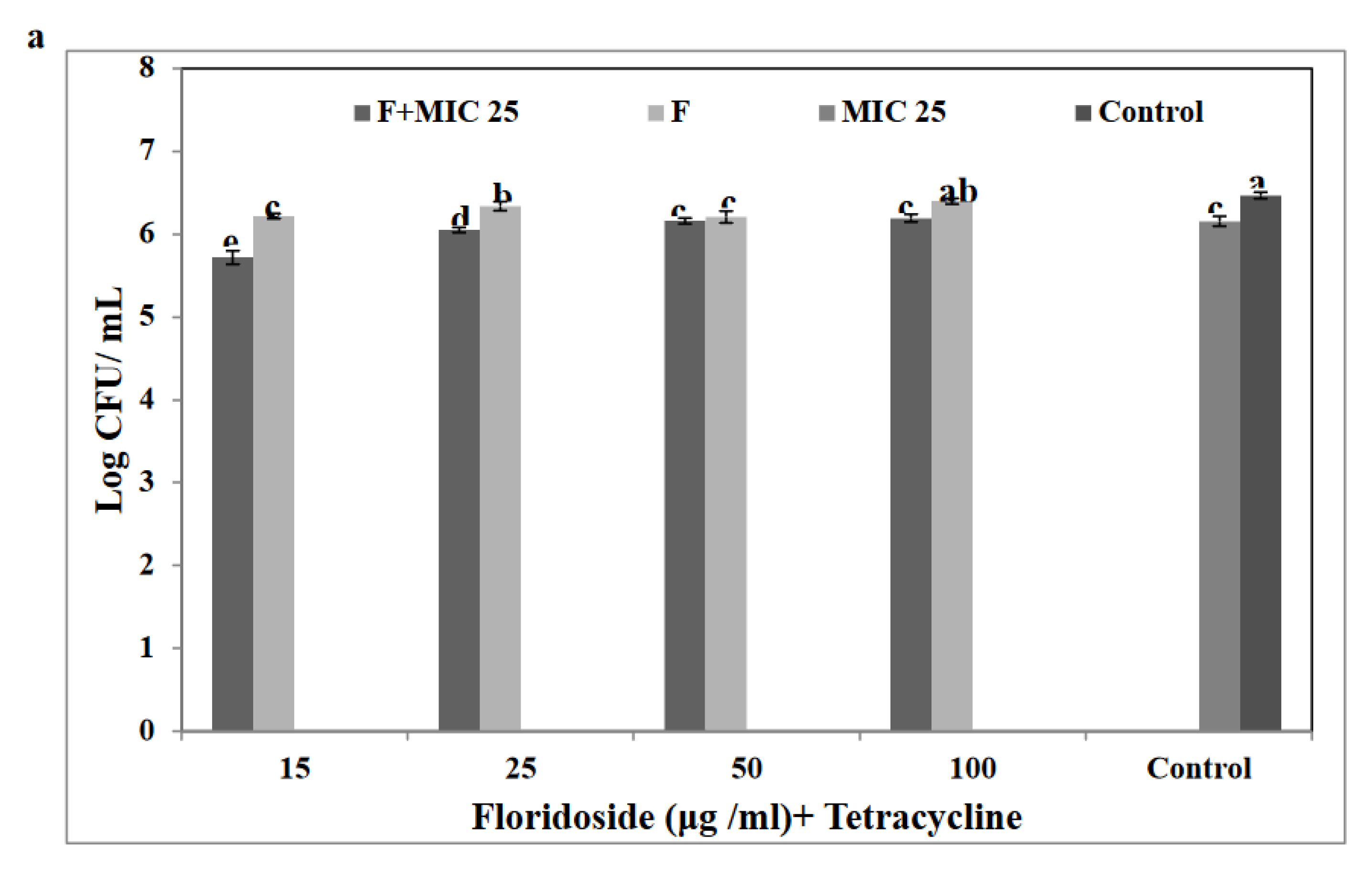
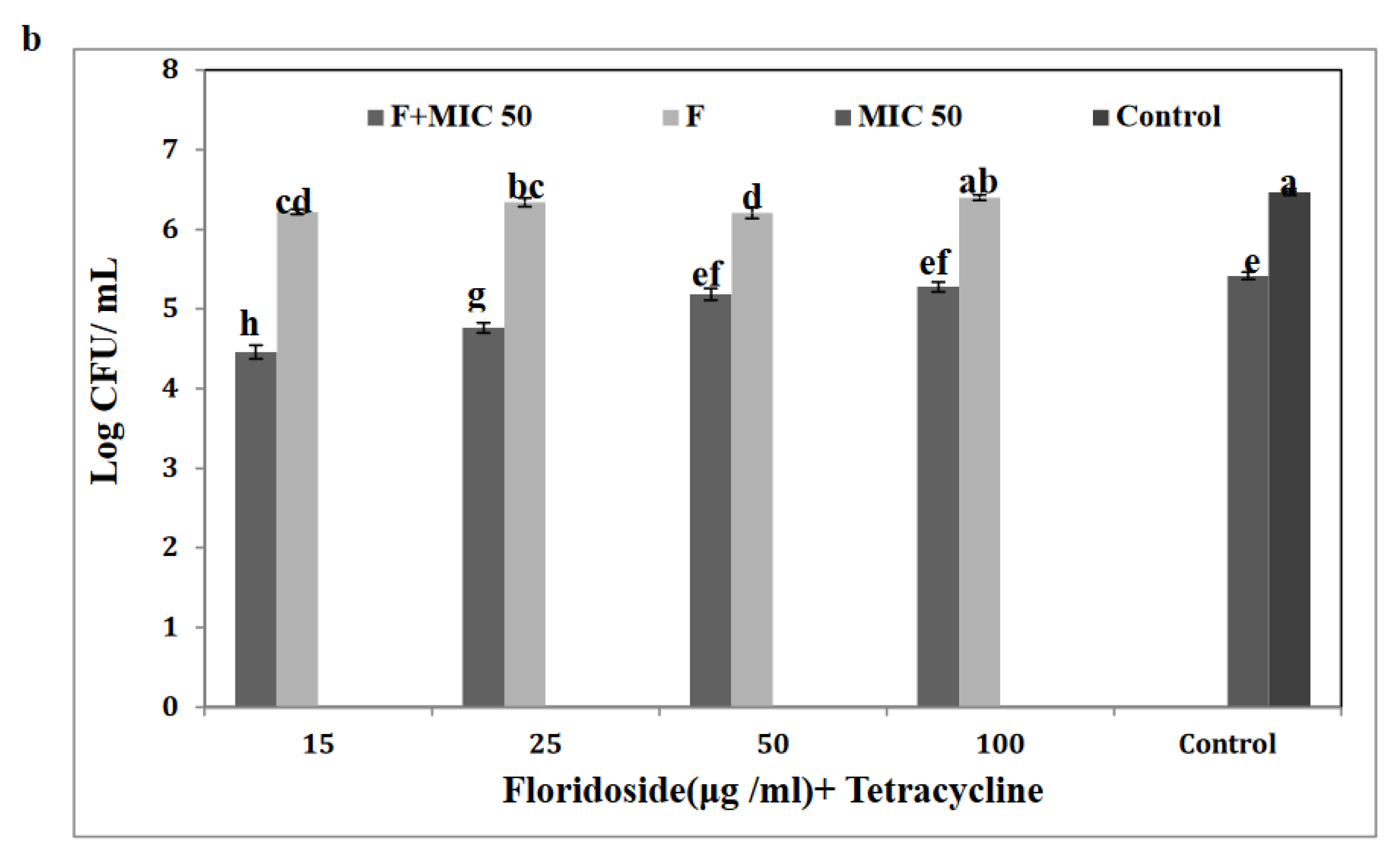
3.6. Floridoside Potentiated the Activity of Tetracycline against S. Enteritidis
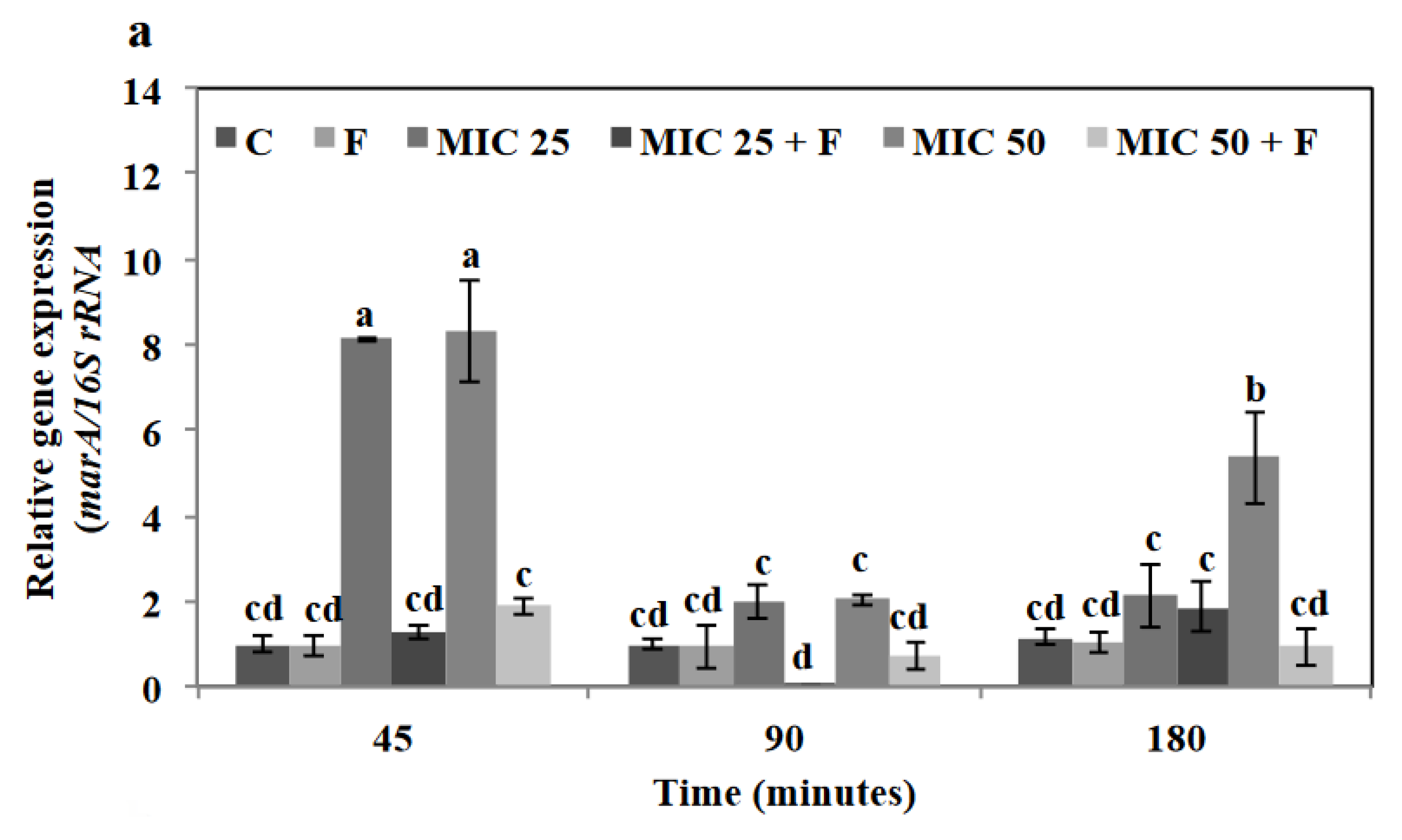
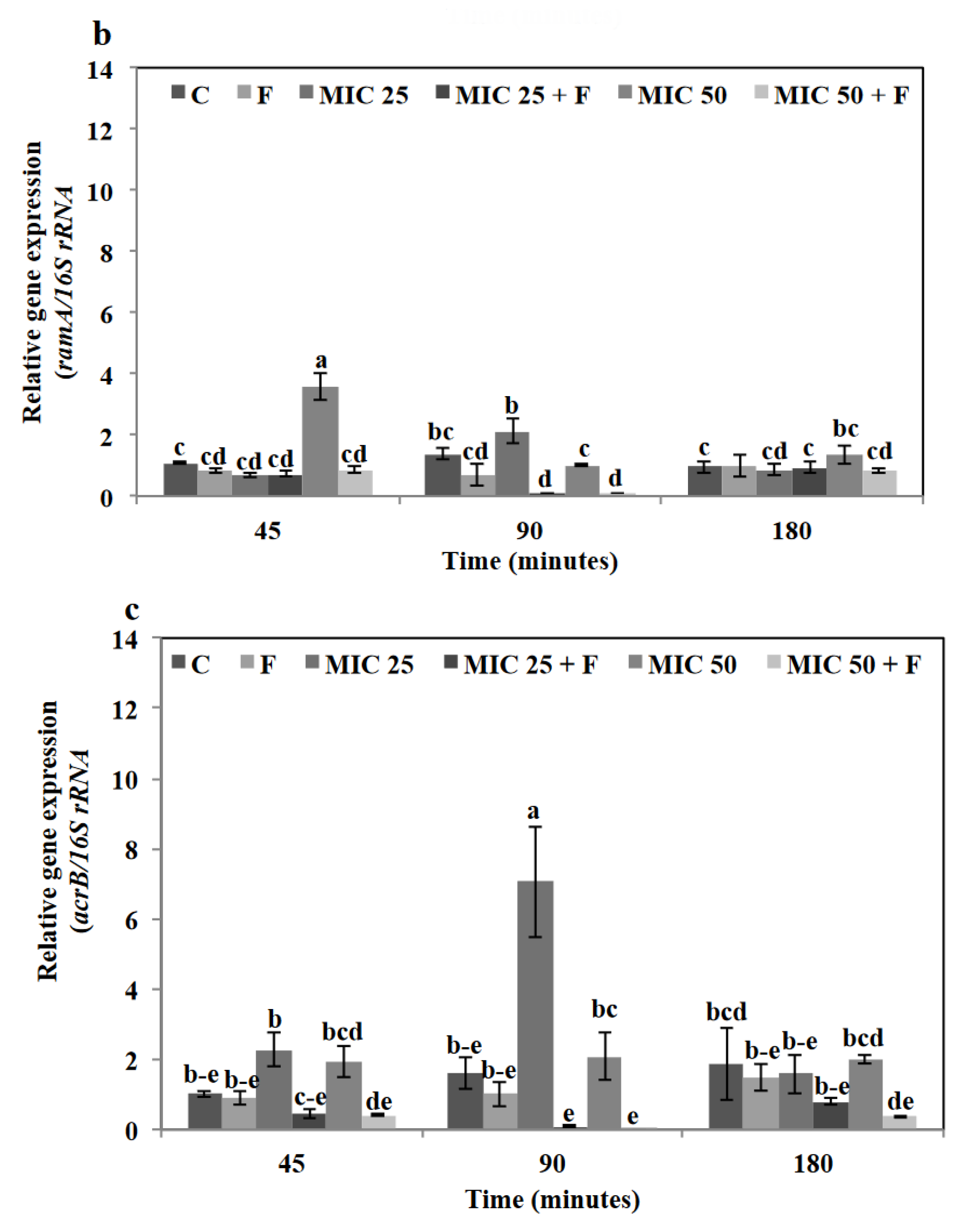
3.7. Floridoside and Tetracycline Suppressed the Expression of Efflux-Pump-Related Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brenes, A.; Roura, E. Essential oils in poultry nutrition: Main effects and modes of action. Anim. Feed Sci. Technol. 2010, 158, 1–14. [Google Scholar] [CrossRef]
- Venkitanarayanan, K.; Kollanoor-Johny, A.; Darre, M.J.; Donoghue, A.M.; Donoghue, D.J. Use of plant-derived antimicrobials for improving the safety of poultry products1. Poult. Sci. 2013, 92, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Aze, N.A.; El-Daly, E.F.; Hassan, H.M.A.; Youssef, A.W.; Mohamed, M.A. Histological Response of Broilers Immune Related Organs to Feeding Different Direct Fed Microbials. Int. J. Poult. Sci. 2015, 14, 331–337. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Food and Food Animals | Antibiotic/Antimicrobial Resistance | CDC. Available online: https://www.cdc.gov/drugresistance/food.html (accessed on 10 May 2020).
- Van Boeckel, T.P.; Gandra, S.; Ashok, A.; Caudron, Q.; Grenfell, B.T.; Levin, S.A.; Laxminarayan, R. Global antibiotic consumption 2000 to 2010: An analysis of national pharmaceutical sales data. Lancet Infect. Dis. 2014, 14, 742–750. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [Green Version]
- Van Boeckel, T.P.; Pires, J.; Silvester, R.; Zhao, C.; Song, J.; Criscuolo, N.G.; Gilbert, M.; Bonhoeffer, S.; Laxminarayan, R. Global trends in antimicrobial resistance in animals in low- and middle-income countries. Science 2019, 365. [Google Scholar] [CrossRef] [Green Version]
- U.S. Food and Drug Administration. FDA Annual Summary Report on Antimicrobials Sold or Distributed in 2012 for Use in Food-Producing Animals. Available online: http://www.fda.gov/AnimalVeterinary/NewsEvents/CVMUpdates/ucm416974.htm (accessed on 10 May 2020).
- U.S. Food and Drug Administration. Poultry and Eggs: Background. Available online: http://www.ers.usda.gov/topics/animal-products/poultry-eggs/background.aspx (accessed on 10 May 2020).
- Canadian Food Inspection Agency. Index of Medicating Ingredients Approved by Livestock Species—Canadian Food Inspection Agency. Available online: https://www.inspection.gc.ca/animal-health/livestock-feeds/medicating-ingredients/mib/livestock-species/eng/1522783196554/1522783196850 (accessed on 10 May 2020).
- Collier, C.T.; Van der Klis, J.D.; Deplancke, B.; Anderson, D.B.; Gaskins, H.R. Effects of tylosin on bacterial mucolysis, Clostridium perfringens colonization, and intestinal barrier function in a chick model of necrotic enteritis. Antimicrob. Agents Chemother. 2003, 47, 3311–3317. [Google Scholar] [CrossRef] [Green Version]
- Roura, E.; Homedes, J.; Klasing, K.C. Prevention of immunologic stress contributes to the growth-permitting ability of dietary antibiotics in chicks. J. Nutr. 1992, 122, 2383–2390. [Google Scholar] [CrossRef]
- Starr, M.P.; Reynolds, D.M. Streptomycin resistance of coliform bacteria from turkeys fed streptomycin. Am. J. Public Health Nations Health 1951, 41, 1375–1380. [Google Scholar] [CrossRef]
- Bates, J.; Jordens, J.Z.; Griffiths, D.T. Farm animals as a putative reservoir for vancomycin-resistant enterococcal infection in man. J. Antimicrob. Chemother. 1994, 34, 507–514. [Google Scholar] [CrossRef]
- Castanon, J.I.R. History of the use of antibiotic as growth promoters in European poultry feeds. Poult. Sci. 2007, 86, 2466–2471. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.; Inman, C.F.; Bailey, M. Review: Postnatal development of the mucosal immune system and consequences on health in adulthood. Can. J. Anim. Sci. 2010, 90, 129–136. [Google Scholar] [CrossRef]
- Yan, G.L.; Guo, Y.M.; Yuan, J.M.; Liu, D.; Zhang, B.K. Sodium alginate oligosaccharides from brown algae inhibit Salmonella Enteritidis colonization in broiler chickens. Poult. Sci. 2011, 90, 1441–1448. [Google Scholar] [CrossRef] [PubMed]
- Filho, R.A.C.P.; de Paiva, J.B.; Argüello, Y.M.S.; da Silva, M.D.; Gardin, Y.; Resende, F.; Junior, A.B.; Sesti, L. Efficacy of several vaccination programmes in commercial layer and broiler breeder hens against experimental challenge with Salmonella enterica serovar Enteritidis. Avian Pathol. 2009, 38, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention (CDC). Biggest Threats and Data, Antibiotic Antimicrobial Resistance. Available online: https://www.cdc.gov/drugresistance/biggest-threats.html (accessed on 10 May 2020).
- Amábile-Cuevas, C.F. Antibiotic resistance: From Darwin to Lederberg to Keynes. Microb. Drug Resist. 2013, 19, 73–87. [Google Scholar] [CrossRef]
- Hawkey, P.M. Why are bacteria resistant to antibiotics? Intensive Care Med. 2000, 26, S009–S013. [Google Scholar]
- Nikaido, H. Multidrug Resistance in Bacteria. Annu. Rev. Biochem. 2009, 78, 119–146. [Google Scholar] [CrossRef] [Green Version]
- Nikaido, H. Prevention of drug access to bacterial targets: Permeability barriers and active efflux. Science 1994, 264, 382–388. [Google Scholar] [CrossRef] [Green Version]
- Mah, T.F.; Pitts, B.; Pellock, B.; Walker, G.C.; Stewart, P.S.; O’Toole, G.A. A genetic basis for Pseudomonas aeruginosa biofilm antibiotic resistance. Nature 2003, 426, 306–310. [Google Scholar] [CrossRef]
- Khan, S.; Tøndervik, A.; Sletta, H.; Klinkenberg, G.; Emanuel, C.; Onsøyen, E.; Myrvold, R.; Howe, R.A.; Walsh, T.R.; Hill, K.E.; et al. Overcoming drug resistance with alginate oligosaccharides able to potentiate the action of selected antibiotics. Antimicrob. Agents Chemother. 2012, 56, 5134–5141. [Google Scholar] [CrossRef] [Green Version]
- Hussin, W.A.; El-Sayed, W.M. Synergic interactions between selected botanical extracts and tetracycline against gram positive and gram negative bacteria. J. Biol. Sci. 2011, 11, 433–441. [Google Scholar] [CrossRef]
- Singh, A.P.; Prabha, V.; Rishi, P. Value Addition in the Efficacy of Conventional Antibiotics by Nisin against Salmonella. PLoS ONE 2013, 8, e76844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hierowski, M. Inhibition of protein synthesis by chlortetracycline in the E. coli in In vitro system. Proc. Natl. Acad. Sci. USA 1965, 53, 594–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chovanová, R.; Mezovská, J.; Vaverková, Š.; Mikulášová, M. The inhibition the Tet(K) efflux pump of tetracycline resistant Staphylococcus epidermidis by essential oils from three Salvia species. Lett. Appl. Microbiol. 2015, 61, 58–62. [Google Scholar] [CrossRef]
- Reed, R.H.; Collins, J.C.; Russell, G. Effects of Salinity upon Galactosyl-Glycerol Content and Concentration of the Marine Red Alga Porphyra purpurea (Roth) C.Ag. J. Exp. Bot. 1980, 31, 1539–1554. [Google Scholar] [CrossRef]
- Pardoe, I.; Hartley, C. Carbohydrates such as Alpha-D-Galactopyranosyl-1, 2-glycerol and/or Isomers, Obtained by Solvent Extraction from Algae, for Use as Viricides or Anticarcinogenic Agents. U.S. Patent Application 10/344,812, 2001. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically. In Approved Standard, 9th ed.; CLSI document M07-A9; CLSI: Wayne, PA, USA, 2013. [Google Scholar]
- Jorgensen, J.; Ferraro, M. Antimicrobial susceptibility testing: General principles and contemporary practices. J. Clin. Infect. Dis. 2009, 49, 1749–1755. [Google Scholar] [CrossRef]
- Kulshreshtha, G.; Borza, T.; Rathgeber, B.; Stratton, G.S.; Thomas, N.A.; Critchley, A.; Hafting, J.; Prithiviraj, B. Red seaweeds Sarcodiotheca gaudichaudii and Chondrus crispus down regulate virulence factors of Salmonella enteritidis and induce immune responses in Caenorhabditis elegans. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Su, L.-H.; Chiu, C.-H.; Chu, C.; Ou, J.T. Antimicrobial Resistance in Nontyphoid Salmonella Serotypes: A Global Challenge. Clin. Infect. Dis. 2004, 39, 546–551. [Google Scholar] [CrossRef] [Green Version]
- Guerra, B.; Junker, E.; Miko, A.; Helmuth, R.; Mendoza, M.C. Characterization and localization of drug resistance determinants in multidrug-resistant, integron-carrying Salmonella enterica serotype typhimurium strains. Microb. Drug Resist. 2004, 10, 83–91. [Google Scholar] [CrossRef]
- Infection Disease Society of America (ISDA). Review Finds Antibiotic Development Increased, but Insufficient. Available online: https://www.idsociety.org/news--publications-new/articles/2019/review-finds-antibiotic-development-increased-but-insufficient/ (accessed on 10 May 2020).
- Talbot, G.H.; Jezek, A.; Murray, B.E.; Jones, R.N.; Ebright, R.H.; Nau, G.J.; Rodvold, K.A.; Newland, J.G.; Boucher, H.W. The Infectious Diseases Society of America’s 10 × ’20 Initiative (10 New Systemic Antibacterial Agents US Food and Drug Administration Approved by 2020): Is 20 × ’20 a Possibility? Clin. Infect. Dis. 2019, 69, 1–11. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, Y.H.; Seo, Y.W.; Park, S. Quorum sensing inhibitors from the red alga, Ahnfeltiopsis flabelliformis. Biotechnol. Bioprocess. Eng. 2007, 12, 308–311. [Google Scholar] [CrossRef]
- Liu, H.B.; Koh, K.P.; Kim, J.S.; Seo, Y.; Park, S. The effects of betonicine, floridoside, and isethionic acid from the red alga Ahnfeltiopsis flabelliformis on quorum-sensing activity. Biotechnol. Bioprocess. Eng. 2008, 13, 458–463. [Google Scholar] [CrossRef]
- Janssens, J.C.A.; Steenackers, H.; Robijns, S.; Gellens, E.; Levin, J.; Zhao, H.; Hermans, K.; De Coster, D.; Verhoeven, T.L.; Marchal, K.; et al. Brominated furanones inhibit biofilm formation by Salmonella enterica serovar Typhimurium. Appl. Environ. Microbiol. 2008, 74, 6639–6648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.; Bahar, A.A.; Syed, H.; Ren, D. Reverting antibiotic tolerance of Pseudomonas aeruginosa PAO1 Persister Cells by (Z)-4-bromo-5-(bromomethylene)-3-methylfuran-2(5H)-one. PLoS ONE 2012, 7, e45778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharafutdinov, I.S.; Trizna, E.Y.; Baidamshina, D.R.; Ryzhikova, M.N.; Sibgatullina, R.R.; Khabibrakhmanova, A.M.; Latypova, L.Z.; Kurbangalieva, A.R.; Rozhina, E.V.; Klinger-Strobel, M.; et al. Antimicrobial effects of sulfonyl derivative of 2(5H)-Furanone against planktonic and biofilm associated methicillin-resistant and -susceptible Staphylococcus aureus. Front. Microbiol. 2017, 8, 2246. [Google Scholar] [CrossRef]
- Maseda, H.; Sawada, I.; Saito, K.; Uchiyama, H.; Nakae, T.; Nomura, N. Enhancement of the mexAB-oprM Efflux Pump Expression by a quorum-Sensing autoinducer and its cancellation by a regulator, MexT, of the mexEF-oprN efflux pump operon in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2004, 48, 1320–1328. [Google Scholar] [CrossRef] [Green Version]
- Rajić, A.; Waddell, L.A.; Sargeant, J.M.; Read, S.; Farber, J.; Firth, M.J.; Chambers, A. An overview of microbial food safety programs in beef, pork, and poultry from farm to processing in Canada. J. Food Prot. 2007, 70, 1286–1294. [Google Scholar] [CrossRef]
- Zamora-Sanabria, R.; Alvarado, A.M. Preharvest Salmonella Risk Contamination and the Control Strategies. In Current Topics in Salmonella and Salmonellosis; InTechOpen: Rijeka, Croatia, 2017. [Google Scholar]
- Jajere, S.M. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and adaptation and antimicrobial resistance including multidrug resistance. Vet. World 2019, 12, 504–521. [Google Scholar] [CrossRef] [Green Version]
- Okamura, M.; Miyamoto, T.; Kamijima, Y.; Tani, H.; Sasai, K.; Baba, E. Differences in abilities to colonize reproductive organs and to contaminate eggs in intravaginally inoculated hens and in vitro adherences to vaginal explants between Salmonella enteritidis and other Salmonella serovars. Avian Dis. 2001, 45, 962–971. [Google Scholar] [CrossRef]
- Foley, S.L.; Johnson, T.J.; Ricke, S.C.; Nayak, R.; Danzeisen, J. Salmonella Pathogenicity and Host Adaptation in Chicken-Associated Serovars. Microbiol. Mol. Biol. Rev. 2013, 77, 582–607. [Google Scholar] [CrossRef] [Green Version]
- Ebers, K.L.; Zhang, C.Y.; Zhang, M.Z.; Bailey, R.H.; Zhang, S. Transcriptional profiling avian beta-defensins in chicken oviduct epithelial cells before and after infection with Salmonella enterica serovar Enteritidis. BMC Microbiol. 2009, 9, 153. [Google Scholar] [CrossRef] [Green Version]
- Rahmati, S.; Yang, S.; Davidson, A.L.; Zechiedrich, E.L. Control of the AcrAB multidrug efflux pump by quorum-sensing regulator SdiA. Mol. Microbiol. 2002, 43, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Littlejohn, T.G.; Paulsen, I.T.; Gillespie, M.T.; Tennent, J.M.; Midgley, M.; Jones, I.G.; Purewal, A.S.; Skurray, R.A. Substrate specificity and energetics of antiseptic and disinfectant resistance in Staphylococcus aureus. FEMS Microbiol. Lett. 1992, 95, 259–265. [Google Scholar] [CrossRef]
- Poole, K. Multidrug Efflux Pumps and Antimicrobial Resistance in Pseudomonas aeruginosa and Related Organisms. J. Mol. Microbiol. Biotechnol. 2001, 3, 255–263. [Google Scholar]
- Kulshreshtha, G.; Rathgeber, B.; MacIsaac, J.; Boulianne, M.; Brigitte, L.; Stratton, G.; Thomas, N.A.; Critchley, A.T.; Hafting, J.; Prithiviraj, B. Feed supplementation with red seaweeds, Chondrus crispus and Sarcodiotheca gaudichaudii, reduce Salmonella Enteritidis in laying hens. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulshreshtha, G.; Rathgeber, B.; Stratton, G.; Thomas, N.; Evans, F.; Critchley, A.; Hafting, J.; Prithiviraj, B. Immunology, health, and disease: Feed supplementation with red seaweeds, Chondrus crispus and Sarcodiotheca gaudichaudii, affects performance, egg quality, and gut microbiota of layer hens. Poult. Sci. 2014, 93. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′ → 3′) |
|---|---|
| ramA | CGTCATGCGGGGTATTCCAAGTG CGCGCCGCCAGTTTTAGC |
| marA | ATCCGCAGCCGTAAAATGAC TGGTTCAGCGGCAGCATATA |
| acrB | TTTTGCAGGGCGCGGTCAGAATAC TGCGGTGCCCAGCTCAACGAT |
| 16SrRNA | GCGGCAGGCCTAACACAT GCAAGAGGCCCGAACGTC |
| tufA | TGTTCCGCAAACTGCTGGACG ATGGTGCCCGGCTTAGCCAGTA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulshreshtha, G.; Critchley, A.; Rathgeber, B.; Stratton, G.; Banskota, A.H.; Hafting, J.; Prithiviraj, B. Antimicrobial Effects of Selected, Cultivated Red Seaweeds and Their Components in Combination with Tetracycline, against Poultry Pathogen Salmonella Enteritidis. J. Mar. Sci. Eng. 2020, 8, 511. https://doi.org/10.3390/jmse8070511
Kulshreshtha G, Critchley A, Rathgeber B, Stratton G, Banskota AH, Hafting J, Prithiviraj B. Antimicrobial Effects of Selected, Cultivated Red Seaweeds and Their Components in Combination with Tetracycline, against Poultry Pathogen Salmonella Enteritidis. Journal of Marine Science and Engineering. 2020; 8(7):511. https://doi.org/10.3390/jmse8070511
Chicago/Turabian StyleKulshreshtha, Garima, Alan Critchley, Bruce Rathgeber, Glenn Stratton, Arjun H. Banskota, Jeff Hafting, and Balakrishnan Prithiviraj. 2020. "Antimicrobial Effects of Selected, Cultivated Red Seaweeds and Their Components in Combination with Tetracycline, against Poultry Pathogen Salmonella Enteritidis" Journal of Marine Science and Engineering 8, no. 7: 511. https://doi.org/10.3390/jmse8070511