Pro12Ala PPAR-γ2 and +294T/C PPAR-δ Polymorphisms and Association with Metabolic Traits in Teenagers from Northern Mexico

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Phenotyping
2.3. Genotyping
2.4. Statistical Analysis
3. Results
3.1. Population Characteristics and Metabolic Traits among Them
3.2. Pro12Ala PPAR-γ2 and +294T/C PPAR-δ
3.3. Metabolic Traits by Pro12Ala PPAR-γ2 and +294T/C PPAR-δ Polymorphism Genotype: Major Allele Homozygous vs. Minor Allele Carriers in the Total Studied Population
3.4. Association between Pro12Ala PPAR-γ2 and Metabolic Traits
3.5. Association between +294T/C PPAR-δ and Metabolic Traits
3.6. Logistic Regression
4. Discussion
5. Study Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Afshin, A.; Forouzanfar, M.H.; Reitsma, M.B.; Sur, P.; Estep, K.; Lee, A.; Marczak, L.; Mokdad, A.H.; Moradi-Lakeh, M.; Naghavi, M.; et al. Health effects of overweight and obesity in 195 countries over 25 years. New Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [PubMed]
- OECD. The Heavy Burden of Obesity; OECD Health Policy Studies, Ed.; OECD Publishing: Paris, France, 2019. [Google Scholar]
- Ogden, C.L.; Carroll, M.D.; Lawman, H.G.; Fryar, C.D.; Kruszon-Moran, D.; Kit, B.K.; Flegal, K.M. Trends in obesity prevalence among children and adolescents in the United States, 1988-1994 through 2013-2014. JAMA 2016, 315, 2292–2299. [Google Scholar] [CrossRef] [PubMed]
- Instituto Nacional de Salud Pública. Encuesta Nacional de Salud y Nutrición 2018. México: Instituto Nacional de Salud Pública. 2018. Available online: https://www.inegi.org.mx/contenidos/programas/ensanut/2018/doc/ensanut_2018_diseno_conceptual.pdf (accessed on 19 June 2020).
- Willson, T.M.; Brown, P.J.; Sternbach, D.D.; Henke, B.R. The PPARs: From orphan receptors to drug discovery. J. Med. Chem. 2000, 43, 527–550. [Google Scholar] [CrossRef]
- Hong, F.; Pan, S.; Guo, Y.; Xu, P.; Zhai, Y. PPARs as Nuclear Receptors for Nutrients and Energy Metabolism. Molecules 2019, 24, 2545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Jiménez, A.R.; Medh, J.D. Identification and regulation of novel PPAR-γ splice variants in human THP-1 macrophages. Biochim. Biophys. Acta 2006, 1759, 32–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouda, H.N.; Sagoo, G.S.; Harding, A.H.; Yates, J.; Sandhu, M.S.; Higgins, J.P.T. The association between the peroxisome proliferator-activated receptor-γ2 (PPARG2) Pro12Ala gene variant and type 2 diabetes mellitus: A HuGE review and meta-analysis. Am. J. Epidemiol. 2010, 171, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Swarbrick, M.M.; Chapman, C.M.L.; McQuillan, B.M.; Hung, J.; Thompson, P.L. A Pro12Ala polymorphism in the human peroxisome proliferator-activated receptor-γ2 is associated with combined hyperlipidaemia in obesity. Eur. J. Endocrinol. 2001, 144, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Stumvoll, M.; Häring, H. The Peroxisome Proliferator-Activated Receptor-γ2 Pro12Ala Polymorphism. Perspect. Diabetes 2002, 51, 2341–2347. [Google Scholar]
- Stryjecki, C.; Peralta-Romero, J.; Alyass, A.; Karam-Araujo, R.; Suarez, F.; Gomez-Zamudio, J.; Burguete-Garcia, A.; Cruz, M.; Meyre, D. Pro12Ala genotype and insulin resistance is modified by circulating lipids in Mexican children. Sci. Rep. 2016, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Yen, C.; Beamer, B.A.; Negri, C.; Silver, K.; Yarnall, D.P. Molecular Scanning of the Human Peroxisome Proliferator Activated Receptor γ (hPPARγ) Gene in Diabetic Caucasians: Identification of a Pro12Ala γ2 Missense Mutation. Biochem. Biophys. Res. Commun. 1997, 274, 270–274. [Google Scholar] [CrossRef]
- Galbete, C.; Toledo, E.; Martínez-González, M.A.; Martínez, J.A.; Guillén-Grima, F.; Marti, A. Pro12Ala variant of the PPARG2 gene increases body mass index: An updated meta-analysis encompassing 49,092 subjects. Obesity 2013, 21, 1486–1495. [Google Scholar] [CrossRef]
- Morris, A.; Voight, B.; Teslobich, T.; Ferreira, T.; Segre, A.; Steinthorsdottir, B.; Strawbridge, R.J.; Khan, H.; Grallert, H.; Mahajan, A.; et al. Large-scale association analysis provides insights into the genetic architecture and pathophysiology of type 2 diabetes. Nat. Genet. 2012, 44, 981–990. [Google Scholar] [PubMed]
- Deeb, S.S.; Fajas, L.; Nemoto, M.; Pihlajamäki, J.; Mykkänen, L.; Kuusisto, J.; Laakso, M.; Fujimoto, W.; Auwerx, J. A Pro12Ala substitution in PPARγ2 associated with decreased receptor activity, lower body mass index and improved insulin sensitivity. Nat. Genet. 1998, 20, 284–287. [Google Scholar] [CrossRef]
- Muller, Y.L.; Bogardus, C.; Beamer, B.A.; Shuldiner, A.R.; Baier, L.J. A Functional Variant in the Peroxisome Proliferator—Activated Receptor γ2 Promoter Is Associated with Predictors of Obesity and Type 2 Diabetes in Pima Indians. Diabetes 2003, 52, 1864–1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canizales-Quinteros, S.; Aguilar-Salinas, C.A.; Ortiz-López, M.G.; Rodríguez-Cruz, M.; Villarreal-Molina, M.T.; Coral-Vázquez, R.; Huertas-Vázquez, A.; Hernández-Caballero, A.; López-Alarcón, M.; Brito-Zurita, O.R.; et al. Association of PPARG2 Pro12Ala Variant with Larger Body Mass Index in Mestizo and Amerindian Populations of Mexico. Hum. Biol. 2007, 79, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Ben Ali, S.; Ben Yahia, F.; Sediri, Y.; Kallel, A.; Ftouhi, B. Gender-specific effect of Pro12Ala polymorphism in peroxisome proliferator-activated receptor γ-2 gene on obesity risk and leptin levels in a Tunisian population. Clin. Biochem. 2009, 42, 1642–1647. [Google Scholar] [CrossRef] [PubMed]
- Baldani, D.P.; Skrgatic, L.; Cerne, J.Z.; Ferk, P.; Simunic, V.; Gersak, K. Association of PPARG Pro12Ala polymorphism with insulin sensitivity and body mass index in patients with polycystic ovary syndrome. Biomed. Rep. 2014, 2, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Tönjes, A.; Stumvoll, M. The role of the Pro12Ala polymorphism in peroxisome proliferator-activated receptor γ in diabetes risk. Lippincott Williams & Wilkins. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 410–414. [Google Scholar]
- Information NC for B. PPARD peroxisome proliferator activated receptor delta [Homo sapiens (human)]. 2018. Available online: https://www.ncbi.nlm.nih.gov/gene/5467#gene-expression (accessed on 2 November 2018).
- Gouni-Berthold, I.; Giannakidou, E.; Faust, M.; Berthold, H.K.; Krone, W. The peroxisome proliferator-activated receptor delta +294T/C polymorphism in relation to lipoprotein metabolism in patients with diabetes mellitus type 2 and in non-diabetic controls. Atherosclerosis 2005, 183, 336–341. [Google Scholar] [CrossRef]
- Sznaidman, M.L.; Haffner, C.D.; Maloney, P.R.; Fivush, A.; Chao, E.; Goreham, D.; Sierra, M.L.; LeGrumelec, C.; Xu, H.E.; Montana, V.G.; et al. Novel selective small molecule agonists for peroxisome proliferator-activated receptor δ (PPARδ) - Synthesis and biological activity. Bioorg Med. Chem. Lett. 2003, 13, 1517–1521. [Google Scholar] [CrossRef]
- Skogsberg, J.; McMahon, A.D.; Karpe, F.; Hamsten, A.; Packard, C.J.; Ehrenborg, E. Peroxisome proliferator activated receptor delta genotype in relation to cardiovascular risk factors and risk of coronary heart disease in hypercholesterolaemic men. J. Intern. Med. 2003, 254, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.Y.; Liu, C.W.; Ge, L.; Pang, G.F.; Yang, M.; Hu, C.Y.; Ze-Ping, L.; Ning-Yuan, C.; Hai-Yan, L.; Hua-Yu, W.; et al. PPARD +294C overrepresentation in general and long-lived population in China Bama longevity area and unique relationships between PPARD +294T/C polymorphism and serum lipid profiles. Lipids Health Dis. 2015, 14, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aberle, J.; Hopfer, I.; Beil, F.U.; Seedorf, U. Association of the T+294C polymorphism in PPAR δ with low HDL cholesterol and coronary heart disease risk in women. Int. J. Med Sci. 2006, 3, 108–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burch, L.R.; Donnelly, L.A.; Doney, A.S.F.; Brady, J.; Tommasi, A.M.; Whitley, A.L.; Goddard, C.; Morris, A.D.; Hansen, M.K.; Palmer, C.N.A. Peroxisome proliferator-activated receptor-δ genotype influences metabolic phenotype and may influence lipid response to statin therapy in humans: A genetics of diabetes audit and research tayside study. J. Clin. Endocrinol. Metab. 2010, 95, 1830–1837. [Google Scholar] [CrossRef] [Green Version]
- Robitaille, J.; Gaudet, D.; Pérusse, L.; Vohl, M.-C. Features of the metabolic syndrome are modulated by an interaction between the peroxisome proliferator-activated receptor-delta -87T>C polymorphism and dietary fat in French-Canadians. Int. J. Obes. 2007, 31, 411–417. [Google Scholar]
- Aberle, J.; Hopfer, I.; Beil, F.U.; Seedorf, U. Association of peroxisome proliferator-activated receptor delta +294T/C with body mass index and interaction with peroxisome proliferator-activated receptor alpha L162V. Int. J. Obes. 2006, 30, 1709–1713. [Google Scholar] [CrossRef] [Green Version]
- Grarup, N.; Albrechtsen, A.; Ek, J.; Borch-Johnsen, K.; Jørgensen, T.; Schmitz, O.; Pedersen, O. Variation in the peroxisome proliferator-activated receptor δ gene in relation to common metabolic traits in 7,495 middle-aged white people. Diabetologia 2007, 50, 1201–1208. [Google Scholar] [CrossRef] [Green Version]
- Villegas, R.; Williams, S.; Gao, Y.; Cai, Q.; Li, H.; Elasy, T.; Cai, H.; Edwards, T.; Xiang, Y.B.; Zheng, W.; et al. Peroxisome proliferator-activated receptor (PPAR) delta genetic variation and type 2 diabetes in middle age Chinese women. Ann. Hum. Genet. 2011, 75, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Lü, Q.; Cao, H.; Yang, Q.; Tong, N. PPARD rs2016520 polymorphism is associated with metabolic traits in a large population of Chinese adults. Gene 2016, 585, 191–195. [Google Scholar] [CrossRef]
- Qian, Y.; Li, P.; Zhang, J.; Shi, Y.; Chen, K.; Yang, J.; Wu, Y.; Ye, X. Association between peroxisome proliferator-activated receptor-alpha, delta, and gamma polymorphisms and risk of coronary heart disease: A case-control study and meta-analysis. Medicine 2016, 95, 1–9. [Google Scholar] [CrossRef]
- Jguirim-Souissi, I.; Jelassi, A.; Hrira, Y.; Najah, M.; Slimani, A.; Addad, F.; Hassine, M.; Hamda, K.B.; Maatouk, F.; Rouis, M.; et al. +294T/C polymorphism in the PPAR-δ gene is associated with risk of coronary artery disease in normolipidemic Tunisians. Genet. Mol. Res. 2010, 9, 1326–1333. [Google Scholar] [CrossRef]
- Miao, L.; Yin, R.-X.; Wu, D.-F.; Cao, X.-L.; Li, Q.; Hu, X.-J.; Yan, T.-T.; Aung, L.H.H.; Yang, D.-Z.; Lin, W.-X.; et al. Peroxisome proliferator-activated receptor delta +294 > C polymorphism and serum lipid levels in the Guangxi Bai Ku Yao and Han populations. Lipids Health Dis. 2010, 9, 145–157. [Google Scholar]
- Rosales-Reynoso, M.A.; Wence-Chavez, L.I.; Arredondo-Valdez, A.R.; Dumois-Petersen, S.; Barros-Núñez, P.; Gallegos-Arreola, M.P.; Flores-Martínez, S.E.; Sánchez-Corona, J. Protective role of +294 T/C (rs2016520) polymorphism of PPARD in Mexican patients with colorectal cancer. Genet. Mol. Res. 2017, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; Weitzman, M.; Auinger, P.; Michael, N.; Dietz, W.H. Prevalence of a Metabolic Syndrome Phenotype in Adolescents. Arch. Pediatr. Adolescent Med. 2003, 157, 821–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mederico, M.; Paoli, M.; Zerpa, Y.; Camacho, N.; Cichetti, R.; Molina, Z.; Camachob, N.; Cichettib, R.; Molinab, Z.; Mora, Y.; et al. Valores de referencia de la circunferencia de la cintura e índice de la cintura/cadera en escolares y adolescentes de Mérida, Venezuela: Comparación con referencias internacionales. Endocrinol. Y Nutr. 2013, 60, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Mokha, J.S.; Srinivasan, S.R.; DasMahapatra, P.; Fernandez, C.; Chen, W.; Xu, J.; Berenson, G.S. Utility of waist-to-height ratio in assessing the status of central obesity and related cardiometabolic risk profile among normal weight and overweight/obese children: The Bogalusa Heart Study. BMC Pediatr. 2013, 10, 73. [Google Scholar]
- CDC. Defining Childhood Obesity. 2018. Available online: https://www.cdc.gov/obesity/childhood/defining.html (accessed on 19 June 2020).
- Must, A.; Anderson, S.E. Body mass index in children and adolescents: Considerations for population-based applications. Int. J. Obes. 2006, 30, 590–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Growth Reference 5-19 years. 2007. Available online: https://www.who.int/growthref/who2007_bmi_for_age/en/ (accessed on 19 June 2020).
- National Heart Lung and Blood Institute. Expert Panel on Integrated Guidelines for Cardiovascular Health and Risk Reduction in Children and Adolescents. 2012. Available online: http://www.nhlbi.nih.gov/files/docs/guidelines/peds_guidelines_full.pdf (accessed on 19 June 2020).
- Millán, J.; Pintó, X.; Muñoz, A.; Zúñiga, M.; Rubiés-Prat, J.; Pallardo, L.F.; Masana, L.; Mangas, A.; Hernández-Mijares, A.; González-Santos, P.; et al. Cocientes lipoproteicos: Significado fisiológico y utilidad clínica de los índices aterogénicos en prevención cardiovascular. Clin. E Investig. En Arterioscler. 2010, 22, 25–32. [Google Scholar] [CrossRef]
- Cossrow, N.; Falkner, B. Race/ethnic issues in obesity and obesity-related comorbidities. J. Clin. Endocrinol. Metab. 2004, 89, 2590–2594. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S.E.; Whitaker, R.C. Prevalence of obesity among US preschool children in different racial and ethnic groups. Arch. Pediatrics Adolesc. Med. 2009, 163, 344–348. [Google Scholar] [CrossRef] [Green Version]
- Taveras, E.M.; Gillman, M.W.; Kleinman, K.; Rich-Edwards, J.W.; Rifas-Shiman, S.L. Racial/Ethnic Differences in Early-Life Risk Factors for Childhood Obesity. Pediatrics 2010, 125, 686–695. [Google Scholar] [CrossRef] [Green Version]
- Salzano, F.M.; Sans, M. Interethnic admixture and the evolution of Latin American populations. Genetics Mol. Biol. 2014, 37 (Suppl. 1), 151–170. [Google Scholar] [CrossRef] [Green Version]
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of Obesity Among Adults and Youth: United States, 2015–2016; NCHS data brief; 2017; 288, pp. 1–8. Available online: https://www.cdc.gov/nchs/products/databriefs/db288.htm (accessed on 19 June 2020).
- Benítez-Hernández, Z.P.; Hernández-Torres, P.; Cabañas, M.D.; De La Torre-Díaz, M.D.L.; López-Ejeda, N.; Marrodán, M.D.; Cervantes-Borunda, M. Composición corporal, estado nutricional y alimentación en escolares Tarahumaras urbanos y rurales de Chihuahua, México. Nutr. Clínica Y Dietética Hosp. 2014, 34, 71–79. [Google Scholar]
- Salcedo-Rocha, A.L.; García de Alba, J.E.; Contreras-Marmolejo, M. Presión arterial en adolescentes mexicanos: Clasificación, factores de riesgo e importancia. Rev. De Salud Pública 2010, 12, 612–622. [Google Scholar]
- Cardoso-Saldaña, G.C.; Yamamoto-Kimura, L.; Medina-Urrutia, A.; Posadas-Sánchez, R.; Caracas-Portilla, N.A.; Posadas-Romero, C. Exceso de peso y síndrome metabólico en adolescentes de la Ciudad de México. Archivos de Cardiología de México 2010, 80, 12–18. [Google Scholar]
- Jackson, S.L.; Zhang, Z.; Wiltz, J.L.; Loustalot, F.; Ritchey, M.D.; Goodman, A.B.; Yang, Q. Hypertension Among Youths—United States, 2001–2016. Morb. Mortal. Wkly. Rep. 2018, 67, 758–762. [Google Scholar] [CrossRef]
- Romero-Velarde, E.; Campollo-Rivas, O.; De La Rosa, A.C.; Vásquez-Garibay, E.M.; Castro-Hernández, J.F.; Cruz-Osorio, R.M. Factores de riesgo de dislipidemia en niños y adolescentes con obesidad. Salud Pública De México 2007, 49, 103–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perea-Martínez, A.; Bárcena-Sobrino, E.; Rodríguez-Herrera, R.; Greenawalt-Rodríguez, S.; Carbajal-Rodríguez, L.; Zarco-Román, J. Obesidad y comorbilidades en niños y adolescentes asistidos en el Instituto Nacional de Pediatría. Acta Pediátrica de México 2009, 30, 167–174. [Google Scholar]
- Camarillo-Romero, E.; García, M.V.D.; Amaya-Chávez, A.; Huitrón-Bravo, G.; Majluf-Cruz, A. Dificultades en la clasificación del síndrome metabólico. El ejemplo de los adolescentes en México. Salud Pública de México 2010, 52, 524–527. [Google Scholar] [PubMed]
- Almeida, S.M.; Furtado, J.M.; Mascarenhas, P.; Ferraz, M.E.; Ferreira, J.C.; Monteiro, M.P.; Vilanova, M.; Ferraz, F.P. Association between LEPR, FTO, MC4R, and PPARG-2 polymorphisms with obesity traits and metabolic phenotypes in school-aged children. Endocrine 2018, 60, 466–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becer, E.; Çlrakoǧlu, A. Effect of the Pro12Ala polymorphism of the peroxisome proliferator-activated receptor γ2 gene on lipid profile and adipokines levels in obese subjects. Balk. J. Med Genet. 2017, 20, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Lagou, V.; Scott, R.A.; Manios, Y.; Chen, T.L.J.; Wang, G.; Grammatikaki, E.; Kortsalioudaki, C.; Liarigkovinos, T.; Moschonis, G.; Roma-Giannikou, E.; et al. Impact of peroxisome proliferator-activated receptors γ and δ on adiposity in toddlers and preschoolers in the GENESIS study. Obesity 2008, 16, 913–918. [Google Scholar] [CrossRef]
- Akhmetov, I.; Astranenkove, I.; Togozkin, V. Association of PPARD gene polymorphism with human physical performance. Mol. Biol. 2007, 41, 852–857. [Google Scholar]
- Shin, H.D.; Park, B.L.; Kim, L.H.; Jung, H.S.; Cho, Y.M.; Moon, M.K.; Park, Y.J.; Lee, H.K.; Park, K.S. Genetic Polymorphisms in Peroxisome Proliferator-Activated Receptor δ Associated with Obesity. Diabetes 2004, 53, 847–851. [Google Scholar] [CrossRef] [Green Version]
- Hasan, N.S.; Kamel, S.A.; Hamed, M.; Awadallah, E.; Rahman, A.H.A.; Musa, N.I.; Hussein, G.H.S. Peroxisome proliferator-activated receptor-γ polymorphism (rs1801282) is associated with obesity in Egyptian patients with coronary artery disease and type 2 diabetes mellitus. J. Genet. Eng. Biotechnol. 2017, 15, 409–414. [Google Scholar] [CrossRef]
- Hasstedt, S.J.; Ren, Q.F.; Teng, K.; Elbein, S.C. Effect of the peroxisome proliferator-activated receptor-γ2 Pro12Ala variant on obesity, glucose homeostasis, and blood pressure in members of familial type 2 diabetic kindreds. J. Clin. Endocrinol. Metab. 2001, 86, 536–541. [Google Scholar] [CrossRef] [Green Version]
- Douglas, J.A.; Erdos, M.R.; Watanabe, R.M.; Braun, A.; Johnston, C.L.; Oeth, P.; Mohlke, K.L.; Valle, T.T.; Ehnholm, C.; Buchanan, T.A.; et al. The Peroxisome Proliferator-Activated Receptor-γ2 Pro12Ala Variant Association with Type 2 Diabetes and Trait Differences. Diabetes 2001, 50, 886–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Esparragón, F.J.; Rodríguez-Pérez, J.C.; Macías-Reyes, A.; Alamo-Santana, F. Peroxisome proliferator-activated receptor-gamma2-Pro12Ala and endothelial nitric oxide synthase-4a/b gene polymorphisms are associated with essential hypertension. J. Hypertens. 2003, 21, 1649–1655. [Google Scholar] [CrossRef] [PubMed]
- Al-Safar, H.; Hassoun, A.; Almazrouei, S.; Kamal, W.; Afandi, B.; Rais, N. Association of the genetic polymorphisms in transcription factor 7-like 2 and peroxisome proliferator-activated receptors-γ2 with type 2 diabetes mellitus and its interaction with obesity status in Emirati population. J. Diabetes Res. 2015, 2015, 129695. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Jia, W.; Fang, Q.; Zhang, R.; Wang, C.; Lu, J.; Xiang, K. Peroxisome proliferator-activated receptor (PPAR) delta genetic polymorphism and its association with insulin resistance index and fasting plasma glucose concentrations in Chinese subjects. Diabet. Med. 2006, 23, 1307–1312. [Google Scholar] [CrossRef]
- Ye, H.; Li, Y.; Hong, Q.; Zhou, A.; Zhao, Q.; Xu, L.; Xu, M.Q.; Xu, X.T.; Tang, L.L.; Dai, J.; et al. Positive association between PPARD rs2016520 polymorphism and coronary heart disease in a Han Chinese population. Genet. Mol. Res. 2015, 14, 12254–12261. [Google Scholar] [CrossRef] [PubMed]
- Sözen, M.A.; Özcan, M.U.; Çıldır, M.; Doğru, I.H.; Aygök, A.G.; Balkan, K. Association of the human PPARγ2 Pro12Ala polymorphism with obesity in a population from Turkey. Acta Endocrinol. 2018, 14, 459–465. [Google Scholar]
- Meirhaeghe, A.; Fajas, L.; Helbecque, N.; Cottel, D.; Auwerx, J.; Deeb, S.S.; Amouyel, P. Impact of the Peroxisome Proliferator Activated Receptor γ2 Pro12Ala polymorphism on adiposity, lipids and non-insulin-dependent diabetes mellitus. Int. J. Obes. 2000, 24, 195–199. [Google Scholar] [CrossRef] [Green Version]
- Masud, S.; Ye, S. Effect of the peroxisome proliferator activated receptor-y gene Pro12Ala variant on body mass index: A meta-analysis. J. Med. Genetics 2003, 40, 773–780. [Google Scholar] [CrossRef] [Green Version]
- Mansoori, A.; Amini, M.; Kolahdooz, F.; Seyedrezazadeh, E. Obesity and Pro12Ala polymorphism of peroxisome proliferator-activated receptor-gamma gene in healthy adults: A systematic review and meta-analysis. Ann. Nutr. Metab. 2015, 67, 104–118. [Google Scholar] [CrossRef]
- Luo, W.; Guo, Z.; Wu, M.; Hao, C.; Hu, X.; Zhou, Z.; Zhou, Z.; Yao, X.; Zhang, L.; Liu, J. Association of Peroxisome Proliferator-Activated Receptor α/δ/γ with Obesity, and Gene-Gene Interaction, in the Chinese Han Population. J. Epidemiol. 2013, 23, 187–194. [Google Scholar] [CrossRef] [Green Version]
 female,
female,  male.
female, male.
male.
female, male.

{kind=link}
| Characteristic | Mestizo-1 n = 96 (23%) | Mestizo-2 n = 37 (9%) | Tarahumara n = 173 (42%) | Mennonite n = 106 (26%) | Total (n = 412) |
|---|---|---|---|---|---|
| Md (IQR) | Md (IQR) | Md (IQR) | Md (IQR) | Md (IQR) | |
| Age (years) | 13 (13–14) a | 13 (12–13) a | 14 (13–15) b | 15 (14–15) c | 14 (13–15) |
| Weight (kg) | 47 (43–56) a | 59 (46–69) b,d | 48 (42–55) a | 57 (51–66) c,d | 51 (44–59) |
| Height (cm) | 158 (152–164) a | 157 (152–162) a | 152 (148–158) b | 171 (164–175) c | 158 (151–166) |
| BMI (kg/m2) | 19 (17–21) a | 24 (20–25) b | 20 (18–23) c,d | 20 (18–22) a,d | 20 (18–23) |
| n (%) | n (%) | n (%) | n (%) | n (%) | |
| Sex | |||||
| Male | 41 (43)a | 21 (57)a | 52 (30)b | 47 (44)a | 161 (39) |
| Female | 55 (57) | 16 (43) | 121 (70) | 59 (56) | 251 (61) |
| pBMI ≥ 85th (kg/m2) | 17 (18)a | 26 (70)b | 30 (17)a | 12 (11)a | 85 (21) |
| Characteristic | Mestizo-1 | Mestizo-2 | Tarahumara | Mennonite | Total |
|---|---|---|---|---|---|
| Mean ± SD/Md (IQR) | Mean ± SD/Md (IQR) | Mean ± SD/Md (IQR) | Mean ± SD/Md (IQR) | Mean ± SD/Md (IQR) | |
| BMI (Z-score) | 0.05 (−0.66–0.70) a | 1.45 (.75–1.64) b | 0.41 (-0.19–0.88) a,c | -0.05 (-0.63–0.77) a,d | 0.25 (-0.50–0.92) |
| SBP (mmHg) | 110 ± 11 a | 115 ± 11 a | 108 ± 10 a | 108 ± 10 a | 112 ± 10 |
| DBP (mmHg) | 70 (63–80) a | 70 (65–80) a,c | 70 (60–75) a | 75 (68–80) b,c | 70 (64–80) |
| WC (cm) | 64 (61–70) a | 80 (72–85) b,e | 70 (65–75) c | 68 (64–74) d,e | 68 (63–74) |
| WHR | 0.41 (0.38–0.44) a | 0.52 (0.47–0.54) b | 0.46 (0.43–0.49) c | 0.40 (0.38–0.42) a | 0.43 (0.40–0.48) |
| Glucose (mg/dL) | 87 ± 10 a | 75 ± 6 b | 83 ± 11 a | 78 ± 9 c | 82 ± 10 |
| TG (mg/dL) | 77 (57–109) a | 93 (72–129) a,c | 92 (75–115) b,c | 75 (60–92) a,d | 83 (64–111) |
| TC (mg/dL) | 145 ± 22 a | 156 ± 26 a | 133 ± 25 a | 144 ± 21 a | 140 ± 24 |
| HDL (mg/dL) | 48 (40–55) a | 51 (44–56) a | 37 (32–42) b | 50 (43–56) a | 43 (36–52) |
| LDL (mg/dL) | 79 ± 17 a | 84 ± 19 a | 76 ± 20 a | 77 ± 16 a | 78 ± 18 |
| VLDL (mg/dL) | 15 (11–22) a | 19 (14–26) a,c | 18 (15–23) b,c | 15 (12–18) a,d | 17 (13–22) |
| AI (Index) | 3.01 (2.59–3.48) a | 3.02 (2.55–3.74) a | 3.59 (3.12–4.20) b | 2.90 (2.56–3.19) a | 3.17 (2.76–3.69) |
| PPAR-γ2 | Mestizo-1 n (%) | Mestizo-2 n (%) | Tarahumara n (%) | Mennonite n (%) | Total n (%) |
|---|---|---|---|---|---|
| CC | 79 (82) a | 25 (68) a,c | 103 (60) b,c | 83 (78) a | 290 (70) |
| CG | 14 (15) | 10 (27) | 61 (35) | 20 (19) | 105 (26) |
| GG | 3 (3) | 2 (5) | 9 (5) | 3 (3) | 17 (4) |
| HWE p | 0.09 | 0.75 | 1 | 0.45 | 0.18 |
| C | 172 (90) a | 60 (81) a,c | 267 (77) b,c | 186 (88) a | 685 (83) |
| G | 20 (10) | 14 (19) | 79 (23) | 26 (12) | 139 (17) |
| PPAR-δ | Mestizo-1 n (%) | Mestizo-2 n (%) | Tarahumara n (%) | Mennonite n (%) | Total n (%) |
|---|---|---|---|---|---|
| TT | 62 (65) a | 23 (62) a | 153 (88) b | 81 (76) a | 319 (77) |
| TC | 33 (34) | 13 (35) | 20 (12) | 23 (22) | 89 (22) |
| CC | 1 (1) | 1 (3) | 0 (0) | 2 (2) | 4 (1) |
| HWE p | 0.32 | 0.88 | 0.71 | 0.97 | 0.72 |
| T | 157 (82) a | 59 (80) a | 326 (94) b | 185 (87) a | 727 (88) |
| C | 35 (18) | 15 (20) | 20 (6) | 27 (13) | 97 (12) |
| Metabolic Traits | Pro12Ala PPAR-γ2 | +294T/C PPAR-δ | ||||
|---|---|---|---|---|---|---|
| CC (n = 290) | CG/GG (n = 122) | TT (n = 319) | TC/CC (n = 93) | |||
| Mean ± SD/Md (IQR) | Mean ± SD/Md (IQR) | p | Mean ± SD/Md (IQR) | Mean ± SD/Md (IQR) | p | |
| BMI (Z-score) | 0.22 (−0.54–0.99) | 0.28 (-0.37–0.77) | 0.46 # | 0.21 (-0.52–0.87) | 0.39 (−0.41–1.18) | 0.06 # |
| SBP (mmHg) | 111 ± 11 | 109 ± 10 | 0.08 * | 110 ± 11 | 112 ± 11 | 0.15 * |
| DBP (mmHg) | 70 (65–80) | 70 (60–78) | 0.10 # | 70 (60–80) | 70 (65–78) | 0.78 # |
| WC (cm) | 68 (63–75) | 69 (65–74) | 0.37 # | 68 (63–74) | 69 (65–77) | 0.30 # |
| WHR | 0.42 (0.39–0.48) | 0.44 (0.41–0.48) | 0.04 # | 0.43 (0.40–0.48) | 0.42 (0.40–0.47) | 0.82 # |
| Glucose (mg/dL) | 82 ± 10 | 82 ± 10 | 0.90 * | 82 ± 11 | 80 ± 8 | 0.07 * |
| TG (mg/dL) | 83 (64–112) | 87 (66–110) | 0.51 # | 87 (68–112) | 75 (59–109) | 0.01 # |
| TC (mg/dL) | 142 ± 24 | 138 ± 24 | 0.14 * | 140 ± 25 | 143 ± 21 | 0.25 * |
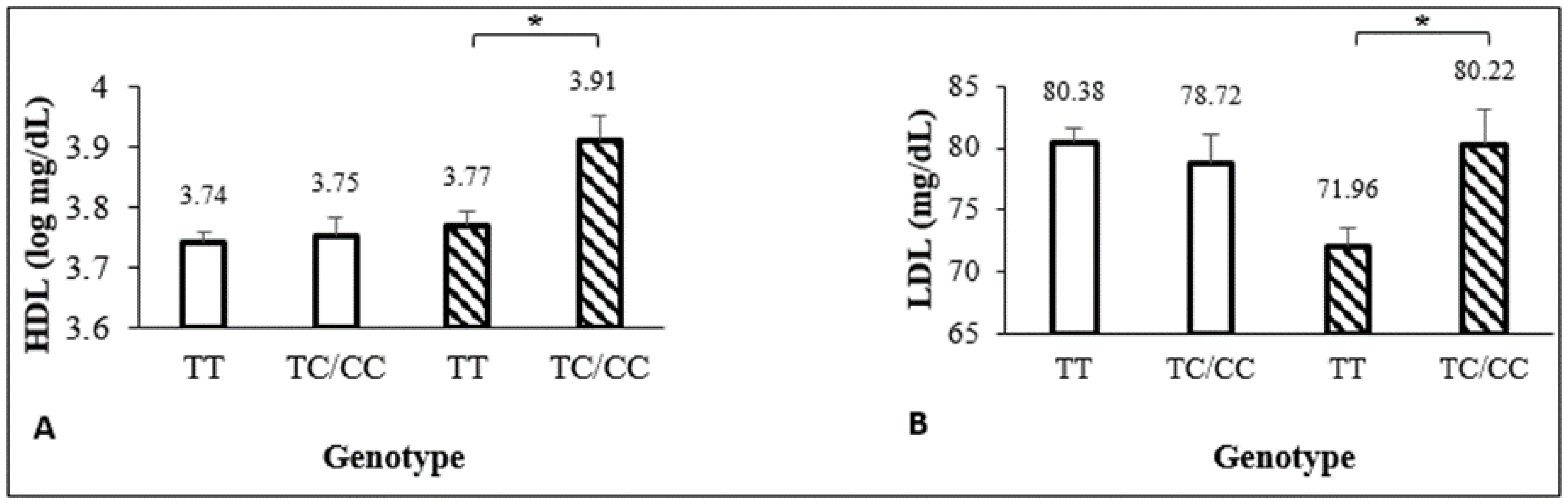
| HDL (mg/dL) | 45 (37–52) | 39 (35–48) | <0.01 # | 43 (36–51) | 48 (36–54) | 0.03 # |
| LDL (mg/dL) | 78 ± 19 | 76 ± 17 | 0.37 * | 77 ± 19 | 80 ± 16 | 0.24 * |
| VLDL (mg/dL) | 17 (13–22) | 17 (13–22) | 0.51 # | 17 (14–22) | 15 (12–22) | 0.01 # |
| AI (Index) | 3.14 (2.70–3.66) | 3.23 (2.82–3.77) | 0.15 # | 3.18 (2.81–3.68) | 3.12 (2.66–3.70) | 0.37 # |
| Metabolic Trait | Mestizo-1 | Mestizo-2 | Tarahumara | Mennonite | Total | Total (Adjusted) |
|---|---|---|---|---|---|---|
| n = (96) | n = (37) | n = (173) | n = (106) | n = (412) | n = (412) | |
| β ± SE | β ± SE | β ± SE | β ± SE | β ± SE | β ± SE | |
| (p) | (p) | (p) | (p) | (p) | (p) | |
| SBP (mmHg) * | −5.59 ± 2.60 | −0.62 ± 3.03 | −1.37 ± 1.44 | 0.61 ± 2.64 | −2.05 ± 1.07 | −1.41 ± 1.08 (0.191) *, ++ |
| (0.035) | (0.840) | (0.341) | (0.817) | (0.057) + | ||
| DBP (mmHg) * | −2.14 ± 2.34 | −0.70 ± 3.36 | −0.34 ± 1.37 | 1.25 ± 2.10 | −1.68 ± 0.98 | −0.73 ± 0.98 (0.455) ‡ |
| (0.363) | (0.835) | (0.802) | (0.554) | (0.086) | ||
| Glucose (mg/dL) * | −4.77 ± 2.71 | 1.74 ± 2.20 | 0.73 ± 1.53 | 1.46 ± 1.81 | −18.77 ± 9.20 | −13.82 ± 8.48 (0.104) *, -- |
| (0.082) | (0.436) | (0.632) | (0.423) | (0.042)- | ||
| TG (log mg/dL) * | −0.02 ± 0.11 | −0.07 ± 0.14 | −0.03 ± 0.05 | 0.10 ± 0.09 | 0.03 ± 0.04 | -0.008 ± 0.04 (0.84) § |
| (0.863) | (0.654) | (0.586) | (0.276) | (0.475) | ||
| TC (mg/dL) # | −4.07 ± 5.85 | −10.67 ± 9.27 | −1.66 ± 3.74 | 8.01 ± 4.57 | −3.91 ± 2.59 | −1.48 ± 2.53 (0.558) ¶ |
| (0.489) | (0.258) | (0.658) | (0.083) | (0.132) | ||
| HDL (log mg/dL) ^ | −0.11 ± 0.05 | <−0.01 ± 0.08 | 0.03 ± 0.03 | 0.01 ± 0.04 | −0.06 ± 0.03 | −0.005 ± 0.02 (0.825) ‡ |
| (0.033) | (0.961) | (0.250) | (0.815) | (0.031) ~ | ||
| LDL (mg/dL) * | 2.07 ± 4.71 | −9.94 ± 6.74 | −2.77 ± 2.87 | 4.98 ± 3.66 | −1.94 ± 1.92 | −9.63 ± 6.19 (0.119) ^ |
| (0.660) | (0.150) | (0.336) | (0.176) | (0.312) | ||
| VLDL (log mg/dL) * | −0.02 ± 0.11 | −0.07 ± 0.14 | −0.03 ± 0.05 | 0.10 ± 0.09 | 0.03 ± 0.04 | −0.28 ± 0.78 (0.720) § |
| (0.863) | (0.654) | (0.586) | (0.276) | (0.475) | ||
| AI (log) ^ | 0.08 ± 0.05 | −0.07 ± 0.07 | −0.05 ± 0.03 | 0.05 ± 0.04 | 0.40 ± 0.19 | 0.39 ± 0.17 (0.021) ^, // |
| (0.114) | (0.367) | (0.084) | (0.244) | (0.033) / |
| Metabolic Trait | Mestizo-1 | Mestizo-2 | Tarahumara | Mennonite | Total | Total (Adjusted) |
|---|---|---|---|---|---|---|
| n = (96) | n = (37) | n = (173) | n = (106) | n = (412) | n = (412) | |
| β ± SE | β ± SE | β ± SE | β ± SE | β ± SE | β ± SE | |
| (p) | (p) | (p) | (p) | (p) | (p) | |
| SBP (mmHg) * | 2.07 ± 2.09 | 0.74 ± 2.97 | 1.71 ± 2.26 | −1.54 ± 2.55 | 1.94 ± 1.19 | 1.34 ± 1.20 |
| (0.326) | (0.806) | (0.451) | (0.546) | (0.103) | (0.263) * | |
| DBP (mmHg) # | −0.25 ± 1.84 | −0.38 ± 3.25 | 0.29 ± 2.10 | −0.67 ± 2.03 | 0.85 ± 1.08 | 0.01 ± 1.08 |
| (0.894) | (0.907) | (0.889) | (0.742) | (0.432) | (0.994) # | |
| Glucose (mg/dL) * | −2.62 ± 2.15 | 0.04 ± 2.17 | −3.91 ± 2.39 | −0.75 ± 1.75 | −2.57 ± 1.18 | −2.22 ± 1.12 (0.048) * |
| (0.227) | (0.984) | (0.103) | (0.669) | (0.031) | ||
| TG (log mg/dL) * | −0.10 ± 0.09 | −0.12 ± 0.14 | −0.07 ± 0.08 | −0.15 ± 0.09 | −0.12 ± 0.05 | −0.08 ± 0.05 (0.08) # |
| (0.273) | (0.415) | (0.374) | (0.086) | (0.008) | ||
| TC (mg/dL) ^ | −0.002 ± 4.60 | 2.12 ± 9.35 | −4.89 ± 5.87 | −5.76 ± 4.48 | 79.02 ± 29.61 | 49.8 ± 28.9 (0.085) &, ++ |
| (1) | (0.822) | (0.406) | (0.202) | (0.008) + | ||
| HDL (log mg/dL) * | 0.02 ± 0.04 | 0.11 ± 0.07 | −0.10 ± 0.05 | 0.01 ± 0.04 | <0.01 ± 0.04 | 0.08 ± 0.07 (0.256) ‡ |
| (0.594) | (0.144) | (0.046) | (0.685) | (0.904) - | ||
| LDL (mg/dL) * | −6.75 ± 4.64 | 2.66 ± 6.6 | 0.47 ± 4.52 | −4.15 ± 3.49 | −1.67 ± 2.66 | −2.89 ± 2.68 (0.281) ‡,~~ |
| (0.149) / | (0.688) | (0.917) ^ | (0.238) & | (0.531) ‡,~ | ||
| VLDL (log mg/dL) * | 0.10 ± 0.09 | −0.12 ± 0.14 | −0.07 ± 0.08 | −0.15 ± 0.09 | −0.12 ± 0.05 | −0.08 ± 0.05 (0.082) # |
| (0.273) | (0.415) | (0.374) | (0.086) | (0.008) | ||
| AI (log) * | −0.02 ± 0.04 | −0.08 ± 0.07 | 0.08 ± 0.05 | −0.06 ± 0.04 | −0.03 ± 0.02 | −0.03 ± 0.06 (0.599) # |
| (0.643) | (0.223) | (0.093) | (0.133) | (0.208) |
| Metabolic Traits | Pro12Ala PPAR-γ2 | +294T/C PPAR-δ | ||||||
|---|---|---|---|---|---|---|---|---|
| CC | CG/GG | Adjusted OR (95%CI) | p | TT | C/CC | Adjusted OR (95%CI) | p | |
| (n = 290) | (n = 122) | (n = 319) | (n = 93) | |||||
| n (%) | n (%) | n (%) | n (%) | |||||
| BMI (Z-score > 1.04) | 68 (23) | 15 (12) | 0.46 (0.25–0.84) | 0.011 | 57 (18) | 26 (28) | 1.78 (1.04–3.05) | 0.034 |
| pBP ≥ 90th | 100 (34) | 33 (27) | 0.70 (0.44–1.12) | 0.142 | 103 (32) | 30 (32) | 1 (0.61–1.64) | 0.996 |
| pWC ≥ 90th | 9 (3) | 5 (4) | 1.33 (0.44–4.07) | 0.612 | 9 (3) | 5 (5) | 1.96 (0.64–5.99) | 0.239 |
| WHR > 0.45 * | 108 (37) | 55 (45) | 1.79 (1.03–3.08) | 0.036 ¶ | 131 (41) | 32 (34) | 0.74 (0.45–1.21) | 0.231 |
| Glucose ≥ 110 (mg/dL) ** | 4 (1) | 1 (1) | -- | 5 (2) | 0 (0) | -- | ||
| TG ≥ 110 ^ (mg/dL) | 76 (26) | 30 (25) | 0.91 (0.56–1.50) | 0.721 | 84 (26) | 22 (24) | 0.77 (0.44–1.34) | 0.350 |
| TC ≥ 150 # (mg/dL) | 41 (14) | 9 (7) | 0.47 (0.22–1) | 0.050 - | 39 (12) | 11 (12) | 0.87 (0.43–1.81) | 0.726 ~ |
| HDL ≤ 40 @ (mg/dL) | 92 (32) | 65 (53) | 2.50 (1.60–3.89) | <0.01 | 128 (40) | 29 (31) | 1 (0.55–1.82) | 0.995 / |
| LDL ≥ 110 ‡ (mg/dL) | 13 (4) | 3 (2) | 0.54 (0.15–1.96) | 0.351 | 15 (5) | 1 (1) | 0.20 (0.03–1.52) | 0.118 |
| VLDL ≥ 30 ‡ (mg/dL) | 22 (8) | 7 (6) | 0.74 (0.31–1.80) | 0.514 | 23 (7) | 6 (6) | 0.82 (0.32–2.10) | 0.679 |
| AI (M > 4 F > 3.5) & | 86 (30) | 39 (32) | 1.07 (0.64–1.78) | 0.798 | 99 (31) | 26 (28) | 0.80 (0.45–1.41) | 0.435 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrillo-Venzor, M.A.; Erives-Anchondo, N.R.; Moreno-González, J.G.; Moreno-Brito, V.; Licón-Trillo, A.; González-Rodríguez, E.; Hernández-Rodríguez, P.d.C.; Reza-López, S.A.; Loera-Castañeda, V.; Leal-Berumen, I. Pro12Ala PPAR-γ2 and +294T/C PPAR-δ Polymorphisms and Association with Metabolic Traits in Teenagers from Northern Mexico. Genes 2020, 11, 776. https://doi.org/10.3390/genes11070776
Carrillo-Venzor MA, Erives-Anchondo NR, Moreno-González JG, Moreno-Brito V, Licón-Trillo A, González-Rodríguez E, Hernández-Rodríguez PdC, Reza-López SA, Loera-Castañeda V, Leal-Berumen I. Pro12Ala PPAR-γ2 and +294T/C PPAR-δ Polymorphisms and Association with Metabolic Traits in Teenagers from Northern Mexico. Genes. 2020; 11(7):776. https://doi.org/10.3390/genes11070776
Chicago/Turabian StyleCarrillo-Venzor, Martín A., Nancy R. Erives-Anchondo, Janette G. Moreno-González, Verónica Moreno-Brito, Angel Licón-Trillo, Everardo González-Rodríguez, Pilar del Carmen Hernández-Rodríguez, Sandra A. Reza-López, Verónica Loera-Castañeda, and Irene Leal-Berumen. 2020. "Pro12Ala PPAR-γ2 and +294T/C PPAR-δ Polymorphisms and Association with Metabolic Traits in Teenagers from Northern Mexico" Genes 11, no. 7: 776. https://doi.org/10.3390/genes11070776