Endozoochory by Goats of Two Invasive Weeds with Contrasted Propagule Traits

 , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Species
2.2. Fruit Collection and Seed Characteristics
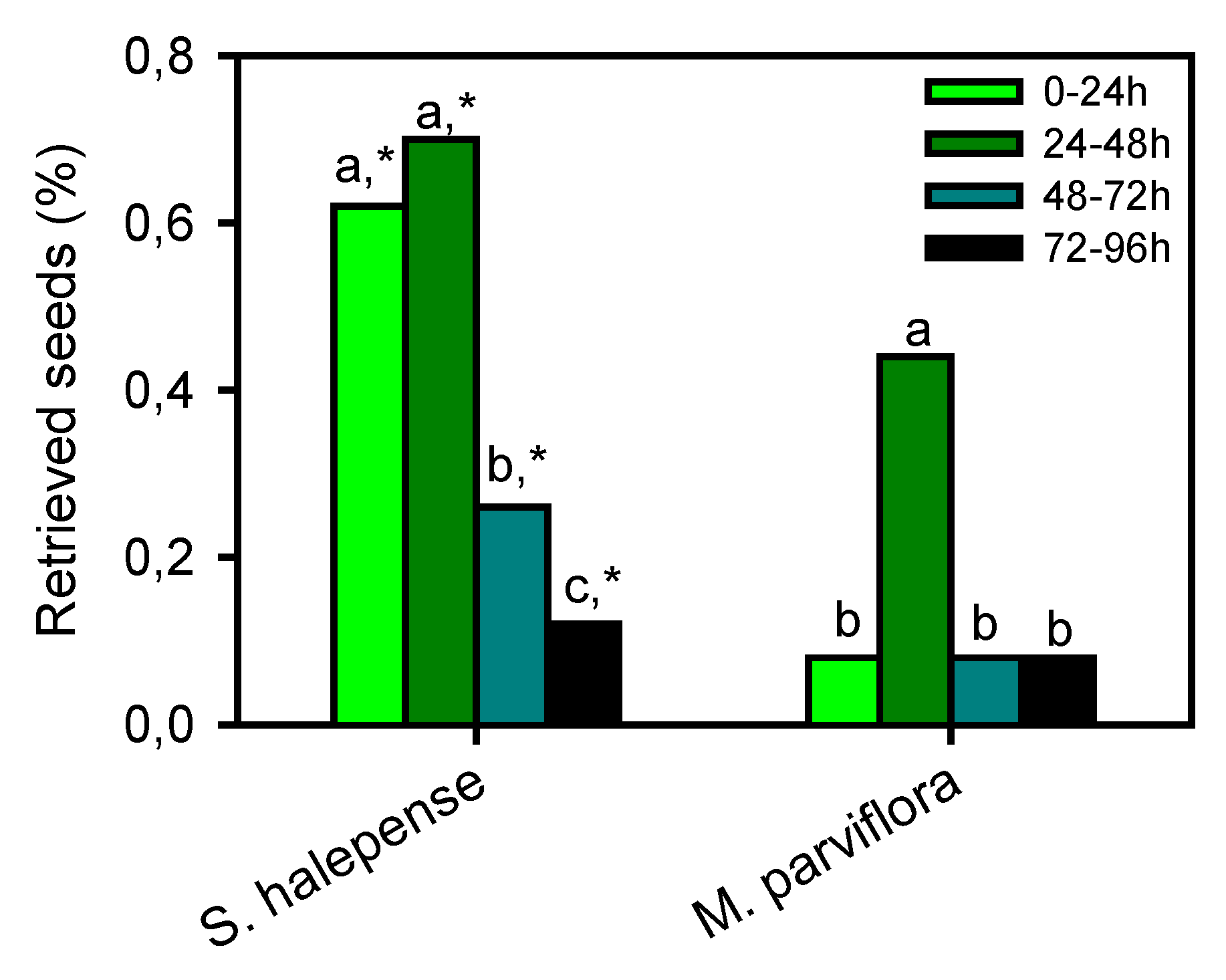
2.3. Seeds Retrieved after Gut Passage
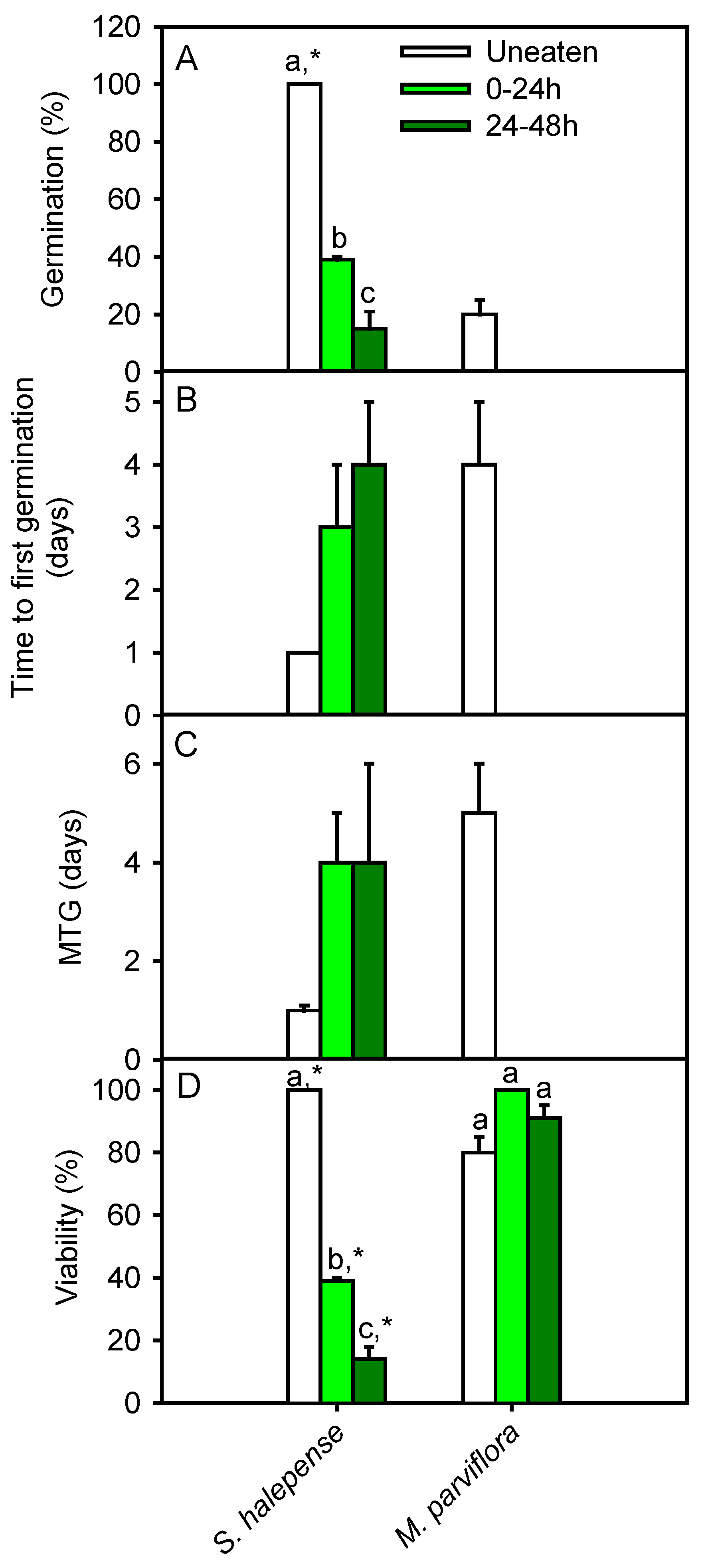
2.4. Seed Germination after Gut Passage
2.5. Data Analyses
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rai, P.K.; Singh, J.S. Invasive alien plant species: Their impact on environment, ecosystem services and human health. Ecol. Indic. 2020, 111, 106020. [Google Scholar]
- Iqbal, N.; Bajwa, A.A.; Manalil, S.; Khan, A.M.; Kebaso, L.; Frimpong, D.; Ali, H.H.; Jha, P.; Chauhan, B.S. Biology and management of two Hordeum weedy species: A review. Crop Prot. 2019, 125, 104908. [Google Scholar] [CrossRef]
- Novoa, A.; Richardson, D.M.; Pysek, P.; Meyerson, L.A.; Bacher, S.; Canavan, S.; Catford, J.A.; Cuda, J.; Essl, F.; Foxcroft, L.C. Invasion syndromes: A systematic approach for predicting biological invasions and facilitating effective management. Biol. Invasions 2020, 22, 1801–1820. [Google Scholar] [CrossRef] [Green Version]
- Chuang, A.; Peterson, C.R. Expanding population edges: Theories, traits, and trade-offs. Glob. Change Biol. 2016, 22, 494–512. [Google Scholar] [CrossRef]
- Gioria, M.; Pysek, P. Early bird catches the worm: Germination as a critical step in plant invasion. Biol. Invasions 2017, 19, 1055–1080. [Google Scholar] [CrossRef] [Green Version]
- Howe, H.F.; Smallwod, J. Ecology of seed dispersal. Ecol. Evol. Syst. Rev. Annu. 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Gokbulak, F.; Call, C.A. Grass seedling recruitment in cattle dungpats. Rangel. Ecol. Manag. 2004, 57, 649–655. [Google Scholar] [CrossRef]
- Gallandt, E.R. How can we target the weed seedbank? Weed Sci. 2006, 54, 588–596. [Google Scholar] [CrossRef]
- Paulose, J.; Hallatschek, O. The impact of long-range dispersal on gene surfing. Proc. Natl. Acad. Sci. USA 2020, 117, 7584–7593. [Google Scholar] [CrossRef]
- Lessa, L.G.; Geise, L.; Costa, F.N. Effects of gut passage on the germination of seeds ingested by didelphid marsupials in a neotropical savanna. Acta Bot. Bras. 2013, 27, 519–525. [Google Scholar] [CrossRef] [Green Version]
- Verdú, M.; Traveset, A. Early emergence enhances plant fitness: A phylogenetically controlled meta-analysis. Ecology 2005, 86, 1385–1394. [Google Scholar] [CrossRef]
- Zwolak, R. How intraspecific variation in seed-dispersing animals matters for plants. Biol. Invasion 2018, 93, 897–913. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.L.; Wang, D.; Wang, S.P.; Bonser, F.; Sun, Y.; Zhou, Y.; Gao, X.T. Plants can benefit from herbivory: Stimulatory effects of sheep saliva on growth of Leymus chinensis. PLoS ONE 2012, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Gardener, C.J.; McIvor, J.G.; Jansen, A. Survival of seeds of tropical grassland species subjected to bovine digestion. J. Appl. Ecol. 1993, 30, 75–85. [Google Scholar] [CrossRef]
- Mancilla-Leytón, J.M.; Fernández-Alés, R.; Martín Vicente, A. Plant-ungulate interaction: Goat gut passage effect on survival and germination of Mediterranean shrub seeds. J. Veg. Sci. 2011, 22, 1031–1037. [Google Scholar] [CrossRef]
- Albert, A.; Marell, A.; Picard, M.; Baltzinger, C. Using basic plant traits to predict ungulate seed dispersal potential. Ecography 2015, 38, 440–449. [Google Scholar] [CrossRef]
- Archer, S.; Pyke, D.A. Plant-animal interactions affecting plant establishment and persistence on revegetated rangeland. J. Range Manag. 1991, 44, 558–565. [Google Scholar] [CrossRef]
- Gökbulak, F. Recovery and germination of grass seeds ingested by cattle. J. Biol. Sci. 2006, 6, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Kettenring, K.M.; Adams, C.R. Lessons learned from invasive plant control experiments: A systematic review and meta-analysis. J. Appl. Ecol. 2011, 48, 970–979. [Google Scholar] [CrossRef]
- Shapero, M.; Huntsinger, L.; Becchetti, T.; Mashiri, F.; James, J. Land manager perceptions of opportunities and constraints of using livestock to manage invasive plants. Rangel. Ecol. Manag. 2018, 71, 603–611. [Google Scholar] [CrossRef]
- Whitacre, M.K.; Call, C.A. Recovery and germinability of native seed fed to cattle. West. N. Am. Nat. 2006, 66, 121–128. [Google Scholar] [CrossRef]
- Kuiters, A.T.; Huiskes, H.P.J. Potential of endozoochorous seed dispersal by sheep in calcareous grasslands: Correlations with seed traits. Appl. Veg. Sci. 2010, 13, 163–172. [Google Scholar] [CrossRef]
- Grande, D.; Mancilla-Leytón, J.M.; Martín Vicente, A.; Delgado-Pertíñez, M. Can goats disperse seeds of herbaceous pasture plants in Mediterranean grasslands? Small Rumin. Res. 2016, 143, 67–74. [Google Scholar] [CrossRef]
- Hart, S. Recent perspectives in using goats for vegetation management in the USA. J. Dairy Sci. 2001, 84, 170–176. [Google Scholar] [CrossRef]
- Michael, P.J.; Steadman, K.J.; Plummer, J.A. Climatic regulation of seed dormancy and emergence of diverse Malva parviflora populations from a Mediterranean-type environment. Seed Sci. Res. 2006, 16, 273–281. [Google Scholar] [CrossRef]
- Michael, P.J.; Steadman, K.J.; Plummer, J.A. Seed development in Malva parviflora: Onset of germinability, dormancy and desiccation tolerance. Aust. J. Exp. Agric. 2007, 47, 683–688. [Google Scholar] [CrossRef]
- Wan, J.Z.; Wang, C.J. Contribution of environmental factors toward distribution of ten most dangerous weed species globally. App. Ecol. Environ. Res. 2019, 17, 14835–14846. [Google Scholar] [CrossRef]
- Kaur, R.; Soodan, A.S. Reproductive biology of Sorghum halepense (L.) Pers. (Poaceae; Panicoideae; Andropogoneae) in relation to invasibility. Flora 2017, 229, 32–49. [Google Scholar] [CrossRef]
- Boulos, L. Flora of Egypt: Monocotyledons (Alismataceae-Orchidaceae); Al Hadara Publishing: Cairo, Egypt, 2005; Volume 4, p. 617. [Google Scholar]
- Fryxell, P.A. Malvaceae of México (Systematic Botany Monographs); The American Society of Plant Taxonomists: Ann Arbor, MI, USA, 1988. [Google Scholar]
- Chauhan, B.S.; Gill, G.; Preston, C. Factors affecting seed germination of little mallow (Malva parviflora) in southern Australia. Weed Sci. 2006, 54, 1045–1050. [Google Scholar] [CrossRef]
- Daugovish, O.; Mochizuki, M.; Fennimore, S. Control of difficult weeds for California strawberry production. Acta Hortic. 2009, 842, 207–210. [Google Scholar] [CrossRef]
- Al-Johani, N.S.; Aytah, A.A.; Boutraa, T. Allelopathic impact of two weeds, Chenopodium murale and Malva parviflora on growth and photosynthesis of barley (Hordeum vulgare L.). Pak. J. Bot. 2012, 44, 1865–1872. [Google Scholar]
- Badry, M.O. Biosystematic Studies on Some Genera of Malvaceae (S.L.). Ph.D. Thesis, South Valley University, Qena Governorate, Egypt, 2018. [Google Scholar]
- Michael, P.J.; Steadman, K.J.; Plummer, J.A. The Biology of Australian Weeds 52. Malva parviflora L. Plant Prot. Q. 2009, 24, 2. [Google Scholar]
- Boulos, L. Flora of Egypt: Geraniaceae-Boraginaceae; Al Hadara Publishing: Cairo, Egypt, 2000; Volume 2, p. 352. [Google Scholar]
- Boulos, L. Flora of Egypt Checklist; Al-Hadara Publishing: Cairo, Egypt, 2009; p. 410. [Google Scholar]
- Abbas, A.M.; Mancilla-Leytón, J.M.; Castillo, J.M. Can camels disperse seeds of the invasive tree Prosopis juliflora? Weed Res. 2018, 58, 221–228. [Google Scholar] [CrossRef]
- Manoto, M.M.; Ferreira, M.; Agenbag, G. The effect of temperature on the germination of six selected weed species. S. Afr. J. Plant Soil 2004, 21, 214–219. [Google Scholar] [CrossRef]
- Michael, P.J.; Steadman, K.J.; Plummer, J.A.; Vercoe, P. Sheep rumen digestion and transmission of weedy Malva parviflora seeds. Aust. J. Exp. Agric. 2006, 46, 1251–1256. [Google Scholar] [CrossRef]
- Abbas, A.M.; Rubio-Casal, A.E.; De Cires, A.; Figueroa, M.E.; Lambert, A.M.; Castillo, J.M. Effects of flooding on germination and establishment of the invasive cordgrass Spartina densiflora. Weed Res. 2012, 52, 269–276. [Google Scholar] [CrossRef]
- Mackay, D.B. The measurement of viability. In Viability of Seeds; Roberts, E.H., Ed.; Chapman and Hall: London, UK, 1972; pp. 172–208. [Google Scholar]
- Jaganathan, G.K.; Yule, K.; Liu, B. On the evolutionary and ecological value of breaking physical dormancy by endozoochory. Perspect. Plant Ecol. Evol. Syst. 2016, 22, 11–22. [Google Scholar] [CrossRef]
- Marchetto, K.M.; Heuschele, D.J.; Larkin, D.J.; Wolf, T.M. Goat digestion leads to low survival and viability of common buckthorn (Rhamnus cathartica) seeds. Nat. Areas J. 2020, 40, 150–154. [Google Scholar] [CrossRef]
- Frost, R.A.; Mosley, J.C.; Roeder, B.L. Recovery and viability of sulfur cinquefoil seeds from the feces of sheep and goats. Rangel. Ecol. Manag. 2013, 66, 51–55. [Google Scholar] [CrossRef]
- Traveset, A. Effect of seed passage through vertebrate frugivores’ guts on germination: A review. Perspect. Plant Ecol. Evol. Syst. 1998, 1, 151–190. [Google Scholar] [CrossRef] [Green Version]
- Blackshaw, R.E.; Rode, L.M. Effect of ensiling and rumen digestion by cattle on weed seed viability. Weed Sci. 1991, 39, 104–108. [Google Scholar] [CrossRef]
- Krenchinski, F.H.; Albrecht, A.J.P.; Albrecht, L.P.; Villetti, H.L.; Orso, G.; Barroso, A.A.M.; Victoria Filho, R. Germinação e dormência em sementes de Sorghum halepense e Sorghum arundinaceum. Planta Daninha 2015, 33, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Kneuper, C.L.; Scott, C.B.; Pinchak, W.E. Consumption and dispersion of Mesquite seeds by ruminants. J Range Manag. 2003, 56, 255–259. [Google Scholar] [CrossRef]
- Piltz, J.W.; Stanton, R.A.; Wu, H. Effect of ensiling and in sacco digestion on the viability of seeds of selected weed species. Weed Res. 2017, 57, 382–389. [Google Scholar] [CrossRef]
- Scarabel, L.; Panozzo, S.; Savoia, W.; Sattin, M. Target-site ACCase-Resistant Johnsongrass (Sorghum halepense) selected in summer dicot crops. Weed Technol. 2014, 28, 307–315. [Google Scholar] [CrossRef]
- Travlos, I.S.; Montull, J.M.; Kukorelli, G.; Malidza, G.; Dogan, M.N.; Cheimona, N.; Antonopoulos, N.; Kanatas, P.J.; Zannopoulos, S.; Peteinatos, G. Key aspects on the biology, ecology and impacts of Johnsongrass [Sorghum halepense (L.) Pers] and the role of glyphosate and non-chemical alternative practices for the management of this weed in Europe. Agronomy 2019, 9, 717. [Google Scholar] [CrossRef] [Green Version]
- Nóbrega, J.E.; Riet-Correa, F.; Mederisos, R.M.T.; Dantas, A.F.M. Poisoning by Sorghum halepense (Poaceae) in cattle in the Brazilian semiarid. Pesqui. Vet. Bras. 2006, 26, 201–204. [Google Scholar]
- Auffret, A.G.; Cousins, S.A.O. Grassland connectivity by motor vehicles and grazing livestock. Ecography 2013, 36, 1150–1157. [Google Scholar] [CrossRef]
- Dovrat, G.; Perevolotsky, A.; Ne’eman, G. Wild boars as seed dispersal agents of exotic plants from agricultural lands to conservation areas. J. Arid Environ. 2012, 78, 49–54. [Google Scholar] [CrossRef]
- Farmer, J.A.; Webb, E.B.; Pierce, R.A.; Bradley, K.W. Evaluating the potential for weed seed dispersal based on waterfowl consumption and seed viability. Pest Manag. Sci. 2017, 73, 2592–2603. [Google Scholar] [CrossRef] [PubMed]
- Benvenuti, S. Weed seed movement and dispersal strategies in the agricultural environment. Weed Biol. Manag. 2007, 7, 141–157. [Google Scholar] [CrossRef]
- Ginman, E.; Prider, J.; Matthews, J.; Virtue, J.; Watling, J. Sheep as vectors for branched broomrape (Orobanche ramosa subsp mutelii [FW Schultz] Cout.) seed dispersal. Weed Biol. Manag. 2015, 15, 61–69. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Malva parviflora | Sorghum halepense | |||||||
|---|---|---|---|---|---|---|---|---|
| Length | Width | Height | Mass | Length | Width | Height | Mass | |
| Uneaten seeds | 1.56 ± 0.02 a | 1.72 ± 0.02 a | 1.51 ± 0.03 a | 6 ± 0 a | 4.25 ± 0.08 a | 2.23 ± 0.04 a | 1.43 ± 0.03 a | 16 ± 2 a |
| Retrieved seeds | 1.74 ± 0.05 a | 1.74 ± 0.05 a | 1.28 ± 0.06 b | 13 ± 3 b | 4.64 ± 0.14 b | 2.29 ± 0.04 a | 1.44 ± 0.03 a | 13 ± 0 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbas, A.M.; Al-Kahtani, M.; Abdelazeem Mousa, M.; Badry, M.O.; Hassaneen, A.S.A.; Ezzat-Ahmed, A.; Mancilla-Leytón, J.M.; Castillo, J.M. Endozoochory by Goats of Two Invasive Weeds with Contrasted Propagule Traits. Sustainability 2020, 12, 5450. https://doi.org/10.3390/su12135450
Abbas AM, Al-Kahtani M, Abdelazeem Mousa M, Badry MO, Hassaneen ASA, Ezzat-Ahmed A, Mancilla-Leytón JM, Castillo JM. Endozoochory by Goats of Two Invasive Weeds with Contrasted Propagule Traits. Sustainability. 2020; 12(13):5450. https://doi.org/10.3390/su12135450
Chicago/Turabian StyleAbbas, Ahmed M., Mohammed Al-Kahtani, Mohamed Abdelazeem Mousa, Mohammed O. Badry, Ahmed S.A. Hassaneen, Ahmed Ezzat-Ahmed, Juan Manuel Mancilla-Leytón, and Jesús M. Castillo. 2020. "Endozoochory by Goats of Two Invasive Weeds with Contrasted Propagule Traits" Sustainability 12, no. 13: 5450. https://doi.org/10.3390/su12135450