Machine Learning Algorithms to Predict Tree-Related Microhabitats using Airborne Laser Scanning

 ,
,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
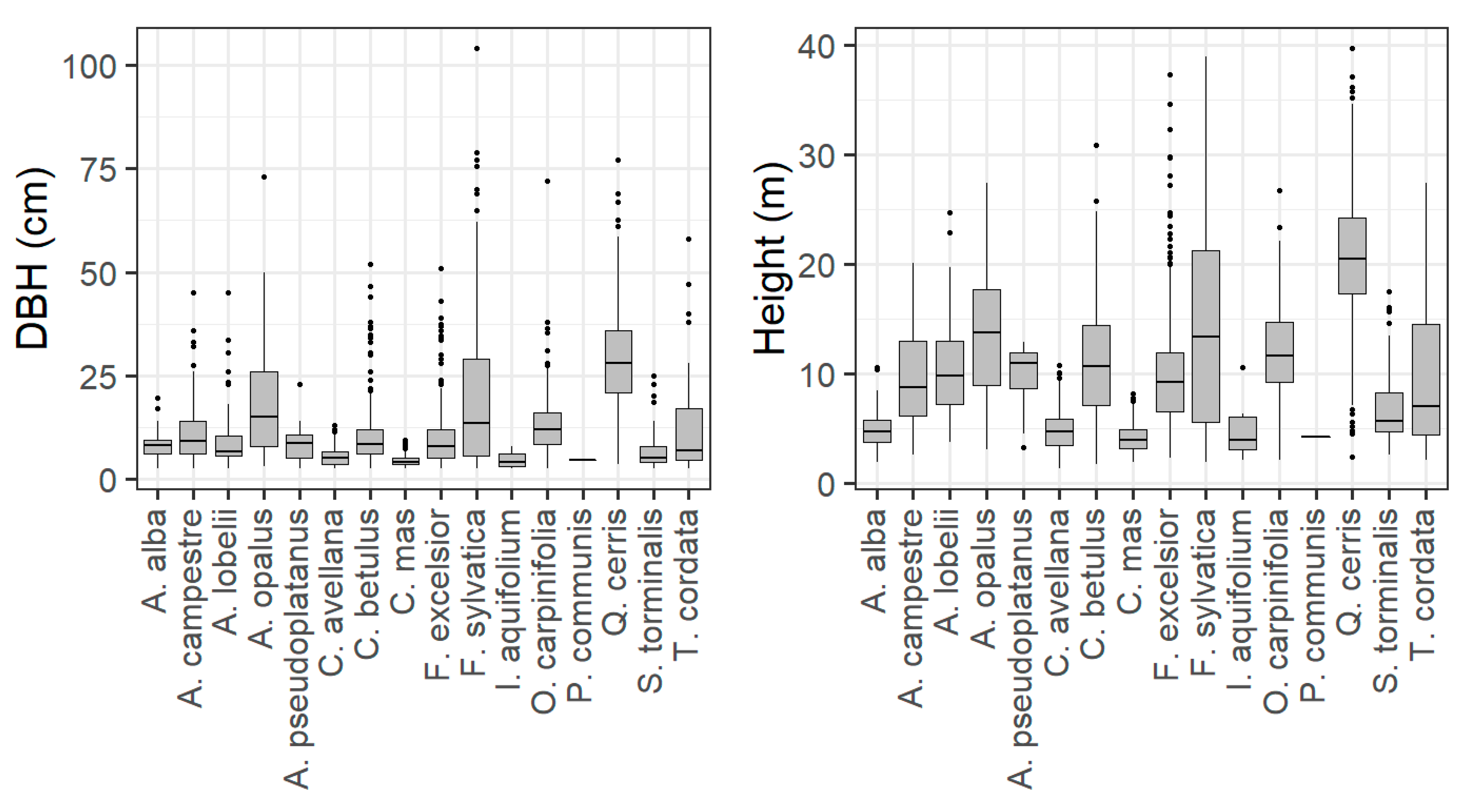
2.2. Field Data
2.3. ALS Data Acquisition and Processing
2.4. Machine Learning Algorithm Implementation
3. Results
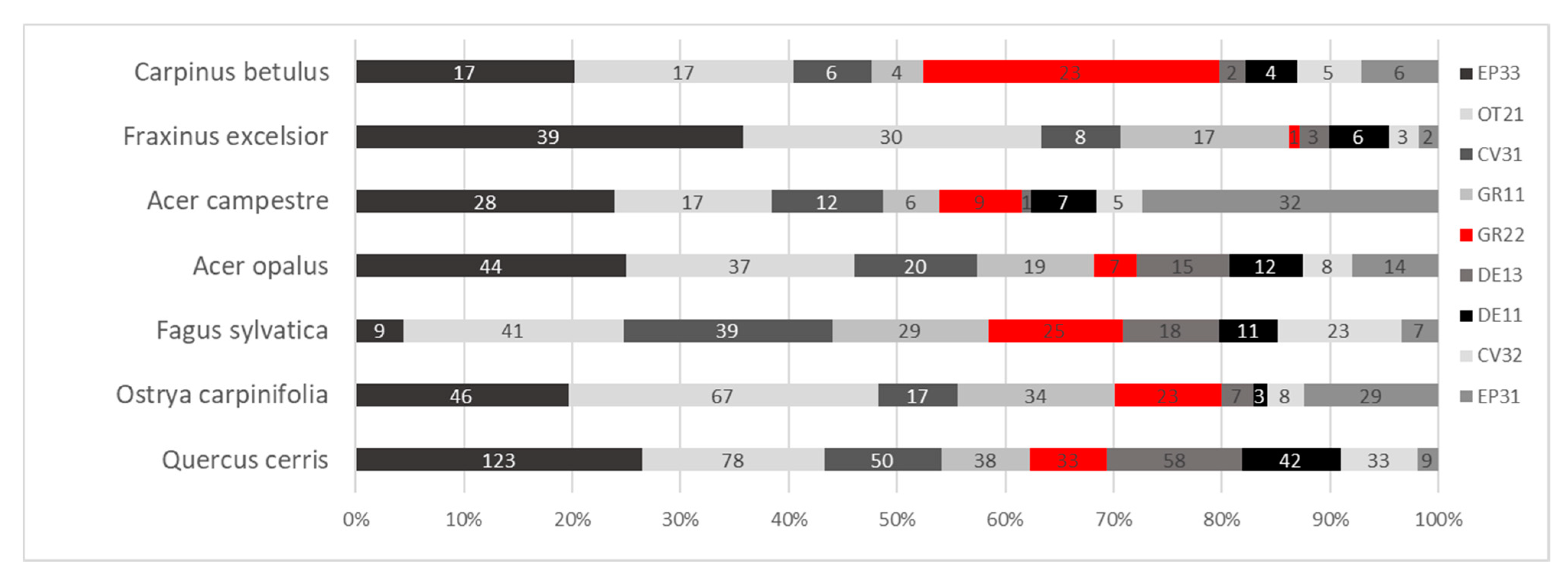
3.1. TreMs Occurrence and Abundance
3.2. Predicting TreMs and Habitat Trees Using ALS Metrics
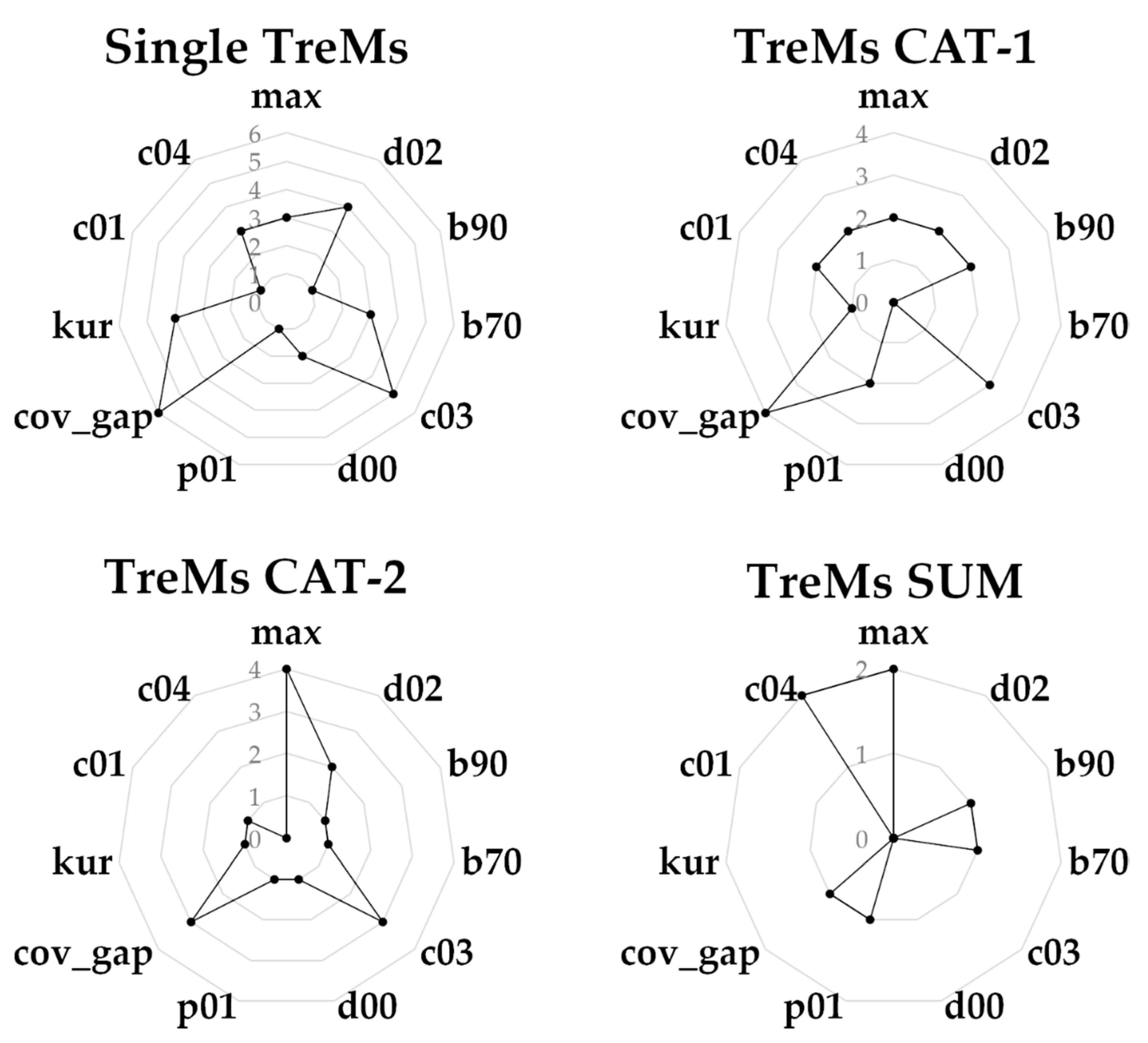
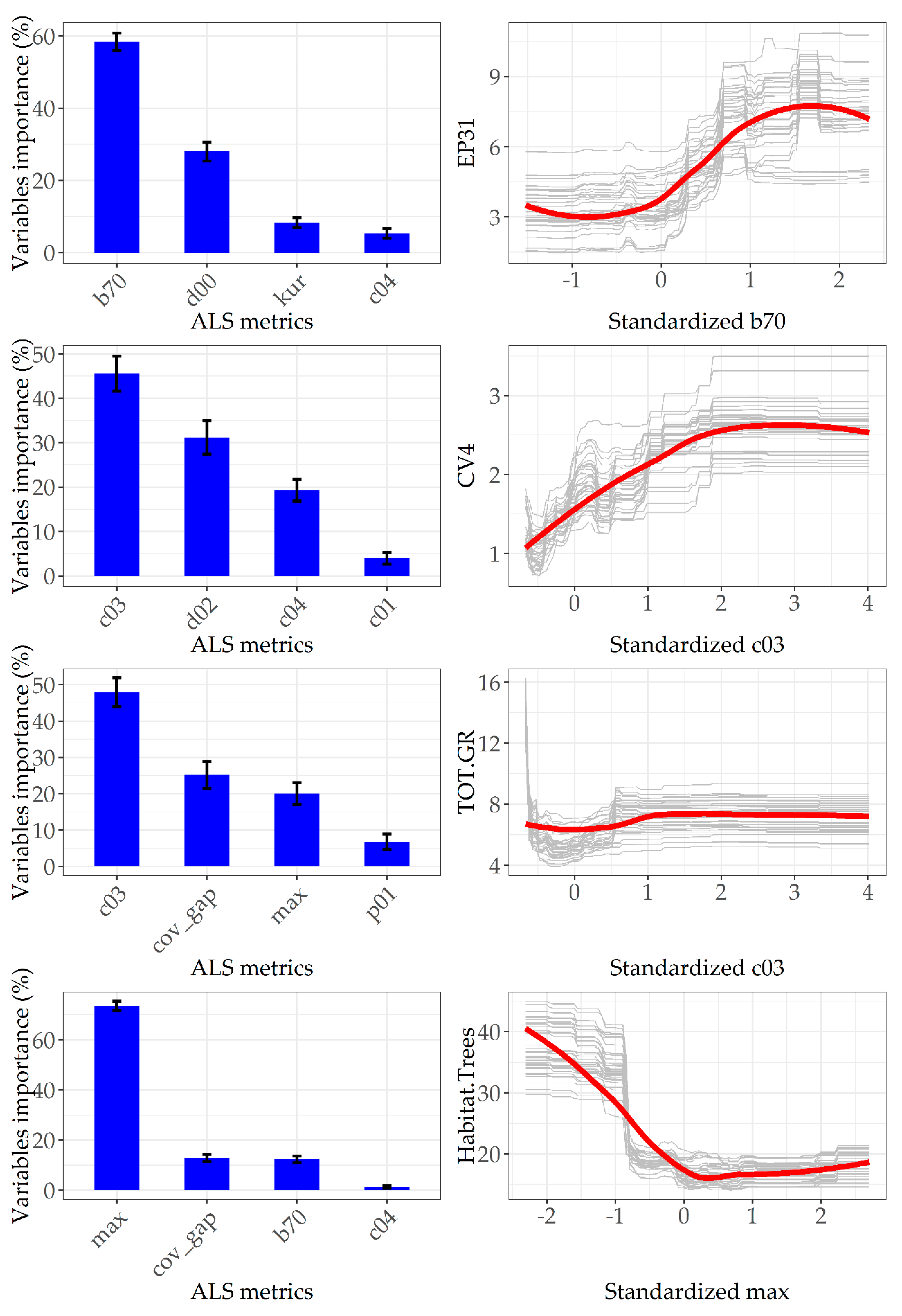
3.3. Ranking Metrics Contribution in Predicting Microhabitat Abundance
4. Discussion
4.1. TreMs Prediction and Forest Structure
4.2. TreMs Hierarchical Prediction
4.3. TreMs as Biodiversity Indicators
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix A
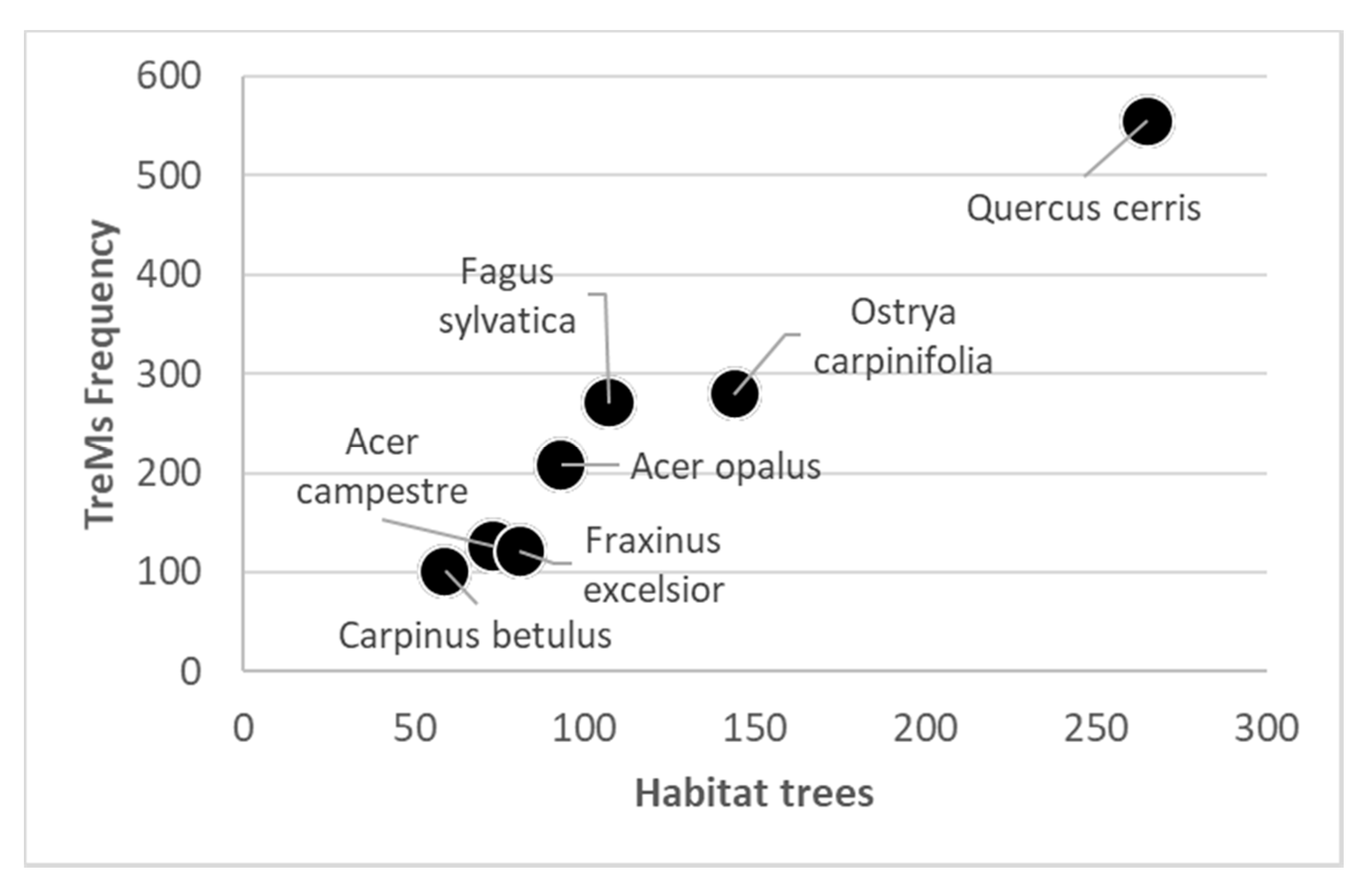
Forest Structure and TreMs Frequency According to Field Data


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volume (m3/ha) | Basal Area (m2/ha) | Tree Species (no.) | Trees ha−1 (no.) | DBH (cm) | Height Max (m) | Habitat Trees (no.) | TreMs (no.) | |
|---|---|---|---|---|---|---|---|---|
| Min | 183 | 19.94 | 3 | 454 | 11.33 | 17.55 | 8 | 14 |
| 1st Qu. | 302 | 301.6 | 7 | 690 | 17.74 | 24.92 | 16 | 31 |
| Median | 367 | 367.4 | 7 | 1020 | 21.25 | 28.44 | 20 | 44 |
| Mean | 387 | 387.4 | 7.5 | 1285 | 21.21 | 28.3 | 24 | 49 |
| 3rd Qu. | 464 | 463.7 | 9 | 1673 | 23.7 | 30.88 | 31 | 65 |
| Max | 647 | 647.2 | 11 | 3705 | 31.96 | 39.63 | 51 | 101 |


Appendix B
| Metrics Name | Metrics Description |
|---|---|
| max* | Maximum height of all returns above the cutoff |
| min | Minimum height of all returns above the cutoff |
| avg | Average height of all returns above the cutoff |
| Std | Standard deviation of height of all returns above the cutoff |
| Ske | Returns height skewness of points above the height cutoff |
| kur* | Returns height kurtosis of points above the height cutoff |
| Qav | Average square height of points above the height cutoff |
| dns.gap | Inverse of the canopy density. Canopy density is the number of points above the height cutoff divided by the number of all returns |
| d00* | Relative height density as the total number of returns between 1.30 and 5 m divided by the total number of points and scaled to a percentage |
| d01 | Relative height density as the total number of returns between 5 and 10 m divided by the total number of points and scaled to a percentage |
| d02* | Relative height density as the total number of returns between 10 and 20 m divided by the total number of points and scaled to a percentage |
| d03 | Relative height density as the total number of returns between 20 and 30 m divided by the total number of points and scaled to a percentage |
| d04 | Relative height density as the total number of returns > 30 m divided by the total number of points and scaled to a percentage |
| cov.gap* | Inverse in the canopy cover. Canopy cover is the number of first returns above the height cutoff divided by the number of all first returns and output as a percentage |
| c00 | Number of returns between 1.30 and 5 m |
| c01* | Number of returns between 5 and 10 m |
| c02 | Number of returns between 10 and 20 m |
| c03* | Number of returns between 20 and 30 m |
| c04* | Number of returns > 30 m |
| p01* | Percentile 1 of height distribution |
| p05 | Percentile 5 of height distribution |
| p10 | Percentile 10 of height distribution |
| p25 | Percentile 25 of height distribution |
| p50 | Percentile 50 of height distribution |
| p75 | Percentile 75 of height distribution |
| p90 | Percentile 90 of height distribution |
| P95 | Percentile 95 of height distribution |
| p99 | Percentile 99 of height distribution |
| b10 | The percentage of ALS returns whose heights are below 10% of the maximum tree height, after the subtraction of the height cut-off value |
| b20 | The percentage of ALS returns whose heights are below 20% of the maximum tree height, after the subtraction of the height cut-off value |
| b30 | The percentage of ALS returns whose heights are below 30% of the maximum tree height, after the subtraction of the height cut-off value |
| b40 | The percentage of ALS returns whose heights are below 40% of the maximum tree height, after the subtraction of the height cut-off value |
| b50 | The percentage of ALS returns whose heights are below 50% of the maximum tree height, after the subtraction of the height cut-off value |
| b60 | The percentage of ALS returns whose heights are below 60% of the maximum tree height, after the subtraction of the height cut-off value |
| b70* | The percentage of ALS returns whose heights are below 70% of the maximum tree height, after the subtraction of the height cut-off value |
| b80 | The percentage of ALS returns whose heights are below 80% of the maximum tree height, after the subtraction of the height cut-off value |
| b90* | The percentage of ALS returns whose heights are below 90% of the maximum tree height, after the subtraction of the height cut-off value |
References
- Bütler, R.; Lachat, T.; Larrieu, L.; Paillet, Y. Habitat trees: Key elements for forest biodiversity. In Integrative Approaches as an Opportunity for the Conservation of Forest Biodiversity; Kraus, D., Krumm, F., Eds.; European Forest Insititute: Joensuu, Finland, 2013; pp. 84–91. [Google Scholar]
- Großmann, J.; Schultze, J.; Bauhus, J.; Pyttel, P. Predictors of Microhabitat Frequency and Diversity in Mixed Mountain Forests in South-Western Germany. Forests 2018, 9, 104. [Google Scholar] [CrossRef] [Green Version]
- Leidinger, J.; Weisser, W.W.; Kienlein, S.; Blaschke, M.; Jung, K.; Kozak, J.; Fischer, A.; Mosandl, R.; Michler, B.; Ehrhardt, M.; et al. Formerly managed forest reserves complement integrative management for biodiversity conservation in temperate European forests. Biol. Conserv. 2020, 242, 108437. [Google Scholar] [CrossRef]
- Asbeck, T.; Pyttel, P.; Frey, J.; Bauhus, J. Predicting abundance and diversity of tree-related microhabitats in Central European montane forests from common forest attributes. For. Ecol. Manag. 2019, 432, 400–408. [Google Scholar] [CrossRef]
- Santopuoli, G.; di Cristofaro, M.; Kraus, D.; Schuck, A.; Lasserre, B.; Marchetti, M. Biodiversity conservation and wood production in a Natura 2000 Mediterranean forest. A trade-off evaluation focused on the occurrence of microhabitats. iForest-Biogeosciences For. 2019, 12, 76–84. [Google Scholar] [CrossRef]
- Paillet, Y.; Debaive, N.; Archaux, F.; Cateau, E.; Gilg, O.; Guilbert, E. Nothing else matters? Tree diameter and living status have more effects than biogeoclimatic context on microhabitat number and occurrence: An analysis in French forest reserves. PLoS ONE 2019, 14, e0216500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, A.K.; Winter, S. Tree microhabitat structures as indicators of biodiversity in Douglas-Fir forests of different stand ages and management histories in the Pacific Northwest, U.S.A. For. Ecol. Manag. 2009, 257, 1453–1464. [Google Scholar] [CrossRef]
- Kraus, D.; Schuck, A.; Bebi, P.; Blaschke, M.; Bütler, R.; Flade, M.; Heintz, W.; Krumm, F.; Lachat, T.; Larrieu, L.; et al. Spatially Explicit Database of Tree Related Microhabitats (TreMs). Version 1.2. Integrate+ Project; Institut National de la Recherche Agronomique (INRA): Paris, France, 2017. [Google Scholar]
- Larrieu, L.; Paillet, Y.; Winter, S.; Bütler, R.; Kraus, D.; Krumm, F.; Lachat, T.; Michel, A.K.; Regnery, B.; Vandekerkhove, K. Tree related microhabitats in temperate and Mediterranean European forests: A hierarchical typology for inventory standardization. Ecol. Indic. 2018, 84, 194–207. [Google Scholar] [CrossRef]
- Larrieu, L.; Cabanettes, A. Species, live status, and diameter are important tree features for diversity and abundance of tree microhabitats in subnatural montane beech-fir forests. Can. J. For. Res. 2012, 42, 1433–1445. [Google Scholar] [CrossRef]
- Vuidot, A.; Paillet, Y.; Archaux, F.; Gosselin, F. Influence of tree characteristics and forest management on tree microhabitats. Biol. Conserv. 2011, 144, 441–450. [Google Scholar] [CrossRef]
- Paillet, Y.; Bergès, L.; Hjältén, J.; Odor, P.; Avon, C.; Bernhardt-Römermann, M.; Bijlsma, R.-J.; De Bruyn, L.; Fuhr, M.; Grandin, U.; et al. Biodiversity differences between managed and unmanaged forests: Meta-analysis of species richness in Europe. Conserv. Biol. 2010, 24, 101–112. [Google Scholar] [CrossRef]
- Johann, F.; Schaich, H. Land ownership affects diversity and abundance of tree microhabitats in deciduous temperate forests. For. Ecol. Manag. 2016, 380, 70–81. [Google Scholar] [CrossRef]
- Regnery, B.; Paillet, Y.; Couvet, D.; Kerbiriou, C. Which factors influence the occurrence and density of tree microhabitats in Mediterranean oak forests? For. Ecol. Manag. 2013, 295, 118–125. [Google Scholar] [CrossRef]
- Cosyns, H.; Kraus, D.; Krumm, F.; Schulz, T.; Pyttel, P. Reconciling the Tradeoff between Economic and Ecological Objectives in Habitat-Tree Selection: A Comparison between Students, Foresters, and Forestry Trainers. For. Sci. 2019, 65, 223–234. [Google Scholar] [CrossRef]
- McRoberts, R.E.; Winter, S.; Chirici, G.; LaPoint, E. Assessing forest naturalness. For. Sci. 2012, 58, 294–309. [Google Scholar] [CrossRef]
- Antonucci, S.; Rossi, S.; Deslauriers, A.; Morin, H.; Lombardi, F.; Marchetti, M.; Tognetti, R. Large-scale estimation of xylem phenology in black spruce through remote sensing. Agric. For. Meteorol. 2017, 233, 92–100. [Google Scholar] [CrossRef]
- Congedo, L.; Sallustio, L.; Munafò, M.; Ottaviano, M.; Tonti, D.; Marchetti, M. Copernicus high-resolution layers for land cover classification in Italy. J. Maps 2016, 12, 1195–1205. [Google Scholar] [CrossRef] [Green Version]
- Montaghi, A.; Corona, P.; Dalponte, M.; Gianelle, D.; Chirici, G.; Olsson, H. Airborne laser scanning of forest resources: An overview of research in Italy as a commentary case study. Int. J. Appl. Earth Obs. Geoinf. 2013, 23, 288–300. [Google Scholar] [CrossRef] [Green Version]
- Mura, M.; McRoberts, R.E.; Chirici, G.; Marchetti, M. Estimating and mapping forest structural diversity using airborne laser scanning data. Remote Sens. Environ. 2015, 170, 133–142. [Google Scholar] [CrossRef]
- Santi, E.; Paloscia, S.; Pettinato, S.; Fontanelli, G.; Mura, M.; Zolli, C.; Maselli, F.; Chiesi, M.; Bottai, L.; Chirici, G. The potential of multifrequency SAR images for estimating forest biomass in Mediterranean areas. Remote Sens. Environ. 2017, 200, 63–73. [Google Scholar] [CrossRef]
- Chirici, G.; McRoberts, R.E.; Fattorini, L.; Mura, M.; Marchetti, M. Comparing echo-based and canopy height model-based metrics for enhancing estimation of forest aboveground biomass in a model-assisted framework. Remote Sens. Environ. 2016, 174, 1–9. [Google Scholar] [CrossRef]
- Giannetti, F.; Puletti, N.; Puliti, S.; Travaglini, D.; Chirici, G. Assessment of UAV photogrammetric DTM-independent variables for modelling and mapping forest structural indices in mixed temperate forests. Ecol. Indic. 2020, 117. [Google Scholar] [CrossRef]
- Giannetti, F.; Chirici, G.; Gobakken, T.; Næsset, E.; Travaglini, D.; Puliti, S. A new approach with DTM-independent metrics for forest growing stock prediction using UAV photogrammetric data. Remote Sens. Environ. 2018, 213, 195–205. [Google Scholar] [CrossRef]
- Marcelli, A.; Mattioli, W.; Puletti, N.; Chianucci, F.; Gianelle, D.; Grotti, M.; Chirici, G.; D’Amico, G.; Francini, S.; Travaglini, D.; et al. Large-Scale two-phase estimation of wood production by poplar plantations exploiting sentinel-2 data as auxiliary information. Silva Fenn. 2020, 54, 15. [Google Scholar] [CrossRef] [Green Version]
- Maselli, F.; Chiesi, M.; Mura, M.; Marchetti, M.; Corona, P.; Chirici, G. Combination of optical and LiDAR satellite imagery with forest inventory data to improve wall-to-wall assessment of growing stock in Italy. Int. J. Appl. Earth Obs. Geoinf. 2014, 26, 377–386. [Google Scholar] [CrossRef]
- Lasserre, B.; Chirici, G.; Chiavetta, U.; Garfì, V.; Tognetti, R.; Drigo, R.; DiMartino, P.; Marchetti, M. Assessment of potential bioenergy from coppice forests trough the integration of remote sensing and field surveys. Biomass Bioenergy 2011, 35, 716–724. [Google Scholar] [CrossRef]
- Giannetti, F.; Puletti, N.; Quatrini, V.; Travaglini, D.; Bottalico, F.; Corona, P.; Chirici, G. Integrating terrestrial and airborne laser scanning for the assessment of single-tree attributes in Mediterranean forest stands. Eur. J. Remote Sens. 2018, 51, 795–807. [Google Scholar] [CrossRef] [Green Version]
- Rehush, N.; Abegg, M.; Waser, L.T.; Brändli, U.-B. Identifying Tree-Related Microhabitats in TLS Point Clouds Using Machine Learning. Remote Sens. 2018, 10, 1735. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Popescu, S.C.; Lawing, A.M.; Eriksson, M.; Strimbu, B.M.; Bürkner, P.C. Bayesian and classical machine learning methods: A Comparison for tree species classification with LiDAR waveform signatures. Remote Sens. 2018, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Barabesi, L.; Franceschi, S.; Marcheselli, M. Properties of design-based estimation under stratified spatial sampling with application to canopy coverage estimation. Ann. Appl. Stat. 2012, 6, 210–228. [Google Scholar] [CrossRef] [Green Version]
- Frate, L.; Carranza, M.L.; Garfì, V.; Di Febbraro, M.; Tonti, D.; Marchetti, M.; Ottaviano, M.; Santopuoli, G.; Chirici, G. Spatially explicit estimation of forest age by integrating remotely sensed data and inverse yield modeling techniques. IForest 2016, 9, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Kraus, D.; Bütler, R.; Krumm, F.; Lachat, T.; Larrieu, L.; Mergner, U.; Paillet, Y.; Rydkvist, T.; Schuck, A.; Winter, S. Catalogue of Tree Microhabitats—Reference Field List; Integrate+TechnicalPaper.16p; European Forest Institute: Freiburg, Germany, 2016. [Google Scholar]
- Hengl, T.; Walsh, M.G.; Sanderman, J.; Wheeler, I.; Harrison, S.P.; Prentice, I.C. Global mapping of potential natural vegetation: An assessment of machine learning algorithms for estimating land potential. PeerJ 2018, 6, e5457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Tompalski, P.; White, J.C.; Coops, N.C.; Wulder, M.A. Demonstrating the transferability of forest inventory attribute models derived using airborne laser scanning data. Remote Sens. Environ. 2019, 227, 110–124. [Google Scholar] [CrossRef]
- Kuhn, M. Building predictive models in R using the caret package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Vabalas, A.; Gowen, E.; Poliakoff, E.; Casson, A.J. Machine learning algorithm validation with a limited sample size. PLoS ONE 2019, 14, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Harrell, F.E., Jr.; Lee, K.L.; Mark, D.B. Multivariable prognostic models: Issues in developing models, evaluating assumptions and adequacy, and measuring and reducing errors. Stat. Med. 1996, 15, 361–387. [Google Scholar] [CrossRef]
- Breiner, F.T.; Guisan, A.; Bergamini, A.; Nobis, M.P. Overcoming limitations of modelling rare species by using ensembles of small models. Methods Ecol. Evol. 2015, 6, 1210–1218. [Google Scholar] [CrossRef]
- Di Febbraro, M.; D’Amen, M.; Raia, P.; De Rosa, D.; Loy, A.; Guisan, A. Using macroecological constraints on spatial biodiversity predictions under climate change: The modelling method matters. Ecol. Model. 2018, 390, 79–87. [Google Scholar] [CrossRef]
- Botchkarev, A. A new typology design of performance metrics to measure errors in machine learning regression algorithms. Interdiscip. J. Inf. Knowl. Manag. 2019, 14, 45–76. [Google Scholar] [CrossRef] [Green Version]
- Parisi, F.; Di Febbraro, M.; Lombardi, F.; Biscaccianti, A.B.; Campanaro, A.; Tognetti, R.; Marchetti, M. Relationships between stand structural attributes and saproxylic beetle abundance in a Mediterranean broadleaved mixed forest. For. Ecol. Manag. 2019, 432, 957–966. [Google Scholar] [CrossRef]
- Sačkov, I.; Santopuoli, G.; Bucha, T.; Lasserre, B.; Marchetti, M. Forest inventory attribute prediction using lightweight aerial scanner data in a selected type of multilayered deciduous forest. Forests 2016, 7, 307. [Google Scholar] [CrossRef] [Green Version]
- Király, I.; Nascimbene, J.; Tinya, F.; Ódor, P. Factors influencing epiphytic bryophyte and lichen species richness at different spatial scales in managed temperate forests. Biodivers. Conserv. 2013, 22, 209–223. [Google Scholar] [CrossRef] [Green Version]
- Maesano, M.; Lasserre, B.; Masiero, M.; Tonti, D.; Marchetti, M. First mapping of the main high conservation value forests (HCVFs) at national scale: The case of Italy. Plant Biosyst. 2016, 150, 208–216. [Google Scholar] [CrossRef]
- Santopuoli, G.; Ferranti, F.; Marchetti, M. Implementing Criteria and Indicators for Sustainable Forest Management in a Decentralized Setting: Italy as a Case Study. J. Environ. Policy Plan. 2016, 18, 177–196. [Google Scholar] [CrossRef]
- Wolfslehner, B.; Baycheva-Merger, T. Evaluating the implementation of the Pan-European Criteria and indicators for sustainable forest management—A SWOT analysis. Ecol. Indic. 2016, 60, 1192–1199. [Google Scholar] [CrossRef]
- Winter, S.; Möller, G.C. Microhabitats in lowland beech forests as monitoring tool for nature conservation. For. Ecol. Manag. 2008, 255, 1251–1261. [Google Scholar] [CrossRef]
- Lombardi, F.; Lasserre, B.; Chirici, G.; Tognetti, R.; Marchetti, M. Deadwood occurrence and forest structure as indicators of old-growth forest conditions in Mediterranean mountainous ecosystems. ECOSCIENCE 2012, 19, 344–355. [Google Scholar] [CrossRef]
- Pioli, S.; Antonucci, S.; Giovannelli, A.; Traversi, M.L.; Borruso, L.; Bani, A.; Brusetti, L.; Tognetti, R. Community fingerprinting reveals increasing wood-inhabiting fungal diversity in unmanaged Mediterranean forests. For. Ecol. Manag. 2018, 408, 202–210. [Google Scholar] [CrossRef]
- Vizzarri, M.; Chiavetta, U.; Santopuoli, G.; Tonti, D.; Marchetti, M. Mapping forest ecosystem functions for landscape planning in a mountain Natura2000 site, Central Italy. J. Environ. Plan. Manag. 2014, 0568, 1–25. [Google Scholar] [CrossRef]
- Pastorella, F.; Giacovelli, G.; Maesano, M.; Paletto, A.; Vivona, S.; Veltri, A.; Pellicone, G.; Mugnozza, G.S. Social perception of forest multifunctionality in southern Italy: The case of Calabria Region. J. For. Sci. 2016, 62, 366–379. [Google Scholar] [CrossRef] [Green Version]
- Gasparini, P.; Tabacchi, G. L’Inventario Nazionale delle Foreste e dei serbatoi forestali di Carbonio INFC 2005. Secondo inventario forestale nazionale italiano. In Metodi e Risultati; Edagricole-Il Sole 24 ore: Bologna, Italy, 2011; p. 653. [Google Scholar]
- Santopuoli, G.; Requardt, A.; Marchetti, M. Application of indicators network analysis to support local forest management plan development: A case study in Molise, Italy. iForest-Biogeosci. For. 2012, 5, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Maesano, M.; Ottaviano, M.; Lidestav, G.; Lasserre, B.; Matteucci, G.; Mugnozza, G.S.; Marchetti, M. Forest certification map of Europe. IForest 2018, 11, 526–533. [Google Scholar] [CrossRef]
- Santopuoli, G.; Lasserre, B.; Di Martino, P.; Marchetti, M. Dynamics of the silver fir (Abies alba Mill.) natural regeneration in a mixed forest in the Central Apennine. Plant Biosyst. 2016, 150, 217–226. [Google Scholar] [CrossRef]




| Single TreMs | TreMs CAT-1 | TreMs CAT-2 | Single TreMs | TreMs CAT-1 | TreMs CAT-2 |
|---|---|---|---|---|---|
| CV11* | Woodpecker cavities | Cavities (CV) | BA11 | Bark pockets | Bark (BA) |
| CV12 | BA12 | ||||
| CV13* | BA21* | Bark structure | |||
| CV14 | DE11 | Dead branches and limbs/crown deadwood | Deadwood (DE) | ||
| CV15* | DE12 | ||||
| CV21 | Trunk and mold cavities | DE13 | |||
| CV22* | DE14 | ||||
| CV23 | DE15* | ||||
| CV24 | GR11 | Root buttress cavities | Deformation/growth form (GR) | ||
| CV25 | GR12 | ||||
| CV26* | GR13* | ||||
| CV31 | Branch hole | GR21 | Witches broom | ||
| CV32 | GR22 | ||||
| CV33 | GR31 | Cankers and burrs | |||
| CV41 | Dendrotelms and water-filled holes | GR32 | |||
| CV42 | EP11* | Fruiting bodies fungi | Epiphytes (EP) | ||
| CV43 | EP12* | ||||
| CV44 | EP13* | ||||
| CV51 | Insect galleries and bore holes | EP14* | |||
| CV52* | EP21 | Myxomycetes | |||
| IN11* | Bark loss/exposed sapwood | Injuries and wounds (IN) | EP31 | Epiphytic crypto- andphanerogams | |
| IN12* | EP32 | ||||
| IN13 | EP33 | ||||
| IN14* | EP34* | ||||
| IN21 | Exposed heartwood/trunk and crown breakage | EP35* | |||
| IN22 | NE11* | Vertebrate nests | Nests (NE) | ||
| IN23 | NE12* | ||||
| IN24* | NE21 | Invertebrate nests | |||
| IN31 | Cracks and scar | OT11* | Sap and resin run | Other (OT) | |
| IN32 | OT12* | ||||
| IN33 | OT21 | Microsoil | |||
| IN34* | OT22 |
| TreMs Code | TreMs Type | Frequency of TreMs | Frequency (%) |
|---|---|---|---|
| EP33 | Lianas, coverage > 25 % | 324 | 18.89 |
| OT21 | Crown microsoil | 296 | 17.26 |
| CV31 | Branch hole ø ≥ 5 cm | 153 | 8.92 |
| GR11 | Root buttress cavities ≥ 5 cm | 151 | 8.80 |
| GR22 | Water sprout | 121 | 7.06 |
| DE13 | Dead branches and limbs ø 10–20 cm, not sun-exposed | 107 | 6.24 |
| EP31 | Epiphytic bryophyte coverage > 25 % | 100 | 5.83 |
| CV32 | Branch hole ø ≥ 10 cm | 88 | 5.13 |
| DE11 | Dead branches and limbs ø 10–20 cm, sun-exposed | 88 | 5.13 |
| GR12 | Root buttress cavities ≥ 10 cm | 66 | 3.85 |
| CV44 | Dendrotelms and water-filled holes ø ≥ 15 cm/crown | 38 | 2.22 |
| GR32 | Decayed canker, ø > 20 cm | 30 | 1.75 |
| CV33 | Hollow branch, ø ≥ 10 cm | 28 | 1.63 |
| CV12 | Woodpecker cavities ø = 5–6 cm | 24 | 1.40 |
| DE14 | Dead branches and limbs ø > 20 cm, not sun-exposed | 16 | 0.93 |
| CV43 | Dendrotelms and water-filled holes ø ≥ 5 cm/crown | 13 | 0.76 |
| DE12 | Dead branches and limbs ø > 20 cm, sun exposed | 10 | 0.58 |
| Others | Other TreMs | 62 | 3.6 |
| TreMs | NMAE | SE | |
|---|---|---|---|
| Single TreMs | CV31 | 0.265 | 0.069 |
| CV32 | 0.287 | 0.103 | |
| DE11 | 0.289 | 0.076 | |
| DE13 | 0.249 | 0.067 | |
| EP31 | 0.429 | 0.268 | |
| EP33 | 0.270 | 0.067 | |
| GR11 | 0.272 | 0.098 | |
| GR12 | 0.355 | 0.111 | |
| GR22 | 0.259 | 0.102 | |
| OT21 | 0.267 | 0.097 | |
| TreMs CAT-1 | CV3 | 0.251 | 0.072 |
| CV4 | 0.364 | 0.174 | |
| EP3 | 0.278 | 0.061 | |
| GR1 | 0.269 | 0.082 | |
| GR2 | 0.275 | 0.086 | |
| TreMs CAT-2 | CV | 0.252 | 0.079 |
| DE | 0.282 | 0.048 | |
| EP | 0.263 | 0.051 | |
| GR | 0.283 | 0.099 | |
| OT | 0.269 | 0.076 | |
| TreMs SUM | Habitat Trees | 0.287 | 0.079 |
| Count TreMs | 0.286 | 0.073 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santopuoli, G.; Di Febbraro, M.; Maesano, M.; Balsi, M.; Marchetti, M.; Lasserre, B. Machine Learning Algorithms to Predict Tree-Related Microhabitats using Airborne Laser Scanning. Remote Sens. 2020, 12, 2142. https://doi.org/10.3390/rs12132142
Santopuoli G, Di Febbraro M, Maesano M, Balsi M, Marchetti M, Lasserre B. Machine Learning Algorithms to Predict Tree-Related Microhabitats using Airborne Laser Scanning. Remote Sensing. 2020; 12(13):2142. https://doi.org/10.3390/rs12132142
Chicago/Turabian StyleSantopuoli, Giovanni, Mirko Di Febbraro, Mauro Maesano, Marco Balsi, Marco Marchetti, and Bruno Lasserre. 2020. "Machine Learning Algorithms to Predict Tree-Related Microhabitats using Airborne Laser Scanning" Remote Sensing 12, no. 13: 2142. https://doi.org/10.3390/rs12132142