Waxy Gene-Orthologs in Wheat × Thinopyrum Amphidiploids
by
 , , , and
, , , and
Marina V. Klimushina
1,† ,
,
Pavel Yu. Kroupin
2,3,*,†,
Mikhail S. Bazhenov
2,
Gennady I. Karlov
2 and
Mikhail G. Divashuk
2,3,* 1
National Medical Research Center for Therapy and Preventive Medicine, Ministry of Healthcare of The Russian Federation, Petrovergskiy Lane, 10, 101990 Moscow, Russia
2
Laboratory of Applied Genomics and Crop Breeding, All-Russia Research Institute of Agricultural Biotechnology, Timiryazevskaya Street, 42, 127550 Moscow, Russia
3
Kurchatov Genomics Center of ARRIAB, All-Russia Research Institute of Agricultural Biotechnology, Timiryazevskaya Street, 42, 127550 Moscow, Russia
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Agronomy 2020, 10(7), 963; https://doi.org/10.3390/agronomy10070963
Submission received: 30 April 2020
/
Revised: 20 June 2020
/
Accepted: 2 July 2020
/
Published: 3 July 2020
(This article belongs to the Section Crop Breeding and Genetics)
Abstract
:Starch, as the main component of grain in cereals, serves as the major source of calories in staple food and as a raw material for industry. As the technological and digestive properties of starch depend on its content, the management of its components, amylose and amylopectin, is of great importance. The starch properties of wheat grain can be attuned using allelic variations of genes, including granule-bound starch synthase I (GBSS I), or Wx. The tertiary gene pool, including wheatgrass (Thinopyrum) species, provides a wide spectrum of genes-orthologs that can be used to increase the allelic diversity of wheat genes by wide hybridization. Octaploid partial wheat–wheatgrass hybrids (amphidiploids, WWGHs) combine the complete genome of bread wheat (BBAADD), and a mixed genome from the chromosomes of intermediate wheatgrass (Thinopyrum intermedium, genomic composition JrJrJvsJvsStSt) and tall wheatgrass (Th. ponticum, JJJJJJJsJsJsJs). Thus, WWGHs may carry Wx genes not only of wheat (Wx-B1, Wx-A1 and Wx-D1) but also of wheatgrass origin. We aimed to assess the level of amylose in starch and investigate the polymorphism of Wx genes in 12 accessions of WWGHs. Additionally, we characterized orthologous Wx genes in the genomes of wild wheat-related species involved in the development of the studied WWGHs, Th. intermedium and Th. ponticum, as well as in the putative donors of their subgenomes, bessarabian wheatgrass (Th. bessarabicum, JbJb) and bluebunch wheatgrass (Pseudoroegneria stipifolia, St1St1St2St2). Although no significant differences in amylose content were found between different WWGH accessions, SDS-PAGE demonstrated that at least two WWGHs have an additional band. We sequenced the Wx gene-orthologs in Th. bessarabicum, P. stipifolia, Th. intermedium and Th. ponticum, and developed a WXTH marker that can discriminate the Thinopyrum Wx gene in the wheat background, and localized it to the 7E chromosome in Th. elongatum. Using the WXTH marker we revealed the allelic polymorphism of the Thinopyrum Wx gene in the studied WWGHs. The applicability of Thinopyrum Wx genes in wheat breeding and their effect on starch quality are discussed.
1. Introduction
Wheat is one of the most important crops in the world, and plays a significant role in human nutrition. The major component of wheat grain endosperm is starch, accounting for about two-thirds of the weight of flour, thus starch could significantly affect the products made from wheat. Starch is composed of two types of glucose polymers: branched amylopectin and essentially linear amylose. To characterize the physico-chemical and technological properties of starch, its amylose concentration (%) is often used [1]. Amylose content in the cereal endosperm depends on the enzymes involved in starch synthesis [2]. Granule-bound starch synthase I (GBSSI) or waxy protein is the key enzyme, providing the biosynthesis of amylose; it is encoded by Wx genes [3].
In hexaploid bread wheat, there are three Wx loci, Wx-A1, Wx-B1, and Wx-D1, located on chromosome arms 7AS, 4AL and 7DS, respectively. Studies on the composition of GBSSI proteins with SDS and 2D electrophoresis have demonstrated low polymorphism in hexaploid wheat, in comparison to diploid wheat species [3,4,5,6,7,8,9]. Null mutations at the Wx genes prevent the expression of the functional Wx protein, which results in a decrease in the amylose ratio in partial Wx-mutants (null alleles in one or two Wx loci), and virtually amylose-free starch in complete Wx-mutants (null alleles in three Wx loci). Different combinations of Wx alleles result in different ratios of amylose and amylopectin, thereby causing differences in the chemical and physical properties of starch and, therefore, influencing the quality of the consumed product. Complete Wx-mutants of wheat with the waxy phenotype and decreased (or no) content of amylose are demanded in local markets for various food processing purposes, and for biofuel production. Low-amylose flour is suitable for the production of Japanese udon noodles. Flour blends with waxy flour improve the palatability of noodles, and adding waxy flour keeps baked goods fresh longer, increasing their tolerance to staling and freezing. The starch of waxy wheat is characterized by a faster fermentation rate than starch in the wild-type wheat in fuel ethanol production [10,11].
The contribution of the Wx-B1 gene to amylose content is the most significant, followed by Wx-D1, the contribution of which, in turn, is more than that of Wx-A1 [12]. To date, 19, 15 and 7 allelic variants have been found and described for Wx-A1, Wx-B1 and Wx-D1, respectively [10,13,14,15,16]. Further polymorphism has been detected in Wx genes as SNPs, and insertions and deletions in introns, exons and non-coding regions, which were all associated with both decreases and increases in amylose content [10,13,14,15,16]. For example, Wx-A1g alleles had a 160-bp insertion within the fourth intron that was associated with a lower low level of the Wx-A1 protein [14], while Wx-B1e and Wx-B1f, associated with higher amylose content, have two amino acid substitutions that differentiate them from the wild-type allele Wx-B1a [15,17]. Different allelic variants of Wx genes, with different effects in amylose synthesis, can be either created using mutagenesis [18,19] or found in the natural variability of wheat and its relative species [20,21,22].
To increase the genetic diversity of wheat, genes from primary, as well as secondary and tertiary, pools are used. The efforts in searching for naturally evolved waxy alleles are mainly concentrated on the primary and secondary gene pool, i.e., bread and durum wheats, einkorn, emmer and spelt collections, and wild Aegilops and Triticum germplasm [8,14,17,23,24,25,26,27,28,29,30,31,32].
The tertiary pool of starch synthase gene allele diversity in wheat-related species and their hybrids has been studied insufficiently [33]. The studies of starch in triticale, and starch synthase variability in triticale, are ongoing [34,35,36]. Recently, complete sequences of the Wx gene in Taeniatherum and Hordeum chilense Roem. et Schult. have been published [37,38]. Studies on the Wx gene in the tertiary gene pool, such as Elymus, Thinopyrum, Leymus, Pseudoreogneria and other grasses, are limited and, in general, have partial sequences, and are aimed at phylogenetic studies rather than discovering complete gene sequence [39,40]. It is promising to study Wx genes in wild cereals for several reasons, one of which is the increasing of the diversity of allelic variants in order to later involve them in the selection process. Novel alleles of Wx genes, with altered expression activity of those encoding enzymes, may be used to produce wheat cultivars with different amylose levels.
The first step in using tertiary pool genes is the development of amphidiploids, combining genomes of wheat and wheat-related species; then, translocations are obtained, which are gradually reduced to small fragments of chromatin [41]. Based on the commercial-scale introduction of triticale, amphidiploids themselves can be considered as a prospective crop, particularly due to the wide range of amylose content in the seeds [42]. Additionally, amphidiploids demonstrate complex resistance and tolerance to adverse environmental factors, as well as perenniality, which can be one of the components of sustainable agriculture [43]. The more intensive research is carried out on such prospective amphidiploid crops as tritordeum (Triticum × Hordeum) [44,45,46] and trititrigia (Triticum × Thinopyrum) [47,48,49,50,51,52,53]. Trititrigia cziczinii Tsvel. [wheat–wheatgrass (Thinopyrum) hybrids, hereafter WWGH] was developed to combine the bread-making quality of wheat with the resistance and perennial quality of the Thinopyrum species [43,53]. Further studies revealed new aspects in grain compounds and bread-making quality that can be transferred from wheatgrass, such as higher protein and gluten content, a high content of carotenoids, essential amino acids and dietary fiber, special peptides suitable for celiac patients, and longer amylose chains and internal chains [54,55,56]. Intermediate wheatgrass [Thinopyrum intermedium (Host) Barkworth & D.R. Dewey, 2n = 6x = 42, JrJrJvsJvsStSt] and tall wheatgrass [Th. ponticum (Podp.) Z.-W. Liu & R.-C. Wang, 2n = 10x = 70 JJJJJJJsJsJsJs] have been extensively used in wheat breeding as sources of different resistance genes in wheat genome [57,58,59,60,61]. Although the phylogenetic relationships between these and other Triticeae species are still under discussion, it was demonstrated that the Jr and Jvs subgenomes of Th. Intermedium, and the J and Js subgenomes of Th. ponticum, are close to the genome of bessarabian wheatgrass [Th. bessarabicum (Savul. & Rayss) Á. Löve, Jb, 2n = 14], while the St genome of Th. intermedium is close to the genome of bluebunch wheatgrass [Pseudoroegneria stipifolia (Czern. ex Nevski) A. Löve, St, 2n = 2x = 14; St1St2, 2n = 4x = 28]; the indices r, v and s stand for partial homology of the wheatgrass subgenomes to the R genome of Secale, the V genome of Dasypyrum and the St genome of Pseudoroegneria, respectively [62,63,64,65,66,67,68].
On the other hand, the development of amphidiploids using wide hybridization is a bridge to transferring genes that carry agronomically valuable characteristics to the wheat genome [41,48]. The wide transfer of large chromosomal segments containing genes of interest may be associated with linkage drag, which can reduce wheat grain quality. Thus, it is important to know if the donor genome carries functional genes that can influence (either positively or negatively) the quality of a target wheat grain. Moreover, recently, there has been growing interest in WWGH as a distinct crop, in the form of perennial wheat, in the context of transformative technology [69,70,71].
Wheat–wheatgrass hybrids (WWGH) are partial amphidiploids that have 42 chromosomes of bread wheat and 14 chromosomes of wheatgrass (Thinopyrum species) in their genome [72]. It was shown that, as a rule, wheatgrass chromosomes represent all homeologous groups of the genome. Thus, it is highly likely that the Wx orthologous genes of Thinopyrum (hereafter designated Wx-Th) are present in the WWGH complex genome, which can influence the amylose content in the starch grain of WWGH. The aim of this study was to assess the level of amylose in the starch of different WWHG accessions, to investigate their polymorphism as dependent on the wheatgrass waxy genes, and to characterize the orthologous Wx genes in the genomes of Th. intermedium, Th. ponticum, Th. bessarabicum and Pseudoroegneria stipifolia.
2. Materials and Methods
2.1. Plant Material
The following accessions of wild grasses were used in the study: Th. bessarabicum W6 21890 (2x = 2n = 14, Jb), Pseudoroegneria stipifolia W6 21759 (2x = 2n = 28, St1St1St2St2), Thinopyrum intermedium PI 383573 (2n = 6x = 42, JrJvsSt), Th. ponticum 1158A/19 (2n = 10x = 70, JJJJJJJsJsJsJs). The following accessions of wheat–wheatgrass hybrids (WWGH) were used: 1416, 1432, 150, 1512, 1783, 4044/4, 548, ZP26/1, 166, 1765, 1779 and 1865. Common wheat accessions carrying Wx-B1e, Wx-B1b (null allele) and Wx-B1a (wild-type allele, cv. Nemchinovskaya 56) were used as controls in the amylose content measurement. The set of seven disomic addition lines of the bread wheat Chinese Spring, carrying chromosomes of diploid tall wheatgrass (Th. elongatum, from 1Je to 7Je, developed by Jan Dvorak at UC Davis), was used for the chromosomal localization of the Wx-Th gene.
Accessions W and PI were ordered via Germplasm Resources Information Network (National Genetic Resources Program of USDA). Th. ponticum accession 1158A/19 (kindly provided by Dr. L.I. Glukhova) and WWGH accessions (kindly provided by Dr. V.I. Belov) were developed at the Department of Distant Hybridization, Tsitsin Main Botanical Garden of Russian Academy of Sciences, Moscow, Russia. The set of Th. elongatum addition lines were kindly provided by Dr. Jon Raupp (Wheat Genetic & Genomic Resources Center, Kansas State University, Manhattan KS).
2.2. DNA Isolation, Wx Cloning and Sequencing
DNA extraction from the leaves was performed using the cetyltrimethylammonium bromide (CTAB) method as described in [73]. DNA concentration was determined using the NanoDrop 1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
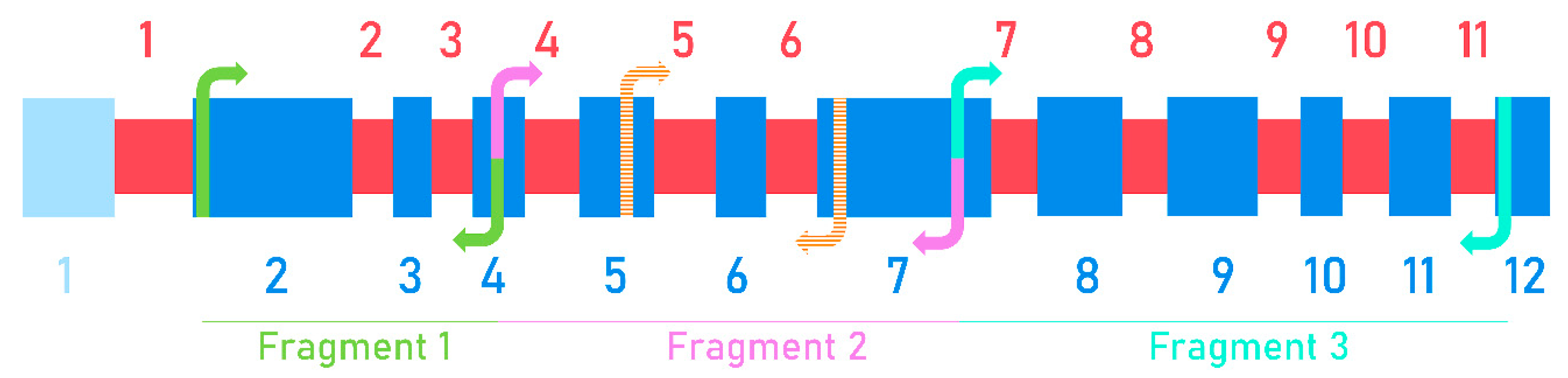
The sequence of the Wx gene was divided into three regions: fragment I (exons 2–4, introns 2–3; exon 1 is in the untranslated region); fragment II (exons 4–7, introns 4–6); and fragment III (exons 7–12, introns 7–11). To amplify these fragments the following primers were used accordingly for each region: WxF3/WxVT1R, WxBAF/WxBAR and WxVT1F/WxVTR (Table 1, Figure 1) [74].
Polymerase chain reaction was carried out using the Tetrad 2 Peltier Thermal Cycler (Bio-Rad, Hercules, CA, USA) in the conditions recommended by the primer designers ([74], Table 1). Amplicons were separated on 2% (m/v) agarose gels in Tris-borate buffer (TBE). The “100 bp Ladder” (Fermentas, Vilnius, Lithuania) was used as a molecular size marker. The PCR products were ligated into pGEM®-T Easy Vectors (Promega, Madison, WI, USA) to generate target plasmids. After transforming plasmids into Escherichia coli strain DH10B cells, three positive clones were isolated for sequencing. Sequencing was performed by a sequencer ABI 3130xl Genetic Analyzer (Applied Biosystems, Foster City, CA, USA) using a set of Big Dye v 3.1 (Thermo Fisher Scientific, Waltham, MA, USA) in accordance with the manufacturer’s recommendations.
2.3. Sequence Data Analysis
DNA sequence assembly reads from Sanger sequencing and their alignment was performed using GeneDoc2.7 software [75]. Spanning of the exons was predicted by aligning the mRNA sequences of the wheat Wx genes (GenBank accessions: EU719608, EU719610, EU719612) to the sequences of the wild grasses. Exon and intron borders for each of the sequenced fragments were predicted using Augustus online tool (http://augustus.gobics.de/) and taking into account the known borders in the bread wheat genes Wx-A1, Wx-B1 and Wx-D1, in the A, B and D subgenomes of wheat, respectively. Translation of the coding DNA sequences into amino acid protein sequences was also performed in GeneDoc2.7. The significance of amino acid substitutions in terms of their impact on protein function was predicted using PROVEAN software tool (J. Craig Venter Institute, Rockville, MD, USA) [76].
2.4. Amylose Content Analysis
The starch granules were extracted from the grains by splitting them using a mortar and pestle and performing the purification procedure described in [77]. Amylose content in the purified starch was determined using the Amylose/Amylopectin Assay Kit (Megazyme, Wicklow, Ireland) and the procedure described by Gibson et al., 1997 [78]. The experiment was performed in double replication, and the differences between variants were estimated using confidence intervals (t-test, p = 0.05).
2.5. Protein Electrophoresis
The extraction of the GBSS1 proteins from the purified starch granules and their identification by the one-dimensional sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) was performed using the method of Zhao and Sharp (1996) [77]. PageRuler™ Protein Ladder #SM0671 (Fermentas, Vilnius, Lithuania) was used as a protein size marker.
3. Results
3.1. Analysis of Amylose Content in Starch in Wheat–Wheatgrass Hybrids
We analyzed the amylose content in the starch of 12 WWGH accessions (Table 2). The bread wheat accessions, cultivated under the same conditions as the WWGH plants and carrying the wild-type alleles of the Wx-A1a and Wx-D1a genes, but differing in the Wx-B1 gene alleles [namely Wx-B1b (null allele), Wx-B1a (wild-type allele) and Wx-B1e (functional allele associated with higher amylose content)], were used as controls.
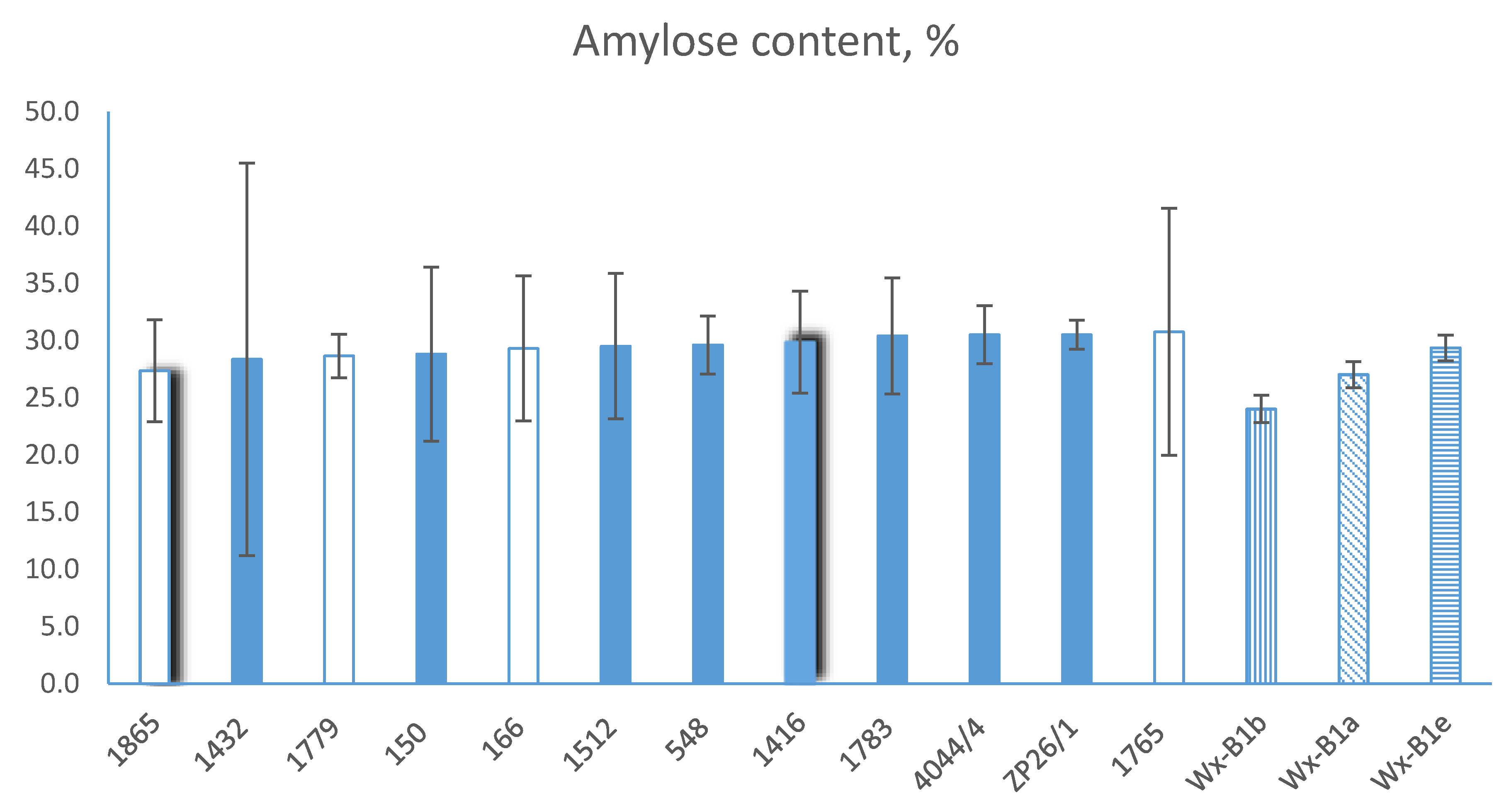
Despite slight variations in the level of amylose between different WWGH accessions, we did not reveal any significant differences (t-test, p = 0.05). At the same time, there were visible differences between the bread wheat control accessions with different allele variants of the Wx-B1 allele, which confirms the accuracy of the experiment. Furthermore, accession ZP26/1 demonstrated a higher level of amylose in comparison to the Wx-B1b and Wx-B1a lines, showing an amylose/amylopectin ratio at the level of Wx-B1e (statistically significant at p = 0.05; Table 2). Generally, we can note that in our experiments, the WWGH accessions had a slightly higher amylose content than that of the bread wheat with the wild-type allele Wx-B1a, and a content similar to that of the wheat line with the Wx-B1e allele (29.4%).
3.2. Analysis of Wx-Th Gene Expression Products in WWGH Accessions
We did not find statistically significant differences in the amylose levels in starches of different WWGH accessions (t-test, p = 0.05; Table 2). This may be due to the lack of significant influence of the wheatgrass waxy genes on the amylose/amylopectin ratio in the WWGH starch, as well as the fact that these genes are not expressed in an alien genomic background. Thus, the next step in our work was to assess the presence of protein products of wheatgrass origin in WWGH starch.
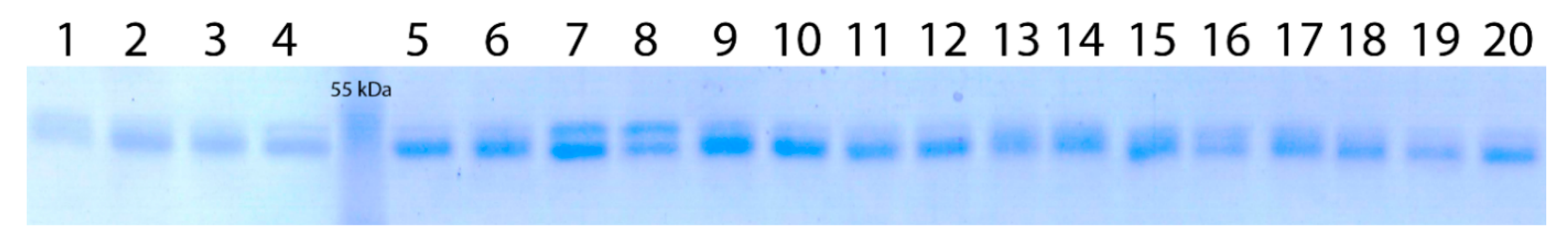
The one-dimensional SDS-PAGE electrophoresis of the protein starch fraction isolated from the WWGH grain revealed that 10 of the 12 analyzed WWGH accessions showed protein patterns similar to that characteristic of bread wheat (Figure 2). However, the lower band had a more intense color. As the upper band of wheat corresponds to the Wx-A1 expression product, and the lower band corresponds to expression products of two loci Wx-D1 and Wx-B1 [26,31], we cannot exclude the possibility that the wheatgrass Wx-Th expression product has the same size as the lower band does. This did not allow us to identify it using one-dimensional electrophoresis. Two accessions, 1416 and 1865, had patterns different from that of wheat: the upper band was consistently more intense than the lower one. This can be explained by the presence of an additional band, close in size to Wx-A1. This is probably the wheatgrass genomic Wx-Th expression product. The different identified patterns, according to the SDS-PAGE electrophoresis of waxy proteins in the WWGH accessions, show, firstly, that wheatgrass waxy genes are expressed in WWGH genomes, at least in some accessions. Secondly, they demonstrate the polymorphism of the studied WWGH accessions, not only in the wheatgrass Wx genes, but also in the level of their expression, and the resultant proteins participating in starch development.
3.3. Development and Verification of the PCR-Marker for Wx-Th
To identify the presence of GBSSI genes of wheatgrass origin in the analyzed WWGH accessions, it was necessary to develop a molecular marker. To develop such a marker is problematic due to the high degree of homology of the orthologous gene sequences in the closely related species. Using the existing sequences for Th. intermedium and Th. ponticum, it was impossible to identify a specific marker that would show numerous allele variants of the waxy gene of wheatgrass origin, and, at the same time, would not produce false positive results due to the amplification from the bread wheat genome. This is because of the fact that only partial sequences of wheatgrass Wx genes are known, and they are highly conservative.
Therefore, we conducted the sequencing of partial nucleotide sequences for four wild grasses. Two species of the four, Th. intermedium and Th. ponticum, were used for the development of WWGH. The other two species (Thinopyrum bessarabicum and Pseudoroegneria stipifolia), are carriers of the main subgenomes (Jb and St) that evolved in the speciation of Th. ponticum and Th. intermedium allopolyploids. The sequences and potential amino acid sequences that we identified are described below.
Based on the Wx gene sequences that we identified in the studied wild grasses, primers for the WXTH marker were developed in such a way that they would not yield positive amplifications with the bread wheat Wx genes (Table 1).
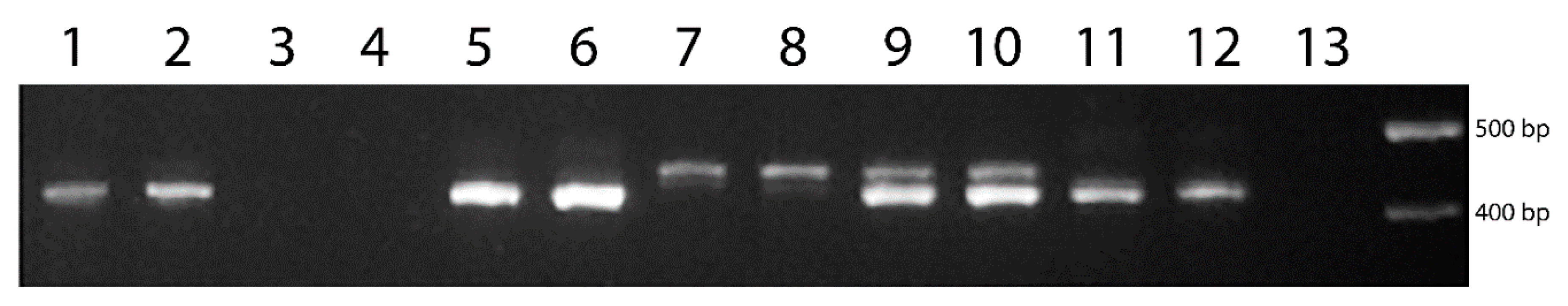
The primers were chosen to provide no amplification of the wheat genes. Figure 3 shows that the developed WXTH marker results in no amplification with wheat DNA, and positive amplification on the DNA of P. stipifolia, Th. bessarabicum, Th. intermedium and Th. ponticum.
Moreover, there is a slight difference in the sizes of the amplicons obtained using WXTH marker (Figure 3). However, further research, with a greater number of different accessions of wild grasses related to wheat, is necessary in order to assess the extent of the specificity of this difference for a particular genome. Therefore, we recommend using this marker only to establish the presence/absence of the waxy genes of wheatgrass, based on the presence/absence of the amplification.
For additional verification of the obtained marker, and assessment of the localization of the Wx gene on the chromosomes of wild relative species, we used a series of seven disomic addition Th. elongatum-wheat lines (2n = 44) that besides 42 wheat chromosomes carried an additional pair of chromosomes of diploid tall wheatgrass (from 1Je to 7 Je). We conducted a PCR with a series of seven addition lines of Chinese spring, each having an additional pair of chromosomes of Th. elongatum from each of the homeologous groups.
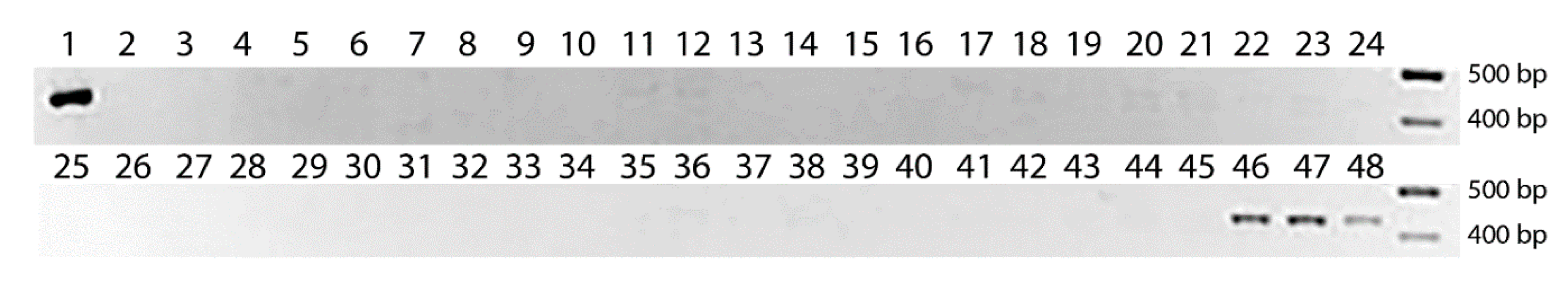
Figure 4 shows that the WXTH marker yielded positive amplification only in the line with substituted 7E chromosome, and nothing in the other lines. Therefore, we can draw the conclusion that, in Th. elongatum (genome Je), the Wx-Je gene is localized on the seventh chromosome. Thus, the verification of our molecular marker with the use of a series of addition lines led us to the conclusion that there is no non-specific amplification from the wheat subgenomes, as well as that there is no non-specific amplification from other chromosomes of wild wheat relatives, at least from those carrying the J genome.
3.4. Study of Allele Polymorphism for the Wx-Th Gene in Partial Wheat–Wheatgrass Hybrids
Our molecular marker WXTH showed specific amplification on the DNA of the Wx orthologous genes of wheatgrass; at the same time, there were no amplification products on the wheat DNA. This permitted the use of it for detection of the Wx-Th genes in the genomes of the 12 investigated partial wheat–Thinopyrum amphidiploids developed using different wheat varieties, Th. ponticum and Th. intermedium. We identified the target amplicon in eight WWGH accessions, and four WWGH accessions lacked this target PCR product, designated as amplification type 1 (charts with blue solid fill-in, Figure 5) and amplification type 2 (charts with no fill-in, Figure 5), respectively. It should be noted that the absence of the amplification product of the WXTH marker does not ultimately exclude the presence of the Wx-Th gene of wheatgrass in the studied WWGH accessions. Since it is impossible to cover the whole allele diversity of wheat’s wild relatives, due to the wide intra- and interpopulation polymorphism, the negative WXTH amplification in certain WWGH accessions may be explained by the nucleotide sequence of the Wx-Th alleles, which prevents WXTH1F/R primer annealing. However, the polymorphism of different WWGH accessions, which was identified using our marker, indicates, at least, the diversity of the Wx-Th genes of wheatgrass in the studied accessions.
Therefore, the identified polymorphisms in the waxy genes of wheatgrass, on the genetic and protein levels, did not have any significant adverse effect on the amylose and amylopectin ratio in the starch of the WWGH grain (Figure 5).
3.5. Sequencing and Characterization of the Nucleotide Sequence of the Wx Gene in Wild Relatives of Wheat
In accordance with previous publications on the production and characterization of the nucleotide sequences of the Wx genes in Triticeae [27,37,38], we designated the obtained sequences in the following manner: Wx-Jb1 for Th. bessarabicum (2x = 2n = 14, JbJb), Wx-Psstip1 for P. stipifolia (2x = 2n = 28, St1St1St2St2), Wx-Thinter1 for Th. intermedium (2n = 6x = 42, JrJvsSt), and Wx-Thpon1 for Th. ponticum (2n = 10x = 70 JJJJsJs). Fragment I covers exons 2 (partially) through 4 (introns 2 and 3, respectively), fragment II covers exons 4–7 and introns 4–6, and fragment III covers exons 7 through 12 (partially) and introns 7–11. It should be noted that the first exon of the Wx gene of wheat contains no protein-encoding sequence, and the initiation codon is located in the second exon. The sequences were registered with the GenBank NCBI database (MT371402-MT371427).
We identified several variants in the sequences for each gene fragment of each species. The different variants are due to both the different subgenomes in polyploids and the probable heterozygosity of the accessions. The resulting fragments differed from each other both in size and nucleotide sequences (Figures S1–S3).
Table 3 shows the results of the analysis of exon and intron size. The comparison of the sizes of the Wx genes from the studied wild species, with each other and with that of bread wheat, showed that the corresponding exons in all the genomes of the wild relative species and wheat are almost similar in size. Exon 6 was the only exception. At the border of intron 5 and exon 6 in P. stipifolia, Th. intermedium and T. ponticum, some of the obtained sequences had a 14-nucleotide deletion. In the transit peptide, we identified the Met68->Val substitution characteristic in the species under study, which is absent in wheat and Aegilops tauschii. Using PROVEAN software, we identified amino acid substitutions that can have a deleterious effect on the enzyme structure and activity. In the case of the preservation of the activity of the GBSSI protein encoded by the Wx genes in wild cereal species, except for Th. bessarabicum, it should be reduced, due to the numerous amino acid substitutions with a PROVEAN score of less than −2.5 (Table S1). We compared the identified amino acid substitutions with the functional motifs described by Leterrier et al., 2008 [79]. In fragment II, we established that there are critical amino acid substitutions, in the Wx-Thinter1 and Wx-Thpon1 sequences, in the 380SLoop/Strech of the polypeptide chain conservative motifs and Motif II. In fragment III, we established that there are critical amino acid substitutions in Wx-Psstip1 and Wx-Thpon1 sequences, in the conservative Motif VII–VIII.
Thus, bioinformatic analysis of the nucleotide sequences confirmed our experimental data. The Wx genes of wheatgrass have no significant effect on the increase of amylose levels in starch in the presence of wheat functional alleles.
4. Discussion
Starch quality management is one of the strategic directions of wheat breeding, as it allows the manufacturing of a wide spectrum of food products that meet the needs of regional markets, as well as of bioethanol production. One of the methods for the extension of the available assortment of Wx gene alleles, responsible for the synthesis of starch, is to involve the tertiary genetic pool in the breeding process. We have identified nucleotide sequences of the Wx-Th genes in P. stipifolia, Th. bessarabicum, Th. ponticum and Th. intermedium. The size of the identified protein is close to that of the wheat GBSSI protein. By their structure, the identified nucleotide and hypothetical amino acid sequences are close to the known sequences of wheat, Aegilops, barley, etc. At the same time, we have identified a number of substitutions, which, as we hypothesized, can impair the performance of these alleles. In their work, Alvarez et al., 2019, analyzed the theoretical Wx amino acid sequence of Hordeum chilense, and revealed that amino acid substitutions in the wheat Wx gene sequence have a neutral character, and exert very limited effects on the protein activity [38]. However, the analysis of the Wx gene sequences of Ae. searsii, Triticum monococcum ssp. monococcum, T. urartu and Taeniatherum revealed amino acid substitutions, which can affect the GBSSI protein activity [27,28,37]. In Motif VII–VIII in Th. ponticum and P. stipifolia, we have found significant amino acid substitutions; the Ala496->Val substitution was revealed in the same motif in Hordeum chilense, in a work by Alvarez et al., 2019 [38]. The presence of amino acid substitutions, which leads to a hypothetical loss of the protein activity, is more characteristic of wild species of Triticum/Aegilops, as well as more distantly related ones, such as Taeniatherum, Pseudoroegneria and Thinopyrum, as compared to domesticated species. This could be due to the artificial selection taking place in the development and domestication of bread and durum wheat when functional variants of the Wx genes passed through the genetic bottleneck, providing the most effective synthesis of amylose, which is a component of the target consumption product, starch. At the same time, the expansion of knowledge regarding the diversity of Wx gene-orthologs, conservation, and the variability of separate gene segments provides a comprehensive platform for a CRIPSR-Cas9 modification of the Wx genes of wheat.
The studied WWGH accessions were developed by multiple-stage crosses between different wheat varieties, Th. intermedium and Th. ponticum. The studied lines are represented by both well-studied old lines and relatively new, uncharacterized lines. In our previous studies, we demonstrated that WWGH 1416 and ZP26 have additional, high molecular weight proteins of wheatgrass origin, that now combine with our present results of GBSSI expression profiles [80]. ZP26 was allowed for industrial cultivation as the grain-forage wheat Ostankinskaya in the late Soviet Union, and was characterized by its good to satisfactory bread-making quality [81]. The estimation of other lines, including the assessment of their bread-making qualities, is ongoing [82]. Our results contribute to an overall understanding of the influence of the wheatgrass genome on the starch and bread-making properties of wheat–wheatgrass amphidiploids, and can be used in further baking tests of WWGHs.
The amylose content analysis of wheat–Thinopyrum hybrids did not reveal any significant differences between the accessions of WWGH. In general, the studied WWGHs had, on average, slightly higher amylose contents than the Wx-B1a wild-type genotype of wheat, close to Wx-B1e; in particular, ZP26/1 had statistically significantly (p = 0.05) higher amylose content than Wx-B1a. Th. intermedium flour has a lower starch concentration than wheat, though the amylose to amylopectin ratio in it is close to that of hard red wheat [55,56,83]. The pasting temperature of wheat–wheatgrass flour blends was higher than that for pure wheat flour, which could be explained by the assembled character of the starch granules’ distribution in Th. intermedium flour [84]. Therefore, our results demonstrate that some wheatgrass starch peculiarities can be inherited and expressed in wheat–wheatgrass amphidiploids. Therefore, the utilization of WWGHs’ grains for bread-making purposes should take into consideration the possible increase in amylose content in relation to amylopectin, and the consequential adaptation of baking technology. When WWGHs are used as a breeding bridge to introgress valuable genes of wheatgrass to perspective bread wheat lines (e.g., for resistance), the concomitant linkage drag transfer, containing Wx orthologous genes, should be monitored using our PCR marker [33,85]. The molecular marker we have developed can be used for the intended transfer of Wx orthologous genes to the bread wheat genome, for assessment of their influence on the wheat starch’s properties. Additionally, it can be used to eliminate alien Wx orthologous genes in intergeneric wheat hybrids, in case of their adverse effect on the grain quality.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4395/10/7/963/s1, Figure S1: Nucleotide sequences of the Wx genes-ortholog fragment I of (from to bottom) Th.bessarabicum (Wx-Jb1-I#1), P.stipifolia (Wx-Psstip1-I#1, Wx-Psstip1-I#2), Th.intermedium (Wx-Thint1-I#1, Wx-Thint1-I#2), Th.ponticum (Wx-Thpon1-I#1) aligned against mRNA of bread wheat Wx genes, Wx-A1a (EU719608.1, mRNA), Wx-B1a (EU719610.1, mRNA), Wx-D1a (EU719612.1, mRNA). Figure S2: Nucleotide sequences of the Wx genes-ortholog fragment II of (from to bottom) of Th.bessarabicum (Wx-Jb1-II#1, Wx-Jb1-II#2), P.stipifolia (Wx-Psstip1-II#1, Wx-Psstip1-II#2, Wx-Psstip1-II#3), Th.intermedium (Wx-Thint1-II#1, Wx-Thint1-II#2, Wx-Thint1-II#3), Th.ponticum (Wx-Thpon1-II#1, Wx-Thpon1-II#2, Wx-Thpon1-II#3) aligned against mRNA of bread wheat Wx genes, Wx-A1a (EU719608.1, mRNA), Wx-B1a (EU719610.1, mRNA), Wx-D1a (EU719612.1, mRNA). Figure S3: Nucleotide sequences of the Wx genes-ortholog fragment III of (from to bottom) Th.bessarabicum (Wx-Jb1-III#1), P.stipifolia (Wx-Psstip1-III#1, Wx-Psstip1-III#2), Th.intermedium (Wx-Thint1-III#1), Th.ponticum (Wx-Thpon1-III#1, Wx-Thpon1-III#2, Wx-Thpon1-III#3, Thpon1-III#4, Thpon1-III#5) aligned against mRNA of bread wheat Wx genes, Wx-A1a (EU719608.1, mRNA), Wx-B1a (EU719610.1, mRNA), Wx-D1a (EU719612.1, mRNA). Table S1: The amino acid changes in Wx genes-orthologs in the studied wheat wild grasses assessed using PROVEAN (in three lists).
Author Contributions
Conceptualization, M.V.K. and M.G.D.; methodology, M.V.K. and M.S.B.; software, M.S.B.; validation, M.V.K. and M.G.D.; formal analysis, M.V.K. and M.S.B.; investigation, M.V.K., P.Y.K., and M.S.B.; resources, P.Y.K. and M.G.D.; data curation, P.Y.K. and M.G.D.; writing—original draft preparation, P.Y.K.; writing—review and editing, M.V.K., G.I.K., and M.G.D.; visualization, P.Y.K.; supervision, M.G.D.; project administration, G.I.K. and M.G.D.; funding acquisition, P.Y.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, grant number 18-76-00035.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, W.; Johnson, J.W.; Graybosch, R.A.; Gaines, C.S. Physicochemical properties and end-use quality of wheat starch as a function of waxy protein alleles. J. Cereal Sci. 2003, 37, 195–204. [Google Scholar] [CrossRef] [Green Version]
- James, M.G.; Denyer, K.; Myers, A.M. Starch synthesis in the cereal endosperm. Curr. Opin. Plant Biol. 2003, 6, 215–222. [Google Scholar] [CrossRef]
- Nakamura, T.; Yamamori, M.; Hirano, H.; Hidaka, S. Identification of three Wx proteins in wheat (Triticum aestivum L.). Biochem. Genet. 1993, 31, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Urbano, M.; Margiotta, B.; Colaprico, G.; Lafiandra, D. Waxy proteins in diploid, tetraploid and hexaploid wheats. Plant Breed. 2002, 121, 465–469. [Google Scholar] [CrossRef]
- Caballero, L.; Bancel, E.; Debiton, C.; Branlard, G. Granule bound starch synthase (GBSS) diversity of ancient wheat and related species. Plant Breed. 2008, 127, 548–553. [Google Scholar] [CrossRef]
- Yamamori, M.; Yamamoto, K. Effects of two novel Wx-A1 alleles of common wheat (Triticum aestivum L.) on amylose and starch properties. J. Cereal Sci. 2011, 54, 229–235. [Google Scholar] [CrossRef]
- Rodríguez-Quijano, M.; Nieto-Taladriz, M.T.; Carrillo, J.M. Polymorphism of waxy proteins in Iberian hexaploid wheats. Plant Breed. 1998, 117, 341–344. [Google Scholar] [CrossRef]
- Guzmán, C.; Caballero, L.; Moral, A.; Alvarez, J. Genetic variation for waxy proteins and amylose content in Spanish spelt wheat (Triticum spelta L.). Genet. Resour. Crop Evol. 2010, 57, 721–725. [Google Scholar]
- McIntosh, R.A.; Yamazaki, Y.; Dubcovsky, J.; Rogers, W.J.; Morris, G.; Appels, R.; Xia, X.C. Catalogue of Gene Symbols for Wheat. 2013. Available online: http://www.shigen.nig.ac.jp/wheat/komugi/genes/macgene/2013/GeneSymbol.pdf (accessed on 30 April 2020).
- Guzmán, C.; Alvarez, J.B. Wheat waxy proteins: Polymorphism, molecular characterization and effects on starch properties. Theor. Appl. Genet. 2016, 129, 1–16. [Google Scholar] [CrossRef]
- Zi, Y.; Ding, J.; Song, J.; Humphreys, G.; Peng, Y.; Li, C.; Zhu, X.; Guo, W. Grain yield, starch content and activities of key enzymes of waxy and non-waxy wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4548. [Google Scholar] [CrossRef]
- Nakamura, T.; Vrinten, P.; Saito, M.; Konda, M. Rapid classification of partial waxy wheats using PCR-based markers. Genome 2002, 45, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Konda, M.; Vrinten, P.; Nakamura, K.; Nakamura, T. Molecular comparison of Waxy null alleles in common wheat and identification of a unique null allele. Theor. Appl. Genet. 2004, 108, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, C.; Caballero, L.; Yamamori, M.; Alvarez, J. Molecular Characterization of a new Waxy allele with partial expression in spelt wheat. Planta 2012, 235, 1331–1339. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Taladriz, M.T.; Rodriguez-Quijano, M.; Carrillo, J.M. Polymorphism of waxy proteins in Spanish durum wheats. Plant Breed. 2000, 119, 277–279. [Google Scholar] [CrossRef]
- Shariflou, M.R.; Sharp, P.J. A polymorphic microsatellite in the 3 ‘end of ‘waxy’ genes of wheat, Triticum aestivum. Plant Breed. 1999, 118, 275–277. [Google Scholar] [CrossRef]
- Ortega, R.; Guzmán, C.; Alvarez, J. Molecular characterization of several Wx alleles in durum wheat. Biol. Plant 2015, 59, 220–226. [Google Scholar] [CrossRef]
- Li, S.; Zhong, X.; Zhang, X.; Rahman, M.; Lan, J.; Tang, H.; Qi, P.; Ma, J.; Wang, J.; Chen, G.; et al. Production of waxy tetraploid wheat (Triticum turgidum durum L.) by EMS mutagenesis. Genet. Resour. Crop Evol. 2020, 67, 433–443. [Google Scholar] [CrossRef]
- Lan, J.; Li, Y.; Xu, K.; Zhang, X.; Tang, H.; Qi, P.; Ma, J.; Wang, J.; Chen, G.; Pu, Z.; et al. EMS induced SNP changes led to mutation of Wx protein in common wheat. Cereal Res. Commun. 2020, 48, 233–238. [Google Scholar] [CrossRef]
- Divashuk, M.; Klimushina, M.; Karlov, G. Molecular genetic characteristics of the Wx-B1e allele from common wheat and applicability of the DNA markers for its identification. Russ. J. Genet. 2011, 47, 1428–1432. [Google Scholar] [CrossRef]
- Yan, L.; Bhave, M. Characterization of waxy proteins and waxy genes of Triticum timopheevii and T. zhukovskyi and implications for evolution of wheat. Genome 2001, 44, 582–588. [Google Scholar] [CrossRef]
- Guzmán, C.; Ortega, R.; Yamamori, M.; Peña, R.; Alvarez, J. Molecular characterization of two novel null waxy alleles in Mexican bread wheat landraces. J. Cereal Sci. 2015, 62, 8–14. [Google Scholar] [CrossRef]
- Guzmán, C.; Caballero, L.; Alvarez, J. Molecular characterisation of the Wx-B1 allelic variants identified in cultivated emmer wheat and comparison with those of durum wheat. Mol. Breed. 2010, 28, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Guzmán, C.; Caballero, L.; Gutierrez, M.; Alvarez, J. Polymorphism of waxy proteins in Spanish hulled wheats. Plant Genet. Resour. 2011, 9, 330–333. [Google Scholar] [CrossRef] [Green Version]
- Guzmán, C.; Caballero, L.; Alvarez, J.; Yamamori, M. Amylose content and starch properties in emmer and durum wheat lines with different waxy proteins composition. J. Sci. Food Agric. 2011, 91, 1625–1629. [Google Scholar] [CrossRef]
- Guzmán, C.; Alvarez, J. Molecular characterization of a novel waxy allele (Wx-A u 1a) from Triticum urartu Thum. ex Gandil. Genet. Resour. Crop Evol. 2012, 59, 971–979. [Google Scholar] [CrossRef]
- Guzmán, C.; Caballero, L.; Martín, L.; Alvarez, J. Waxy genes from spelt wheat: New alleles for modern wheat breeding and new phylogenetic inferences about the origin of this species. Ann. Bot. 2012, 110, 1161–1171. [Google Scholar] [CrossRef] [Green Version]
- Ortega, R.; Alvarez, J.; Guzmán, C. Characterization of the Wx gene in diploid Aegilops species and its potential use in wheat breeding. Genet. Resour. Crop Evol. 2013, 61, 369–382. [Google Scholar] [CrossRef]
- Ortega, R.; Guzmán, C.; Alvarez, J. Wx gene in diploid wheat: Molecular characterization of five novel alleles from einkorn (Triticum monococcum L. ssp. monococcum) and T. urartu. Mol. Breed. 2014, 34, 1137–1146. [Google Scholar] [CrossRef]
- Maryami, Z.; Fazeli, A. Molecular diversity and detection of Waxy genes in the Iranian wheat populations by multiplex PCR. Biotechnol. Biotechnol. Equip. 2015, 29, 869–875. [Google Scholar] [CrossRef]
- Ayala, M.; Alvarez, J.; Yamamori, M.; Guzmán, C. Molecular characterization of Waxy alleles in three subspecies of hexaploid wheat and identification of two novel Wx-B1 alleles. Theor. Appl. Genet. 2015, 128, 2427–2435. [Google Scholar] [CrossRef]
- Li, W.; Fu, B.; Li, Z.; Liu, Y.; Pu, Z.; Qi, P.; Jiang, Q.; Chen, G.; Wang, J.; Wei, Y.; et al. Characterization of the waxy gene in diploid Triticum L. and Aegilops L. species and its geographic distribution. Genet. Resour. Crop Evol. 2015, 63, 987–1002. [Google Scholar] [CrossRef]
- Alvarez, J.; Guzmán, C. Interspecific and intergeneric hybridization as a source of variation for wheat grain quality improvement. Theor. Appl. Genet. 2018, 131, 225–251. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Contreras, A.; Chaires-González, C.; Rosas-Burgos, E.; Borboa-Flores, J.; Wong-Corral, F.; Cortez-Rocha, M.; Cinco-Moroyoqui, F. Comparison of protein and starch content of substituted and complete triticales (× Triticosecale Wittmack): Contribution To Functional Properties. Int. J. Food Prop. 2014, 17, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Cornejo-Ramírez, Y.; Ramírez-Reyes, F.; Cinco-Moroyoqui, F.; Rosas-Burgos, E.; Martínez-Cruz, O.; Carvajal-Millán, E.; Cárdenas-López, J.; Torres-Chavez, P.; Osuna-Amarillas, P.; Borboa-Flores, J.; et al. Starch debranching enzyme activity and its effects on some starch physicochemical characteristics in developing substituted and complete triticales (× Triticosecale Wittmack). Cereal Chem. J. 2016, 93, 64–70. [Google Scholar] [CrossRef]
- Mergoum, M.; Sapkota, S.; ElDoliefy, A.; Naraghi, S.; Pirseyedi, S.; Alamri, M.; AbuHammad, W. Triticale (× Triticosecale Wittmack) Breeding. In Advances in Plant Breeding Strategies: Cereals; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer: Cham, Switzerland, 2019; pp. 404–452. [Google Scholar]
- Dai, S.; Jiang, J.; Jia, Y.; Xue, X.; Liu, D.; Wei, Y.; Zheng, Y.; Yan, Z. Molecular characterization and phylogenetic analysis of Wx genes from three Taeniatherum diploid species. Biol. Plant 2016, 60, 505–512. [Google Scholar] [CrossRef]
- Alvarez, J.; Castellano, L.; Recio, R.; Cabrera, A. Wx gene in Hordeum chilense: Chromosomal location and characterisation of the allelic variation in the two main ecotypes of the species. Agronomy 2019, 9, 261. [Google Scholar] [CrossRef] [Green Version]
- Mason-Gamer, R. Phylogeny of a genomically diverse group of Elymus (Poaceae) allopolyploids reveals multiple levels of reticulation. PLoS ONE 2013, 8, e78449. [Google Scholar] [CrossRef] [Green Version]
- Sha, L.; Fan, X.; Li, J.; Liao, J.; Zeng, J.; Wang, Y.; Kang, H.; Zhang, H.; Zheng, Y.; Zhou, Y. Contrasting evolutionary patterns of multiple loci uncover new aspects in the genome origin and evolutionary history of Leymus (Triticeae; Poaceae). Mol. Phylogenet. Evol. 2017, 114, 175–188. [Google Scholar] [CrossRef]
- Gill, B.S.; Friebe, B.; Koo, D.H.; Li, W. Crop species origins, the impact of domestication and the potential of wide hybridisation for crop improvement. In Sustaining Global Food Security: The Nexus of Science and Policy; Zeigler, R., Ed.; CSIRO Publishing: Victoria, Australia, 2019; pp. 2–35. [Google Scholar]
- Dennett, A.; Schofield, P.; Roake, J.; Howes, N.; Chin, J. Starch swelling power and amylose content of triticale and Triticum timopheevii germplasm. J. Cereal Sci. 2009, 49, 393–397. [Google Scholar] [CrossRef]
- Cui, L.; Ren, Y.; Murray, T.; Yan, W.; Guo, Q.; Niu, Y.; Sun, Y.; Li, H. Development of perennial wheat through hybridization between wheat and wheatgrasses: A review. Engineering 2018, 4, 507–513. [Google Scholar] [CrossRef]
- Alvarez, J.; Ballesteros, J.; Sillero, J.; Martin, L. Tritordeum: A new crop of potential importance in the food industry. Hereditas 1992, 116, 193–197. [Google Scholar] [CrossRef]
- Martín, A.; Alvarez, J.; Martín, L.; Barro, F.; Ballesteros, J. The development of Tritordeum: A novel cereal for food processing. J. Cereal Sci. 1999, 30, 85–95. [Google Scholar] [CrossRef]
- Alvarez, J.; Campos, L.; Martin, A.; Martin, L. Influence of HMW and LMW glutenin subunits on gluten strength in hexaploid Tritordeum. Plant Breed. 1999, 118, 456–458. [Google Scholar] [CrossRef]
- Shapovalova, N. Two New Agricultural Crops will Appear on Russian Fields at Once (In Russian). Agro XXI (Agroindustrial Portal). 2019. Available online: https://www.agroxxi.ru/zhurnal-agromir-xxi/novosti/na-rossiiskih-poljah-pojavjatsja-srazu-dve-novye-agrokultury.html (accessed on 30 April 2020).
- Kroupin, P.; Divashuk, M.; Karlov, G. Gene resources of perennial wild cereals involved in breeding to improve wheat crop. Sel’skokhozyaistvennaya Biol. 2019, 54, 409–425. [Google Scholar] [CrossRef]
- Kocheshkova, A.; Kroupin, P.; Bazhenov, M.; Karlov, G.; Pochtovyy, A.; Upelniek, V.; Belov, V.; Divashuk, M. Pre-harvest sprouting resistance and haplotype variation of ThVp-1 gene in the collection of wheat-wheatgrass hybrids. PLoS ONE 2017, 12, e0188049. [Google Scholar] [CrossRef] [Green Version]
- Hayes, R.; Wang, S.; Newell, M.; Turner, K.; Larsen, J.; Gazza, L.; Anderson, J.; Bell, L.; Cattani, D.; Frels, K.; et al. The performance of early-generation perennial winter cereals at 21 sites across four continents. Sustainability 2018, 10, 1124. [Google Scholar] [CrossRef] [Green Version]
- Trifonova, A.; Boris, K.; Dedova, L.; Melnik, V.; Ivanova, L.; Kuzmina, N.; Zavgorodniy, S.; Upelniek, V. genome polymorphism of the synthetic species X Trititrigia cziczinii Tsvel. inferred from AFLP analysis. Vavilov J. Genet. Breed. 2018, 22, 648–653. [Google Scholar] [CrossRef]
- Larkin, P.; Newell, M.; Hayes, R.; Aktar, J.; Norton, M.; Moroni, S.; Wade, L. Progress in developing perennial wheats for grain and grazing. Crop Pasture Sci. 2014, 65, 1147. [Google Scholar] [CrossRef]
- Tsitsin, N.V. Mnogoletnyaya Pshenitsa; Nauka: Moscow, Russia, 1987. (In Russian) [Google Scholar]
- Gazza, L.; Galassi, E.; Ciccoritti, R.; Cacciatori, P.; Pogna, N. Qualitative traits of perennial wheat lines derived from different Thinopyrum species. Genet. Resour. Crop Evol. 2016, 63, 209–219. [Google Scholar] [CrossRef]
- Zhong, Y.; Mogoginta, J.; Gayin, J.; Annor, G. Structural characterization of intermediate wheatgrass (Thinopyrum intermedium) starch. Cereal Chem. 2019, 96, 927–936. [Google Scholar] [CrossRef]
- Marti, A.; Bock, J.; Pagani, M.; Ismail, B.; Seetharaman, K. Structural characterization of proteins in wheat flour doughs enriched with intermediate wheatgrass (Thinopyrum intermedium) flour. Food Chem. 2016, 194, 994–1002. [Google Scholar] [CrossRef]
- Li, H.; Wang, X. Thinopyrum ponticum and Th. intermedium: The promising source of resistance to fungal and viral diseases of wheat. J. Genet. Genom. 2009, 36, 557–565. [Google Scholar] [CrossRef]
- Mago, R.; Zhang, P.; Xia, X.; Zhang, J.; Hoxha, S.; Lagudah, E.; Graner, A.; Dundas, I. Transfer of stem rust resistance gene Srb from Thinopyrum ponticum into wheat and development of a closely linked PCR-based marker. Theor. Appl. Genet. 2018, 132, 371–382. [Google Scholar] [CrossRef]
- Wang, S.; Wang, C.; Wang, Y.; Wang, Y.; Chen, C.; Ji, W. Molecular cytogenetic identification of two wheat–Thinopyrum ponticum substitution lines conferring stripe rust resistance. Mol. Breed. 2019, 39, 143. [Google Scholar] [CrossRef]
- Zheng, X.; Tang, C.; Han, R.; Zhao, J.; Qiao, L.; Zhang, S.; Qiao, L.; Ge, C.; Zheng, J.; Liu, C. Identification, characterization, and evaluation of novel stripe rust-resistant wheat–Thinopyrum intermedium chromosome translocation lines. Plant Dis. 2020, 104, 875–881. [Google Scholar] [CrossRef]
- Ali, N. Wheat–Thinopyrum intermedium introgression lines enhancing wheat streak mosaic virus (WSMV) resistance. In Climate Change and Food Security with Emphasis on Wheat; Ozturk, M., Gul, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 243–255. [Google Scholar]
- Mahelka, V.; Kopecký, D.; Paštová, L. On the genome constitution and evolution of intermediate wheatgrass (Thinopyrum intermedium: Poaceae, Triticeae). BMC Evol. Biol. 2011, 11, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Larson, S.; Jensen, K.; Bushman, B.; DeHaan, L.; Wang, S.; Yan, X. genome evolution of intermediate wheatgrass as revealed by EST-SSR markers developed from its three progenitor diploid species. Genome 2015, 58, 63–70. [Google Scholar] [CrossRef]
- Guo, J.; Yu, X.; Yin, H.; Liu, G.; Li, A.; Wang, H.; Kong, L. Phylogenetic relationships of thinopyrum and triticum species revealed by Scot and CDDP markers. Plant Syst. Evol. 2016, 302, 1301–1309. [Google Scholar] [CrossRef]
- Divashuk, M.; Khuat, T.; Kroupin, P.; Kirov, I.; Romanov, D.; Kiseleva, A.; Khrustaleva, L.; Alexeev, D.; Zelenin, A.; Klimushina, M.; et al. Variation in copy number of Ty3/Gypsy centromeric retrotransposons in the genomes of Thinopyrum intermedium and its diploid progenitors. PLoS ONE 2016, 11, e0154241. [Google Scholar] [CrossRef] [Green Version]
- Linc, G.; Gaál, E.; Molnár, I.; Icsó, D.; Badaeva, E.; Molnár-Láng, M. Molecular cytogenetic (FISH) and genome analysis of diploid wheatgrasses and their phylogenetic relationship. PLoS ONE 2017, 12, e0173623. [Google Scholar] [CrossRef] [Green Version]
- Divashuk, M.; Karlov, G.; Kroupin, P. Copy number variation of transposable elements in Thinopyrum intermedium and its diploid relative species. Plants 2019, 9, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroupin, P.; Kuznetsova, V.; Nikitina, E.; Martirosyan, Y.; Karlov, G.; Divashuk, M. Development of new cytogenetic markers for Thinopyrum ponticum (Podp.) Z.-W. Liu & R.-C. Wang. Comp. Cytogenet. 2019, 13, 231–243. [Google Scholar] [PubMed]
- Adebiyi, J.; Schmitt Olabisi, L.; Snapp, S. Understanding perennial wheat adoption as a transformative technology: Evidence from the literature and farmers. Renew. Agric. Food Syst. 2016, 31, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Marquardt, K.; Vico, G.; Glynn, C.; Weih, M.; Eksvärd, K.; Dalin, P.; Björkman, C. Farmer perspectives on introducing perennial cereal in Swedish farming systems: A sustainability analysis of plant traits, farm management, and ecological implications. Agroecol. Sustain. Food Syst. 2016, 40, 432–450. [Google Scholar] [CrossRef]
- Snapp, S.; Rogé, P.; Okori, P.; Chikowo, R.; Peter, B.; Messina, J. Perennial grains for Africa: Possibility or pipedream? Exp. Agric. 2018, 55, 251–272. [Google Scholar] [CrossRef] [Green Version]
- Kroupin, P.; Divashuk, M.; Belov, V.; Glukhova, L.; Aleksandrov, O.; Karlov, G. Comparative Molecular Cytogenetic Characterization of Partial Wheat-Wheatgrass Hybrids. Russ. J. Genet. 2011, 47, 432–437. [Google Scholar] [CrossRef]
- Litvinov, D.; Chernook, A.; Kroupin, P.; Bazhenov, M.; Karlov, G.; Avdeev, S.; Divashuk, M. A Convenient co-dominant marker for height-reducing Ddw1 allele useful for marker-assisted selection. Agriculture 2020, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Monari, A.; Simeone, M.; Urbano, M.; Margiotta, B.; Lafiandra, D. Molecular characterization of new waxy mutants identified in bread and durum wheat. Theor. Appl. Genet. 2005, 110, 1481–1489. [Google Scholar] [CrossRef]
- Nicholas, K.B.; Nikolas, H.B., Jr. GeneDoc: A Tool for Editing and Annotating Multiple Sequence Alignments. 1997. Available online: https://genedoc.software.informer.com/download/ (accessed on 30 April 2020).
- Choi, Y.; Chan, A.P. PROVEAN web server: A tool to predict the functional effect of amino acid substitutions and indels. Bioinformatics 2015, 31, 2745–2747. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Sharp, P. An improved 1-D SDS–PAGE method for the identification of three bread wheat «Waxy» proteins. J. Cereal Sci. 1996, 23, 191–193. [Google Scholar] [CrossRef]
- Gibson, T.; Solah, V.; McCleary, B. A procedure to measure amylose in cereal starches and flours with Concanavalin A. J. Cereal Sci. 1997, 25, 111–119. [Google Scholar] [CrossRef]
- Leterrier, M.; Holappa, L.; Broglie, K.; Beckles, D. Cloning, characterisation and comparative analysis of a starch synthase IV gene in wheat: Functional and evolutionary implications. BMC Plant Biol. 2008, 8, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Divashuk., M.; Krupin, P.; Bazhenov., M.; Klimushina, M.; Belov, V.; Semyonova, E.; Karlov, I. Molecular-genetic characterization of seed storage protein composition in partial wheat-wheatgrass hybrids. Izv. Tskha 2012, 5, 29–37. (In Russian) [Google Scholar]
- Lubimova, V.; Belov, V. Grain-Forage Wheat Cultivars Zernokormovaya 169 and Zernokormovaya 26 (Recommendations for Cultivating Grain-Forage Wheat); Tsitsin Main Botanical Garden, Academy of Sciences of USSR: Moscow, Russia, 1990. [Google Scholar]
- Upelniek, V.; Belov, V.; Ivanova, l.; Dolgova, S.; Demidov, A. Heritage of academician N.V. Tsitsin: State-of-the-art and potential of the collection of intermediate wheat × couch-grass hybrids. Vavilov J. Genet. Breed. 2020, 6, 85–89. (In Russian) [Google Scholar]
- Schoenfuss, T.; Seetharaman, K.; Peterson, D. Incorporation of intermediate wheat grass in food products. (Abstr.). Cereal Foods World 2014, 59, A12. [Google Scholar]
- Marti, A.; Qiu, X.; Schoenfuss, T.; Seetharaman, K. Characteristics of perennial wheatgrass (Thinopyrum intermedium) and refined wheat flour blends: Impact on rheological properties. Cereal Chem. J. 2015, 92, 434–440. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Dundas, I.; Xu, S.; Friebe, B.; McIntosh, R.; Raupp, W. Chromosome engineering techniques for targeted introgression of rust resistance from wild wheat relatives. Methods Mol. Biol. 2017, 1659, 163–172. [Google Scholar]
Figure 1.
The schematic structure of the Wx gene of Triticeae. Blue boxes and numbers show exons, red boxes and numbers show introns. Scheme of the Wx gene is presented in accordance with [38], exon 1 (pale blue) is in the untranslated region. Fully colored arrows show primers taken from [74] for cloning Wx genes of the studied wild grasses: WxF3/WxVT1R, green arrows; WxBAF/WxVT1F, violet arrows; WxVT1F/WxVTR, turquoise arrows. The respective fragments of amplification for cloning are shown. Peach striped patterned arrows show the WXTH F/WXTH R primers developed in the current study for the identification of Wx genes-orthologs of the studied wild grasses in the wheat background.
Figure 1.
The schematic structure of the Wx gene of Triticeae. Blue boxes and numbers show exons, red boxes and numbers show introns. Scheme of the Wx gene is presented in accordance with [38], exon 1 (pale blue) is in the untranslated region. Fully colored arrows show primers taken from [74] for cloning Wx genes of the studied wild grasses: WxF3/WxVT1R, green arrows; WxBAF/WxVT1F, violet arrows; WxVT1F/WxVTR, turquoise arrows. The respective fragments of amplification for cloning are shown. Peach striped patterned arrows show the WXTH F/WXTH R primers developed in the current study for the identification of Wx genes-orthologs of the studied wild grasses in the wheat background.

Figure 2.
One-dimensional SDS–PAGE patterns of waxy proteins of wheat–wheatgrass hybrids: 1—50; 2, 3—ZP26/1; 4—1783; 5, 6—4044/4; 7, 8—1416; 9, 10—1779.1; 11, 12—166; 13—1512; 14, 15—1765; 16—1865; 17, 18—1432; 19, 20—548. PageRuler™ Protein Ladder #SM0671 (Fermentas) was used as a protein size marker.
Figure 2.
One-dimensional SDS–PAGE patterns of waxy proteins of wheat–wheatgrass hybrids: 1—50; 2, 3—ZP26/1; 4—1783; 5, 6—4044/4; 7, 8—1416; 9, 10—1779.1; 11, 12—166; 13—1512; 14, 15—1765; 16—1865; 17, 18—1432; 19, 20—548. PageRuler™ Protein Ladder #SM0671 (Fermentas) was used as a protein size marker.

Figure 3.
Amplification products obtained using WXTH marker in wheat and the studied related wild species: 1,2—WWGH (ZP26/1); 3, 4—T. aestivum; 5, 6—P. stipifolia; 7, 8—Th. bessarabicum; 9, 10—Th. intermedium; 11, 12—Th. ponticum; 13—T. aestivum. 100 bp DNA Ladder (Fermentas) was used as a DNA size marker.
Figure 3.
Amplification products obtained using WXTH marker in wheat and the studied related wild species: 1,2—WWGH (ZP26/1); 3, 4—T. aestivum; 5, 6—P. stipifolia; 7, 8—Th. bessarabicum; 9, 10—Th. intermedium; 11, 12—Th. ponticum; 13—T. aestivum. 100 bp DNA Ladder (Fermentas) was used as a DNA size marker.

Figure 4.
PCR analysis of chromosomal location of Th. elongatum Wx gene-orhtolog using WXTH marker in the series of wheat–Th. elongatum addition lines: 1—Th. elongatum; 2–8—1E add, 9–19—2E add; 20–26—3E add; 21–32—4E add; 33–39—5E add; 40–45—6E add; 46–48—7E add. 100 bp DNA Ladder (Fermentas) was used as a DNA size marker.
Figure 4.
PCR analysis of chromosomal location of Th. elongatum Wx gene-orhtolog using WXTH marker in the series of wheat–Th. elongatum addition lines: 1—Th. elongatum; 2–8—1E add, 9–19—2E add; 20–26—3E add; 21–32—4E add; 33–39—5E add; 40–45—6E add; 46–48—7E add. 100 bp DNA Ladder (Fermentas) was used as a DNA size marker.

Figure 5.
The amylose content in the accessions of the wheat–wheatgrass hybrids (%). Blue solid fill, type 1 of WXTH amplification; no fill, type 2 of WXTH amplification; pattern fill, common wheat control accessions: vertical pattern, Wx-B1b (null allele); diagonal pattern, Wx-B1a (Nemchinovskaya 56); horizontal pattern, Wx-B1e. Chart columns with shadows (1416 and 1865) show wheat–wheatgrass accessions with putative wheatgrass bands in SDS-PAGE electrophoregram. The differences are estimated using t-test, bars show confidence interval (p = 0.05).
Figure 5.
The amylose content in the accessions of the wheat–wheatgrass hybrids (%). Blue solid fill, type 1 of WXTH amplification; no fill, type 2 of WXTH amplification; pattern fill, common wheat control accessions: vertical pattern, Wx-B1b (null allele); diagonal pattern, Wx-B1a (Nemchinovskaya 56); horizontal pattern, Wx-B1e. Chart columns with shadows (1416 and 1865) show wheat–wheatgrass accessions with putative wheatgrass bands in SDS-PAGE electrophoregram. The differences are estimated using t-test, bars show confidence interval (p = 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers used for cloning Wx genes of the studied wild grasses (taken from [74]) and WXTH primers developed in the present study for the identification of Wx genes-orthologs of the studied wild grasses in the wheat background.
Table 1.
Primers used for cloning Wx genes of the studied wild grasses (taken from [74]) and WXTH primers developed in the present study for the identification of Wx genes-orthologs of the studied wild grasses in the wheat background.
| Primer | Sequence | Annealing Temperature, °C |
|---|---|---|
| WxF3 | 5′-TCT GGT CAC GTC CCA GCT CGC CAC CT-3′ | 62 |
| WxVT1R | 5′-ACC CCG CGC TTG TAG CAG TGG AAG T-3′ | |
| WxBAF | 5′-ACT TCC ACT GCT ACA AGC GCG GGG T-3′ | 62 |
| WxBAR | 5′-GCT GAC GTC CAT GCC GTT GAC GAT G-3′ | |
| WxVT1F | 5′-CAT CGT CAA CGG CAT GGA CGT CAG C-3′ | 64 |
| WxVTR | 5′-CCA GAA GCA CGT CCT CCC AGT TCT TG-3′ | |
| WXTH F | 5’-AGG ATC CTG AAC CTC AAC AA-3’ | 64 |
| WXTH R | 5’-GAA GTC GTC GAA GGA GAA GC-3’ |
Table 2.
Amylose content in wheat–wheatgrass accessions, common wheat lines with Wx-B1b (null allele), Wx-B1a (wild-type allele, Nemchinovskaya 56) and Wx-B1e are used as controls. Confidence interval after ± is shown (t-test, p = 0.05).
Table 2.
Amylose content in wheat–wheatgrass accessions, common wheat lines with Wx-B1b (null allele), Wx-B1a (wild-type allele, Nemchinovskaya 56) and Wx-B1e are used as controls. Confidence interval after ± is shown (t-test, p = 0.05).
| № | Accession | Mean Amylose Content (% in Starch) |
|---|---|---|
| 1 | 1865 | 27.4 ± 4.4 |
| 2 | 1432 | 28.4 ± 17.2 |
| 3 | 1779 | 28.7 ± 1.9 |
| 4 | 150 | 28.8 ± 7.6 |
| 5 | 166 | 29.3 ± 6.4 |
| 6 | 1512 | 29.5 ± 6.4 |
| 7 | 548 | 29.6 ± 2.5 |
| 8 | 1416 | 29.9 ± 4.4 |
| 9 | 1783 | 30.4 ± 5.1 |
| 10 | 4044/4 | 30.5 ± 2.5 |
| 11 | ZP26/1 | 30.5 ± 1.3 |
| 12 | 1765 | 30.8 ± 10.8 |
| 13 | Wx-B1b | 24.0 ± 1.2 |
| 14 | Wx-B1a | 27.0 ± 1.2 |
| 15 | Wx-B1e | 29.4 ± 1.1 |
Table 3.
Exon and intron sizes of the Wx sequences in bread wheat (Wx-A1, Wx-B1, Wx-D1) and wheat wild relatives (Wx-Jb1—Thinopyrum bessarabicum, Wx-Psstip1—Pseudoroegneria stipifolia, Wx-Thinter1– Th. intermedium, Wx-Thpon1—Th. ponticum).
Table 3.
Exon and intron sizes of the Wx sequences in bread wheat (Wx-A1, Wx-B1, Wx-D1) and wheat wild relatives (Wx-Jb1—Thinopyrum bessarabicum, Wx-Psstip1—Pseudoroegneria stipifolia, Wx-Thinter1– Th. intermedium, Wx-Thpon1—Th. ponticum).
| Exon/Intron | Wx-A1 | Wx-B1 | Wx-D1 | Wx-Jb1 | Wx-Psstip1 | Wx-Thinter1 | Wx-Thpon1 |
|---|---|---|---|---|---|---|---|
| Exon 2 ** | 321 | 324 | 321 | 321 | 318/321 | 321/321 | 321 |
| Exon 3 | 81 | 81 | 81 | 81 | 81/81 | 81/81 | 81 |
| Exon 4 | 99 | 99 | 99 | 99 | 99/99 | 99/99 | 99 |
| Exon 5 | 154 | 154 | 154 | 154/154 | 154/154/154 | 154/154/154 | 154/154/154 |
| Exon 6 | 101 | 101 | 101 | 101/101 | 80 */80 */80 * | 80 */80 */80 * | 80 */80 */101 |
| Exon 7 | 354 | 354 | 354 | 354/354 | 354/354 | 354/348/354 | 354/354/354 |
| Exon 8 | 180 | 180 | 180 | 180 | 180/180 | 180 | 180/180/180/180/180 |
| Exon 9 | 192 | 192 | 192 | 192 | 192/192 | 192 | 192/192/192/192/192 |
| Exon 10 | 87 | 87 | 87 | 87 | 87/87 | 87 | 87/87/87/87/87 |
| Exon 11 | 129 | 129 | 129 | 129 | 129/129 | 129 | 129/129/129/129/129 |
| Exon 12 ** | 117 | 117 | 117 | 117 | 117/117 | 117 | 117/117/117/117/117 |
| Intron 2 | 82 | 99 | 90 | 81 | 81/84 | 74/81 | 81 |
| Intron 3 | 84 | 88 | 95 | 83 | 81/84 | 80/84 | 84 |
| Intron 4 | 109 | 113 | 104 | 105/108 | 106/106/106 | 106/106/106 | 106/106/105 |
| Intron 5 | 125 | 133 | 152 | 125/132 | 144 */144 */144 * | 144 */149 */144 * | 149 */145 */125 |
| Intron 6 | 99 | 69 | 141 | 99/99 | 103/103/103 | 103/103/103 | 103/103/96 |
| Intron 7 | 91 | 92 | 85 | 80 | 80/89 | 80 | 89/89/89/93/80 |
| Intron 8 | 95 | 86 | 82 | 82 | 82/99 | 82 | 82/82/82/86/82 |
| Intron 9 | 90 | 84 | 84 | 83 | 84/84 | 83 | 89/84/84/90/85 |
| Intron 10 | 98 | 97 | 98 | 98 | 98/96 | 98 | 99/98/94/96/98 |
| Intron 11 | 93 | 115 | 116 | 110 | 110/111 | 110 | 115/115/115/115/115 |
*—In case of alternative splicing. **—Sequenced partially, as the primer was located in the given region.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Klimushina, M.V.; Kroupin, P.Y.; Bazhenov, M.S.; Karlov, G.I.; Divashuk, M.G. Waxy Gene-Orthologs in Wheat × Thinopyrum Amphidiploids. Agronomy 2020, 10, 963. https://doi.org/10.3390/agronomy10070963
AMA Style
Klimushina MV, Kroupin PY, Bazhenov MS, Karlov GI, Divashuk MG. Waxy Gene-Orthologs in Wheat × Thinopyrum Amphidiploids. Agronomy. 2020; 10(7):963. https://doi.org/10.3390/agronomy10070963
Chicago/Turabian StyleKlimushina, Marina V., Pavel Yu. Kroupin, Mikhail S. Bazhenov, Gennady I. Karlov, and Mikhail G. Divashuk. 2020. "Waxy Gene-Orthologs in Wheat × Thinopyrum Amphidiploids" Agronomy 10, no. 7: 963. https://doi.org/10.3390/agronomy10070963
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.