
Abstract
Different point stimulations can induce brain activity in specific regions, and however whether these stimulations affect unique neurotransmitter transmission remains unknown. Therefore, we aimed to investigate the effect of point specificity to the brain by resolving the metabolite profiles. Eighteen Sprague–Dawley rats were randomly divided into three groups: (1) the sham group: sham acupuncture at Zusanli (ST36) without electric stimulation; (2) the Zusanli (ST36) group: electroacupuncture (EA) at ST36; and (3) the Neiguan (PC6) group: EA at PC6. Then, the metabolites from rat brain samples were measured by LC–ESI–MS. The results of a partial least squares discriminant analysis revealed the differences among the sham, ST36, and PC6 groups regarding the relative content of metabolites in the cerebral cortex, hippocampus, and hypothalamus. EA at PC6 resulted in downregulation of adenosine, adrenaline, γ-aminobutyric acid, glycine, and glutamate majorly in hippocampus, and then in cerebral cortex. Otherwise, EA at ST6 resulted in upregulation of adrenaline and arginine in hippocampus, and all stimulations showed barely change of identified neurotransmitters in hypothalamus. These differential metabolite and neurotransmitter profiles prove that brain areas can be modulated by point specificity and may provide a maneuver to understand more details of meridian.
Similar content being viewed by others

Introduction
Acupuncture stimulation can affect brain activity, including the somatosensory system, motor area, and limbic system; additionally, it can modulate the activity of specific brain areas. Hence, different points produce different effects, which is a phenomenon referred to as point specificity1. Different points on the same meridian produce brain activity in similar area. For instance, acupuncture at the liver meridian Taichong (LR3) and Zhongdu (LR6) induces activity in the ipsilateral superior parietal lobe2. Acupuncture at adjacent points LR3 and Neiting (ST44) at different meridians induces distinct cerebral activation patterns, and this accounts for the mechanism of point specific therapeutic effects3. Overall, acupoints are specific to induce certain downstream modulation.
A possible mechanism of acupuncture is the regulation of dopamine release to affect other neurotransmitters such as γ-aminobutyric acid (GABA), glutamate, serotonin, and opioids, which contribute to the balance of the central nervous system (CNS)4. Acupuncture facilitated synaptic dopamine release in a mouse model of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced Parkinson’s disease5. Acupuncture, including manual acupuncture, electroacupuncture (EA), transcutaneous electrical nerve stimulation, and laser stimulation at PC6, can reduce the incidence of postoperative nausea and vomiting, which is a similar effect to that observed from the use of antiemetics6. Acupuncture at PC6 modulates activity in the cardiovascular system by influencing the brain to secrete inhibitory neurotransmitters such as GABA, opioids, and 5-hydroxytryptamine to attenuate the activity of sympathetic neurons. This effect of acupuncture involves a long-loop pathway that includes the hypothalamic rostral ventrolateral medulla, arcuate nucleus, ventrolateral periaqueductal gray, and medullary raphe7. EA at PC6 can induce inhibition of GABA transmission to the dorsal motor nucleus of the vagus and thereby reduce the inhibition of efferent vagal motor fiber, resulting in an increase of gastric motility8. Therefore, acupuncture at PC6 can produce a protective cardiovascular effect and increase gastric motility related to the inhibition of GABA. Acupuncture at ST36 during the remaining needle phase can increase motor cortical excitation and reduce motor cortical inhibition through transcranial magnetic stimulation9. A transcriptomic analysis in mice revealed that EA at ST36 involves the GABAergic neurotransmission network of the brain10. Collectively, these findings indicate that points are specific and that the acupuncture effect is related to neurotransmitter transmission.
Different frequencies of EA applied to body specific point can induce the release of different neuropeptides in central nervous system, e.g. 2 Hz EA mediates μ receptor to induce the release of β-endorphin, endomorphin and enkephalins, 100 Hz EA is via κ receptor to cause the release of dynorphin, whereas 15 Hz EA is through μ, δ and κ receptors to induce the release of β-endorphin, endomorphin, enkephalins and dynorphin11. Our previous studies have known that 2 Hz EA applied to ST36 produces more sustained effect in reducing heart rate than 100 Hz EA in human12. By consecutive stimulation, EA at Dazhui (CV14) and Baihui (CV20) points 1 day after reperfusion then once daily for 6 consecutive days can reduce infarction volume and neurological deficit13, and also can increase long-term potentiation14 in ischemia–reperfusion injured rats.
Therefore, this study aimed to investigate the point specificity of 2 Hz EA at ST36 and PC6 for 5 consecutive days in Sprague–Dawley (SD) rats to evaluate the neurotransmitter changes in the cerebral cortex, hippocampus, and hypothalamus by examining their metabolomic profiles.
Results
Differences between the sham, PC6, and ST36 groups in the cerebral cortex, hippocampus, and hypothalamus according to PLS-DA
Dansylated brain extracts of the cerebral cortices, hippocampus, and hypothalamus were resolved by liquid chromatography–mass spectrometry and presented as a base peak ion chromatogram (Fig. 1A). All data were further subjected to Progenesis QI for PLS-DA analysis. The distribution of each sample in the PLS-DA plot indicated differences of metabolite contents among the sham, PC6, and ST36 groups in the cerebral cortices, hippocampus, and hypothalamus (Fig. 1B).

Metabolite signatures of acupoints. (A) Base-peak-ion chromatograms of electroacupuncture in the sham, ST36, and PC6 groups for the cerebral cortex, hippocampus, and hypothalamus, respectively. (B) PLS-DA plot of the cerebral cortex, hippocampus, and hypothalamus of rats presenting the difference of metabolite profiles in the sham, ST36, and PC6 groups.
Effect of EA at ST36 and PC6 on neurotransmitters in the cerebral cortex
Multiple small molecular neurotransmitters were further examined by measuring the integrated chromatogram peak area in the cerebral cortex (Table 1).
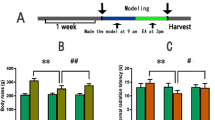
The adenosine levels of the cerebral cortices were greater in the sham group than in the ST36 and PC6 groups (both p < 0.01; Fig. 2A), whereas the adenosine levels of the cerebral cortices were similar between the ST36 and PC6 groups (p > 0.05; Fig. 2A). The adrenaline levels in the cerebral cortices were greater in the ST36 group than in the sham and PC6 groups (both p < 0.05; Fig. 2A). The adrenaline levels of the cerebral cortices were lower in the PC6 groups than sham (p < 0.01; Fig. 2A). The arginine levels of the cerebral cortices were not significantly different among the sham, ST36, and PC6 groups (all p > 0.05; Fig. 2A). The GABA levels of the cerebral cortices were greater in the sham and ST36 groups than in the PC6 groups (both p < 0.05; Fig. 2A), whereas the GABA levels of the cerebral cortices were similar between the sham and ST36 groups (p > 0.05; Fig. 2A). The glycine levels of the cerebral cortices were greater in the sham (p < 0.05) and ST36 groups than in the PC6 groups (Fig. 2A), whereas the glycine levels of cerebral cortices were similar between the sham and ST36 groups (p > 0.05; Fig. 2A). The glutamate and arginine levels of the cerebral cortices were not significantly different among the three groups (all p > 0.05; Fig. 2A).

Neurotransmitter changes induced by electroacupuncture at ST36 and PC6. The relative levels of six molecules in the (A) cerebral cortex, (B) hippocampus and (C) hypothalamus were determined by LC–MS analysis. GABA, γ-aminobutyric acid; **p < 0.05; ***p < 0.01.
Effect of EA at ST36 and PC6 on the neurotransmitters in the hippocampus
A set of signals for neurotransmitters was also obtained from the hippocampus samples during examination of the cerebral cortex (Table 2).
The adenosine levels of the hippocampi were similar among the groups (all p > 0.01; Fig. 2B). The adrenaline levels of the hippocampi in the ST36 group were greater than those in the sham and PC6 groups (both p < 0.05; Fig. 2B), whereas the adrenaline levels were similar between the sham and PC6 groups (p > 0.05; Fig. 2B). The arginine levels of the hippocampi in the ST36 group were greater than those in the sham and PC6 groups (both p < 0.01; Fig. 2B), whereas the arginine levels were similar between the sham and PC6 groups (p > 0.05; Fig. 2B). The GABA levels of the hippocampi in the sham and ST36 groups were greater than those in the PC6 groups (both p < 0.05; Fig. 2B), whereas the GABA levels of the hippocampi were similar between the sham and ST36 groups (p > 0.05; Fig. 2B). The glycine levels of the hippocampi in the sham and ST36 groups were greater than those in the PC6 groups (both p < 0.01; Fig. 2B), and the glycine levels of the hippocampi were lower in the sham than ST36 groups (p < 0.05; Fig. 2B). The glutamate levels of the hippocampi in the sham and ST36 groups were greater than those in the PC6 group (both p < 0.01; Fig. 2B), whereas the glutamate levels of hippocampi were similar between the sham and ST36 groups (p > 0.05; Fig. 2B).
Effect of EA at PC6 and ST36 on neurotransmitters in the hypothalamus
Finally, data for the same set of neurotransmitters were acquired from the hypothalamus samples (Table 3).
The adenosine, arginine, glutamate, GABA, and glycine levels of the hypothalamus were similar among the three groups (all p > 0.05; Fig. 2C). The adrenaline levels of the hypothalamus were greater in the PC6 group than in the sham and ST36 groups (p > 0.05), whereas the adrenaline levels were similar between the sham and PC6 groups and between the sham and ST36 groups (both p > 0.05; Fig. 2C).
Discussion
The results of PLS-DA in this study indicated metabolite differences among the sham, ST36, and PC6 groups in the cerebral cortices, hippocampi, and hypothalamus. This finding suggested that the signals from the sham, ST36, and PC6 points were transmitted through different pathways according to their specific location in the cerebral cortex, hippocampus, and hypothalamus (Fig. 3). In addition, our results also indicated that EA at ST36 increased adrenaline levels in the cerebral cortex and hippocampus, whereas EA at PC6 could not produce similar results. EA at PC6 could reduce the GABA and glycine levels of the cerebral cortex and hippocampus, whereas EA at ST36 could not produce similar results, suggesting that the EA at ST36 and PC6 could cause different changes in the neurotransmitters. Collectively, the results were consistent with the idea that different points modulate the activity of a corresponding brain area and have specificity1. GABA is a neurotransmitter that can rapidly inhibit synaptic transmission. The GABA system has been widely used as a target in the CNS for various drugs, such as anxiolytic, sedative–hypnotic, and anticonvulsant medications15,16. Glycine is a secondary rapid inhibitory neurotransmitter, and its receptor involves motor reflexes and nociceptive pathways, especially in the spinal cord15,16. GABA-A controls inhibitory synaptic neurotransmission, whereas glutamate controls excitatory synaptic neurotransmission. A reduction in the number of GABA-A receptors reduces synaptic inhibition, and an increase in the number of glutamatergic receptors causes an imbalance between excitation and inhibition, resulting in epileptic seizure17. GABA dysfunction and glycine disruption involve the mechanism of neuropathic pain18. The results of the present study indicated that EA at PC6 reduces GABA and glycine levels in the cerebral cortex and hippocampus (Fig. 2A, B). Further studies are required to determine whether the reduction of GABA and glycine by EA at PC6 results in epileptic seizure or pain thresholds to cause seizures or neuropathic pain to occur.

Additionally, our present results also indicated that the glutamate levels in the hippocampus were also lower in the PC6 group than in the sham and ST36 groups (Fig. 2B). Glutamate is mainly an excitatory neurotransmitter in the mammalian CNS. Glutamatergic excitotoxicity plays a critical role in the pathogenesis following an ischemic stroke; additionally, it contributes to neuropsychiatric disorders such as schizophrenia and major depressive disorder19. The results imply that applying EA at PC6 may be feasible to treat ischemic stroke or neuropsychiatric diseases.
Our results also indicated that adenosine levels of the cerebral cortices in the ST36 and PC6 groups were lower than those in the sham group (Fig. 2A). Adenosine is a neurotransmitter that serves as an endogenous distress signal and plays a modulatory role in tissue damage and repair in the nervous system20,21. The formation of adenosine mainly results from the breakdown of intracellular or extracellular adenine nucleotides when cell-membrane damage occurs, such as when transient global ischemia causes a massive increase in the extracellular adenosine triphosphate (ATP), which results in the rapid formation of extracellular adenosine. By contrast, extracellular adenosine levels decrease with the increase in intracellular adenosine formation21,22. Acupuncture may induce increases in extracellular ATP, adenosine diphosphate, adenosine monophosphate, and adenosine levels; however, the increase in adenosine level only lasts for a short duration because of the facilitated uptake of nucleoside transporters21,23. Collectively, these findings indicate that the reduction of adenosine levels in the ST36 and PC6 groups may be attributable to the movement of adenosine from the extracellular to intracellular space.
Our results also indicted that the adrenaline levels of the cerebral cortices and hippocampi in the ST36 group were greater than those in the sham and PC6 groups (Fig. 2A, B). Adrenaline is one of the principal neurotransmitters in the CNS and is involved in many CNS functions. It is suggested that noradrenaline can control the neuronal activity of the cerebral cortex or amygdala in the consolidation process of long-term memory formation24. Adrenaline can enhance hippocampal glucose metabolism after stress to support stress-related memory processing25. Therefore, EA at ST36 is perhaps more suitable than EA at PC6 for improving memory.
From the results, arginine levels in the hippocampus were greater in the ST36 group than in the PC6 group, whereas the arginine levels in the cerebral cortex were not significantly different between the two groups (Fig. 2A, B). l-arginine involves nitric oxide (NO) synthase, which is related to the generation of endogenous NO. l-arginine induces vasodilation, and its long-term oral administration can alleviate the clinical symptoms of cardiovascular disorders26. NO levels are elevated in acupoints, inducing vasodilation and leading to an increase in local blood flow and also contributes to the specificity of acupoints27. The change of arginine level in these results partly support to explain of specificity of acupoints.
One of our results requires further explanation, namely why no significant difference was observed in the adenosine, arginine, GABA, and glycine levels of the hypothalamus among the three groups, except for higher adrenaline levels of the hypothalamus in the PC6 group than in the ST36 group (Fig. 3). The specificity of acupoints and the meridian in the hypothalamic paraventricular nucleus was reported in a study using acupuncture stimulation at different 33 points28. EA at PC6 or ST36 can elevate gonadotropin-releasing hormone (GnRH) mRNA levels in the hypothalamus of female rats29. GnRH acts as a primary brain signal in the hypothalamic–pituitary–ovarian axis response to the release of follicle-stimulating hormone, luteinizing hormone, and neurotransmitters and neuropeptides such as GABA and glutamate29,30. The functional connectivity between the amygdala and hypothalamus is observed in patients with carpal tunnel syndrome subjected to acupuncture stimuli31. The hypothalamus centrally communicates acupuncture stimulation because the 2 Hz EA at LI4 can induce hypothalamus activation, which in turn extends to the insula, anterior cingulate cortex, and other brain regions32. Although acupuncture stimulation reaches hypothalamus, only moderate change has been observed on these neurotransmitters in this study. Therefore, hypothalamus may mainly coordinate signals and transmit to the cerebral cortex and hippocampus. Considering all of these findings together, we suggest that an acupuncture signal transfers from the point to the hypothalamus, after which it transfers to the cerebral cortex and hippocampus, producing physiological action possibly playing a partially critical role in acupuncture stimulation, but further study is still needed..
In conclusion, our results indicated differences among the sham, PC6, and ST36 groups in the cerebral cortex, hippocampus, and hypothalamus. EA produces different changes at ST36 and PC6 in the aforementioned neurotransmitters in the cerebral cortex and hippocampus, suggesting point specificity. The hypothalamus is a characterized central expression of acupuncture stimulation; some signals from acupuncture stimulation extend to other brain areas, possibly through the hypothalamus, but further study is needed in the future.
Methods
Animals
Male SD rats, weighing 200–300 g, were purchased from BioLASCO Taiwan Co., Ltd. The rats were kept at the animal center of China Medical University (CMU) in a 12:12 h light–dark cycle environment. The room temperature was controlled at 25 ± 1 °C. The rats had free access to food and sufficient water. The laboratory use of animals was approved by the Institutional Animal Care and Use Committee of CMU, and the Guide for the Use of Laboratory Animals (National Academy Press) was followed.
Grouping
A total of 18 SD rats were randomly divided into 3 groups, with 6 rats in each group. Rats in the sham group were anesthetized with 2% isoflurane, and acupuncture needles were subcutaneously inserted into their bilateral Zusanli (ST36) (cathode) and Shangjuxu (ST37) (anode), which have similar characteristics to those in humans. Thereafter, the needles were connected to a Trio 300 electrical stimulator (Grand Medical Instrument Co. Ltd, Ito, Japan), but electrical stimulation was not administered. This routine was performed 20 min each day for 5 consecutive days; finally, the rats were sacrificed immediately after final EA under 3–5% isoflurane anesthesia, and the rat brains were collected for metabolomic analyses. Rats in the Zusanli (ST36) group underwent the same procedure as those in the sham group, except the acupuncture needles were inserted intramuscularly at ST36 (cathode) and ST37 (anode) and an electric stimulation of 2 Hz (constant rectangle in output mode and 150 μs in wave width) was administered. The intensity of electrical stimulation was sufficient to cause visible muscle twitching. Rats in the Neiguan (PC6) group underwent the same procedure as those in the Zusanli group, except the acupuncture needles were inserted in the bilateral PC6 (cathode) and PC5 (anode).
Metabolite sample preparation and derivatization
Rat tissue samples were processed with similar procedures as our previous work33. Cerebral cortex, hippocampus, and hypothalamus tissues were respectively homogenized using 1.0-mm zirconium oxide bead grinding with a ratio of 1 mg of tissue to 10 μL of ultrapure water and then centrifuged at 14,000 rpm for 10 min. Each supernatant was collected as an aliquot of 100 μL and mixed with 300 μL of 100% methanol. After 14,000 rpm centrifugation for 10 min, 150 μL of supernatant from methanol extraction was transferred to a new microtube and then subjected to vacuum drying. Each dry metabolite sample was dissolved in a reaction solution consisting of 16 μL of ultrapure water, 2 μL of 0.5 M carbonate buffer (pH 9.4), and 2 μL of 10 mg/mL dansyl chloride prepared in acetone. The reaction was conducted at 60 °C for 2 h, and then 80 μL of ultrapure water was added before the solution was incubated for 30 min at 60 °C. After being subjected to 14,000 rpm centrifugation for 10 min, the supernatant of the dansylated sample was transferred to an insert vial and kept in an autosampler at 10 °C for further analysis.
Liquid chromatography–mass spectrometry
Samples were analyzed by our previously established methods33. The liquid chromatography–mass spectrometry (LC–MS) system consisted of an ultraperformance liquid chromatography (UPLC) system (ACQUITY UPLC I-Class, Waters) and an ESI/APCI source of 4 kDa quadrupole time-of-flight mass spectrometer (Waters VION, Waters). A BEH C18 column (2.1 × 100 mm, Walters) was used to perform reversed-phase LC. The flow rate was set at 0.2 mL/min with a column temperature of 35 °C and 5-μL sample injection. The sample contents were eluted by 99% mobile phase A (ultrapure water + 0.1% formic acid) and 99% mobile phase B (100% methanol + 0.1% formic acid), then held at 1% B for 0.5 min, raised to 90% B at 5.5 min, held at 90% B for 1 min, and then lowered to 1% B at 1 min. The column was equilibrated by pumping 1% B for 4 min. An LC–MS chromatogram was acquired using ESI + mode under the following conditions: capillary voltage of 2.5 kV, source temperature of 100 °C, desolvation temperature of 250 °C, cone gas maintained at 10 L/h, desolvation gas maintained at 600 L/h, and acquisition by MSE mode with a range of 100–1,000 m/z and a 0.5-s scan time. The acquired data were processed using UNIFI software (Waters) to summarize the integrated area of signals, and Progenesis QI with EZinfo (Waters) was used to perform partial least squares discriminant analysis (PLS-DA).
Statistical analysis
All data are expressed as the mean ± standard error. Significant differences were analyzed using one-way analysis of variance followed by Tukey’s test (post hoc). A p value less than 0.05 was considered statistically significant.
Ethics statement
The protocol was approved by the Animal Care and Use Committee of China Medical University (study on the relationship among acupoints, the brain, and visceral organs; CMUIACUC-2017-349).
Consent for publication
This study did not include any human data.
References
Huang, W. et al. Characterizing acupuncture stimuli using brain imaging with FMRI—a systematic review and meta-analysis of the literature. PLoS ONE 7, e32960 (2012).
Li, L. et al. The human brain response to acupuncture on same-meridian acupoints: evidence from an fMRI study. J. Altern. Complement. Med. 14, 673–678 (2008).
Liu, H. et al. FMRI evidence of acupoints specificity in two adjacent acupoints. Evid.-Based Complement. Altern. Med. 2013, 532581 (2013).
Yang, C. H., Lee, B. H. & Sohn, S. H. A possible mechanism underlying the effectiveness of acupuncture in the treatment of drug addiction. Evid.-Based Complement. Altern. Med. 5, 257–266 (2008).
Kim, S.-N. et al. Acupuncture enhances the synaptic dopamine availability to improve motor function in a mouse model of Parkinson’s disease. PLoS ONE 6, e27566 (2011).
Lee, A., Chan, S. K. & Fan, L. T. Stimulation of the wrist acupuncture point PC6 for preventing postoperative nausea and vomiting. Cochrane Database Syst. Rev., CD003281 (2015).
Li, J. et al. The influence of PC6 on cardiovascular disorders: a review of central neural mechanisms. Acupunct. Med. 30, 47–50 (2012).
Lu, M., Chen, C., Li, W., Yu, Z. & Xu, B. EA at PC6 promotes gastric motility: role of brainstem vagovagal neurocircuits. Evid.-Based Complement. Altern. Med. 2019, 7457485 (2019).
Sun, Z. G. et al. Effect of acupuncture at ST36 on motor cortical excitation and inhibition. Brain Behav. 9, e01370 (2019).
Wu, J.-S. et al. Comprehensive evaluation of gene expression signatures in response to electroacupuncture stimulation at Zusanli (ST36) acupoint by transcriptomic analysis. BMC Complement. Altern. Med. 17, 406 (2017).
Han, J. S. Acupuncture: neuropeptide release produced by electrical stimulation of different frequencies. Trends Neurosci 26, 17–22. https://doi.org/10.1016/s0166-2236(02)00006-1 (2003).
Hsieh, C. L., Lin, J. G., Li, T. C. & Chang, Q. Y. Changes of pulse rate and skin temperature evoked by electroacupuncture stimulation with different frequency on both Zusanli acupoints in humans. Am. J. Chin. Med. 27, 11–18. https://doi.org/10.1142/S0192415X99000033 (1999).
Cheng, C. Y., Lin, J. G., Tang, N. Y., Kao, S. T. & Hsieh, C. L. Electroacupuncture-like stimulation at the Baihui (GV20) and Dazhui (GV14) acupoints protects rats against subacute-phase cerebral ischemia-reperfusion injuries by reducing S100B-mediated neurotoxicity. PLoS ONE 9, e91426. https://doi.org/10.1371/journal.pone.0091426 (2014).
Lin, Y. W. & Hsieh, C. L. Electroacupuncture at Baihui acupoint (GV20) reverses behavior deficit and long-term potentiation through N-methyl-d-aspartate and transient receptor potential vanilloid subtype 1 receptors in middle cerebral artery occlusion rats. J. Integr. Neurosci. 9, 269–282. https://doi.org/10.1142/s0219635210002433 (2010).
Olsen, R. W. GABA. In Neuropsychopharmacology: The Fifth Generation of Progress, Chap. 12, 159–168 (2002).
Bowery, N. & Smart, T. GABA and glycine as neurotransmitters: a brief history. Br. J. Pharmacol. 147, S109–S119 (2006).
Naylor, D. E. Glutamate and GABA in the balance: convergent pathways sustain seizures during status epilepticus. Epilepsia 51, 106–109 (2010).
Alves, J. M. & Lin, K. Neuropathic pain: a review of interneuronal disinhibition. Arch. Neurosci. 5, e12290 (2018).
Niciu, M. J., Kelmendi, B. & Sanacora, G. Overview of glutamatergic neurotransmission in the nervous system. Pharmacol. Biochem. Behav. 100, 656–664 (2012).
Sebastião, A. M. & Ribeiro, J. A. Fine-tuning neuromodulation by adenosine. Trends Pharmacol. Sci. 21, 341–346 (2000).
Fredholm, B. Adenosine, an endogenous distress signal, modulates tissue damage and repair. Cell Death Differ. 14, 1315 (2007).
Zimmermann, H. Extracellular metabolism of ATP and other nucleotides. Naunyn-Schmiedeberg’s Arch. Pharmacol. 362, 299–309 (2000).
Ren, W., Tu, W., Jiang, S., Cheng, R. & Du, Y. Electroacupuncture improves neuropathic pain: adenosine, adenosine 5′-triphosphate disodium and their receptors perhaps change simultaneously. Neural Regener. Res. 7, 2618 (2012).
Kobayashi, K. Role of catecholamine signaling in brain and nervous system functions: new insights from mouse molecular genetic study. J. Invest. Dermatol. Symp. Proc. 6, 115–121 (2001).
Osborne, D. M., Pearson-Leary, J. & McNay, E. C. The neuroenergetics of stress hormones in the hippocampus and implications for memory. Front. Neurosci. 9, 164 (2015).
Böger, R. H. & Bode-Böger, S. M. The clinical pharmacology of L-arginine. Annu. Rev. Pharmacol. Toxicol. 41, 79–99 (2001).
Ma, S.-X. Nitric oxide signaling molecules in acupoints: toward mechanisms of acupuncture. Chin. J. Integr. Med. 23, 812–815 (2017).
Wang, S.-J., Zhang, J.-J., Yang, H.-Y., Wang, F. & Li, S.-T. Acupoint specificity on acupuncture regulation of hypothalamic–pituitary–adrenal cortex axis function. BMC Complement. Altern. Med. 15, 87 (2015).
Zhu, H. et al. Electro-acupuncture affects the activity of the hypothalamic–pituitary–ovary axis in female rats. Front. Physiol. 10, 466 (2019).
Tobari, Y., Son, Y. L., Ubuka, T., Hasegawa, Y. & Tsutsui, K. A new pathway mediating social effects on the endocrine system: female presence acting via norepinephrine release stimulates gonadotropin-inhibitory hormone in the paraventricular nucleus and suppresses luteinizing hormone in quail. J. Neurosci. 34, 9803–9811 (2014).
Napadow, V. et al. Hypothalamus and amygdala response to acupuncture stimuli in carpal tunnel syndrome. Pain 130, 254–266 (2007).
Hsieh, J.-C. et al. Activation of the hypothalamus characterizes the acupuncture stimulation at the analgesic point in human: a positron emission tomography study. Neurosci. Lett. 307, 105–108 (2001).
Huang, C. P., Lin, Y. W., Lee, D. Y. & Hsieh, C. L. Electroacupuncture relieves CCI-induced neuropathic pain involving excitatory and inhibitory neurotransmitters. Evid. Based Complement. Alternat. Med. 2019, 6784735. https://doi.org/10.1155/2019/6784735 (2019).
Acknowledgements
This work was financially supported by Grants DMR-109-007 and CMU108-S-32 from China Medical University Hospital and by the Chinese Medicine Research Center, China Medical University under The Featured Areas Research Center Program within the framework of the Higher Education Sprout Project by the Ministry of Education in Taiwan (CMRC-CENTER-0). Experiments and data analysis were performed in part by Metabolomics platform of the Medical Research Core Facilities, Office of Research and Development at China medical University, Taichung, Taiwan, ROC.
Author information
Authors and Affiliations
Contributions
U.-R.J. performed animal experiments; D.-Y.L. performed metabolomic analysis, including liquid chromatography–electrospray ionization–tandem mass spectrometry analysis; and C.-L.H. designed the protocol and wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lee, DY., Jiu, YR. & Hsieh, CL. Electroacupuncture at Zusanli and at Neiguan characterized point specificity in the brain by metabolomic analysis. Sci Rep 10, 10717 (2020). https://doi.org/10.1038/s41598-020-67766-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-67766-0
This article is cited by
-
Excitatory neurons in paraventricular hypothalamus contributed to the mechanism underlying acupuncture regulating the swallowing function
Scientific Reports (2022)
-
Electro-Acupuncture Pretreatment Ameliorates Anesthesia and Surgery-Induced Cognitive Dysfunction Via Inhibiting Mitochondrial Injury and nEuroapoptosis in Aged Rats
Neurochemical Research (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
