Spatiotemporal Pattern Formation in a Prey-Predator System: The Case Study of Short-Term Interactions Between Diatom Microalgae and Microcrustaceans


Abstract
:1. Introduction
2. Mathematical Model
3. Numerical Study
3.1. Numerical Approximation
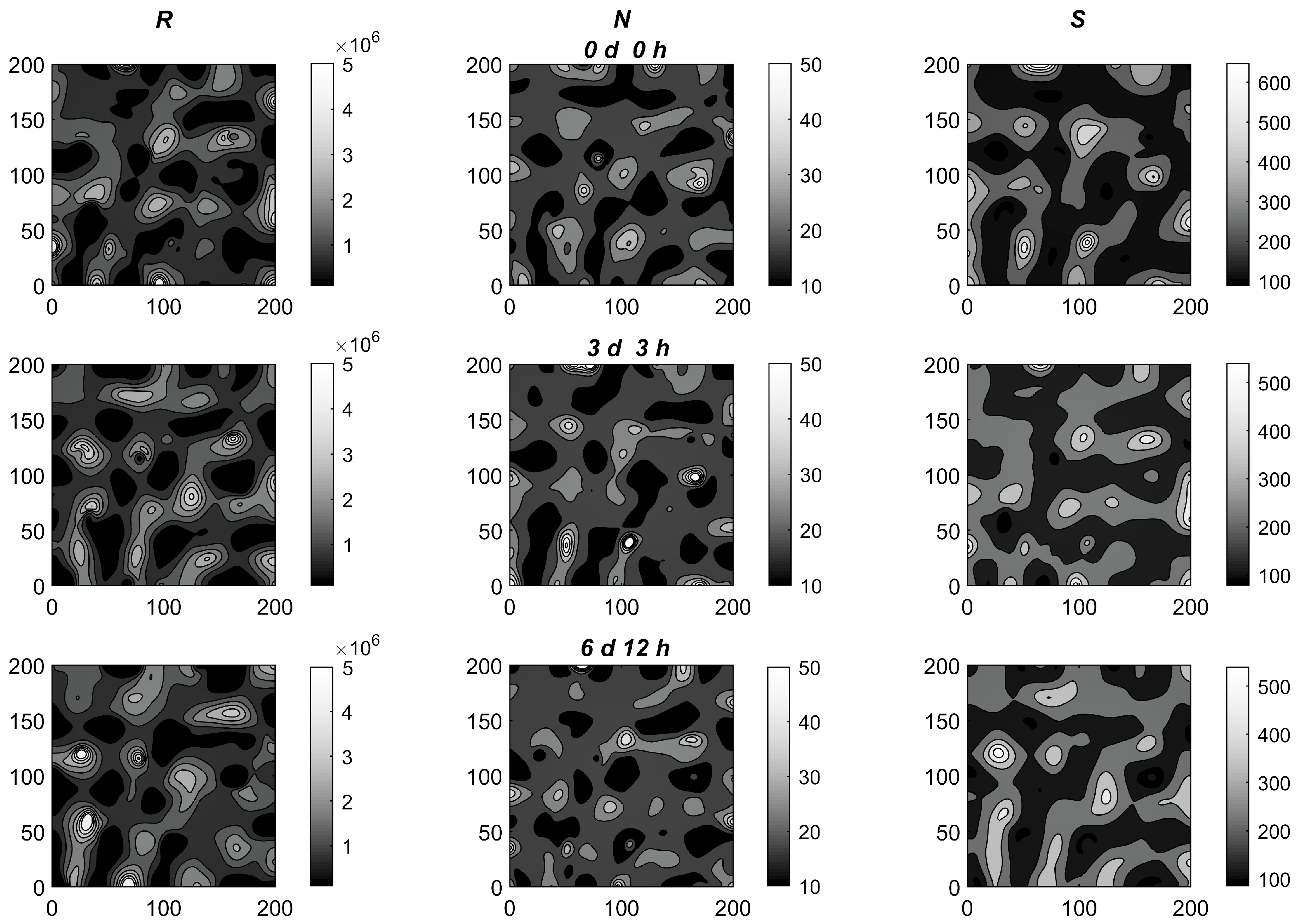
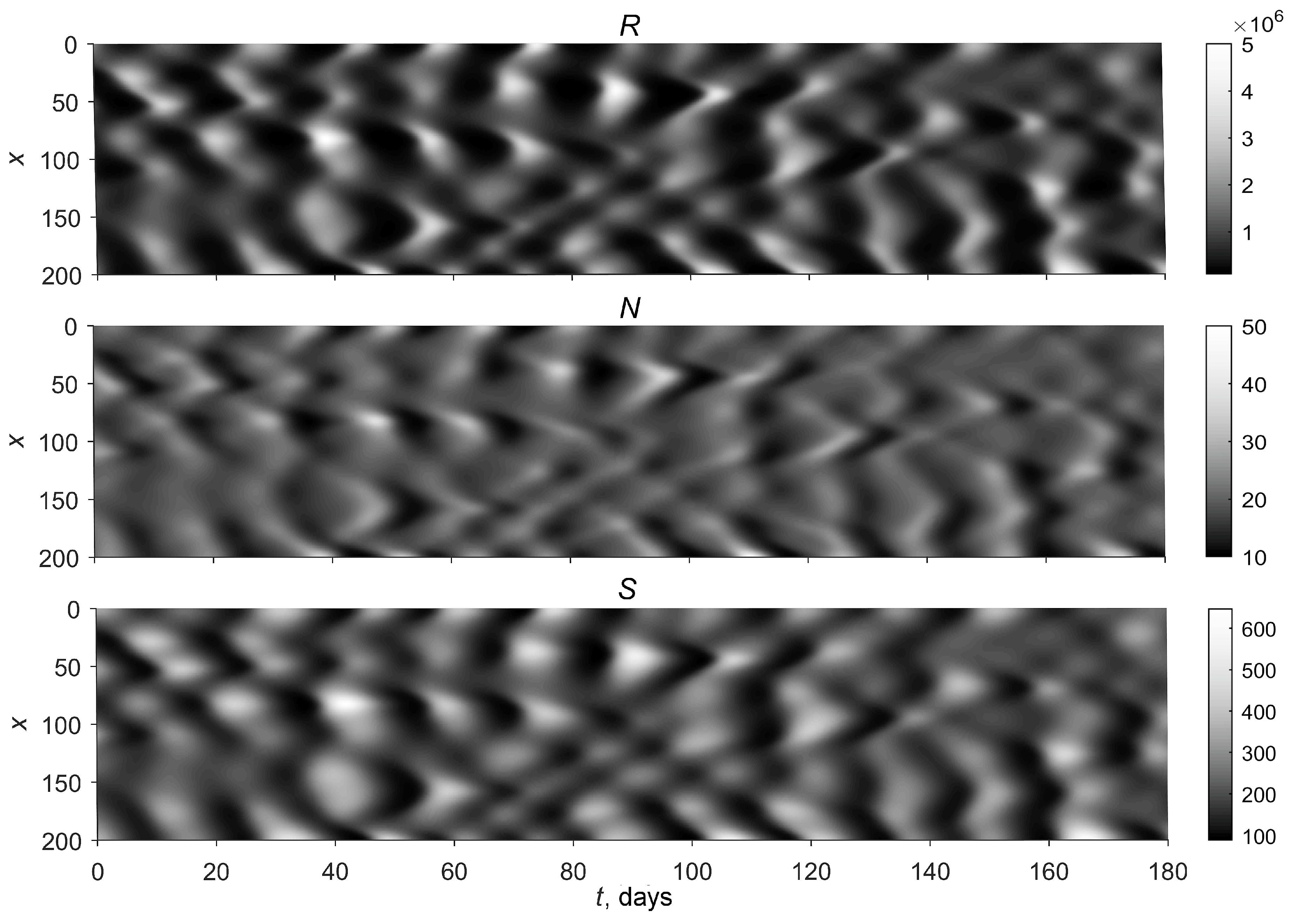
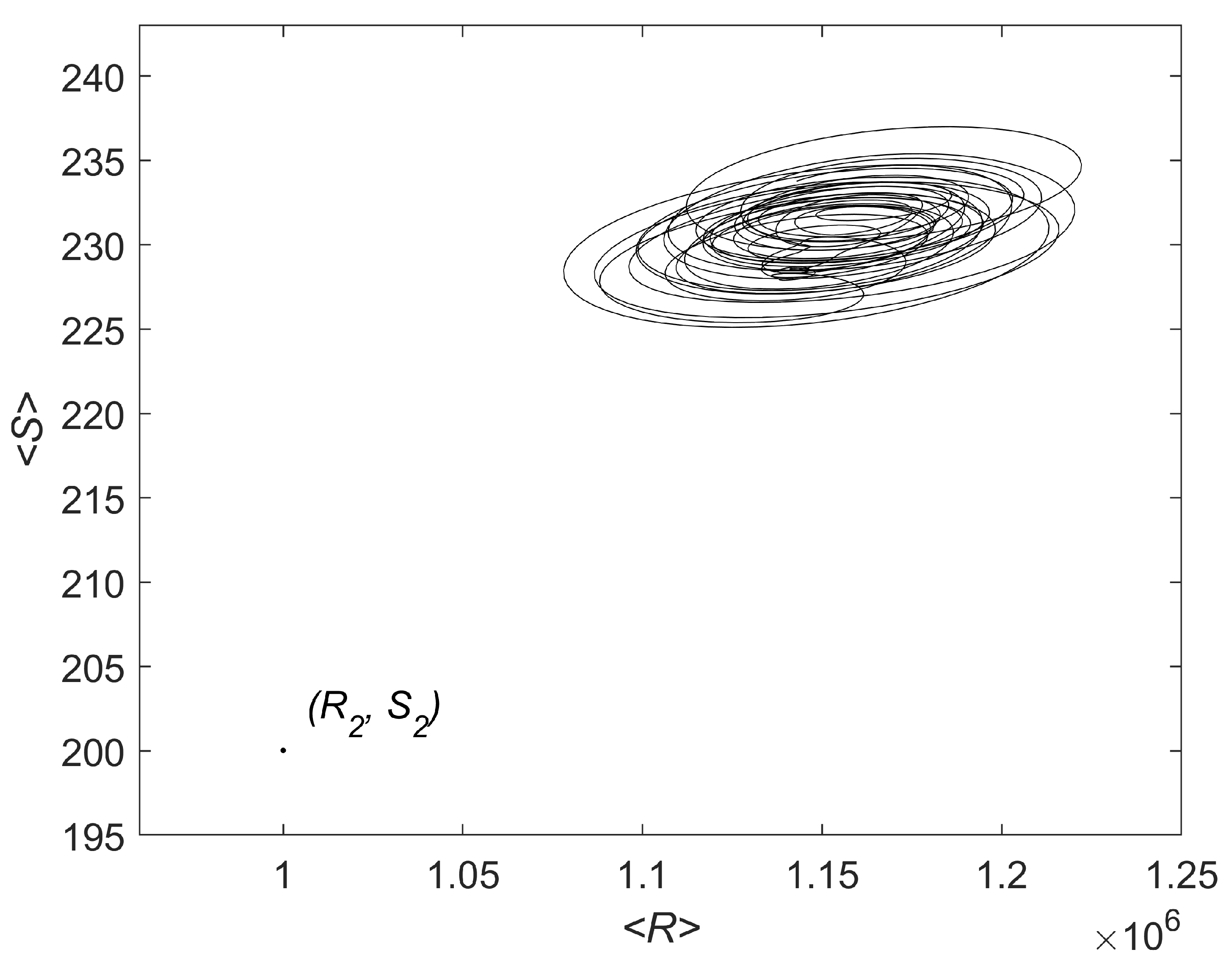
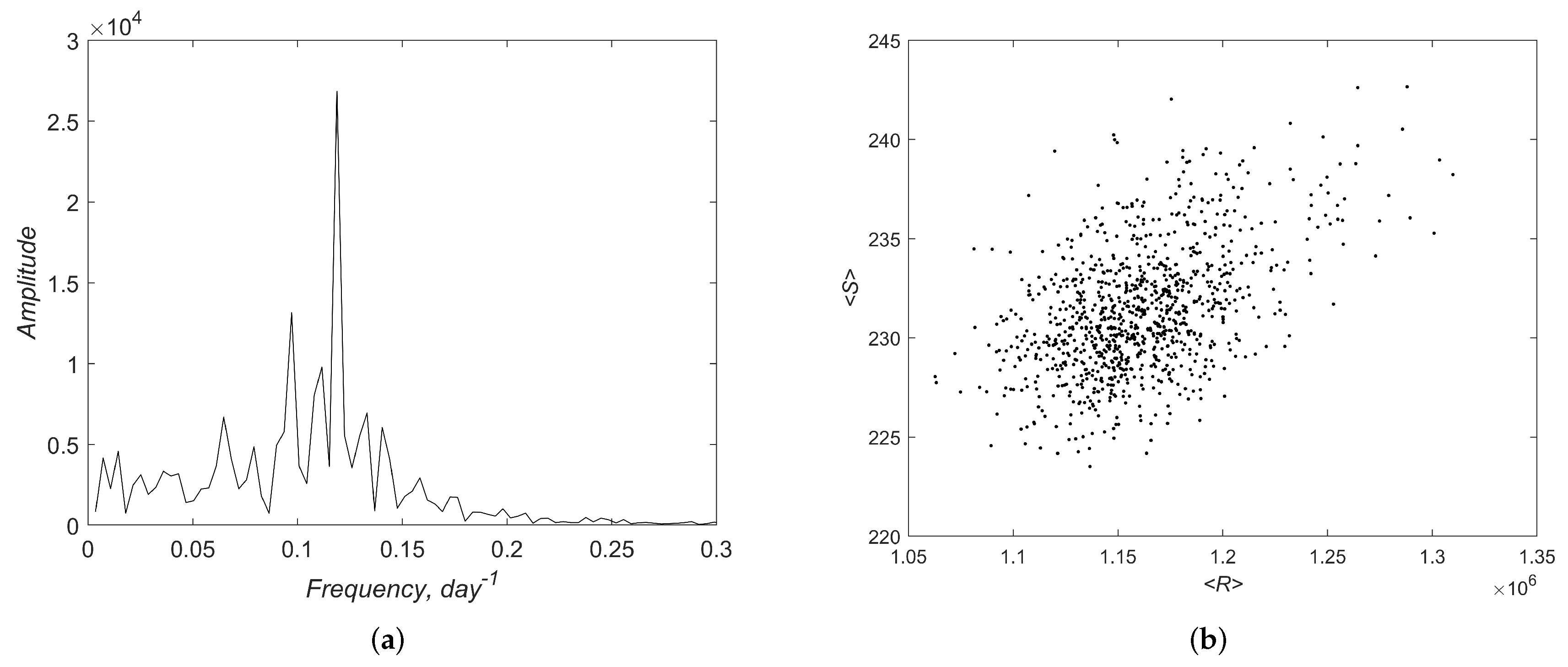
3.2. Simulations
4. Discussion and Conclusions
- The TZA model is continuous in both space and time, while the cellular automata-based SA model is spatially discrete;
- Migration process is continuous in TZA but periodic (semidiurnal) in SA model, imitating the tidal rhythm;
- in TZA, the environment is spatially homogenous, while both cases (homogeneous and heterogeneous environment for prey) are considered in SA model;
- In both models, the predator migrations are simulated as random walk; the distances of movement are unrestricted (normally distributed) in TZA, while restricted (either equiprobably distributed in a fixed neighborhood of a starting point or partly directed by a taxis) in SA model;
- Functional response of predator is the Lotka–Voltera (linear) in TZA but the Holling type III (sigmoid) in SA model;
- The relationship between the frequency of predator migrations and their degree of starvation (stimulus-response function) is described by inverse parabolic function (15) in TZA but by S-shaped function in SA model.
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PKS | Patlak—Keller—Segel |
| PDE | Partial differential equation |
| ODE | Ordinary differential equation |
| TZA | Tyutyunov—Zagrebneva—Azovsky |
| SA | Smirnova—Azovsky |
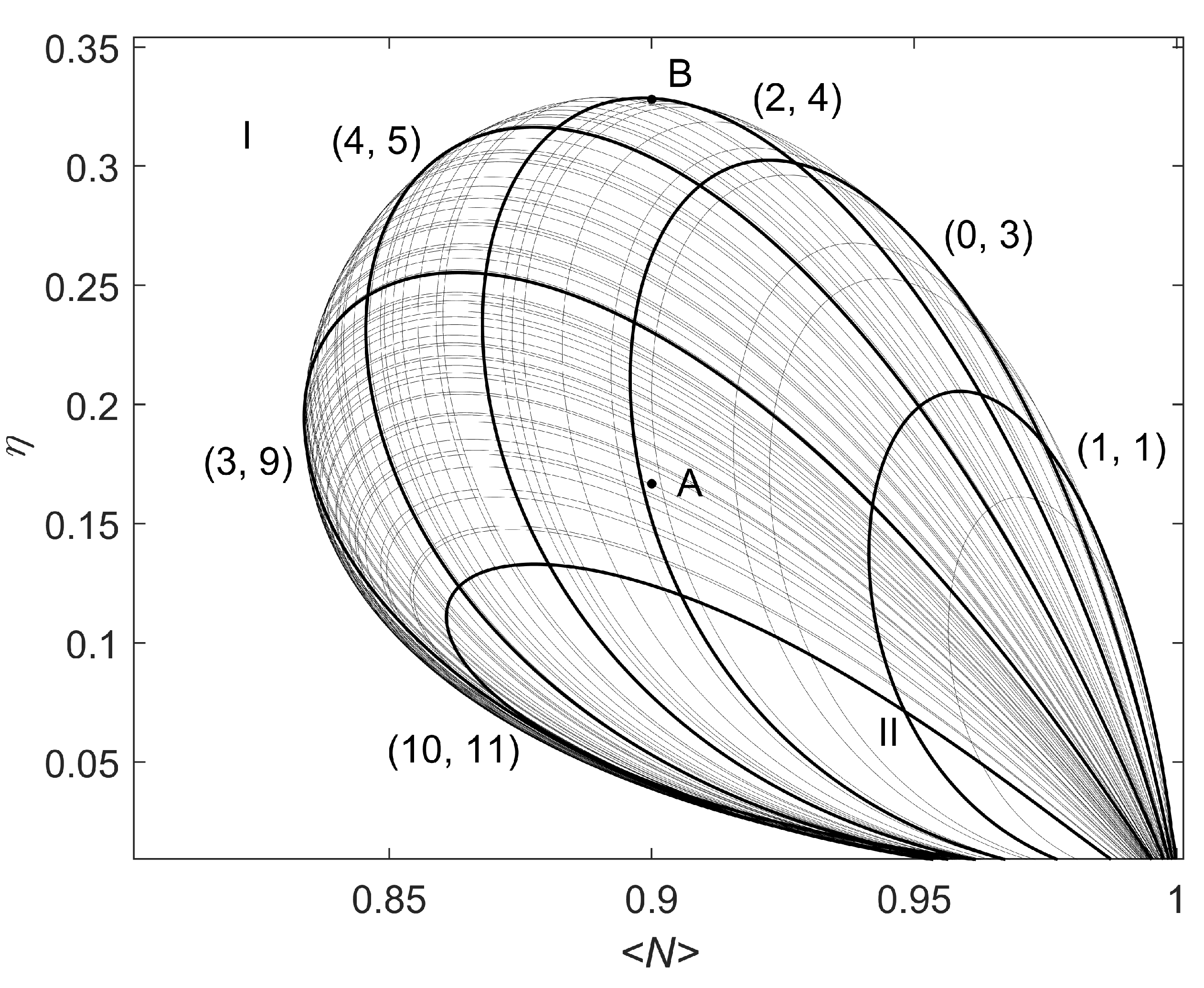
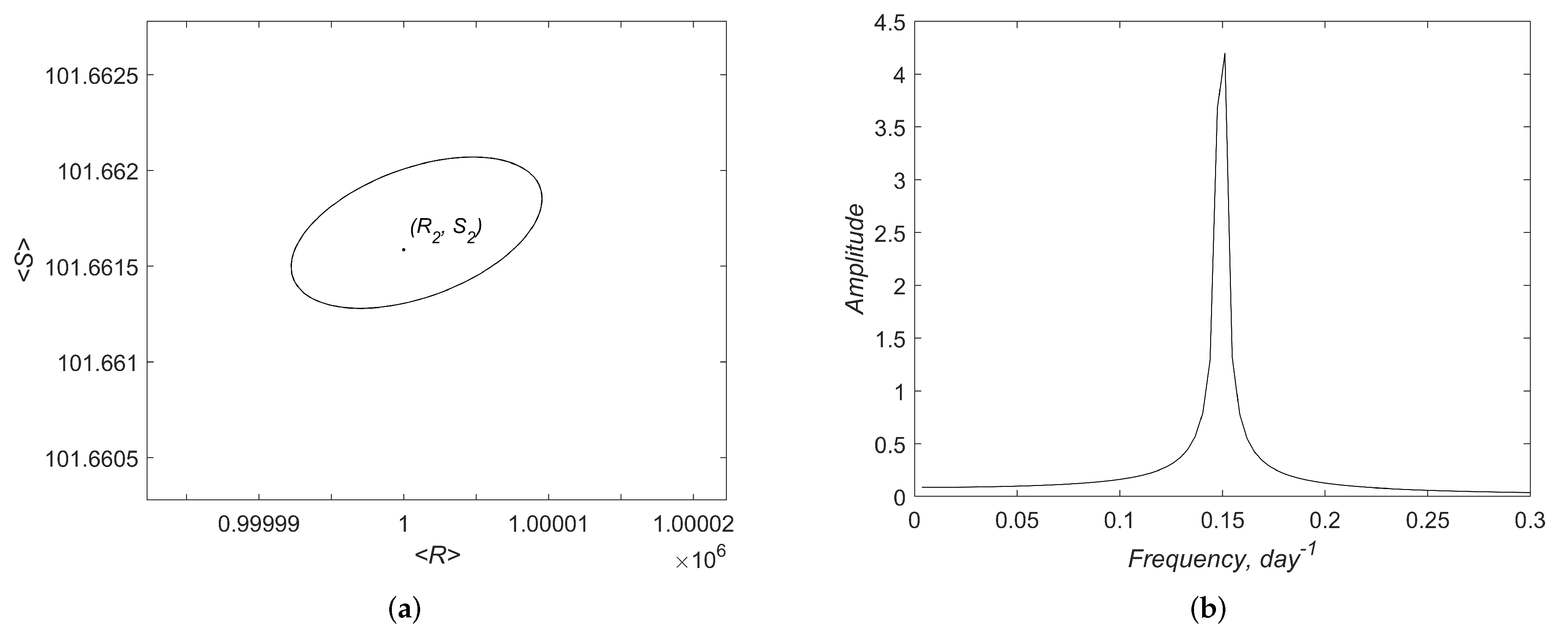
Appendix A. Additional Results of Linear Analysis and Numerical Simulations



References
- Azovsky, A.I.; Chertoprud, E.S. Spatio-temporal dynamics of the White Sea littoral Harpacticoid community. Oceanology 2003, 43, 103–111. [Google Scholar] [CrossRef]
- Fleecer, J.W.; Palmer, M.A.; Moser, E.B. On the scale of aggregation of meio-bentic copepodes on a tidal mudflat. Mar. Ecol. 1990, 11, 227–237. [Google Scholar] [CrossRef]
- Azovsky, A.; Chertoprood, E.; Saburova, M.; Polikarpov, G. Selective feeding of littoral harpacticoids on diatom algae: Hungry gourmands? Mar. Biol. 2005, 148, 327–337. [Google Scholar] [CrossRef]
- Sach, G.; Bernem, H. Spatial patterns of Harpacticoida copepods on tidal flats. Senchenberg. Mar. 1996, 26, 97–106. [Google Scholar]
- Sun, B.; Fleeger, J.W. Spatial and temporal patterns of dispersion in meiobenthic copepods. Mar. Ecol. Prog. Ser. 1991, 71, 1–11. [Google Scholar] [CrossRef]
- Woods, D.R.; Tietjen, J.H. Horizontal and vertical distribution of meiofauna in the Venezuela Basin. Mar. Geol. 1985, 68, 233–241. [Google Scholar] [CrossRef]
- Smirnova, E.A.; Azovsky, A.I. Modelling of spatially distributed predator-prey system with periodically migrating predator (Case study of the White Sea inertidal harpacticoids and benthic microalgae). Oceanology 2020, 60, 89–97. [Google Scholar] [CrossRef]
- Tyutyunov, Y.V.; Zagrebneva, A.D.; Surkov, F.A.; Azovsky, A.I. Microscale patchiness of the distribution of copepods (Harpacticoida) as a result of trophotaxis. Biophysics 2009, 54, 355–360. [Google Scholar] [CrossRef]
- Keller, E.F.; Segel, L.A. Model for Chemotaxis. J. Theor. Biol. 1971, 30, 225–234. [Google Scholar] [CrossRef]
- Patlak, C.S. Random walk with persistence and external bias. Bull. Math. Biophys. 1953, 15, 311–338. [Google Scholar] [CrossRef]
- Tyutyunov, Y.V.; Zagrebneva, A.D.; Surkov, F.A.; Azovsky, A.I. Derivation of density flux equation for intermittently migrating population. Oceanology 2010, 50, 67–76. [Google Scholar] [CrossRef]
- Painter, K.J. Mathematical models for chemotaxis and their applications in self-organisation phenomena. J. Theor. Biol. 2019, 481, 162–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeschke, J.M.; Kopp, M.; Tollrian, R. Consumer-food systems: Why type I functional responses are exclusive to filter feeders. Biol. Rev. 2004, 79, 337–349. [Google Scholar] [CrossRef] [PubMed]
- De Troch, M.; Grego, M.; Chepurnov, V.A.; Vincx, M. Food patch size, food concentration and grazing efficiency of the harpacticoid Paramphiascella fulvofasciata (Crustacea, Copepoda). J. Exp. Mar. Biol. Ecol. 2007, 343, 210–216. [Google Scholar] [CrossRef]
- Leising, A.W.; Franks, P.J. Does Acartia clausi use an area-restricted search foraging strategy to find food. Hydrobiologia 2002, 480, 193–207. [Google Scholar] [CrossRef]
- Kern, J.C. Active and passive aspects of meiobenthic copepod dispersal at two sites near Mustang Island, Texas. Mar. Ecol. Prog. Ser. 1990, 60, 211–223. [Google Scholar] [CrossRef]
- Morton, K.W.; Mayers, D.F. Numerical Solution of Partial Differential Equations; Cambridge University Press: Cambridge, UK, 1994; 227p. [Google Scholar] [CrossRef]
- Samarsky, A.; Gulin, A. Numerical Methods; Publishing House Nauka: Moscow, Russia, 1989; 430p. (In Russian) [Google Scholar]
- Zagrebneva, A.; Tyutyunov, Y.; Surkov, F.; Azovsky, A. Numerical realization of taxis-reaction-diffusion model describing population dynamics in predator-prey system. IZvestiya Vuzov. Sev. Kavk. Reg. Nat. Sci. Ser. 2010, 2, 12–16. (In Russian) [Google Scholar]
- Kahaner, D.; Moler, C.; Nash, S. Numerical Methods and Software; Prentice-Hall Inc.: Upper Saddle River, NJ, USA, 1989; 495p. [Google Scholar]
- Moon, F. Chaotic and Fractal Dynamics: Introduction for Applied Scientists and Engineers; Wiley-VCH: Weinheim, Germany, 1992; 508p. [Google Scholar]
- Arditi, R.; Tyutyunov, Y.; Morgulis, A.; Govorukhin, V.; Senina, I. Directed movement of predators and the emergence of density-dependence in predator–prey models. Theor. Popul. Biol. 2001, 59, 207–221. [Google Scholar] [CrossRef] [Green Version]
- Govorukhin, V.; Morgulis, A.; Tyutyunov, Y. Slow taxis in a predator–prey model. Dokl. Math. 2000, 61, 420–422. [Google Scholar]
- Morgulis, A.; Ilin, K. A remark on the disorienting of species due to the fluctuating environment. arXiv 2019, arXiv:1808.02091v4. [Google Scholar]
- Rai, V.; Upadhyay, R.K.; Thakur, N.K. Complex population dynamics in heterogeneous environments: Effects of random and directed animal movements. Int. J. Nonlin. Sci. Num. 2012, 13, 299–309. [Google Scholar] [CrossRef]
- Tello, J.I.; Wrzosek, D. Predator–prey model with diffusion and indirect prey-taxis. Math. Mod. Meth. Appl. Sci. 2016, 26, 2129–2162. [Google Scholar] [CrossRef]
- Tyutyunov, Y.; Sen, D.; Titova, L.I.; Banerjee, M. Predator overcomes the Allee effect due to indirect prey–taxis. Ecol. Complex. 2019, 39, 100772. [Google Scholar] [CrossRef]
- Tyutyunov, Y.; Titova, L.; Senina, I. Prey–taxis destabilizes homogeneous stationary state in spatial Gause–Kolmogorov-type model for predator–prey system. Ecol. Complex. 2017, 31, 170–180. [Google Scholar] [CrossRef]
- Tyutyunov, Y.; Zagrebneva, A.D.; Govorukhin, V.N.; Titova, L.I. Numerical Study of Bifurcations Occurring at Fast Time-scale in a Predator–prey Model with Inertial Prey–taxis. In Advance Mathematical Method in Biosciences and Applications; Berezovskaya, F., Toni, B., Eds.; STEAM-H Science, Technology, Engineering, Agriculture, Mathematics & Health; Springer: Cham, Switzerland, 2019; pp. 221–239. [Google Scholar] [CrossRef]
- Sapoukhina, N.; Tyutyunov, Y.; Arditi, R. The role of prey taxis in biological control: A spatial theoretical model. Am. Nat. 2003, 162, 61–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Meaning | Dimension Units |
|---|---|---|
| x, y | Spatial coordinates | |
| t | Time | |
| Density of diatom microalgae population | ||
| Density of harpacticoid population | ||
| Satiety of harpacticoid |
| Parameter | Meaning | Dimension Units |
|---|---|---|
| r | Reproduction rate of algal population | |
| K | Carrying capacity of algal population | |
| a | Searching efficiency of individual copepod | |
| Assimilation coefficient of algal cells | – | |
| Digestion rate of algal cells | ||
| Diffusion coefficient of algal population | ||
| Diffusion coefficient of harpacticoid population | ||
| Minimum temporal resolution of the model | min | |
| Mean squared length of individual replacement within | ||
| Frequency of copepod egress into water | ||
| Taxis coefficient of harpacticoid population |
| Variables | Parameters | Initial Conditions |
|---|---|---|
| , | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyutyunov, Y.V.; Zagrebneva, A.D.; Azovsky, A.I. Spatiotemporal Pattern Formation in a Prey-Predator System: The Case Study of Short-Term Interactions Between Diatom Microalgae and Microcrustaceans. Mathematics 2020, 8, 1065. https://doi.org/10.3390/math8071065
Tyutyunov YV, Zagrebneva AD, Azovsky AI. Spatiotemporal Pattern Formation in a Prey-Predator System: The Case Study of Short-Term Interactions Between Diatom Microalgae and Microcrustaceans. Mathematics. 2020; 8(7):1065. https://doi.org/10.3390/math8071065
Chicago/Turabian StyleTyutyunov, Yuri V., Anna D. Zagrebneva, and Andrey I. Azovsky. 2020. "Spatiotemporal Pattern Formation in a Prey-Predator System: The Case Study of Short-Term Interactions Between Diatom Microalgae and Microcrustaceans" Mathematics 8, no. 7: 1065. https://doi.org/10.3390/math8071065