Behavioral Responses of Thrips hawaiiensis (Thysanoptera: Thripidae) to Volatile Compounds Identified from Gardenia jasminoides Ellis (Gentianales: Rubiaceae)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects and Plants
2.2. Behavioural Responses of T. hawaiiensis to Flower Volatiles
2.3. Collection and Analysis of Volatile Organic Compounds (VOCs)
2.4. Behavioural Responses of T. hawaiiensis to the Main G. jasminoides Volatile Organic Compounds (VOCs)
2.5. Odour Stimuli
2.6. Six-arm Olfactometer Bioassays
2.7. Y-Tube Bioassays
2.8. Statistical Analyses
3. Results
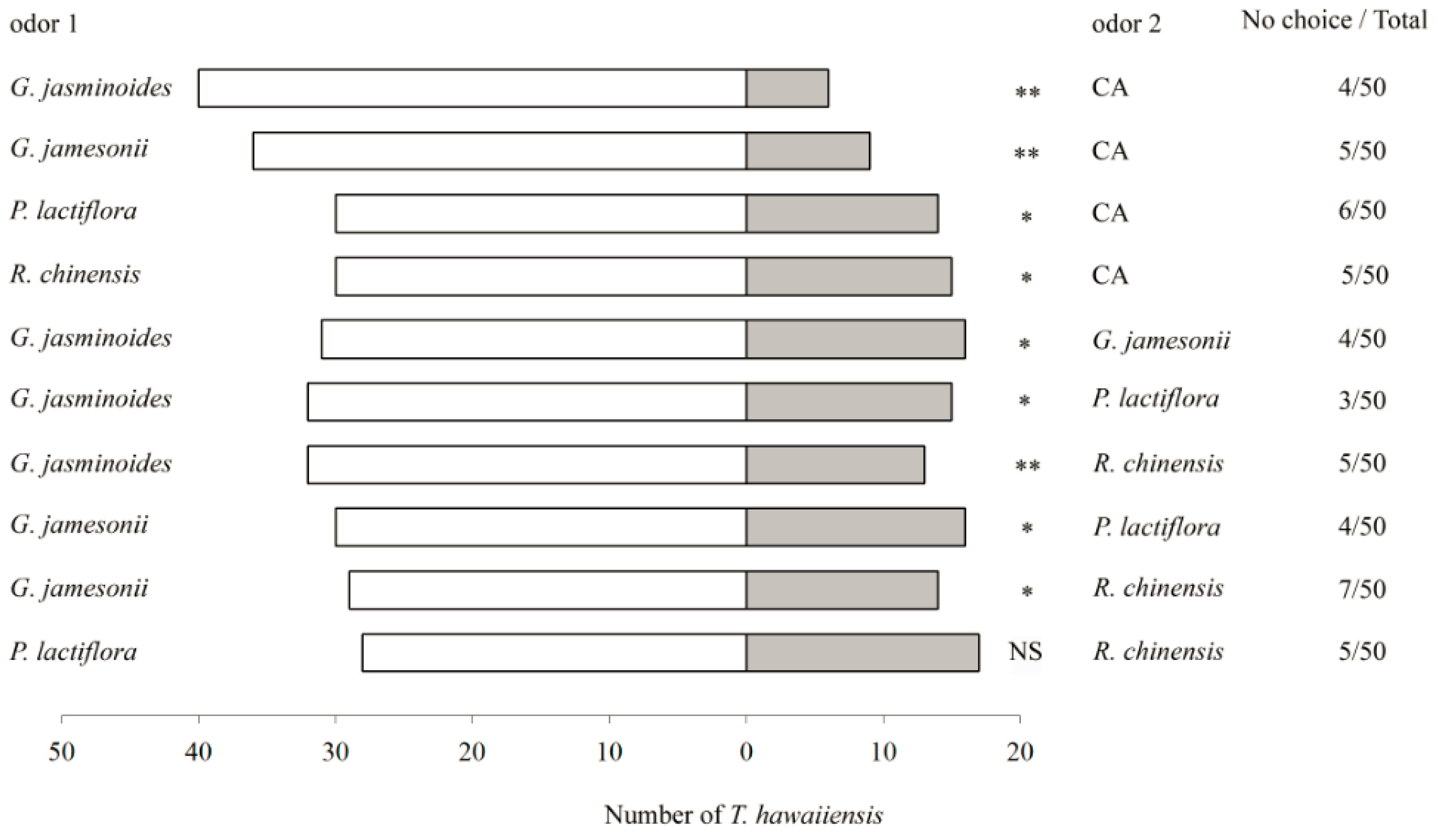
3.1. Behavioural Responses of T. hawaiiensis to Flower Volatiles
3.2. Analysis of G. jasminoides Volatiles
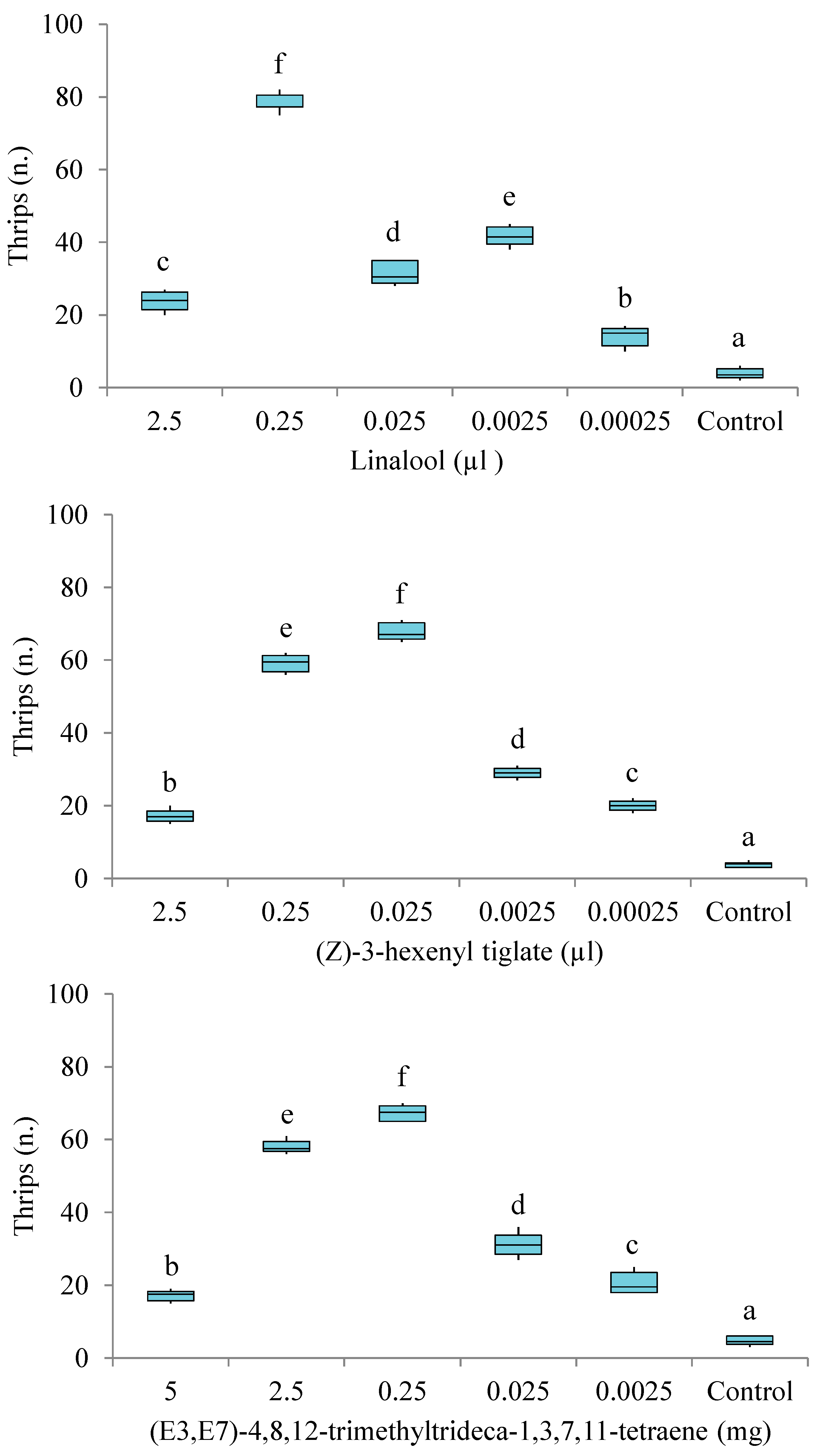
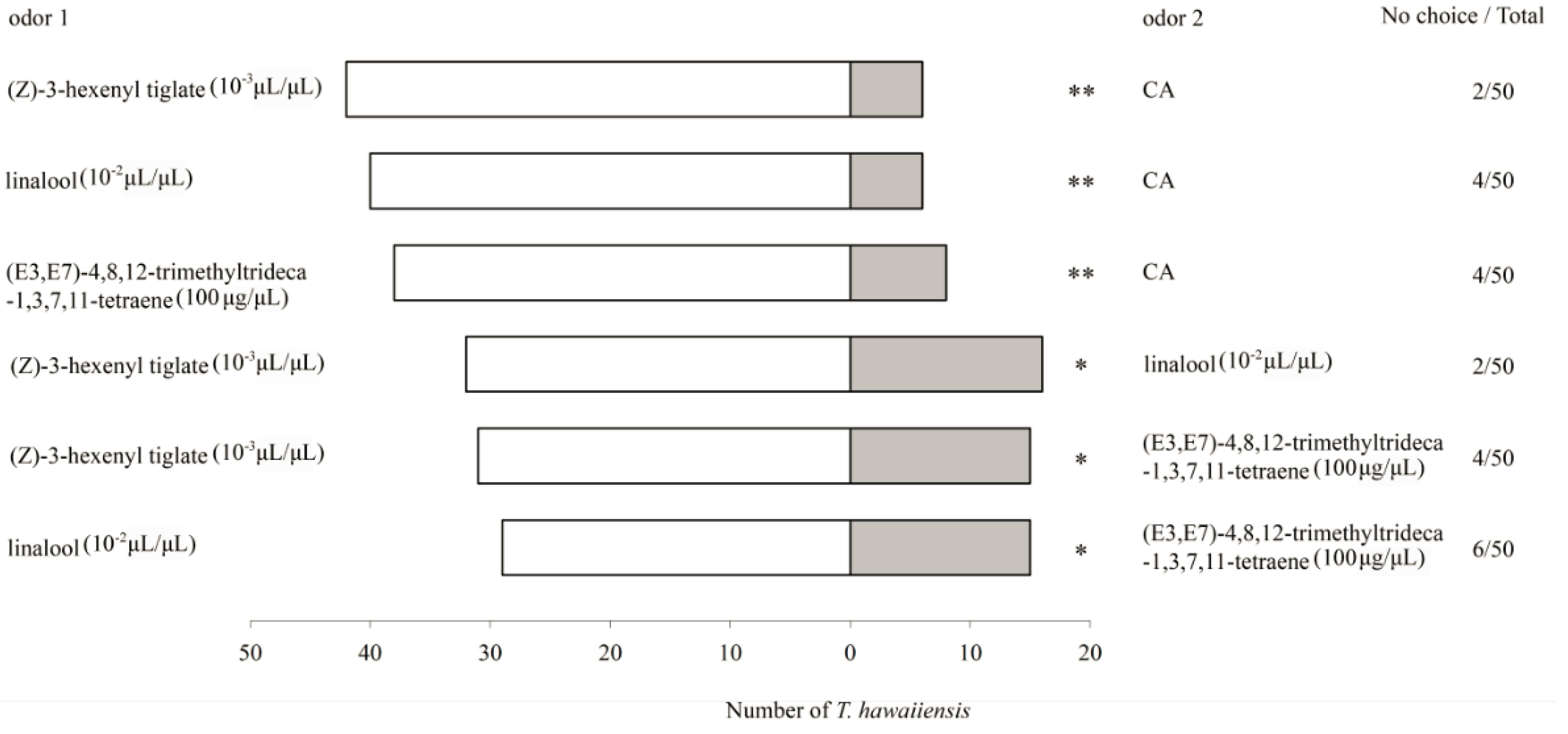
3.3. Behavioural Responses of T. hawaiiensis to the Main Components of G. jasminoides VOCs
3.3.1. Six-Arm Olfactometer Bioassays
3.3.2. Y-Tube Olfactometer Bioassays
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mound, L.A.; Kibby, G. Thysanoptera: An Identification Guide, 2nd ed.; CAB International: Oxon, UK, 1998. [Google Scholar]
- Mound, L.A. Thysanoptera: Diversity and interaction. Annu. Rev. Entomol. 2005, 50, 247–269. [Google Scholar] [CrossRef] [PubMed]
- Murai, T. Development and reproductive capacity of Thrips hawaiiensis (Thysanoptera: Thripidae) and its potential as a major pest. Bull. Entomol. Res. 2001, 91, 193–198. [Google Scholar] [PubMed]
- Reynaud, P.; Balmès, V.; Pizzol, J. Thrips hawaiiensis (Morgan, 1913) (Thysanoptera: Thripidae), an Asian pest thrips now established in Europe. Bull OEPP 2008, 1, 155–160. [Google Scholar] [CrossRef]
- Aliakbarpour, H.; Salmah, M.R.C. Seasonal abundance and spatial distribution of larval and adult thrips (Thysanoptera) on weed host plants in mango orchards in Penang, Malaysia. Appl. Entomol. Zool. 2011, 2, 185–194. [Google Scholar] [CrossRef]
- Goldarazena, A. First record of Thrips hawaiiensis (Morgan, 1913) (Thysanoptera: Thripidae), an Asian pest thrips in Spain. Bull. OEPP 2011, 41, 170–173. [Google Scholar] [CrossRef]
- Atakan, E.; Ölçülü, M.; Pehlivan, S.; Satar, S. A new thrips species recorded in Turkey: Thrips hawaiiensis (Morgan, 1913) (Thysanoptera: Thripidae). Türk. Entomol. Bült. 2015, 2, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Fu, B.L.; Qiu, H.Y.; Li, Q.; Tang, L.D.; Zeng, D.Q.; Liu, K.; Gao, Y.L. Analysis of seasonal and annual field-evolved insecticide resistance in populations of Thrips hawaiiensis in banana orchards. J. Pest Sci. 2019, 92, 1293–1307. [Google Scholar] [CrossRef]
- Fu, B.L.; Tang, L.D.; Qiu, H.Y.; Liu, J.F.; Zeng, D.Q.; Xie, Y.X.; Zeng, D.Q.; Liu, K. Screening of high effect and low toxicity insecticides for controlling Thrips hawaiiensis (Morgan). Chin. J. Fruit Sci. 2016, 33, 257–267. [Google Scholar]
- Fu, B.L.; Li, Q.; Xia, X.Y.; Tang, L.D.; Qiu, H.Y.; Xie, Y.X.; Zeng, D.Q.; Liu, K. Moderate resistance to spinetoram reduces the fitness of Thrips hawaiiensis (Thysanoptera: Thripidae). Acta Entomol. Sin. 2017, 60, 180–188. [Google Scholar]
- Fu, B.L.; Li, Q.; Qiu, H.Y.; Tang, L.D.; Zeng, D.Q.; Liu, K.; Gao, Y.L. Resistance development, stability, cross-resistance potential, biological fitness and biochemical mechanisms of spinetoram resistance in the Thrips hawaiiensis (Thysanoptera: Thripidae). Pest Manag. Sci. 2018, 74, 1564–1574. [Google Scholar] [CrossRef]
- Terry, I. Host Selection Communication and Reproductive Behaviour. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: Oxon, UK, 1997; pp. 65–118. [Google Scholar]
- Teulon, D.A.J.; Hollister, B.; Butler, R.C.; Cameron, E.A. Colour and odour responses of flying western flower thrips: Wind tunnel and greenhouse experiments. Entomol. Exp. Appl. 1999, 93, 9–19. [Google Scholar] [CrossRef]
- Koschier, E.H.; De Kogel, W.J.; Visser, J.H. Assessing the attractiveness of volatile plant compounds to western flower thrips (Frankliniella occidentalis (Pergande). J. Chem. Ecol. 2000, 26, 2643–2655. [Google Scholar] [CrossRef]
- Cao, Y.; Zhi, J.R.; Li, C.; Zhang, R.Z.; Wang, C.; Shang, B.Z.; Gao, Y.L. Behavioral responses of Frankliniella occidentalis to floral volatiles combined with different background visual cues. Arthropod Plant Interact. 2018, 12, 31–39. [Google Scholar] [CrossRef]
- Cao, Y.; Li, C.; Yang, H.; Li, J.; Li, S.; Wang, Y.W.; Gao, Y.L. Laboratory and field investigation on the orientation of Frankliniella occidentalis (Thysanoptera: Thripidae) to more suitable host plants driven by volatiles and component analysis of volatiles. Pest Manag. Sci. 2019, 75, 598–606. [Google Scholar] [CrossRef]
- Mainali, B.P.; Lim, U.T. Behavioral response of western flower thrips to visual and olfactory cues. J. Insect. Behav. 2011, 24, 436–446. [Google Scholar] [CrossRef]
- Abdullah, Z.S.; Ficken, K.J.; Greenfield, B.P.J.; Butt, T.M. Innate responses to putative ancestral hosts: Is the attraction of western flower thrips to pine pollen a result of relict olfactory receptors? J. Chem. Ecol. 2014, 40, 534–540. [Google Scholar] [CrossRef] [Green Version]
- Avellaneda, J.; Marco, D.; Ericsson, C.B.; Daniel, R.; Coralia, O. Rose volatile compounds allow the design of new control strategies for the western flower thrips (Frankliniella occidentalis). J. Pest Sci. 2019, 1–14. [Google Scholar] [CrossRef]
- Zhang, F.; Fu, B.L.; Liu, K.; Qiu, H.Y.; Wu, W. The effect of temperature on the development and survival of Thrips hawaiiensis (Morgan). Acta Ecol. Sin. 2014, 14, 3895–3899. [Google Scholar]
- Cao, Y.; Zhi, J.R.; Zhang, R.Z.; Li, C.; Liu, Y.; Lv, Z.Y.; Gao, Y.L. Different population performances of Frankliniella occidentalis and Thrips hawaiiensis on flowers of two horticultural plants. J. Pest. Sci. 2018, 1, 79–91. [Google Scholar] [CrossRef]
- Yuan, C.M.; Zhi, J.R.; Li, J.Z.; Zhang, Y. Investigation on the species of thrips in fields of vegetable in Guizhou Province. China Plant Prot. 2008, 7, 8–10. [Google Scholar]
- Cao, Y.; Zhi, J.R.; Cong, C.L.; Margolies, D.C. Olfactory cues used in host selection by Frankliniella occidentalis (Thysanoptera: Thripidae) in relation to host suitability. J. Insect Behav. 2014, 27, 41–56. [Google Scholar] [CrossRef]
- Liu, X.F.; Chen, H.H.; Li, J.K.; Zhang, R.; Turlings, T.C.; Chen, L. Volatiles released by Chinese liquorice roots mediate host location behaviour by neonate Porphyrophora sophorae (Hemiptera: Margarodidae). Pest Manag Sci. 2016, 72, 1959–1964. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Benelli, G.; Germinara, G.S.; Maggi, F.; Zhang, Y.J.; Luo, S.L.; Yang, H.; Li, C. Innate positive chemotaxis to paeonal from highly attractive Chinese medicinal herbs in the cigarette beetle, Lasioderma serricorne. Sci Rep. 2019, 6995, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tianyi, K.; Yao, J.P.; Wang, R.H.; Lin, J.; Zhou, Y.; Zuo, G.Q.; Wang, Z.B.; Dai, L.L.; Ren, J.L.; et al. Liquid-solid phase-inversion plga implant for the treatment of residual tumor tissue after hifu ablation. PLoS ONE 2015, 10, e0117358. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Engelwood Cliffs, NJ, USA, 1984. [Google Scholar]
- Tryfonidis, M.; Charalambous, C.P.; Mills, Y.S.P.; Jass, Z.G.K.; Jacob, S.; Stanleyx, J.K.; Haytonx, M.J. Distal radial and ulnar landmarks used in percutaneous pin fixation: Anatomical relationship to the superficial radial and ulnar nerves. Hand Surg. 2010, 15, 161–164. [Google Scholar] [CrossRef]
- Knolhoff, L.M.; Heckel, D.G. Behavioral assays for studies of host plant choice and adaptation in herbivorous insects. Ann. Rev. Entomol. 2014, 59, 263–278. [Google Scholar] [CrossRef]
- Beyaert, I.; Hilker, M. Plant odour plumes as mediators of plant–insect interactions. Biol. Rev. 2014, 89, 68–81. [Google Scholar] [CrossRef]
- Teulon, D.A.J.; Penman, D.R.; Ramakers, P.M.J. Volatile chemical for thrips (Thysanoptera: Thripidae) host-finding and applications for thrips pest management. J. Econ. Entomol. 1993, 86, 1405–1415. [Google Scholar] [CrossRef]
- Frey, J.E.; Cortada, R.V.; Helbling, H. The potential of flower odours for use in population monitoring of western flower thrips, Frankliniella occidentalis (Perg.) (Thysanoptera: Thripidae). Biocontrol. Sci. Technol. 1994, 4, 177–186. [Google Scholar] [CrossRef]
- Davidson, M.M.; Butler, R.C.; Winkler, S.; Tuelon, D.A.J. Pyridine compounds increase trap capture of Frankliniella occidentalis (Pergande) in a covered crop. NZ Plant Prot. 2007, 60, 56–60. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, Z.S.; Greenfield, B.P.; Ficken, K.J.; Taylor, J.W.; Wood, M.; Butt, T.M. A new attractant for monitoring western flower thrips, Frankliniella occidentalis in protected crops. SpringerPlus 2015, 4, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Li, C.; Yang, W.J.; Meng, Y.L.; Wang, L.J.; Shang, B.Z.; Gao, Y.L. Effects of temperature on the development and reproduction of Thrips hawaiiensis (Thysanoptera: Thripidae). J. Econ. Entomol. 2018, 111, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.T.; Xue, S.; Tang, H.Z. Analysis of aroma constituents in Gardenia jasminoides at different flowering stages. Food Sci. 2012, 33, 227–233. [Google Scholar]
- Davidson, M.M.; Perry, N.B.; Larsen, L.; Green, V.C.; Butler, R.C.; Teulon, D.A.J. 4-Pyridyl carbonyl compounds as thrips lures: Effectiveness for western flower thrips in y-tube bioassays. J. Agric. Food Chem. 2008, 56, 6554–6561. [Google Scholar] [CrossRef]
- Najar-Rodriguez, A.J.; Galizia, C.G.; Stierle, J.; Dorn, S. Behavioral and neurophysiological responses of an insect to changing ratios of constituents in host plant-derived volatile mixtures. J. Exp. Biol. 2010, 213, 3388–3397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, D.H.; Linn, C.E.; Teal, P.E.A.; Zhang, A.; Roelofs, W.L.; Loeb, G.M. Eavesdropping on plant volatiles by a specialist moth: Significance of ratio and concentration. PLoS ONE 2011, 6, e17033. [Google Scholar] [CrossRef] [Green Version]
- Hulshof, J.; Ketoja, E.; Vanninen, L. Life history characteristics of Frankliniella occidentalis on cucumber leaves with and without supplemental food. Entomol. Exp. Appl. 2003, 108, 19–32. [Google Scholar] [CrossRef]
- Riley, D.G.; Angelella, G.M.; McPherson, R.M. Pine pollen dehiscence relative to thrips population dynamics. Entomol. Exp. Appl. 2011, 138, 223–233. [Google Scholar] [CrossRef]
- Zhi, J.R.; Fitch, G.K.; Margolies, D.C.; Nechols, J.R. Apple pollen as a supplemental food for the western flower thrips, Frankliniella occidentalis: Response of individuals and populations. Entomol. Exp. Appl. 2005, 117, 185–192. [Google Scholar] [CrossRef]
- Vernon, R.S.; Gillespie, D.R. Influence of trap shape, size, and background color on captures of Frankliniella occidentalis (Thysanoptera: Thripidae) in a cucumber greenhouse. J. Econ. Entomol. 1995, 88, 288–293. [Google Scholar] [CrossRef]
- Brown, A.S.S.; Simmonds, M.S.J.; Blaney, W.M. Relationship between nutritional composition of plant species and infestation levels of thrips. J. Chem. Ecol. 2003, 12, 2399–2409. [Google Scholar]
- Mainali, B.P.; Lim, U.T. Circular yellow sticky trap with black background enhances attraction of Frankliniella occidentalis (pergande) (Thysanoptera: Thripidae). Appl. Entomol. Zool. 2010, 45, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Dicke, M. Are herbivore-induced plant volatiles reliable indicators of herbivore identity to foraging carnivorous arthropods? Entomol. Exp. Appl. 1999, 92, 131–142. [Google Scholar] [CrossRef]
- Zhong, F.; He, Y.R.; Gao, Y.; Qi, G.J.; Zhao, C.Y.; Lu, L.H. Olfactory responses of Neoseiulus cucumeris (Acari: Phytoseiidae) to odors of host plants and Frankliniella occidentalis (Thysanoptera: Thripidae)–plant complexes. Arthropod Plant Interact. 2011, 5, 307–314. [Google Scholar] [CrossRef]
- Joo, Y.; Schuman, M.C.; Goldberg, J.K.; Sang-Gyu, K.; Yon, F.; Brütting, C.; Baldwin, I.T.; Rasmann, S. Herbivore-induced volatile blends with both “fast” and “slow” components provide robust indirect defence in nature. Funct. Ecol. 2018, 32, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Takemoto, S.; Shimoda, T. Olfactory responses of the predatory mites (Neoseiulus cucumeris) and insects (Orius strigicollis) to two different plant species infested with onion thrips (Thrips tabaci). J. Chem. Ecol. 2008, 34, 605–613. [Google Scholar] [CrossRef]
- Turlings, T.C.J.; Bernasconi, M.; Bertossa, R.; Bigler, F.; Caloz, G.; Dorn, S. The induction of volatile emissions in maize by three herbivore species with different feeding habits: Possible consequences for their natural enemies. Biol. Control. 1998, 11, 122–129. [Google Scholar] [CrossRef]
- Mattiacci, L.; Dicke, M. Host-age discrimination during host location by Cotesia glomerata, a larval parasitoid of Pieris brassicae. Entomol. Exp. Appl. 1995, 76, 37–48. [Google Scholar] [CrossRef]
- Röse, U.S.R.; Lewis, W.J.; Tumlinson, J.H. Specificity of systemically released cotton volatiles as attractants for specialist and generalist parasitic wasps. J. Chem. Ecol. 1998, 24, 303–319. [Google Scholar] [CrossRef]
- Germinara, G.S.; De Cristofaro, A.; Rotundo, G. Electrophysiological and behavioral responses of Theocolax elegans (Westwood) (Hymenoptera: Pteromalidae) to cereal grain volatiles. BioMed Res. Int. 2016, 2016, 5460819. [Google Scholar] [CrossRef] [Green Version]
- Fu, B.L.; Qiu, H.Y.; Li, Q.; Sun, Y.T.; Zhou, S.H.; Yang, S.Y.; LI, S.G.; Tang, L.D.; Zhang, F.P.; Liu, K. Predation of Orius sauteri on Thrips hawaiiensis in the laboratory. Chin. J. Appl. Entomol. 2019, 56, 91–98. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Number | Compounds | Molecular Formula | Molecular Weight | Content (%) |
|---|---|---|---|---|
| 1 | Methyl tiglate | C6H10O2 | 114 | 0.39 |
| 2 | (Z)-3-Hexen-1-ol | C6H12O | 100 | 3.59 |
| 3 | β-Myrcene | C10H16 | 136 | 0.15 |
| 4 | (Z)-3-Hexenyl acetate | C8H14O2 | 142 | 0.18 |
| 5 | (E)-Ocimene | C10H16 | 136 | 1.38 |
| 6 | γ-Caprolactone | C6H10O2 | 114 | 0.10 |
| 7 | trans-Linalool oxide | C10H18O2 | 170 | 0.32 |
| 8 | Methyl benzoate | C8H8O2 | 136 | 1.64 |
| 9 | Linalool | C10H18O | 154 | 27.45 |
| 10 | (Z)-3-hexenyl iso-butyrate | C10H18O2 | 134 | 0.22 |
| 11 | Benzyl acetate | C9H10O2 | 150 | 0.25 |
| 12 | (Z)-3-hexenyl butanoate | C10H18O2 | 170 | 0.38 |
| 13 | Methyl salicylate | C8H8O3 | 152 | 0.20 |
| 14 | (Z)-3-hexenyl 2-methylbutanoate | C11H20O2 | 184 | 1.28 |
| 15 | Hexyl 2-Methylbutyrate | C11H22O2 | 186 | 0.61 |
| 16 | Benzyl propionate | C10H12O2 | 164 | 0.12 |
| 17 | (Z)-3-hexenyl tiglate | C11H18O2 | 182 | 14.38 |
| 18 | Hexyl tiglate | C11H20O2 | 184 | 4.01 |
| 19 | (Z)-3-hexenyl hexanoate | C12H22O2 | 198 | 0.11 |
| 20 | Isoamyl benzoate | C12H16O2 | 192 | 0.28 |
| 21 | γ-Decanolactone | C10H18O2 | 170 | 0.18 |
| 22 | Jasmine lactone | C10H16O2 | 168 | 6.93 |
| 23 | Benzyl tiglate | C12H14O2 | 190 | 0.23 |
| 24 | α-Farnesene | C15H24 | 204 | 3.58 |
| 25 | Octyl (E)-2-methylbut-2-enoate | C13H24O2 | 212 | 0.58 |
| 26 | E-Nerolidol | C15H26O | 222 | 0.27 |
| 27 | (Z)-3-Hexenyl phenylacetate | C14H18O2 | 218 | 0.41 |
| 28 | (E3,E7)-4,8,12-trimethyltrideca-1,3,7,11-tetraene | C16H26 | 218 | 24.67 |
| 29 | Geranyl tiglate | C15H24O2 | 236 | 0.41 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Wang, J.; Germinara, G.S.; Wang, L.; Yang, H.; Gao, Y.; Li, C. Behavioral Responses of Thrips hawaiiensis (Thysanoptera: Thripidae) to Volatile Compounds Identified from Gardenia jasminoides Ellis (Gentianales: Rubiaceae). Insects 2020, 11, 408. https://doi.org/10.3390/insects11070408
Cao Y, Wang J, Germinara GS, Wang L, Yang H, Gao Y, Li C. Behavioral Responses of Thrips hawaiiensis (Thysanoptera: Thripidae) to Volatile Compounds Identified from Gardenia jasminoides Ellis (Gentianales: Rubiaceae). Insects. 2020; 11(7):408. https://doi.org/10.3390/insects11070408
Chicago/Turabian StyleCao, Yu, Jie Wang, Giacinto Salvatore Germinara, Lijuan Wang, Hong Yang, Yulin Gao, and Can Li. 2020. "Behavioral Responses of Thrips hawaiiensis (Thysanoptera: Thripidae) to Volatile Compounds Identified from Gardenia jasminoides Ellis (Gentianales: Rubiaceae)" Insects 11, no. 7: 408. https://doi.org/10.3390/insects11070408