High-Value Compounds in Fruit, Vegetable and Cereal Byproducts: An Overview of Potential Sustainable Reuse and Exploitation

 , and
, and
Abstract
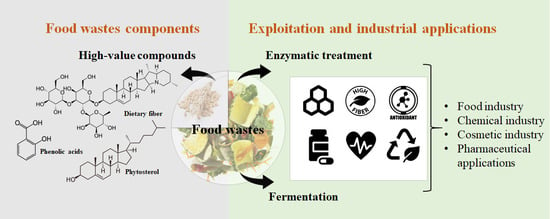
:
1. Introduction
2. High-Value Compounds (HVC) in Plant Byproducts
2.1. Phenolic Compounds
2.2. Carotenoids
2.3. Phytosterols
2.4. Phytic Acid
2.5. Dietary Fibers
3. Main Plant Byproducts
3.1. Apple Byproducts
3.2. Avocado Byproducts
3.3. Banana Byproducts
3.4. Pomegranate Byproducts
3.5. Date Byproducts
3.6. Mango Byproducts
3.7. Citrus Byproducts
3.8. Tomato Byproducts
3.9. Carrot Byproducts
3.10. Cauliflower Byproducts
3.11. Wheat Byproducts
3.12. Rice Byproducts
3.13. Corn Byproducts
3.14. Barley Byproducts
4. Exploitation of Plant Byproducts through Enzymatic Treatment and Fermentation
4.1. Enzymatic Treatment
4.1.1. Enzymatic Treatments of Fruit/Vegetable Byproducts
4.1.2. Enzymatic Treatment of Cereal Byproducts
4.2. Microbial Fermentation
4.2.1. Fermented Fruit Byproducts
4.2.2. Fermented Vegetable Byproducts
4.2.3. Fermented Cereal Byproducts
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chen, H.; Jiang, W.; Yang, Y.; Yang, Y.; Man, X. State of the art on food waste research: A bibliometrics study from 1997 to 2014. J. Clean. Prod. 2017, 140, 840–846. [Google Scholar] [CrossRef]
- Gustavsson, J.; Cederberg, C.; Sonesson, U. Global Food Losses and Food Waste: Extent, Causes and Prevention; study conducted for the International Congress Save Food! at Interpack 2011, [16–17 May], Düsseldorf, Germany; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011; ISBN 978-92-5-107205-9. [Google Scholar]
- Cecilia, J.A.; García-Sancho, C.; Maireles-Torres, P.J.; Luque, R. Industrial Food Waste Valorization: A General Overview. In Biorefinery; Springer Science and Business Media LLC: Berlin, Germany, 2019; pp. 253–277. [Google Scholar]
- Saqib, N.U.; Sharma, H.B.; Baroutian, S.; Dubey, B.; Sarmah, A.K. Valorisation of food waste via hydrothermal carbonisation and techno-economic feasibility assessment. Sci. Total. Environ. 2019, 690, 261–276. [Google Scholar] [CrossRef] [PubMed]
- European Union. Directive 2008/98/EC of the European Parliament and of the Council of 19 November 2008 on Waste and Repealing Certain Directives (Text with EEA Relevance); 2008; Volume OJL 312, pp. 3–30. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32008L0098 (accessed on 28 May 2020).
- European Union. Commission Directive 2008/38/EC of 5 March 2008 Establishing a List of Intended Uses of Animal Feedingstuffs for Particular Nutritional Purposes (Codified Version) (Text with EEA Relevance); 2008; Volume OJL 62, pp. 9–22. Available online: https://www.eumonitor.eu/9353000/1/j9vvik7m1c3gyxp/vi8rm2zu9oyx (accessed on 28 May 2020).
- Coman, V.; Teleky, B.-E.; Mitrea, L.; Martău, G.A.; Szabo, K.; Călinoiu, L.-F.; Vodnar, D.C. Chapter Five—Bioactive potential of fruit and vegetable wastes. In Advances in Food and Nutrition Research; Toldrá, F., Ed.; Academic Press: Amsterdam, The Netherland, 2020; Volume 91, pp. 157–225. [Google Scholar]
- Trigo, J.P.; Alexandre, E.M.C.; Saraiva, J.A.; Pintado, M.E. High value-added compounds from fruit and vegetable by-products – Characterization, bioactivities, and application in the development of novel food products. Crit. Rev. Food Sci. Nutr. 2019, 60, 1388–1416. [Google Scholar] [CrossRef] [PubMed]
- OECD. Food and Agriculture Organization of the United Nations OECD-FAO Agricultural Outlook 2019–2028; OECD-FAO Agricultural Outlook; OECD: Paris, France, 2019; ISBN 978-92-64-31245-6. [Google Scholar]
- Viuda-Martos, M.; Fernández-López, J.; Pérez-Álvarez, J.A. Valuable Compounds Extraction from Cereal Waste and By-Products. Green Extr. Valorization Prod. Food Process. 2019, 153–186. [Google Scholar] [CrossRef]
- Gunes, R.; Palabiyik, I.; Toker, O.S.; Konar, N.; Kurultay, S. Incorporation of defatted apple seeds in chewing gum system and phloridzin dissolution kinetics. J. Food Eng. 2019, 255, 9–14. [Google Scholar] [CrossRef]
- Călinoiu, L.F.; Mitrea, L.; Precup, G.; Bindea, M.; Rusu, B.; Dulf, F.V.; Ştefănescu, B.E.; Vodnar, D.C. Characterization of Grape and Apple Peel Wastes’ Bioactive Compounds and Their Increased Bioavailability After Exposure to Thermal Process. Bull. Univ. Agric. Sci. Veter-Med. Cluj-Napoca. Food Sci. Technol. 2017, 74, 80. [Google Scholar] [CrossRef] [Green Version]
- Nile, S.H.; Nile, A.; Liu, J.; Kim, D.H.; Kai, G. Exploitation of apple pomace towards extraction of triterpenic acids, antioxidant potential, cytotoxic effects, and inhibition of clinically important enzymes. Food Chem. Toxicol. 2019, 131, 110563. [Google Scholar] [CrossRef]
- Tremocoldi, M.A.; Rosalen, P.L.; Franchin, M.; Massarioli, A.P.; Denny, C.; Daiuto, E.R.; Paschoal, J.A.R.; Melo, P.S.; De Alencar, S.M. Exploration of avocado by-products as natural sources of bioactive compounds. PLoS ONE 2018, 13, e0192577. [Google Scholar] [CrossRef] [Green Version]
- Arun, K.B.; Persia, F.; Aswathy, P.S.; Chandran, J.; Sajeev, M.S.; Jayamurthy, P.; Nisha, P. Plantain peel—A potential source of antioxidant dietary fibre for developing functional cookies. J. Food Sci. Technol. 2015, 52, 6355–6364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandhya, S.; Khamrui, K.; Prasad, W.; Kumar, M.; Kumar, C.M. Preparation of pomegranate peel extract powder and evaluation of its effect on functional properties and shelf life of curd. LWT 2018, 92, 416–421. [Google Scholar] [CrossRef]
- Jridi, M.; Souissi, N.; Ben Salem, M.; Ayadi, M.; Nasri, M.; Azabou, S. Tunisian date (Phoenix dactylifera L.) by-products: Characterization and potential effects on sensory, textural and antioxidant properties of dairy desserts. Food Chem. 2015, 188, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Bernal-Mercado, A.T.; Acevedo-Hernandez, C.; Silva-Espinoza, B.A.; Cruz-Valenzuela, M.R.; Gonzalez-Aguilar, G.A.; Nazzaro, F.; Siddiqui, M.W.; Ayala-Zavala, J.F.; Fratianni, F.; Vazquez-Armenta, F.J. Antioxidant and Antimicrobial Capacity of Phenolic Compounds of Mango (Mangifera indica L.) Seed depending upon the Extraction Process. J. Med. Plants Prod. 2018, 7, 209–219. [Google Scholar] [CrossRef]
- Baddi, J.; Vijayalakshmi, D.; Kapale, M. Extraction of Total Polyphenols and Dietary Fiber from Mango Peel - As Potential Sources of Natural Phytonutrients. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1196–1205. [Google Scholar] [CrossRef]
- Sudha, M.L.; Indumathi, K.; Sumanth, M.S.; Rajarathnam, S.; Shashirekha, M. Mango pulp fibre waste: Characterization and utilization as a bakery product ingredient. J. Food Meas. Charact. 2015, 9, 382–388. [Google Scholar] [CrossRef]
- Inan, Ö.; Özcan, M.M.; Aljuhaimi, F. Effect of location and Citrus species on total phenolic, antioxidant, and radical scavenging activities of some Citrus seed and oils. J. Food Process. Preserv. 2017, 42, e13555. [Google Scholar] [CrossRef]
- Pacheco, M.T.; Moreno, F.J.; Villamiel, M. Chemical and physicochemical characterization of orange by-products derived from industry. J. Sci. Food Agric. 2018, 99, 868–876. [Google Scholar] [CrossRef]
- Nour, V.; Ionica, M.E.; Trandafir, I. Bread enriched in lycopene and other bioactive compounds by addition of dry tomato waste. J. Food Sci. Technol. 2015, 52, 8260–8267. [Google Scholar] [CrossRef] [Green Version]
- Gulsunoglu, Z.; Karbancioglu-Guler, F.; Raes, K.; Kilic-Akyilmaz, M. Soluble and insoluble-bound phenolics and antioxidant activity of various industrial plant wastes. Int. J. Food Prop. 2019, 22, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Zhang, W.; Zhang, R.; Jiao, J.; Fu, C.; Tong, X.; Zhang, W.; Qin, L. Cereal fiber improves blood cholesterol profiles and modulates intestinal cholesterol metabolism in C57BL/6 mice fed a high-fat, high-cholesterol diet. Food Nutr. Res. 2019, 63. [Google Scholar] [CrossRef]
- Lin, Y.; Chen, K.; Tu, D.; Yu, X.; Dai, Z.; Shen, Q. Characterization of dietary fiber from wheat bran (Triticum aestivum L.) and its effect on the digestion of surimi protein. LWT 2019, 102, 106–112. [Google Scholar] [CrossRef]
- Lei, L.; Chen, J.; Liu, Y.; Wang, L.; Zhao, G.; Chen, Z.-Y. Dietary Wheat Bran Oil Is Equally as Effective as Rice Bran Oil in Reducing Plasma Cholesterol. J. Agric. Food Chem. 2018, 66, 2765–2774. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.; Madl, R.L.; Takemoto, L.J.; Baybutt, R.C.; Wang, W. Lignans Are Involved in the Antitumor Activity of Wheat Bran in Colon Cancer SW480 Cells. J. Nutr. 2005, 135, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Grases, F. Effects of Phytic Acid on Renal Stone Formation in Rats. Scand. J. Urol. Nephrol. 1998, 32, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.G.; Stone, N.J. Antiatherosclerotic and Antithrombotic Effects of Omega-3 Fatty Acids. Am. J. Cardiol. 2006, 98, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Zanzottera, F.; Bizzaro, G.; Michelotti, A.; Nobile, V. Efficacy of a Nutritional Supplement, Standardized in Fatty Acids and Phytosterols, on Hair Loss and Hair Health in both Women and Men. J. Cosmetol. Trichol. 2017, 3, 03. [Google Scholar] [CrossRef]
- Kim, E.O.; Min, K.J.; Kwon, T.K.; Um, B.H.; Moreau, R.; Choi, S.-W. Anti-inflammatory activity of hydroxycinnamic acid derivatives isolated from corn bran in lipopolysaccharide-stimulated Raw 264.7 macrophages. Food Chem. Toxicol. 2012, 50, 1309–1316. [Google Scholar] [CrossRef]
- Lerma-García, M.; Herrero-Martínez, J.M.; Simó-Alfonso, E.F.; Mendonça, C.R.; Ramis-Ramos, G. Composition, industrial processing and applications of rice bran γ-oryzanol. Food Chem. 2009, 115, 389–404. [Google Scholar] [CrossRef]
- Padhi, E.M.; Liu, R.; Hernandez, M.; Tsao, R.; Ramdath, D. Total polyphenol content, carotenoid, tocopherol and fatty acid composition of commonly consumed Canadian pulses and their contribution to antioxidant activity. J. Funct. Foods 2017, 38, 602–611. [Google Scholar] [CrossRef]
- Wu, X.; Kim, E.; Choi, K.; Kim, H. Antioxidative and Physical Characteristics of Rice Germ added Sweet Pumpkin Rice Snack for Infants. J. Korean Soc. Food Cult. 2018, 33, 62–69. [Google Scholar] [CrossRef]
- Giusti, M.; Wrolstad, R.E. Acylated anthocyanins from edible sources and their applications in food systems. Biochem. Eng. J. 2003, 14, 217–225. [Google Scholar] [CrossRef]
- Rose, D.J.; Inglett, G.E.; Liu, S.X. Utilisation of corn (Zea mays ) bran and corn fiber in the production of food components. J. Sci. Food Agric. 2010, 90, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Cheung, N.-K.V.; Modak, S.; Vickers, A.J.; Knuckles, B. Orally administered β-glucans enhance anti-tumor effects of monoclonal antibodies. Cancer Immunol. Immunother. 2002, 51, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Nocente, F.; Taddei, F.; Galassi, E.; Gazza, L. Upcycling of brewers’ spent grain by production of dry pasta with higher nutritional potential. LWT 2019, 114, 108421. [Google Scholar] [CrossRef]
- Babbar, N.; Oberoi, H.S.; Sandhu, S.K. Therapeutic and Nutraceutical Potential of Bioactive Compounds Extracted from Fruit Residues. Crit. Rev. Food Sci. Nutr. 2014, 55, 319–337. [Google Scholar] [CrossRef]
- Akhtar, S.; Ismail, T.; Fraternale, D.; Sestili, P. Pomegranate peel and peel extracts: Chemistry and food features. Food Chem. 2015, 174, 417–425. [Google Scholar] [CrossRef]
- Maiani, G.; Castón, M.J.P.; Catasta, G.; Toti, E.; Cambrodón, I.G.; Bysted, A.; Granado-Lorencio, F.; Olmedilla-Alonso, B.; Knuthsen, P.; Valoti, M.; et al. Carotenoids: Actual knowledge on food sources, intakes, stability and bioavailability and their protective role in humans. Mol. Nutr. Food Res. 2008, 53, S194–S218. [Google Scholar] [CrossRef]
- Moreau, R.; Nyström, L.; Whitaker, B.D.; Winkler-Moser, J.K.; Baer, D.J.; Gebauer, S.K.; Hicks, K.B. Phytosterols and their derivatives: Structural diversity, distribution, metabolism, analysis, and health-promoting uses. Prog. Lipid Res. 2018, 70, 35–61. [Google Scholar] [CrossRef]
- Kornsteiner-Krenn, M.; Wagner, K.-H.; Elmadfa, I. Phytosterol Content and Fatty Acid Pattern of Ten Different Nut Types. Int. J. Vitam. Nutr. Res. 2013, 83, 263–270. [Google Scholar] [CrossRef]
- Crews, C.; Hough, P.; Godward, J.; Brereton, P.; Lees, M.; Guiet, S.; Winkelmann, W. Study of the Main Constituents of Some Authentic Hazelnut Oils. J. Agric. Food Chem. 2005, 53, 4843–4852. [Google Scholar] [CrossRef]
- Osuntokun, O.T.; Oluduro, A.; Idowu, T.; Omotuyi, A. Assessment of Nephrotoxicity, Anti-Inflammatory and Antioxidant Properties of Epigallocatechin, Epicatechin and Stigmasterol Phytosterol (Synergy) Derived from Ethyl Acetate Stem Bark Extract of Spondias Mombin on Wistar Rats Using Molecular Method of Analysis. J. Mol. Microbiol. 2017, 11. [Google Scholar]
- Silva, E.O.; Bracarense, A.P.F. Phytic Acid: From Antinutritional to Multiple Protection Factor of Organic Systems. J. Food Sci. 2016, 81, R1357–R1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galanakis, C.M. Dietary Fiber: Properties, Recovery, and Applications; Academic Press: London, UK, 2019; ISBN 978-0-12-816496-9. [Google Scholar]
- Alba, K.; Campbell, G.; Kontogiorgos, V. Dietary fibre from berry-processing waste and its impact on bread structure: A review. J. Sci. Food Agric. 2019, 99, 4189–4199. [Google Scholar] [CrossRef] [Green Version]
- Products, N.E.P.O.D.; Nutrition, E.P.O.D.P.; (Nda), A. Scientific Opinion on principles for deriving and applying Dietary Reference Values. EFSA J. 2010, 8, 1458. [Google Scholar] [CrossRef] [Green Version]
- Waldbauer, K.; McKinnon, R.; Kopp, B. Apple Pomace as Potential Source of Natural Active Compounds. Planta Medica 2017, 83, 994–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkhalaf, M.I.; Alansari, W.S.; Ibrahim, E.A.; Elhalwagy, M.E. Anti-oxidant, anti-inflammatory and anti-cancer activities of avocado (Persea americana) fruit and seed extract. J. King Saud Univ. Sci. 2019, 31, 1358–1362. [Google Scholar] [CrossRef]
- Ismail, T.; Akhtar, S.; Riaz, M.; Hameed, A.; Afzal, K.; Sheikh, A.S. Oxidative and Microbial Stability of Pomegranate Peel Extracts and Bagasse Supplemented Cookies. J. Food Qual. 2016, 39, 658–668. [Google Scholar] [CrossRef]
- Xu, Y.; Li, Y.; Bao, T.; Zheng, X.; Chen, W.; Wang, J. A recyclable protein resource derived from cauliflower by-products: Potential biological activities of protein hydrolysates. Food Chem. 2017, 221, 114–122. [Google Scholar] [CrossRef]
- Belc, N.; Mustatea, G.; Apostol, L.; Iorga, S.; Vladut, V.; Mosoiu, C. Cereal supply chain waste in the context of circular economy. E3S Web Conf. 2019, 112, 03031. [Google Scholar] [CrossRef]
- Deroover, L.; Tie, Y.; Verspreet, J.; Courtin, C.; Verbeke, K. Modifying wheat bran to improve its health benefits. Crit. Rev. Food Sci. Nutr. 2019, 60, 1104–1122. [Google Scholar] [CrossRef]
- Apprich, S.; Tirpanalan, Ö.; Hell, J.; Reisinger, M.; Böhmdorfer, S.; Siebenhandl-Ehn, S.; Novalin, S.; Kneifel, W. Wheat bran-based biorefinery 2: Valorization of products. LWT 2014, 56, 222–231. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Cassone, A.; Coda, R.; Gobbetti, M. Antifungal activity of sourdough fermented wheat germ used as an ingredient for bread making. Food Chem. 2011, 127, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Al-Doury, M.K.W.; Hettiararchchy, N.S.; Horax, R. Rice-Endosperm and Rice-Bran Proteins: A Review. J. Am. Oil Chem. Soc. 2018, 95, 943–956. [Google Scholar] [CrossRef]
- Perez-Ternero, C.; De Sotomayor, M.A.; Herrera, M.D. Contribution of ferulic acid, γ-oryzanol and tocotrienols to the cardiometabolic protective effects of rice bran. J. Funct. Foods 2017, 32, 58–71. [Google Scholar] [CrossRef]
- Luna-Vital, D.; Li, Q.; West, L.; West, M.; De Mejia, E.G. Anthocyanin condensed forms do not affect color or chemical stability of purple corn pericarp extracts stored under different pHs. Food Chem. 2017, 232, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.; Motilva, M.-J.; Hazas, M.-C.L.D.L.; Romero, M.-P.; Vaculova, K.; Ludwig, I.A. Phytochemical composition and β-glucan content of barley genotypes from two different geographic origins for human health food production. Food Chem. 2018, 245, 61–70. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to beta-glucans from oats and barley and maintenance of normal blood LDL-cholesterol concentrations (ID 1236, 1299), increase in satiety leading to a reduction in energy intake (ID 851, 852), reduction of post-prandial glycaemic responses (ID 821, 824), and “digestive function”(ID 850) pursuant to Article 13 (1) of Regulation (EC) No 1924/2006. EFSA 2011, 9, 2207. [Google Scholar]
- Karimi, R.; Azizi, M.H.; Xu, Q.; Sahari, M.A.; Hamidi, Z.; Naghizadeh-Raeisi, S. Enzymatic removal of starch and protein during the extraction of dietary fiber from barley bran. J. Cereal Sci. 2018, 83, 259–265. [Google Scholar] [CrossRef]
- Treimo, J.; Westereng, B.; Horn, S.J.; Forssell, P.; Robertson, J.A.; Faulds, C.B.; Waldron, K.W.; Buchert, J.; Eijsink, V.G.H. Enzymatic Solubilization of Brewers’ Spent Grain by Combined Action of Carbohydrases and Peptidases. J. Agric. Food Chem. 2009, 57, 3316–3324. [Google Scholar] [CrossRef]
- Nigam, P.S. An overview: Recycling of solid barley waste generated as a by-product in distillery and brewery. Waste Manag. 2017, 62, 255–261. [Google Scholar] [CrossRef]
- Puri, M.; Sharma, D.; Barrow, C.J. Enzyme-assisted extraction of bioactives from plants. Trends Biotechnol. 2012, 30, 37–44. [Google Scholar] [CrossRef]
- Catalkaya, G.; Kahveci, D. Optimization of enzyme assisted extraction of lycopene from industrial tomato waste. Sep. Purif. Technol. 2019, 219, 55–63. [Google Scholar] [CrossRef]
- Fernandez, K.; Vega, M.; Aspé, E. An enzymatic extraction of proanthocyanidins from País grape seeds and skins. Food Chem. 2015, 168, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Štambuk, P.; Tomašković, D.; Tomaz, I.; Maslov, L.; Stupić, D.; Kontić, J.K. Application of pectinases for recovery of grape seeds phenolics. 3 Biotech. 2016, 6, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meini, M.-R.; Cabezudo, I.; Boschetti, C.E.; Romanini, D. Recovery of phenolic antioxidants from Syrah grape pomace through the optimization of an enzymatic extraction process. Food Chem. 2019, 283, 257–264. [Google Scholar] [CrossRef]
- Ruviaro, A.; Barbosa, P.D.P.M.; Macedo, G.A. Enzyme-assisted biotransformation increases hesperetin content in citrus juice by-products. Food Res. Int. 2018, 124, 213–221. [Google Scholar] [CrossRef]
- Mandalari, G.; Bennett, R.N.; Kirby, A.R.; Curto, R.B.L.; Bisignano, G.; Waldron, K.W.; Faulds, C.B. Enzymatic Hydrolysis of Flavonoids and Pectic Oligosaccharides from Bergamot (Citrus bergamiaRisso) Peel. J. Agric. Food Chem. 2006, 54, 8307–8313. [Google Scholar] [CrossRef]
- Dominiak, M.; Søndergaard, K.M.; Wichmann, J.; Vidal-Melgosa, S.; Willats, W.; Meyer, A.S.; Mikkelsen, J.D. Application of enzymes for efficient extraction, modification, and development of functional properties of lime pectin. Food Hydrocoll. 2014, 40, 273–282. [Google Scholar] [CrossRef]
- Huynh, N.T.; Smagghe, G.; Gonzales, G.B.; Van Camp, J.; Raes, K. Enzyme-Assisted Extraction Enhancing the Phenolic Release from Cauliflower (Brassica oleracea L. var. botrytis) Outer Leaves. J. Agric. Food Chem. 2014, 62, 7468–7476. [Google Scholar] [CrossRef] [PubMed]
- Dranca, F.; Oroian, M. Optimization of Pectin Enzymatic Extraction from Malus domestica ’Fălticeni’ Apple Pomace with Celluclast 1.5L. Molecules 2019, 24, 2158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liew, S.; Chin, N.L.; Yusof, Y.; Sowndhararajan, K.; Sowndhararajan, K. Comparison of Acidic and Enzymatic Pectin Extraction from Passion Fruit Peels and Its Gel Properties. J. Food Process. Eng. 2015, 39, 501–511. [Google Scholar] [CrossRef]
- Moon, H.C.; Song, I.S.; Kim, J.C.; Shirai, Y.; Lee, N.H.; Kim, J.K.; Chung, S.O.; Kim, D.H.; Oh, K.K.; Cho, Y.S. Enzymatic hydrolysis of food waste and ethanol fermentation. Int. J. Energy Res. 2009, 33, 164–172. [Google Scholar] [CrossRef]
- Sungsopha, J.; Moongngarm, A.; Kanesakoo, R. Application of Germination and Enzymatic Treatment to Improve the Concentration of Bioactive Compounds and Antioxidant Activity of Rice Bran. AJBAS 2009, 9. [Google Scholar]
- Wanyo, P.; Meeso, N.; Siriamornpun, S. Effects of different treatments on the antioxidant properties and phenolic compounds of rice bran and rice husk. Food Chem. 2014, 157, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Faulds, C.B.; Mandalari, G.; LoCurto, R.; Bisignano, G.; Waldron, K.W. Arabinoxylan and mono- and dimeric ferulic acid release from brewer?s grain and wheat bran by feruloyl esterases and glycosyl hydrolases from Humicola insolens. Appl. Microbiol. Biotechnol. 2004, 64, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Connolly, A.; Cermeno, M.; Crowley, D.; O’Callaghan, Y.; O’Brien, N.M.; Fitzgerald, R.J. Characterisation of the in vitro bioactive properties of alkaline and enzyme extracted brewers’ spent grain protein hydrolysates. Food Res. Int. 2018, 121, 524–532. [Google Scholar] [CrossRef]
- Vidmantiene, D.; Juodeikiene, G.; Basinskiene, L. Technical ethanol production from waste of cereals and its products using a complex enzyme preparation. J. Sci. Food Agric. 2006, 86, 1732–1736. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, S.H.; Choi, Y.; Kim, G.D.; Chung, T. Effect of enzymatic pretreatment on acid fermentation of food waste. J. Chem. Technol. Biotechnol. 2006, 81, 974–980. [Google Scholar] [CrossRef]
- Gobbetti, M.; Di Cagno, R.; Calasso, M.; Neviani, E.; Fox, P.F.; De Angelis, M. Drivers that establish and assembly the lactic acid bacteria biota in cheeses. Trends Food Sci. Technol. 2018, 78, 244–254. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and functional paths of lactic acid bacteria in plant foods: Get out of the labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef]
- Minnaar, P.; Jolly, N.; Paulsen, V.; Du Plessis, H.; Van Der Rijst, M. Schizosaccharomyces pombe and Saccharomyces cerevisiae yeasts in sequential fermentations: Effect on phenolic acids of fermented Kei-apple (Dovyalis caffra L.) juice. Int. J. Food Microbiol. 2017, 257, 232–237. [Google Scholar] [CrossRef]
- Dey, T.B.; Chakraborty, S.; Jain, K.K.; Sharma, A.; Kuhad, R.C. Antioxidant phenolics and their microbial production by submerged and solid state fermentation process: A review. Trends Food Sci. Technol. 2016, 53, 60–74. [Google Scholar] [CrossRef]
- Madrera, R.R.; Bedriñana, R.P.; Valles, B.S. Enhancement of the nutritional properties of apple pomace by fermentation with autochthonous yeasts. LWT 2017, 79, 27–33. [Google Scholar] [CrossRef]
- Cantatore, V.; Filannino, P.; Gambacorta, G.; De Pasquale, I.; Pan, S.; Gobbetti, M.; Di Cagno, R. Lactic Acid Fermentation to Re-cycle Apple By-Products for Wheat Bread Fortification. Front. Microbiol. 2019, 10, 2574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moccia, F.; Flores-Gallegos, A.C.; Chávez-González, M.L.; Sepúlveda, L.; Marzorati, S.; Verotta, L.; Panzella, L.; Ascacio-Valdés, J.; Aguilar, C.N.; Napolitano, A. Ellagic Acid Recovery by Solid State Fermentation of Pomegranate Wastes by Aspergillus niger and Saccharomyces cerevisiae: A Comparison. Molecules 2019, 24, 3689. [Google Scholar] [CrossRef] [Green Version]
- Godard, J.; Hennequin, D.; Hallier, A.; Demarigny, Y. Evolution of carrot pulps during spontaneous fermentation. Int. J. Appl. Microbiol. Biotechnol. Res. 2018, 6, 95–106. [Google Scholar]
- Ricci, A.; Bernini, V.; Maoloni, A.; Cirlini, M.; Galaverna, G.; Neviani, E.; Lazzi, C. Vegetable By-Product Lacto-Fermentation as a New Source of Antimicrobial Compounds. Microorganisms 2019, 7, 607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagkli, D.M.; Corich, V.; Bovo, B.; Lante, A.; Giacomini, A. Antiradical and antimicrobial properties of fermented red chicory (Cichorium intybus L.) by-products. Ann. Microbiol. 2016, 66, 1377–1386. [Google Scholar] [CrossRef]
- Nanis, I.; Hatzikamari, M.; Katharopoulos, E.; Boukouvala, E.; Ekateriniadou, L.; Litopoulou-Tzanetaki, E.; Gerasopoulos, D. Microbiological and physicochemical changes during fermentation of solid residue of olive mill wastewaters: Exploitation towards the production of an olive paste – type product. LWT 2020, 117, 108671. [Google Scholar] [CrossRef]
- Durante, M.; Bleve, G.; Selvaggini, R.; Veneziani, G.; Servili, M.; Mita, G. Mita Bioactive Compounds and Stability of a Typical Italian Bakery Products “Taralli” Enriched with Fermented Olive Paste. Molecules 2019, 24, 3258. [Google Scholar] [CrossRef] [Green Version]
- Pontonio, E.; Dingeo, C.; Gobbetti, M.; Rizzello, C.G. Maize Milling By-Products: From Food Wastes to Functional Ingredients Through Lactic Acid Bacteria Fermentation. Front. Microbiol. 2019, 10, 561. [Google Scholar] [CrossRef] [Green Version]
- Arte, E.; Rizzello, C.G.; Verni, M.; Nordlund, E.; Katina, K.; Coda, R. Impact of Enzymatic and Microbial Bioprocessing on Protein Modification and Nutritional Properties of Wheat Bran. J. Agric. Food Chem. 2015, 63, 8685–8693. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; De Angelis, M.; Gobbetti, M. Effect of sourdough fermentation on stabilisation, and chemical and nutritional characteristics of wheat germ. Food Chem. 2010, 119, 1079–1089. [Google Scholar] [CrossRef]
- Pontonio, E.; Lorusso, A.; Gobbetti, M.; Rizzello, C.G. Use of fermented milling by-products as functional ingredient to develop a low-glycaemic index bread. J. Cereal Sci. 2017, 77, 235–242. [Google Scholar] [CrossRef]
- Ameur, H.; Cantatore, V.; Filannino, P.; Cavoski, I.; Gobbetti, M.; Di Cagno, R. Use of date-seeds flour as value added ingredient for wheat sourdough bread. 2020; Status (unpublished; manuscript in preparation). [Google Scholar]
- Filannino, P.; Azzi, L.; Cavoski, I.; Vincentini, O.; Rizzello, C.G.; Gobbetti, M.; Di Cagno, R. Exploitation of the health-promoting and sensory properties of organic pomegranate (Punica granatum L.) juice through lactic acid fermentation. Int. J. Food Microbiol. 2013, 163, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.; Matthews, K.R. Exposure of Escherichia coli O157:H7 to soil, manure, or water influences its survival on plants and initiation of plant defense response. Food Microbiol. 2014, 38, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Manini, F.; Brasca, M.; Plumed-Ferrer, C.; Morandi, S.; Erba, D.; Casiraghi, M.C. Study of the Chemical Changes and Evolution of Microbiota During Sourdoughlike Fermentation of Wheat Bran. Cereal Chem. J. 2014, 91, 342–349. [Google Scholar] [CrossRef]
- Zhao, H.-M.; Guo, X.-N.; Zhu, K. Impact of solid state fermentation on nutritional, physical and flavor properties of wheat bran. Food Chem. 2017, 217, 28–36. [Google Scholar] [CrossRef]
- Verni, M.; Rizzello, C.G.; Coda, R. Fermentation Biotechnology Applied to Cereal Industry By-Products: Nutritional and Functional Insights. Front. Nutr. 2019, 6, 42. [Google Scholar] [CrossRef] [Green Version]
- Boros, L.G.; Nichelatti, M.; Shoenfeld, Y. Fermented Wheat Germ Extract (Avemar) in the Treatment of Cancer and Autoimmune Diseases. Ann. N. Y. Acad. Sci. 2005, 1051, 529–542. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Mueller, T.; Coda, R.; Reipsch, F.; Nionelli, L.; Curiel, J.A.; Gobbetti, M. Synthesis of 2-methoxy benzoquinone and 2,6-dimethoxybenzoquinone by selected lactic acid bacteria during sourdough fermentation of wheat germ. Microb. Cell Factories 2013, 12, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
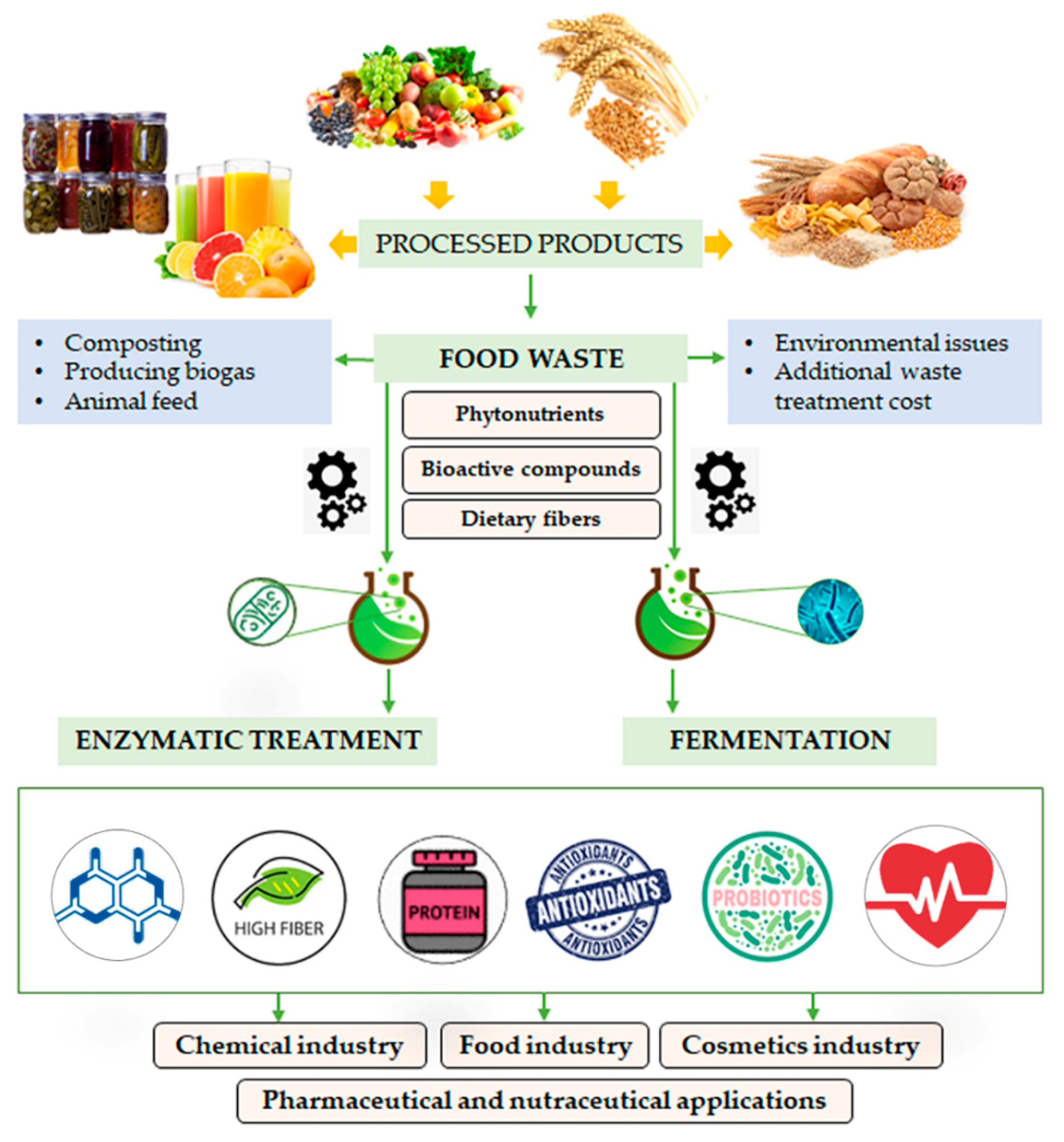{kind=link}
| Products | Byproducts | Total Dietary Fibers (%) | Insoluble Dietary Fibers (%) | Soluble Dietary Fibers (%) | Total Phenol (mg GAE 100 g−1 DM) | Total Flavonoids (mg QE 100 g−1 DM) | Antioxidant Activity | References | ||
|---|---|---|---|---|---|---|---|---|---|---|
| DPPH (mmol TE 100 g−1 DM) | ABTS (mmol TE 100 g−1 DM) | Others | ||||||||
| Apple | Seed | - | - | - | 286.1–514.1 | - | 2.14–4.35 | 9.15–39.79 | - | [11] |
| Peel | - | - | - | 564 | 303 | 22 ± 0.00 a | - | - | [12] | |
| Pomace | - | - | - | 348 | - | 72.6 ± 1.6 a | 84.3 ± 1.6 a | - | [13] | |
| Avocado | Seed | - | - | - | 57.3 | - | 41.07 ± 3.58 | 64.58 ± 1.79 | 65.69 ± 2.6 d (FRAP) | [14] |
| Peel | - | - | - | 63.5 | - | 31.0 ± 3.69 | 79.15 ± 3.59 | 117.51 ± 10.29 d (FRAP) | [14] | |
| Banana | Peel | 64.3 | 56.8 | 7.4 | 1500.2 | 900.3 | IC50: 55.23 b | - | 1.41 e (Cu2+ reduction) | [15] |
| Pomegranate | Peel | - | - | - | 45.7 | - | 92.09 ± 0.36 a | 97.14 ± 0.17 a | - | [16] |
| Dates | Seed | 9.5 | - | - | 442.1 | - | 87.23 ± 1.3 a | - | - | [17] |
| Mango | Seed | - | - | - | ca. 1920 | ca. 290 | ca. 90 c | ca. 620 c | - | [18] |
| Peel | 69.8 | 44.2 | 24.6 | 4500 | - | - | - | 6.96 f | [19] | |
| Pulp | - | 43.1 | 20.9 | 100 | 28.2 | - | - | - | [20] | |
| Citrus | Seed | - | - | - | 7.8 | - | IC50: 5.53 ± 0.08 b | - | 0.92 ± 0.01 g (AC) | [21] |
| Peel | 67.4 | 62.4 | 4.9 | 145.5 | - | 22.4 ± 0.07 | - | 2.24 ± 0.07 g (AC) | [22] | |
| Tomato | Pomace | - | - | - | 86.5 | - | - | - | 2.46 ± 0.3 g (AC) | [23] |
| Carrot | Pomace | - | - | - | ca. 500 | ca. 320 | 35 c | - | 45 c (CUPRAC) | [24] |
| Overview of Main HCV | Cereal Byproducts | |||
|---|---|---|---|---|
| Bioactive and Functional Compound | Beneficial Properties | Example of Case Study Demonstrating Beneficial Properties (referred to *) | Byproduct Containing the HCV | Examples of Industrial Application of Byproducts |
| Dietary fibers | Lowering of blood cholesterol*; improvements in large bowel function; reduction of colon tumors; attenuation of post-prandial blood glucose and insulin levels | Animal in vivo study showing a reduction of blood cholesterol levels at the concentration of 0.8 % in standard diet [25] | Wheat bran | Cosmetic and pharmaceutical formulations for moisturization of skin and mucosa; fortification of bread and surimi seafood [26] |
| Sterols | Reduction of blood cholesterol levels*; anti-inflammatory; anti-atherogenic; antioxidant; anti-carcinogenic | Animal in vivo tests showing a reduction of hypercholesterolemia at the concentration of 2455 ± 127 mg 100g−1 of total phytosterols [27] | ||
| Lignans | Decrease of the production of reactive oxygen species; protection against hormone related breast and prostate cancer; antitumor activity in colon cancer* | In vitro studies showing antitumor activity in colon cancer cells at concentration ≤ 40 µmol L−1 [28] | ||
| Phytic acid | Antioxidant activity; anticancer activity; reduction of serum lipids and inflammatory processes; prevention of kidney stone formation*; prevention of cardiovascular diseases | Animal in vivo studies showing a reduction of renal calcifications at concentration of 1.08·10−4 mol L−1 [29] | ||
| Essential fatty acids | Antiatherosclerotic effect; antioxidant activity; reduction of blood cholesterol; tryglicerides* | Human in vivo studies showing a decrease of plasma triglycerides in patients treated with 1.0-6.0 g day−1 of Omega-3-fatty acids [30] | Wheat germ | Fortification of bread, pasta and biscuits; formulation of nutritional complex for hair loss and health [31] |
| Ferulic acid and phenols | Cardiometabolic protection; anti-diabetic effect; antioxidant activity anti-inflammatory activity*; anti-hypertensive effects | In vitro studies showing the anti-inflammatory effect of hydroxycinnamic acid derivatives at concentration ranging from 1.84 to 378.66 µg g−1 [32] | Rice bran | Cosmetical formulations, production of sunscreen and treatment of skin-related disorders [33] |
| Tocopherols | Antioxidant activity* | In vitro studies showing the antioxidant activity of tocopherol at the concentration of 22.5 ± 0.6–170.5 ± 2.0 μg g−1 dry weight [34] | Rice germ | Incorporation in infant food [35] |
| Anthocyanins | Colorant effect* | In vitro studies showing colorant effect at concentration between 60 and 120 mg per 100 mL syrup [36] | Corn bran | Fortification of bread, cake and muffin; biosynthesis of natural aroma [37] |
| β-glucans | Anti-cancer property*; immune modulating activity; anti-aging and anti-inflammatory properties | Animal in vivo studies showing anti-tumor effect at both 40 µg and 400 µg dose levels [38] | Brewers´ spent grain | Pasta fortification [39] |
| Byproducts | Enymatic Treatment | Functional Components | References |
|---|---|---|---|
| Grape skin and seeds | Pectinase, cellulase and tannase | Proanthocyanidins | [69] |
| Grape seeds | Cellulases and pectinases | Polyphenols | [70] |
| Grape pomace | Tannase Cellulase | Gallic and syringic acids p-coumaric acid and malvidin-3-O-glucoside | [71] |
| Citrus residues | Pectinase, cellulase, tannase and β-glucosidase | Phenolic | [72] |
| Bergamot peel | Pectinase 62L, pectinase 690L and cellulase CO13 from Aspergillus sp. and Trichoderma sp. | Pectic oligosaccharides, glycosylated and deglycosylated flavonoids | [73] |
| Lime peel | Laminex C2K from Penicillium funiculosum | Pectin | [74] |
| Leaves of cauliflower | Viscozyme L and rapidase | Phenolic compounds and kaempferol-glucosides | [75] |
| Tomato residues | Cellulase and pectinase | Lycopene | [68] |
| Apple pomace, passion fruit peels | Celluclast 1.5L | Pectin | [76,77] |
| Fruits, vegetables and grains from cafeteria | Amyloglucosidase and carbohydrase | Ethanol | [78] |
| Rice bran | Protease type II from Aspergillus oryzae and α-amylase type XII-A from Bacillus licheniformis | γ-oryzanol, α-tocopherol and other phenolics | [79] |
| Rice bran and husk | Cellulase | γ-oryzanol and vanillic acid | [80] |
| Brewer’s grain and wheat bran | Ultraflo L | Ferulic acid, p-coumaric acid, cellulose and arabinoxylan | [81] |
| Brewery-spent Grain | Carbohydrases and peptidase | Carbohydrate- and protein-derived fractions | [82] |
| Wheat and rye bran | Amylase, glucoamylase and xylanase | Ethanol | [83] |
| Plant Matrices Byproduct | Microorganisms Involved in the Fermentation Process | Effects | Applications | Main Outcomes | Reference |
|---|---|---|---|---|---|
| Apple pomace | Saccharomyces cerevisiae, Saccharomyces bayanus and Hanseniaspora uvarum | Protein, fat and dietary fiber↑/Phenolic compounds: Quercetin and phloretin derivatives ↑/Oleic and linoleic acid derivatives ↑ | - | - | [89] |
| Apple byproducts | Weissella cibaria and Saccharomyces cerevisiae | Hydration properties ↑/ Total and insoluble dietary fibers ↑ | Fortification wheat bread with 5-10% fermented apple byproducts | Dough water absorption, stability and the content of dietary fibers ↑/Imparted agreeable and specific sensory attributes/Bread hydrolysis index ↓ | [90] |
| Pomegranate waste | Aspergillus niger and Saccharomyces cerevisiae | Punicalagin isomers and granatin B↓/Ellagic acid ↑ | - | - | [91] |
| Carrot peel | Spontaneous fermentation | Ethanol, acetic acid, lactic acid, 2-Butanol ↑ | - | - | [92] |
| Carrot, tomato and melon byproducts | Lactobacillus plantarum, Lactobacillus casei, Lactobacillus paracasei and Lactobacillus rhamnosus | Antimicrobial activity ↑ | - | - | [93] |
| Red chicory leaves | Saccharomyces cerevisiae, Lactobacillus plantarum and Lactobacillus hilgardii | Antimicrobial activity ↑/Antiradical and antioxidant activity ↑/Phenolic compounds: gallic and protocatechuic acids ↑ | - | - | [94] |
| Olive milling waste | Spontaneous fermentation + NaCl (6% w/v) | Oleuropein concentration ↓/Removing the bitter taste | - | - | [95] |
| Saccharomyces cerevisiae and Leuconostoc mesenteroids | Mono and polyunsaturated fatty acids ↑/Fat-soluble antioxidants ↑ | Enriching Taralli, a typical Italian bakery product, with 20% of fermented olive milling waste | Nutritional and health-promoting properties ↑/Polyphenols, triterpenic acids, tocochromanols and carotenoids ↑ | [96] | |
| Maize germ and bran | Lactobacillus plantarum and Weissella confusa | Total free amino acids and peptides ↑/Phytic acid ↓/Radical scavenging activity ↑/Inhibition endogenous lipase activity ↑ | Fortification wheat bread with 25% fermented maize byproducts | Dietary fiber and proteins ↑/Protein digestibility ↑/Starch hydrolysis index ↓ | [97] |
| Wheat bran | Lactobacillus brevis, Candida humilis, and cell-wall-degrading enzymes | Total free amino acids and protein digestibility ↑/Phenolic compounds and antioxidant properties ↑/Phytase activity ↑ | - | - | [98] |
| Wheat germ | Lactobacillus plantarum and Lactobacillus rossiae | Volatile compounds occurring in lipid oxidation ↓/Bioaccesibility of Ca2+, Fe2+, K+, Mn2+, Na+, and Zn2+ ↑/Total free amino acids ↑ Raffinose concentration ↓/Phenol content ↑/Scavenging activity toward free radical DPPH and ABTS ↑ | Fortification of wheat bread with 4% sourdough fermented wheat germ | Antifungal activity ↑/Shelf life ↑ | [58,99] |
| Wheat germ and bran | Lactobacillus plantarum and Lactobacillus rossiae | Improvement of functional properties | Fortification wheat bread with 15% fermented wheat byproducts | Total free amino acids and fiber content ↑/Phenolic compounds and antioxidant activity ↑/Glycaemic index ↓ | [100] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tlais, A.Z.A.; Fiorino, G.M.; Polo, A.; Filannino, P.; Di Cagno, R. High-Value Compounds in Fruit, Vegetable and Cereal Byproducts: An Overview of Potential Sustainable Reuse and Exploitation. Molecules 2020, 25, 2987. https://doi.org/10.3390/molecules25132987
Tlais AZA, Fiorino GM, Polo A, Filannino P, Di Cagno R. High-Value Compounds in Fruit, Vegetable and Cereal Byproducts: An Overview of Potential Sustainable Reuse and Exploitation. Molecules. 2020; 25(13):2987. https://doi.org/10.3390/molecules25132987
Chicago/Turabian StyleTlais, Ali Zein Alabiden, Giuseppina Maria Fiorino, Andrea Polo, Pasquale Filannino, and Raffaella Di Cagno. 2020. "High-Value Compounds in Fruit, Vegetable and Cereal Byproducts: An Overview of Potential Sustainable Reuse and Exploitation" Molecules 25, no. 13: 2987. https://doi.org/10.3390/molecules25132987