Physical and Physiological Monitoring on Red Palm Weevil-Infested Oil Palms

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. RPW Rearing
2.2. E. Guineensis Infestation with RPW Larva
2.3. Plantling Growth Measurement
2.4. Photosynthetic Activity Analysis
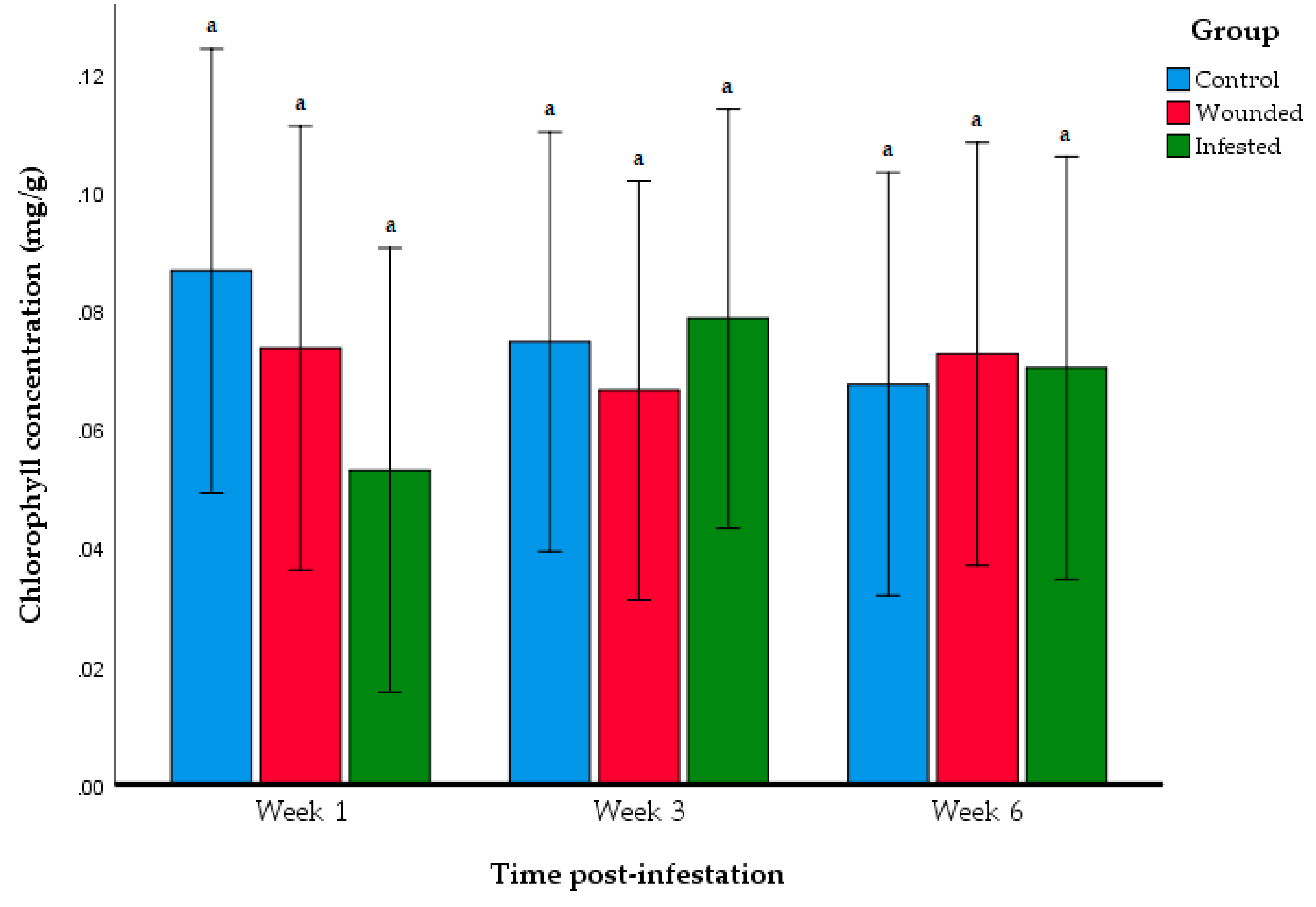
2.5. Chlorophyll Content Analysis
2.6. Statistical Analysis
3. Results
3.1. Symptoms of Infestation
3.2. Effect of RPW Infestation on Plant Growth Performance
3.3. Effect of RPW Infestation on Plant Photosynthesis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Azmi, W.A.; Daud, S.N.; Hussain, M.H.; Wai, Y.K.; Chik, Z.; Sajap, A.S. Field trapping of adult red palm weevil Rhynchophorus ferrugineus Olivier (Coleoptera: Curculionidae) with food baits and synthetic pheromone lure in a coconut plantation. Philipp. Agric. Sci. 2014, 97, 409–415. [Google Scholar]
- Azmi, W.A.; Lian, C.J.; Zakeri, H.A.; Yusuf, N.; Omar, W.B.W.; Wai, Y.K.; Zulkefli, A.N.; Hussain, M.H. The red palm weevil, Rhynchophorus ferrugineus: Current issues and challenges in Malaysia. Oil Palm Bull. 2017, 74, 17–24. [Google Scholar] [CrossRef]
- Murphy, S.; Briscoe, B. The red palm weevil as an alien invasive: Biology and the prospects for biological control as a component of IPM. Biocontrol News Inf. 1999, 20, 35–46. [Google Scholar]
- Rochat, D.; Dembilio, O.; Jaques, J.A.; Suma, P.; La Pergola, A.; Hamidi, R.; Kontodimas, D.; Soroker, V. Rhynchophorus ferrugineus: Taxonomy, distribution, biology, and life cycle. In Handbook of Major Palm Pests: Biology and Management; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 69–101. ISBN 978-1-119-05748-2. [Google Scholar]
- Wahizatul, A.A.; Zazali, C.; Abdul, R.; Nurul’Izzah, A.G. A new invasive coconut pest in Malaysia: The red palm weevil (Curculionidae: Rhynchophorus ferrugineus). Plant. Kuala Lumpur 2013, 89, 97–110. [Google Scholar]
- Kontodimas, D.; Soroker, V.; Pontikakos, C.; Suma, P.; Beaudoin-Ollivier, L.; Karamaouna, F.; Riolo, P. Visual identification and characterization of Rhynchophorus ferrugineus and Paysandisia archon infestation. Handb. Major Palm Pests Biol. Manag. 2016, 187–208. [Google Scholar] [CrossRef]
- Salem, S.A. Accuracy of trained dogs for early detection of red palm weevil, Rhynchophorus ferrugineus Oliv. infestations in date palm plantations. Swift J. Agric. Res. 2015, 1, 1–4. [Google Scholar]
- Hetzroni, A.; Soroker, V.; Cohen, Y. Toward practical acoustic red palm weevil detection. Comput. Electron. Agric. 2016, 124, 100–106. [Google Scholar] [CrossRef]
- Güerri-Agulló, B.; López-Follana, R.; Asensio, L.; Barranco, P.; Lopez-Llorca, L.V. Use of a solid formulation of Beauveria bassiana for biocontrol of the red palm weevil (Rhynchophorus ferrugineus) (Coleoptera: Dryophthoridae) under field conditions in SE Spain. Florida Entomol. Soc. 2011, 94, 737–747. [Google Scholar] [CrossRef]
- Soroker, V.; Suma, P.; Pergola, A.L.; Cohen, Y.; Akchanatis, V.; Golomb, O.; Alchanatis, V.; Golomb, O.; Goldshtein, E.; Hetzroni, A.; et al. Early detection and monitoring of red palm weevil: Approaches and challenges. In Proceedings of the AFPP—Palm Pest Mediterranean Conference, Nice, France, 16–18 January 2013; pp. 1–12. [Google Scholar]
- Stephens, A.E.A.; Westoby, M. Effects of insect attack to stems on plant survival, growth, reproduction and photosynthesis. Oikos 2015, 124, 266–273. [Google Scholar] [CrossRef]
- Golomb, O.; Alchanatis, V.; Cohen, Y.; Levin, N.; Soroker, V. Detection of red palm weevil infected trees using thermal imaging. In Proceedings of the 10th European Conference on Precision Agriculture (ECPA 2015), Tel Aviv, Israel, 12–16 July 2015; pp. 643–650. [Google Scholar]
- Jaafar, H.; Ibrahim, M.H. Photosynthesis and quantum yield of oil palm seedlings to elevated carbon dioxide. In Advances in Photosynthesis—Fundamental Aspects; IntechOpen: London, UK, 2012; pp. 321–340. ISBN 978-953-307-928-8. [Google Scholar]
- Soroker, V.; Suma, P.; Pergola, A.L.; Llopis, V.N. Surveillance techniques and detection methods for Rhynchophorus ferrugineus and Paysandisia archon. In Handbook of Major Palm Pests: Biology and Management; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 209–232. ISBN 978-1-119-05748-2. [Google Scholar]
- Al-Jabr, A.M.; Al-Khateeb, A.A.; Al-Khateeb, S.A.; Al-Ayied, H.Y. Effects of red palm weevil Rynchophorus ferrugineus (Olivier) infestation on gas exchange capacity of two date palm Phoenix dactylifera L. cultivars. J. Biol. Sci. 2007, 7, 1270–1273. [Google Scholar] [CrossRef] [Green Version]
- Hallett, R.H.; Oehlschlager, A.C.; Borden, J.H. Pheromone trapping protocols for the Asian palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae). Int. J. Pest Manag. 1999, 45, 231–237. [Google Scholar] [CrossRef]
- Haris, M.; Nang, M.; Chuah, T.; Azmi, W.A. The efficacy of synthetic food baits in capturing red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae) in campus area of Universiti Malaysia Terengganu. Serangga 2014, 19, 63–81. [Google Scholar]
- Wai, Y.K.; Bakar, A.A.; Azmi, W.A. Fecundity, fertility and survival of red palm weevil (Rhynchophorus ferrugineus) larvae reared on Sago Palm. Sains Malays. 2015, 44, 1371–1375. [Google Scholar]
- Alarcon, F.J.; Martınez, T.F.; Barranco, P.; Cabello, T.; Diaz, M.; Moyano, F. Digestive proteases during development of larvae of red palm weevil, Rhynchophorus ferrugineus (Olivier, 1790) (Coleoptera: Curculionidae). Insect Biochem. Mol. Biol. 2015, 32, 265–274. [Google Scholar] [CrossRef]
- Jalinas, J.; Güerri-Agulló, B.; Mankin, R.W.; López-Follana, R.; Lopez-Llorca, L.V. Acoustic assessment of Beauveria bassiana (Hypocreales: Clavicipitaceae) effects on Rhynchophorus ferrugineus (Coleoptera: Dryophthoridae) larval activity and mortality. J. Econ. Entomol. 2015, 108, 444–453. [Google Scholar] [CrossRef] [Green Version]
- Rasool, K.G.; Khan, M.A.; Tufail, M.; Husain, M.; Mehmood, K.; Mukhtar, M.; Takeda, M.; Aldawood, A.S.; Curculionidae, W.C.; Ghulam, K.; et al. Differential proteomic analysis of date palm leaves infested with the red palm weevil (Coleoptera: Curculionidae). Florida Entomol. 2018, 101, 290–298. [Google Scholar] [CrossRef] [Green Version]
- Rusli, M.H.; Idris, A.S.; Cooper, R.M. Evaluation of Malaysian oil palm progenies for susceptibility, resistance or tolerance to Fusarium oxysporum f. sp. elaeidis and defence-related gene expression in roots. Plant Pathol. 2015, 64, 638–647. [Google Scholar] [CrossRef]
- Lim, S.-L.; Subramaniam, S.; Zamzuri, I.; Amir, H.G. Growth and biochemical profiling of artificially associated micropropagated oil palm plantlets with Herbaspirillum seropedicae. J. Plant Interact. 2018, 13, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Haniff, M.H. Gas exchange of excised oil palm (Elaeis guineensis) fronds. Asian J. Plant Sci. 2006, 5, 9–13. [Google Scholar] [CrossRef]
- Ni, Z.; Kim, E.-D.; Chen, Z.J. Chlorophyll and starch assays. Protoc. Exch. 2009. [Google Scholar] [CrossRef]
- Corley, R.; Tinker, P. Growth, flowering and yield. In The Oil Palm, 4th ed.; Blackwell Science Ltd.: Oxford, UK, 2003; pp. 89–137. ISBN 978-111-895-329-7. [Google Scholar]
- Ralph, S.G.; Yueh, H.; Friedmann, M.; Aeschliman, D.; Zeznik, J.A.; Nelson, C.C.; Butterfield, Y.S.N.; Kirkpatrick, R.; Liu, J.; Jones, S.J.M.; et al. Conifer defence against insects: Microarray gene expression profiling of Sitka spruce (Picea sitchensis) induced by mechanical wounding or feeding by spruce budworms (Choristoneura occidentalis) or white pine weevils (Pissodes strobi). Plant Cell Environ. 2006, 29, 1545–1570. [Google Scholar] [CrossRef] [PubMed]
- Ferry, N.; Stavroulakis, S.; Guan, W.; Davison, G.M.; Bell, H.A.; Weaver, R.J.; Down, R.E.; Gatehouse, J.A.; Gatehouse, A.M.R.R. Molecular interactions between wheat and cereal aphid (Sitobion avenae): Analysis of changes to the wheat proteome. Proteomics 2011, 11, 1985–2002. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Mendes, Y.D.; Cuenca, J.C.; Romero, H.M. Physiological responses of oil palm (Elaeis guineensis Jacq.) seedlings under different water soil conditions. Agron. Colomb. 2016, 34, 163. [Google Scholar] [CrossRef]
- Azzeme, A.M.; Abdullah, S.N.A.; Aziz, M.A.; Wahab, P.E.M. Oil palm leaves and roots differ in physiological response, antioxidant enzyme activities and expression of stress-responsive genes upon exposure to drought stress. Acta Physiol. Plant. 2016, 38, 1–12. [Google Scholar] [CrossRef]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef]
- Rejeb, I.; Pastor, V.; Mauch-Mani, B. Plant responses to simultaneous biotic and abiotic stress: Molecular mechanisms. Plants 2014, 3, 458–475. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level | Figure 1 | Descriptions |
|---|---|---|
| 1 (Early stage) | A, B |
|
| 2 (Intermediate I) | C, D |
|
| 3 (Intermediate II) | E, F, G |
|
| 4 (Final stage) | H, I |
|
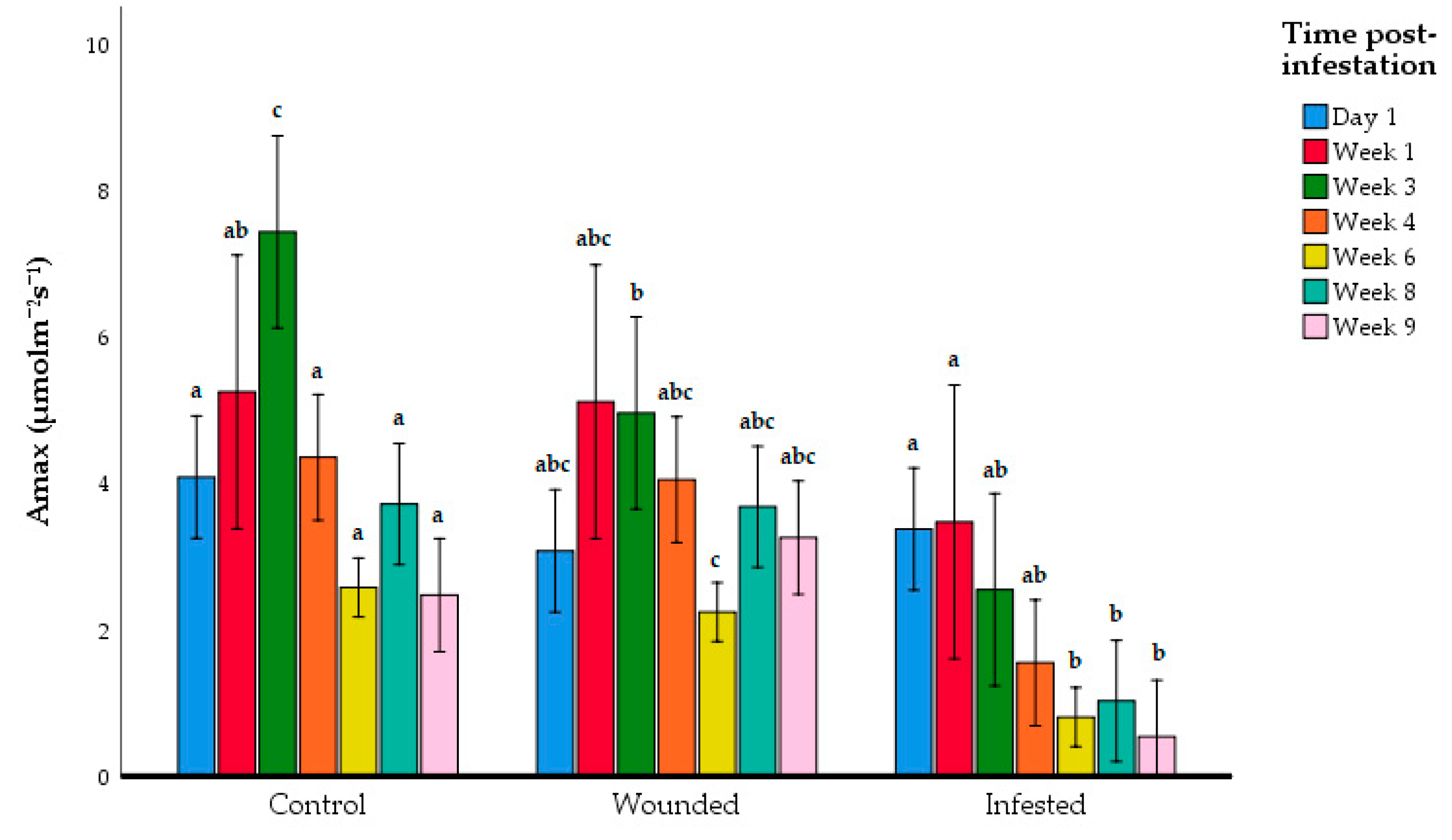
| Group | Day 1 | Week 1 | Week 3 | Week 4 | Week6 | Week 8 | Week 9 |
|---|---|---|---|---|---|---|---|
| Control | 4.19 ± 1.13 a | 3.66 ± 1.10 a | 7.44 ± 1.103 a | 4.362 ± 0.753 a | 2.583 ± 0.326 a | 3.724 ± 1.12 a | 2.48 ± 0.567 a |
| Wounded | 3.083 ± 0.90 a | 5.122 ± 1.09 a | 4.97 ± 1.91 a,b | 4.057 ± 1.34 a | 2.25 ± 0.602 a | 3.687 ± 0.909 a | 3.265 ± 1.23 a |
| Infested | 3.81 ± 1.27 a | 3.19 ± 1.54 a | 2.55 ± 1.408 b | 1.56 ± 0.754 b | 0.812 ± 0.418 b | 1.03 ± 0.79 b | 0.543 ± 0.618 b |
| ANOVA F-value | 1.74 | 2.35 | 15.7 | 14.6 | 24.8 | 15.8 | 14.9 |
| p-value | 0.290 | 0.130 | 2.08 × 10−4 | 3.05 × 10−4 | 1.77 × 10−5 | 2.02 × 10−4 | 2.74 × 10−4 |
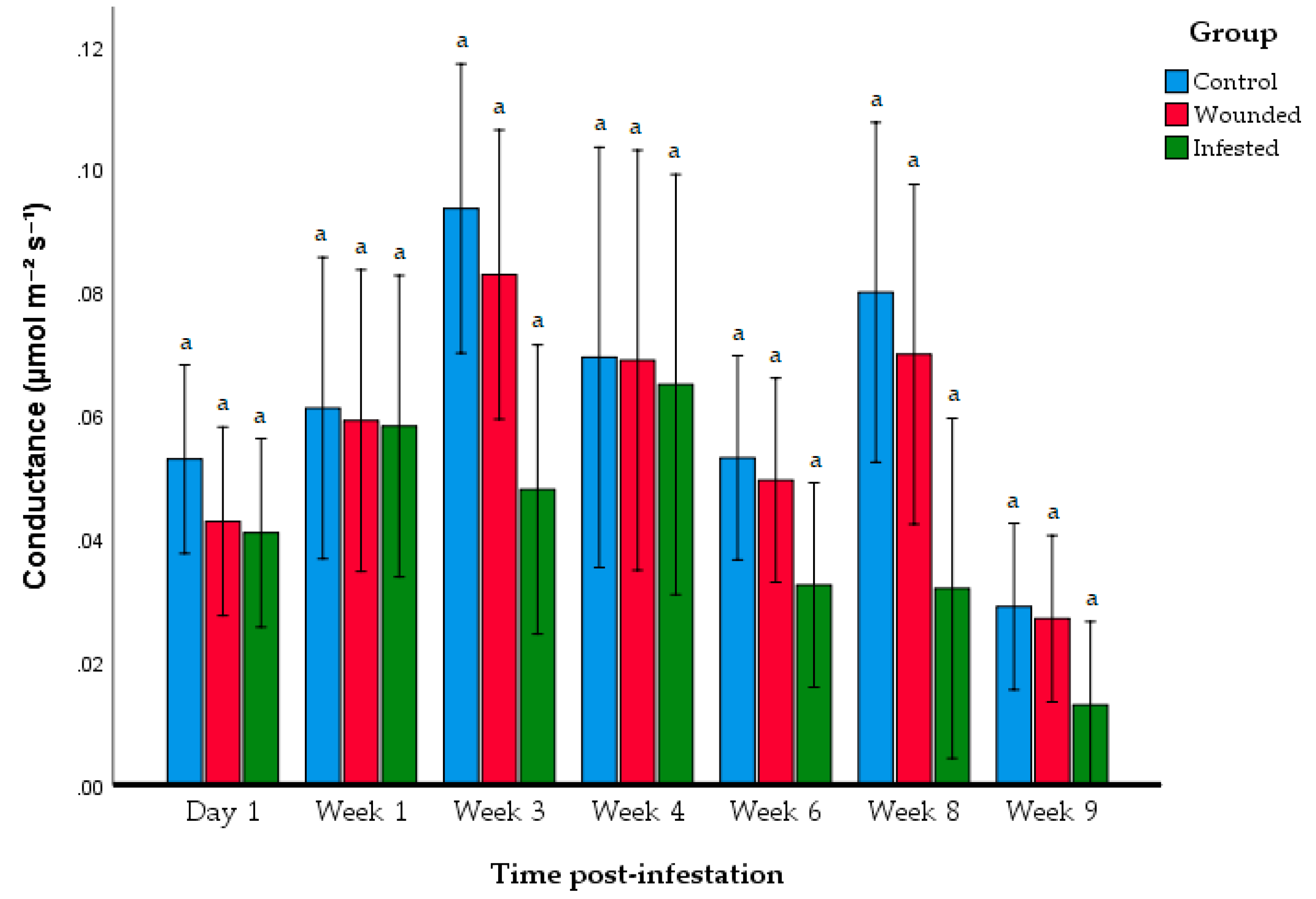
| Group | Day 1 | Week 1 | Week 3 | Week 4 | Week6 | Week 8 | Week 9 |
|---|---|---|---|---|---|---|---|
| Control | 0.053 ± 0.005 a | 0.0612 ± 0.0133 a | 0.0936 ± 0.0151 a | 0.0694 ± 0.02553 a | 0.0531 ± 0.0153 a | 0.0799 ± 0.0197 a | 0.029 ± 0.0128 a |
| Wounded | 0.428 ± 0.233 a | 0.0591 ± 0.0293 a | 0.0828 ± 0.0359 a | 0.0689 ± 0.0418 a | 0.0195 ± 0.0220 a | 0.0699 ± 0.0427 a | 0.027 ± 0.0211 a |
| Infested | 0.049 ± 0.189 a | 0.0582 ± 0.0365 a | 0.0480 ± 0.02577 a | 0.065 ± 0.469 a | 0.0325 ± 0.0192 a | 0.0319 ± 0.0284 a | 0.0130 ± 0.00106 a |
| ANOVA F-value | 0.804 | 0.017 | 4.69 | 0.023 | 2.004 | 3.84 | 1.89 |
| p-value | 0.466 | 0.983 | 0.066 | 0.977 | 0.169 | 0.065 | 0.186 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harith-Fadzilah, N.; Haris-Hussain, M.; Abd Ghani, I.; Zakaria, A.; Amit, S.; Zainal, Z.; Azmi, W.A.; Jalinas, J.; Hassan, M. Physical and Physiological Monitoring on Red Palm Weevil-Infested Oil Palms. Insects 2020, 11, 407. https://doi.org/10.3390/insects11070407
Harith-Fadzilah N, Haris-Hussain M, Abd Ghani I, Zakaria A, Amit S, Zainal Z, Azmi WA, Jalinas J, Hassan M. Physical and Physiological Monitoring on Red Palm Weevil-Infested Oil Palms. Insects. 2020; 11(7):407. https://doi.org/10.3390/insects11070407
Chicago/Turabian StyleHarith-Fadzilah, Nazmi, Mohamad Haris-Hussain, Idris Abd Ghani, Azlina Zakaria, Samsudin Amit, Zamri Zainal, Wahizatul Afzan Azmi, Johari Jalinas, and Maizom Hassan. 2020. "Physical and Physiological Monitoring on Red Palm Weevil-Infested Oil Palms" Insects 11, no. 7: 407. https://doi.org/10.3390/insects11070407