Habitat Models of Focal Species Can Link Ecology and Decision-Making in Sustainable Forest Management


1
Department of Zoology, Institute of Ecology and Earth Sciences, University of Tartu, Vanemuise 46, EE-51005 Tartu, Estonia
2
Department of Ecology, Swedish University of Agricultural Sciences, Box 7044, 750 07 Uppsala, Sweden
*
Author to whom correspondence should be addressed.
Forests 2020, 11(7), 721; https://doi.org/10.3390/f11070721
Submission received: 20 May 2020
/
Revised: 24 June 2020
/
Accepted: 28 June 2020
/
Published: 30 June 2020
(This article belongs to the Special Issue Modeling of Species Distribution and Biodiversity in Forests)
Abstract
:A fundamental problem of sustainability is how to reduce the double complexity of ecological and social systems into simple operational terms. We highlight that the conservation concept of focal species (selected species sensitive to a set of anthropogenic threats to their habitat) links multiple issues of ecological sustainability, and their habitat models can provide a practical tool for solving these issues. A review of the literature shows that most spatial modeling of focal species focuses on vertebrates, lacks the aspect of aquatic and soil habitats, and has been slow in the uptake by actual management planning. We elaborate on a deductive modeling approach that first generalizes the main influential dimensions of habitat change (threats), which are then parameterized as habitat quality estimates for focal species. If built on theoretical understanding and properly scaled, the maps produced with such models can cost-effectively describe the dynamics of ecological qualities across forest landscapes, help set conservation priorities, and reflect on management plans and practices. The models also serve as ecological hypotheses on biodiversity and landscape function. We illustrate this approach based on recent additions to the forest reserve network in Estonia, which addressed the insufficient protection of productive forest types. For this purpose, mostly former production forests that may require restoration were set aside. We distinguished seven major habitat dimensions and their representative taxa in these forests and depicted each dimension as a practical stand-scale decision tree of habitat quality. The model outcomes implied that popular stand-structural targets of active forest restoration would recover passively in reasonable time in these areas, while a critically degraded condition (loss of old trees of characteristic species) required management beyond reserve borders. Another hidden issue revealed was that only a few stands of consistently low habitat quality concentrated in the landscape to allow cost-efficient restoration planning. We conclude that useful habitat models for sustainable forest management have to balance single-species realism with stakeholder expectations of meaningful targets and scales. Addressing such social aspects through the focal species concept could accelerate the adoption of biodiversity distribution modeling in forestry.
1. Introduction
Biodiversity issues in sustainable forest management (SFM) are changing, which brings along the need for new analytical tools. A major change is that sustainability is increasingly defined through connected ecological and social complex systems at multiple scales [1]; this adds adaptive capacity and resilience among key qualities of SFM, along with traditional expectations to sustain a supply of specific forest goods and services [2,3,4]. Thus, biodiversity concerns have transcended traditional nature conservation to become an integrative issue that underpins ecological resilience, adaptive capacity of ecosystems, and many ecosystem services [5,6,7,8]. It is yet unclear how such a perspective will be put into practice (e.g., [9,10,11]). However, given the schism between broad political acceptance of SFM and of forest protection [12,13] versus the continuing loss of forest biodiversity [14], there is an unprecedented need for clear biodiversity targets and tools.
An obvious goal in sustaining biodiversity is to manage harmful environmental pressures and threats rapidly, proactively, and effectively. Geospatial models have long been used to anticipate futures by spatial planning of forests, including biodiversity targets [15,16,17,18,19,20]. Many modeling approaches and techniques have been developed for depicting and accounting for biodiversity across forest landscapes (e.g., [21,22,23]). Recent technological progress enables mass recording of biodiversity variables, e.g., by combining molecular sampling, observations, and remote sensing (e.g., [24,25,26]). However, such advances in biodiversity modeling are not easily picked up by forestry and conservation planners to specify general ecological guidance (e.g., [27,28]). Thus, biodiversity assessment practices for SFM or in protected forest habitats are based mostly on convenient landscape metrics and woody vegetation proxies ([29,30,31,32,33,34], but see [35,36]). The species included in the landscape-scale predictive models of forestry scenarios are defined case-wise for specific purposes (see Section 2.2), while legitimate procedures of setting aside forest stands for biodiversity tend to require laborious field documentation (e.g., [37]).
In this paper, we highlight a biodiversity response variable as a critical issue for useful geospatial models in SFM. The large and diffuse literature on such variables (e.g., [38,39,40]) indicates a narrow disciplinary focus of most spatial models. We identify at least four ‘interdisciplinary gaps’ (sensu [41]) to be considered by the biodiversity modeling community (see also [23,42,43,44]).
- Biodiversity-representation gap—attempts to describe biodiversity comprehensively are common when analyzing current management situations, while most scenario analyses do not address representation beyond woody vegetation (e.g., [45]).
- Scale-relevance gap—typical units of forest management and conservation decisions are either single trees, forest stands, or mosaics of stands (landscapes), while most biodiversity data are collected or modeled in other units (plots; pixels; etc.) that cannot be easily combined for decision support. Local biodiversity patterns, in turn, result from wider and longer-term ecological processes, which are difficult to explicitly incorporate in the models (e.g., [49]).
- Feedback gap—realistic biodiversity models tend to become very complicated (e.g., [17,43]), which undermines their updating, reduces advantages over adaptive management, and limits communication and uptake by the wider public. Infrequent or one-sided communication, in turn, reduces the ability to mobilize knowledge for action [50,51].
In brief, geospatial predictive models can be irreplaceable tools for biodiversity issues in SFM, given the large area of forests, long temporal scales of forest development, and the vast nature of biodiversity. However, there are apparently communication boundaries between the modeling community and other stakeholders (biodiversity researchers; policy-makers; forest and conservation managers; wider public). For example, the first two interdisciplinary gaps above together reflect ‘biodiversity concerns’ that have been recognized as difficult to model [46]. Social and governance studies suggest that the failure to manage communication boundaries by meaningful simplification makes biodiversity difficult to grasp even for professionals [52,53]. The question is how to build models that clarify biodiversity issues and help to plan a meaningful and understandable future.
Here, we revisit the conservation concept of ‘focal species’ as proposed by Lambeck [54,55], who proposed setting environmental standards in a specific context according to the most sensitive species to each threatening process in the environment. Managing for a full set of such species might then encapsulate the biodiversity conservation aim of a landscape. We elaborate this concept in a spatial modeling perspective to demonstrate how it can—mostly through the binding element of ‘threatening process’—operationalize multiple issues of ecological sustainability and bridge the interdisciplinary gaps listed above. We retain Lambeck’s original term for ‘focal species’, while acknowledging that it has been loosely used in the literature and must be routinely rechecked against the original definition [56] (pp. 17–22).
The paper is organized as follows. We first explain the concept and review the literature on spatial habitat modeling of focal species for SFM. We assess the coverage of the current research in terms of biodiversity and the forestry problems it might address. We then illustrate an approach that focuses on the threatening process, conceptualizing it through major dimensions of habitat change. We list the main merits of such an approach from a practical modeling perspective, including parameterizing the model as habitat quality and quantity estimates for focal species. The latter is a well-established modeling field. Finally, we illustrate our approach based on recent additions to the forest reserve network in Estonia, where the practical question is the time scale and expected spatial pattern of recovery of degraded habitats and allocation of management to enhance this.
2. A Spatial Modeling Perspective on Focal Species
2.1. Theoretical Background
Simply put, the practical question of environmental management is where to do what to sustain environmental quality. For the SFM principle of maintaining and restoring biodiversity, it translates to locating and managing threats and opportunities for biodiversity on dynamic landscapes [57,58,59]. There are four classes of spatial management decisions involved: stand-scale management for single or multiple goals; landscape design by combining stand-scale goals for landscape functions (including setting aside protected areas); regulating forest benefits and values in time; and managing for uncertainty at multiple scales.
In our view, the strength of Lambeck’s [54] concept of focal species is that it integrates these strategic aspects in a way that is understandable to the wider public, thus serving the stakeholder participation principle of SFM [60]. Specifically:
- Selecting well-defined sensitive species to represent a full set of threats to biodiversity simplifies practical biodiversity concerns (the representation and goal-setting gaps above). A useful input is the red-listing of species based on the International Union for Conservation of Nature (IUCN) framework, which also considers ‘projected declines’ based on potential threats and changes in habitat area and quality [61,62].
- Managing (avoiding, mitigating, or reversing) each threat to sustain focal species in actual landscapes links the concern with management responses (Figure 1) and implicitly addresses some uncertainty (e.g., maintaining population ‘at the safe side’). A key simplification is that focal species serve simultaneously as biodiversity indicators and management targets (cf. [63]), while such a link is unspecified in other biodiversity schemes for SFM (e.g., [35,64]).
The opportunity for spatial models in this framework is to predict focal species’ distribution or performance in real landscapes subject to expected or designed change. Such spatial predictions are derived from species–habitat relationships, where specific ‘threats’ refer to limiting factors or population processes of the focal species. Note that, for modeling these links, ecological niches of the species must be understood beyond correlative patterns in current distributions [70]; thus, so-called black-box modeling approaches that fit environmental parameters without understanding their ecological meaning [71] are discouraged. Due to the underlying logic that ‘sensitive species illustrate a general threat’, the maps derived from properly parameterized spatial models might then help to depict, analyze, and communicate broader ‘where’ and ‘how’ of sustaining forest biodiversity in the spatial and temporal scales for which the parameter values are available (Figure 1). Depending on how closely the species’ distributions follow environmental threats in time, such maps may also reveal past or present spatial extent, severity, and reversibility of the threats (e.g., [72,73,74]).
The methods for predictive mapping of species distributions can be divided into correlative (inductive) and mechanistic (deductive) models. The inductive methods, where predictions are derived from statistically linking empirical observations with habitat characteristics, include many algorithms and programming tools available [71]. The algorithms basically differ depending on the species data (presence-only, presence–absence, or finer scales) and shapes of its habitat function. The deductive methods are based on prior insight into the species’ requirements, with a wide range of more and less formal approaches, including procedures for systematizing expert knowledge (e.g., [75,76]). For either class of models to enable spatially explicit management guidance, they should be able to depict landscape change (including alternative management scenarios) in terms of the factors that indicate threats.
There are several basic caveats in interpreting predicted distributions of focal species directly for broader biodiversity management. First, each species can be limited by multiple factors, and its distribution is affected by stochastic events and population processes [55,77,78]. This implies that both realized and potential (habitat) distributions of species affected by the same threat only overlap partly and to an extent that varies in time. Hence, increasing model prediction accuracy for a specific species—a major technical aim of distribution modeling [71]—can paradoxically reduce the insight obtained from the model about wider biodiversity. Second, uncertainty of most environmental parameters increases when predicting the future, and data quality is usually reduced toward the past as well. This, too, means general problems with predictions, specifically for complex models. The issue is to find simple and robust habitat characteristics that change predictably in time. Third, most sensitive species may be very rare or even extirpated in the degraded landscapes where habitat improvement is most needed; thus, their habitat prediction may not be practical or reliable for the remaining biodiversity [77]. Instead of completely ignoring such species, a possibility is to add species less vulnerable to the same threat to be able to cover a broad range of environmental change.
An alternative to habitat modeling of threatened species is to map threats (hazards) directly. Such ‘exposure maps’ have been created by remote sensing of whole landscapes (e.g., for fire frequency, deforestation, night-time lights [79,80]) or modeling point observations based on landscape characteristics (for example, poaching threat maps from camera trapping of poached animals [81]). Threat maps, though, are not explicit about likely biodiversity responses, which in turn limits objective-setting and cost-effective spatial analysis of conservation actions [82]. In a structured decision-making process, focal species thus serve as a multi-purpose tool to set the objectives, choose among actions, and to measure the success.
2.2. Published Spatial Models of Focal Species Performance
Modern techniques of creating species habitat maps for forest management prescriptions originate from the rapid development of spatial analysis in environmental protection, wildlife ecology, and threatened species research in the 1980s. Notably, the United States (US) Forest Service developed Habitat Evaluation Procedures in the 1970s, which became increasingly formalized, supported by guidelines of use and computer programs [83]. Along with the appearance and acceptance of GIS-techniques and data, such procedures transformed from individual assessment to automated landscape analysis (e.g., [84]). In the high-profile conservation case of the spotted owl (Strix occidentalis), GIS models were linked with population models in real landscapes first in 1992 [85]. In parallel, there was a development from prescribing forestry activities from the perspective of a single subjectively selected species toward comprehensive sets of species to represent different niche dimensions of habitat specialists ([86,87] or species requiring large areas [88]. In retrospect, these US approaches appear closer to SFM than Lambeck’s [54] Australian view that emphasized protection and restoration. The new aspect brought up by the latter was, however, that species should be used to analyze habitat futures, not just to maintain the present values.
To characterize the field’s development since then, we performed a search of modeling studies that spatially predicted the performance (incidence, trends, or demography) of representative species in real forest landscapes and through time, and in response to threats that could be mitigated by forest management or conservation. We performed initial searches on 8 April 2020, using the Scopus database and two alternative search strings: (i) TITLE-ABS-KEY (“focal species" AND forest) AND TITLE-ABS-KEY (model* OR predict* OR simulat*); (ii) REF (“Lambeck”) AND TITLE-ABS-KEY (forest) AND TITLE-ABS-KEY (model* OR predict* OR simulat*). We excluded irrelevant studies by considering the title and, if unclear, by the abstract. Full texts of the remaining 38 studies were then assessed for whether they included a forestry perspective s. lat. (i.e., including also policies, planning of landscapes and set-asides) and met at least three of the following four criteria: A, addressed environmental threats that also affect wider, at least partly known range of forest taxa (note that, for practical purposes, we here restricted the focal species surrogacy assessment to species diversity only); B, included management options or approaches that also affect wider, known species diversity; C, were a part of a legitimate planning process; D, described how the focal taxa were selected based on specific threats and their surrogate value to represent wider species’ diversity. If these criteria were supported by references only, we also checked the original publications. Finally, we integrated a series of papers by the same research group in the same study system, and assessed potential gaps in the search string for additional searches on specific topics (metapopulation models; forest water bodies).
Thirteen of the 19 focal-species’ modeling studies detected address North American forests (Table 1). Another pattern is that such modeling has remained [89], at least in forestry, largely based on vertebrates. This is despite the problems with cross-taxon congruence being well acknowledged [56]. In fact, many field surveys have addressed potential non-vertebrate surrogate taxa. For example, forest fungal surrogates have been explored in many studies [90], including the matching of selected wood-inhabiting species with threats [91,92]. Specialized lichens appear suitable for guiding multiple management dimensions [93], but spatial models for that remain scarce (Table 1). Modeling for decision-making may have thus contributed to the taxonomic bias in SFM, which is usually attributed to insufficient stakeholder knowledge [94]. A similar gap appears in ecosystem coverage regarding the management of small freshwater bodies, notably headwater streams in forests. Again, there is well-established literature on the indicator value of many aquatic or semi-aquatic taxa, including suggestions to use some invertebrates, fish, amphibians, or birds as broader management targets (e.g., [95,96,97]). Relevant spatial models are, however, rare and tend to focus solely on the species’ indicator value (e.g., [98]) or its conservation perspectives (e.g., [99]).
In our view, most studies listed in Table 1 appear relevant to support decision-making at different levels. This contrasts with an overall scarcity of such studies, of which (judging from the statements in original studies), even fewer were parts of actual decision-making processes. There may be two reasons for such neglect. First, modeling of futures relies on diverse assumptions on the system’s behavior, so that the best focal-species ‘models’ are actually sets of several linked models depicting social, climatic, ecosystem, and population changes. To develop such sets may require expensive study programs, such as the Northwest Forest Plan, Coastal Landscape Analysis and Modeling Study (CLAMS) [121] or the Forest Landscape Disturbance and Succession (LANDIS) model programs in the US [122,123]. Second, there may be broader political inertia in the SFM and forest conservation, which inhibits the practical adoption of new analytical tools for designing futures [124,125,126]. Such inertia is particularly harmful to biodiversity when it suppresses spatial planning under the conditions of increasing timber harvest since spatial solutions are among those few that could mitigate such pressure [127]. Institutional collaboration for mutual understanding of research development might help in both cases (e.g., [125,128]).
2.3. Key Issues for Practical Spatial Models of Focal Species
One way to address the apparently under-used potential of focal-species models in SFM is to clarify the technical issues that could be improved to better contribute to decision-making. For that, we list four major issues that follow from our understanding of focal species as integrating many ecological, social, and cultural aspects of SFM.
- The rationale that focal species serve both as indicators and management goals promotes linking their models strategically with other decision-making tools. The underlying concept of ’threat’ instantly makes sense for ecological risk assessment [129], but some harmonization may be required to also link it with specific ‘pressures’ in DPSIR (drivers–pressures–state–impact–response) and related causal frameworks of biodiversity or environmental management [130,131,132]. For ecosystem analysis, representative sets of focal species can operationalize the issue of ecological integrity [129,133] and help prioritize ecological risks based on irreversible damage. In management, focal species could inspire the development of new forestry approaches if seen as organizational goals subject to the SMART (specific, measurable, attainable, realistic, and time-sensitive) criteria [134] and educational capacity-building.
- The spatial models are most useful when they collectively map most of the risk dimensions of the environment rather than the performance of individual species. More work is needed on how to define such dimensions, how to analyze their ecological trade-offs and optimize solutions that also consider socio-economic aspects. An established practice, which has the advantage of including future threats and recovery of extirpated taxa, is to start from conceptualizing vulnerable niches in the environment (‘ecological profiles’) and selecting species representing such niches [100,133,135,136]. An ecological question is the level of generalization for that, with extremes represented by models based on ‘theoretical species’ (e.g., [72,137,138]) versus complex real-species models to maximize fit with the data [139,140,141]. Based on our experience with broader understandability issues in environmental decision-making (see also [142,143]), we suggest that a middle ground of simplified, limiting-factor based models of real focal species might serve practical goals best. For such generalization, deductive models have advantages over inductive models (e.g., [104,105]), but only in landscapes and ecosystems known well enough.
- Some basic tensions of SFM suggest that at least the following technical qualities are important in focal-species models. (a) Dynamic modeling over decadal time-scales. Static models are of limited use since the main practical challenge is how to balance short- vs. long-term perspectives. (b) Preferring a full range of focal-species responses [144] over quantitative accuracy within a limited range. If managers prioritize actions (scenarios), an ordinal response scale may suffice (e.g., [75,145] and allow less-studied taxa to be modeled. A useful qualitative framework is to distinguish fundamental-niche, realized-niche, source-sink, and dispersal-limited locations [139]. (c) The aspects of time frame and decision-relevance also apply to input data. It is important to utilize data sources that are maintained for wider purposes, over long periods (including historical data), and are legitimate to stakeholders. Stand-structural and tree-composition variables of national forest surveys are specifically promising [146,147,148], also given the general trend to address SFM criteria and indicators at the operational unit (stand) scale [30]. (d) Uncertainty remains a part of any model, but it can be at least described [75]. Such descriptions can be linked with the precautionary principle and safe minimum standards relevant to SFM and conservation management. Uncertainty can also vary in space; usefully, it may be the smallest in the highest-priority locations [117].
- Good maps help to tell a story that matters to people. This recognizes the basic principles of how policy-makers and other stakeholders think and work [149,150]. A dimension worth considering for depicting management scenarios is human activities and personal experiences [151,152], including researcher–stakeholder collaboration in producing the spatial models [153].
3. The Case Study: Protecting Degraded Forests in Estonia
3.1. The Problem and the Setup
We exemplify the potential contribution of focal-species modeling based on a recent decision in Estonia to include into national reserve network some forest types for which natural areas have been largely lost. The situation that the most productive forests are underrepresented in reserves and ecologically impoverished outside due to intensive use is common in developed countries [154,155,156]. Research has shown that some forest structures can spontaneously recover within a few decades (e.g., [157,158]), but demanding species re-colonize with a delay [159,160,161], and it is unclear whether these processes would benefit from active restoration [72,162,163].
The Estonian case followed from an analysis made in 2002 for the national forestry development plan, which identified forest protection gaps for old-growth biodiversity by site type [164]. A 2016 ministerial review set the remaining gaps as quantitative targets, prioritizing to set aside additional eutrophic (149 km2) and meso-eutrophic (147 km2) forests. It was clear that reasonably large patches of such areas only existed in impoverished states, but emerging research suggested that their protection might still pay off in the long run [165,166]. In 2017–2018, the Ministry of the Environment, the State Forest Management Centre, researchers, and environmental NGOs collectively identified a cost-effective selection of state lands that would cover most of the gap. A total of 286 km2 (1.2% of Estonian forest land) was set aside as a result, mostly by a single governmental decision in February 2019 (58 new strict reserves; 267 km2; including 25% meso-eutrophic, 36% eutrophic, and 16% eutrophic-paludified types). Here, we use spatial modeling to analyze the reversibility of the most vulnerable and degraded ecological conditions significant for biodiversity. The models will be used as a basis to assess restoration potential in these new reserves.
The reserves comprise 106 distinct patches all over the country, on average 2.7 km2 (range 0.03–28.5 km2) in size, and with a heavy management footprint. According to historical maps, the area has had >100 km of natural watercourses; ca. 30% remains in its natural streambed, but 56% has been straightened, and 14% is lost due to forestry drainage. Until the mid-20th century, 8% of the forest area was under some agricultural use (arable land; pastures; wooded grasslands). Most other forests have been converted by production forestry into mosaics of forest stands in various successional stages interspersed by networks of drainage ditches and forest roads. Stands >100 years old, which are the ecosystem targets of strict protection [164], cover only 7% of the area. The rest are recent clearcuts (11%), stands <20 years (28%), 21–60 years (28%), or 61–100 years old (27%). Artificial regeneration has been used on 45% of all forest land, mostly with Norway spruce (Picea abies), but the current share of the planted component varies widely among stands. Besides clearcutting, pre-commercial thinning (9% of forest land), and thinnings and sanitation cuttings (24%) have been used within the last 20 years. Of stands >20 years old, 47% are mixed, 30% conifer, and 23% deciduous forests. The main tree species are P. abies (42%), Betula spp. (25%), Pinus sylvestris (17%), Populus tremula (10%), Alnus incana (2%), and A. glutinosa (2%). Nemoral hardwoods, characteristic of such natural forests (Quercus robur, Tilia cordata, Fraxinus excelsior, Acer platanoides, Ulmus spp.), now only occur at small frequencies.
3.2. The Modeling Approach and Inference
We defined major threats as distinct empirically supported habitat dimensions, along which production forestry can reduce natural species pools of eutrophic and meso-eutrophic forests. The expert-based process (including two meetings) involved lead forest biodiversity experts in the country, with knowledge of multiple taxon groups. The forest types under question are well defined by topographic and soil conditions and have diverse species pools [167,168]. In a natural state, these forests have complex uneven-aged or all-aged structure created by gap-dynamics and, depending on moisture, rare stand replacements (mostly due to storm or pathogens) [164,169,170]. This structure is greatly simplified by clearcutting based forestry that uses 30–70-year rotations and a few selected tree-species, notably pioneer deciduous trees and planting of Picea abies over large areas [170,171,172,173]. However, rapid tree growth also accelerates structural recovery after abandoned management or long rotations based on natural regeneration [170,173,174]. Many well-dispersing old-forest species can colonize such forests at longer rotations [92,175], while others remain excluded due to the absence of old-forest structures [93] or (poor dispersers) lack of local refugia [176].
Considering these patterns, we defined seven main ‘ecological profiles’ of focal specialist species and their management-affected limiting factors in priority order (Table 2). The factors were then formalized as decision trees and parameterized based on requirements of the focal species and using practical habitat proxies (available in GIS). The output was designed as eleven threat-related habitat quality scores on the ordinal scale, which can be grouped qualitatively from non-habitat to quality habitat (Figure 2). The ‘ecological profiles’ were (Table 2): D1, a poorly dispersing perennial plant of gap-dynamic eutrophic forest, vulnerable to continuity disruption and dense shade in monoculture stands; D2, a poorly dispersing saproxylic species of natural Picea forests, which is vulnerable to the disrupted continuity of large downed trunks in moderate shade; D3, a rare species inhabiting senescent or dead Populus tremula in mid-succession, which is lost both in intensively managed forests and old-growth without P. tremula recruitment; D4, an epiphyte on old nemoral hardwood trees that are characteristic in natural forests (see above) but suppressed by production forestry; D5, an area-sensitive vertebrate in vertically well-structured stands, vulnerable to structural simplification and patch fragmentation; D6, a terrestrial invertebrate on stable moist ground that suffers from stand-continuity loss and unpalatable litter of forestry-favored conifer and Betula trees; D7, a (semi)-aquatic species of small forest streams, which is threatened by loss of microhabitats due to dredging of stream channel and upstream pollution from agriculture and drainage systems.
We illustrated the mapping approach by predicted changes from 10-years past (2009; based on real data) to current (2019) and 10-years future (2029; predicted by individual variables of the decision trees; Figure 2). We selected these relatively short symmetric time-frames here to assess both the delayed establishing of the reserves (compared to identifying their necessity) and planning and implementing a restoration program (compared to natural succession). Such short time frames also allow us to use a simplified approach to model uncertainty estimation. We ran the decision trees on openly available GIS sources and some critical elements digitalized for this project (notably forest continuity and stream channel changes from historical topographic maps). The basic spatial unit was the forest stand, as defined in the national forest registry. However, since other spatial data divided stands and subsequent forest surveys changed their borders, we performed areal calculations by rasterizing the maps (20 m grid). Certain subjective decisions were made (e.g., we applied the effect of thinning in a 20-year time frame), but the ordinal scale used appeared relatively robust to that. We used QGIS 3.10.2 [177] and R packages, dplyr [178] and sf [179], for the spatial analyses.
We found that (i) although the areas had degraded age and tree-species structure (see above), the reserve selection had been generally successful. Net habitat loss in the last 10 years was only apparent for the perennial plant (D1); it will recover in a decade (Table 2). (ii) Regarding restoration potential, some popular stand-structural targets of active forest restoration (diversification of stand structure; dead wood creation) [162] were likely to be met at reasonable rates also by protection (Table 2; Figure 3b–d). (iii) In contrast, the pronounced lack of old nemoral hardwood trees for D4 will not be healed (Table 2). Since it cannot be rapidly addressed by restoration, too, another perspective is needed—perhaps protecting residual trees in the surrounding landscapes for the long term [200,201,202]. The same factor has degraded the habitat of litter-dwelling invertebrates (D6), but we expect their habitat quality to recover sooner along with undergrowth development. (iv) Another issue revealed was that even though 10% of the area was currently non-habitat for every focal species defined, we expect considerable passive recovery from that status (Figure 3e). Even fewer of such universally degraded stands appear concentrated enough to allow cost-efficient restoration. (v) The forest area containing quality habitats for lotic invertebrates is very small and, thus, potentially vulnerable to occasional disturbance. To sustain this part of biodiversity, we need a better basic understanding of its functioning in degraded forests and in relation to protection regimes.
Uncertainty of our models contains three major components. First, the priority order of the variables (sequence of decision nodes), which can be assessed by field-checking alternative decision trees. Second, parameter values at decision nodes to be analyzed for sensitivity. Third, parameter accuracy in the GIS sources that can be addressed by combining different sources. To exemplify, we report sensitivity of two models to high-priority nodes of tree age: ‘oldest trees >60 yr’ in D2 (Figure 2b) and four classes of ‘mean tree age’ in D5 (Figure 2d). Model projections for these threshold values changed by ±5 years did not yield abrupt changes in habitat quality distributions (quality habitat areas were not affected at all in D2). Trends were least sensitive: only one projection was affected by >1% percent point. Thus, a +5-yr threshold in D2 predicted a 10% decrease in non-habitat by 2029 instead of a 7% decrease at the original threshold and 6% for the 5-yr threshold.
4. Discussion
Within 30 years, species distribution and habitat modeling for forest management and conservation have much developed technically, but such models have not become mainstream in actual planning. There are probably several reasons for that, as outlined below. Our broad conclusion, however, is that the biodiversity criterion of SFM (and other land use) cannot be met without simplifying and visualizing the living environment in both ecologically and socially relevant terms. Despite fair criticism against the misuse of the focal species concept [56,77,78,203,204], there are no clear alternatives for making non-human perspectives of the environment and its impacts meaningful for stakeholders and the wider public. Carefully prepared maps integrate many technical and cultural tools for such perspectives, including spatial modeling as a major technique.
A detailed treatment of the problems with the surrogacy value of focal species is beyond the scope of this paper (see [56]), but we note that the concept seems to have suffered from two frequent misconceptions. First, that it is primarily about species [77,78,204] and, second, that the concept prescribes its prevailing practice to highlight vertebrates [205,206]. Although Lambeck [54] provided vertebrate examples, his main idea was to operationalize threats for conservation action systematically. Our case study demonstrated how the diversity of forest taxa can be considered for threat mapping, following from our long-term study of multi-taxon solutions for SFM [207,208]. In analyses on lichenized and wood-inhabiting fungi, we have concluded that focal-species criteria can be met in ca. 2% of the Estonian forest species pool [92,93]. This points at an order of magnitude of a few hundred focal species to be considered for forestry in this country. There is indeed scarce information on most of these species when compared with birds and other vertebrates (and it is likely to remain so), but the question for SFM and conservation is whether the existing information can be organized for the decision-making processes. Specifically, it may not be feasible to monitor all these species in the field, but spatial analysis of their habitats and perspectives using deductive models may be realistic. That, again, does not mean that focal species should be the sole tools to address threats or that they can be used everywhere. For example, useful tools for biologically mega-diverse tropical regions or for indigenous people are probably distinct [209]. However, as a principle, we maintain that non-human species have some integrated and cultural qualities that are not present in the alternatives of measuring physicochemical environments, functional traits in the assemblages, composition, or structure of ecosystems, or their ‘services’ to humans [38,63,210,211,212]. Such differences may deepen when it comes to society acting for a change.
Coming to the question of how can spatial analysis help to elaborate and represent meaningful information on selected species in SFM, we identified four key issues for future research and development. Perhaps the crucial one is the research on social value of spatial mapping tools for different decision-making processes. We doubt that focal-species models could contribute effectively to the highly formalized field of spatial economic optimization for multi-purpose forestry planning [23], due to general problems with the ‘currency’ of biodiversity and measurement error [213,214]. Focal-species models are probably more effective as heuristic tools for political processes, when ‘windows of opportunity’ shift attention on environmental threats [215]. Our review indicated several enlightening models developed for such policy processes (Table 1), but, in general, there seems to be much unused potential. Noting pronounced problems with uncertainties in biodiversity models [213,216], we emphasize that these analyses make sense in the context of particular decisions. For example, our models initially aimed to locate stands for active restoration for biodiversity, but the results changed the perspective to whether such restoration is feasible overall, and for what purpose. Thus, spatial precision became less an issue compared with field-checking of the qualitative predictions of general habitat availability for different species.
A related issue is a lack of focal-species models on some important ecological dimensions, notably the biodiversity in forest soils and water bodies [217]. We included these dimensions in our model set, although we faced difficulties with obtaining both species’ natural-history information and relevant GIS data. A reason for the former is that the environmental impact research in freshwater and soil domains has traditionally focused on assemblages and taxon groups (including functional groups), not individual species [218,219,220]. Ecological analyses might reconsider whether group-level treatment is always justified for management [218,221,222], given that red-listing of species—an important part of the focal-species scheme (Figure 1)—has become an accepted formal tool. Inconspicuous species are gaining official protection in those jurisdictions that prioritize conservation status over public awareness. Similarly, protecting rare and threatened species is required by forest management certification systems, such as by the Forest Stewardship Council (FSC) [223]. Even from an awareness perspective, it is not self-evident that invertebrate species would be ignored by stakeholders. For example, researchers working through media have succeeded in making a long list of ‘primeval forest relict beetle’ species and their locations attractive for the wider public in Germany [224]. Regarding the GIS data, remote sensing of soil conditions and small water-bodies in biodiversity-relevant terms can probably be better addressed using technological advances already available.
The third issue concerns mapping units and spatial hierarchies, with the basic distinction and different practices of gradient- and patch-based models [225]. We found these approaches complementary and used these in different phases of the analysis. However, it is indeed a caveat of most simple habitat models that they neglect the issues of population structure and viability, although these are primary issues when considering environmental threats to a species. An unanswered question is how much population-specific detail is appropriate to still retain the heuristic value of the model for broad questions.
Finally, we highlight that deductive models based on theoretical understanding are most useful for predicting into the (largely unknown) future. Regarding the present, inductive (empirical) models probably outperform deductive models in spatial prediction, and actual measurements of forest conditions may be even more reliable. This means that ecologists developing decision support for SFM and other land use in human-influenced ecosystems should combine approaches [216]. For modeling, a better understanding of the drivers of future change is necessary. Our review of the literature indicated that studies tend to predict the long-term future by simply extending the current social and economic context and legislature for many decades. Collaboration with social scientists of futures studies might help forest ecologists to understand better how to compile useful spatial long-term scenarios of land use [226,227].
Author Contributions
Conceptualization, A.L.; methodology, A.L., L.R., M.V., and K.R.; formal analysis, R.K. and A.L.; investigation, A.L., R.K., L.R., M.V., and K.R.; data curation, R.K.; writing—original draft preparation, A.L.; writing—review and editing, L.R. and K.R.; visualization, A.L.; project administration, A.L. and M.V.; funding acquisition, A.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Estonian Research Council (grant IUT 34-7) and the Estonian Center for Environmental Investments (grant 16288).
Acknowledgments
We are grateful to the species experts who assisted in defining focal species for the case study, notably Piret Lõhmus, Anneli Palo, and Kai Vellak. The Estonian Environmental Agency kindly provided access to the archives of the national forest registry for the modeling input. We thank the Editors of the Special Issue for inviting this review, and two anonymous reviewers for constructive comments on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Clayton, T.; Radcliffe, N. Sustainability: A Systems Approach; Routledge: Abingdon, UK, 2018. [Google Scholar]
- Filotas, E.; Parrott, L.; Burton, P.J.; Chazdon, R.L.; Coates, K.D.; Coll, L.; Haeussler, S.; Martin, K.; Nocentini, S.; Puettmann, K.J.; et al. Viewing forests through the lens of complex systems science. Ecosphere 2014, 5, 1–23. [Google Scholar] [CrossRef]
- Messier, C.; Puettmann, K.; Chazdon, R.; Andersson, K.P.; Angers, V.A.; Brotons, L.; Filotas, E.; Tittler, R.; Parrott, L.; Levin, S.A. From management to stewardship: Viewing forests as complex adaptive systems in an uncertain world. Conserv. Lett. 2015, 8, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Burton, P.J. The scope and challenge of sustainable forestry. In Achieving Sustainable Management of Boreal and Temperate Forests; Stanturf, J.A., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2020; pp. 1–22. [Google Scholar]
- Folke, C.; Holling, C.S.; Perrings, C. Biological diversity, ecosystems, and the human scale. Ecol. Appl. 1996, 6, 1018–1024. [Google Scholar] [CrossRef] [Green Version]
- Mace, G.M.; Norris, K.; Fitter, A.H. Biodiversity and ecosystem services: A multilayered relationship. Trends Ecol. Evol. 2012, 27, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Oliver, T.H.; Heard, M.S.; Isaac, N.J.; Roy, D.B.; Procter, D.; Eigenbrod, F.; Freckleton, R.; Hector, A.; Orme, C.D.L.; Petchey, O.L.; et al. Biodiversity and resilience of ecosystem functions. Trends Ecol. Evol. 2015, 30, 673–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angeler, D.G.; Fried-Petersen, H.; Allen, C.R.; Garmestani, A.; Twidwell, D.; Chuang, W.; Donovan, V.M.; Eason, T.; Roberts, C.P.; Sundstrom, S.M.; et al. Adaptive capacity in ecosystems. Adv. Ecol. Res. 2019, 60, 1–24. [Google Scholar] [PubMed]
- Baho, D.L.; Allen, C.R.; Garmestani, A.S.; Fried-Petersen, H.B.; Renes, S.E.; Gunderson, L.H.; Angeler, D.G. A quantitative framework for assessing ecological resilience. Ecol. Soc. 2017, 22, 17. [Google Scholar] [CrossRef] [Green Version]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.B.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef] [Green Version]
- Mori, A.S.; Lertzman, K.P.; Gustafsson, L. Biodiversity and ecosystem services in forest ecosystems: A research agenda for applied forest ecology. J. Appl. Ecol. 2017, 54, 12–27. [Google Scholar] [CrossRef]
- Linser, S.; Wolfslehner, B.; Bridge, S.R.; Gritten, D.; Johnson, S.; Payn, T.; Prins, K.; Raši, R.; Robertson, G. 25 years of criteria and indicators for sustainable forest management: How intergovernmental C&I processes have made a difference. Forests 2018, 9, 578. [Google Scholar]
- Miller, D.C.; Nakamura, K.S. Protected areas and the sustainable governance of forest resources. Curr. Opin. Environ. Sustain. 2018, 32, 96–103. [Google Scholar] [CrossRef]
- Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Global Assessment Report on Biodiversity and Ecosystem Services; IPBES: Bonn, Germany, 2019. [Google Scholar]
- Turner, M.G.; Arthaud, G.J.; Engstrom, R.T.; Hejl, S.J.; Liu, J.; Loeb, S.; McKelvey, K. Usefulness of spatially explicit population models in land management. Ecol. Appl. 1995, 5, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Baskent, E.Z.; Keles, S. Spatial forest planning: A review. Ecol. Model. 2005, 188, 145–173. [Google Scholar] [CrossRef]
- Baskent, E.Z. Forest landscape modelling as a tool to develop conservation targets. In Setting Conservation Targets for Managed Forest Landscapes; Villard, M.A., Jonsson, B.G., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 304–327. [Google Scholar]
- Dijak, W.D.; Rittenhouse, C.D. Development and application of habitat suitability models to large landscapes. In Models for Planning Wildlife Conservation in Large Landscapes; Millspaugh, J.J., Thompson, F.R., III, Eds.; Elsevier, Academic Press: Burlington, MA, USA, 2009; pp. 367–389. [Google Scholar]
- Kimmins, H.; Blanco, J.A.; Seely, B.; Welham, C.; Scoullar, K. Forecasting Forest Futures: A Hybrid Modelling Approach to the Assessment of Sustainability of Forest Ecosystems and Their Values; Earthscan: London, UK; New York, NY, USA, 2010. [Google Scholar]
- Snäll, T.; Lehtomäki, J.; Arponen, A.; Elith, J.; Moilanen, A. Green infrastructure design based on spatial conservation prioritization and modeling of biodiversity features and ecosystem services. Environ. Manag. 2016, 57, 251–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, J.M.; Heglund, P.J.; Morrison, M.L.; Haufler, J.B.; Raphael, M.G.; Wall, W.A.; Samson, F.B. (Eds.) Predicting Species Occurrence: Issues of Accuracy and Scale; Island Press: Washington, DC, USA, 2002. [Google Scholar]
- Corona, P.; Köhl, M.; Marchetti, M. (Eds.) Advances in Forest Inventory for Sustainable Forest Management and Biodiversity Monitoring; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003. [Google Scholar]
- Pukkala, T. Measuring non-wood forest outputs in numerical forest planning. In Multi-Objective Forest Planning; Pukkala, T., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 173–207. [Google Scholar]
- Bush, A.; Sollmann, R.; Wilting, A.; Bohmann, K.; Cole, B.; Balzter, H.; Martius, C.; Zlinszky, A.; Calvignac-Spencer, S.; Cobbold, C.A.; et al. Connecting Earth observation to high-throughput biodiversity data. Nat. Ecol. Evol. 2017, 1, 0176. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Levick, S.R.; Heidrich, L.; Magdon, P.; Leutner, B.F.; Wöllauer, S.; Serebryanyk, A.; Nauss, T.; Krzystek, P.; Gossner, M.M.; et al. Radar vision in the mapping of forest biodiversity from space. Nat. Commun. 2019, 10, 4757. [Google Scholar] [CrossRef]
- Randin, C.F.; Ashcroft, M.B.; Bolliger, J.; Cavender-Bares, J.; Coops, N.C.; Dullinger, S.; Dirnböck, T.; Eckert, S.; Ellis, E.; Fernández, N.; et al. Monitoring biodiversity in the Anthropocene using remote sensing in species distribution models. Remote Sens. Environ. 2020, 239, 111626. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Franklin, J.F.; Fischer, J. General management principles and a checklist of strategies to guide forest biodiversity conservation. Biol. Conserv. 2006, 131, 433–445. [Google Scholar] [CrossRef]
- Schulte, L.A.; Mitchell, R.J.; Hunter, M.L., Jr.; Franklin, J.F.; McIntyre, R.K.; Palik, B.J. Evaluating the conceptual tools for forest biodiversity conservation and their implementation in the US. For. Ecol. Manag. 2006, 232, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chertov, O.; Komarov, A.; Mikhailov, A.; Andrienko, G.; Andrienko, N.; Gatalsky, P. Geovisualization of forest simulation modelling results: A case study of carbon sequestration and biodiversity. Comput. Electron. Agric. 2005, 49, 175–191. [Google Scholar] [CrossRef]
- Mäkelä, A.; del Río, M.; Hynynen, J.; Hawkins, M.J.; Reyer, C.; Soares, P.; van Oijen, M.; Tomé, M. Using stand-scale forest models for estimating indicators of sustainable forest management. For. Ecol. Manag. 2012, 285, 164–178. [Google Scholar] [CrossRef] [Green Version]
- Forest Europe. State of Europe’s Forests 2015; Ministerial Conference on the Protection of Forests in Europe: Madrid, Spain, 2015. [Google Scholar]
- Winkel, G.; Blondet, M.; Borrass, L.; Frei, T.; Geitzenauer, M.; Gruppe, A.; Jump, A.; De Koning, J.; Sotirov, M.; Weiss, G.; et al. The implementation of Natura 2000 in forests: A trans- and interdisciplinary assessment of challenges and choices. Environ. Sci. Policy 2015, 52, 23–32. [Google Scholar] [CrossRef]
- Kovač, M.; Kutnar, L.; Hladnik, D. Assessing biodiversity and conservation status of the Natura 2000 forest habitat types: Tools for designated forestlands stewardship. For. Ecol. Manag. 2016, 359, 256–267. [Google Scholar] [CrossRef]
- Thakur, A.K.; Kumar, R.; Verma, R.K. Analysing India’s current national forest inventory for biodiversity information. Biodivers. Conserv. 2018, 27, 3049–3069. [Google Scholar] [CrossRef]
- Criteria and Indicators of Sustainable Forest Management in Canada: National Status 2005; Canadian Council of Forest Ministers; Canadian Forest Service: Ottawa, ON, Canada, 2006.
- Robertson, G.; Gualke, P.; McWilliams, R.; LaPlante, S.; Guldin, R. (Eds.) National Report on Sustainable Forests—2010; Report FS-979; USDA Forest Service: Washington, DC, USA, 2011.
- Timonen, J.; Siitonen, J.; Gustafsson, L.; Kotiaho, J.S.; Stokland, J.N.; Sverdrup-Thygeson, A.; Mönkkönen, M. Woodland key habitats in northern Europe: Concepts, inventory and protection. Scand. J. For. Res. 2010, 25, 309–324. [Google Scholar] [CrossRef] [Green Version]
- Noss, R.F. Assessing and monitoring forest biodiversity: A suggested framework and indicators. For. Ecol. Manag. 1999, 115, 135–146. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Margules, C.R.; Botkin, D.B. Indicators of biodiversity for ecologically sustainable forest management. Conserv. Biol. 2000, 14, 941–950. [Google Scholar] [CrossRef]
- Blattert, C.; Lemm, R.; Thees, O.; Lexer, M.J.; Hanewinkel, M. Management of ecosystem services in mountain forests: Review of indicators and value functions for model based multi-criteria decision analysis. Ecol. Indic. 2017, 79, 391–409. [Google Scholar] [CrossRef]
- Habel, J.C.; Gossner, M.M.; Meyer, S.T.; Eggermont, H.; Lens, L.; Dengler, J.; Weisser, W.W. Mind the gaps when using science to address conservation concerns. Biodivers. Conserv. 2013, 22, 2413–2427. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Landuyt, D.; Perring, M.P.; Seidl, R.; Taubert, F.; Verbeeck, H.; Verheyen, K. Modelling understorey dynamics in temperate forests under global change – Challenges and perspectives. Perspect. Plant Ecol. Evol. Syst. 2018, 31, 44–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Biest, K.; Meire, P.; Schellekens, T.; D’hondt, B.; Bonte, D.; Vanagt, T.; Ysebaert, T. Aligning biodiversity conservation and ecosystem services in spatial planning: Focus on ecosystem processes. Sci. Total Environ. 2020, 712, 136350. [Google Scholar] [CrossRef] [PubMed]
- Blattert, C.; Lemm, R.; Thees, O.; Hansen, J.; Lexer, M.J.; Hanewinkel, M. Segregated versus integrated biodiversity conservation: Value-based ecosystem service assessment under varying forest management strategies in a Swiss case study. Ecol. Indic. 2018, 95, 751–764. [Google Scholar] [CrossRef]
- Moilanen, A. Landscape zonation, benefit functions and target-based planning: Unifying reserve selection strategies. Biol. Conserv. 2007, 134, 571–579. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Guariguata, M.R. Decision Support Tools for Forest Landscape Restoration: Current Status and Future Outlook; Center for International Forest Research: Bogor, Indonesia, 2018. [Google Scholar]
- Map of Biodiversity Importance. Available online: https://www.natureserve.org/conservation-tools/projects/map-biodiversity-importance (accessed on 20 June 2020).
- Brown, D.J.; Ribic, C.A.; Donner, D.M.; Nelson, M.D.; Bocetti, C.I.; Deloria-Sheffield, C.M. Using a full annual cycle model to evaluate long-term population viability of the conservation-reliant Kirtland’s warbler after successful recovery. J. Appl. Ecol. 2017, 54, 439–449. [Google Scholar] [CrossRef]
- Cash, D.W.; Clark, W.C.; Alcock, F.; Dickson, N.M.; Eckley, N.; Guston, D.H.; Jäger, J.; Mitchell, R.B. Knowledge systems for sustainable development. Proc. Natl. Acad. Sci. USA 2003, 100, 8086–8091. [Google Scholar] [CrossRef] [Green Version]
- Bertuol-Garcia, D.; Morsello, C.; El-Hani, C.N.; Pardini, R. A conceptual framework for understanding the perspectives on the causes of the science–practice gap in ecology and conservation. Biol. Rev. 2018, 93, 1032–1055. [Google Scholar] [CrossRef]
- Failing, L.; Gregory, R. Ten common mistakes in designing biodiversity indicators for forest policy. J. Environ. Manag. 2003, 68, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Quarshie, A.; Salmi, A.; Wu, Z. From equivocality to reflexivity in biodiversity protection. Organ. Environ. in press. [CrossRef]
- Lambeck, R.J. Focal species: A multi-species umbrella for nature conservation. Conserv. Biol. 1997, 11, 849–856. [Google Scholar] [CrossRef] [Green Version]
- Lambeck, R.J. Focal species and restoration ecology: Response to Lindenmayer et al. Conserv. Biol. 2002, 16, 549–551. [Google Scholar] [CrossRef]
- Caro, T. Conservation by Proxy: Indicator, Umbrella, Keystone, Flagship, and Other Surrogate Species; Island Press: Washington, DC, USA, 2010. [Google Scholar]
- Angelstam, P.; Persson, R.; Schlaepfer, R. The sustainable forest management vision and biodiversity: Barriers and bridges for implementation in actual landscapes. Ecol. Bull. 2004, 51, 29–49. [Google Scholar]
- Lindenmayer, D.B.; Cunningham, S.A. Six principles for managing forests as ecologically sustainable ecosystems. Landsc. Ecol. 2013, 28, 1099–1110. [Google Scholar] [CrossRef]
- Michanek, G.; Bostedt, G.; Ekvall, H.; Forsberg, M.; Hof, A.R.; De Jong, J.; Rudolphi, J.; Zabel, A. Landscape planning—Paving the way for effective conservation of forest biodiversity and a diverse forestry? Forests 2018, 9, 523. [Google Scholar] [CrossRef] [Green Version]
- Hahn, W.A.; Knoke, T. Sustainable development and sustainable forestry: Analogies, differences, and the role of flexibility. Eur. J. For. Res. 2010, 129, 787–801. [Google Scholar] [CrossRef]
- IUCN Standards and Petitions Committee. Guidelines for Using the IUCN Red List Categories and Criteria. Version 14; IUCN: Gland, Switzerland, 2019; Available online: http://www.iucnredlist.org/documents/RedListGuidelines.pdf (accessed on 19 May 2020).
- Brooks, T.M.; Pimm, S.L.; Akçakaya, H.R.; Buchanan, G.M.; Butchart, S.H.; Foden, W.; Hilton-Taylor, C.; Hoffmann, M.; Jenkins, C.N.; Joppa, L.; et al. Measuring terrestrial area of habitat (AOH) and its utility for the IUCN red list. Trends Ecol. Evol. 2019, 34, 977–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, M., Jr.; Westgate, M.; Barton, P.; Calhoun, A.; Pierson, J.; Tulloch, A.; Beger, M.; Branquinho, C.; Caro, T.; Gross, J.; et al. Two roles for ecological surrogacy: Indicator surrogates and management surrogates. Ecol. Indic. 2016, 63, 121–125. [Google Scholar] [CrossRef]
- Bell, F.W.; Dacosta, J.; Larocque, G.R. Considering forest biodiversity indicators within a pressure, state, benefit, and response framework. In Ecological Forest Management Handbook; Larocque, G.R., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 337–359. [Google Scholar]
- Sutherland, W.J.; Bailey, M.J.; Bainbridge, I.P.; Brereton, T.; Dick, J.T.; Drewitt, J.; Dulvy, N.K.; Dusic, N.R.; Freckleton, R.P.; Gaston, K.J.; et al. Future novel threats and opportunities facing UK biodiversity identified by horizon scanning. J. Appl. Ecol. 2008, 45, 821–833. [Google Scholar] [CrossRef]
- Hines, A.; Bengston, D.N.; Dockry, M.J.; Cowart, A. Setting up a horizon scanning system: A US federal agency example. World Futures Rev. 2018, 10, 136–151. [Google Scholar] [CrossRef]
- Beazley, K.; Cardinal, N. A systematic approach for selecting focal species for conservation in the forests of Nova Scotia and Maine. Environ. Conserv. 2004, 31, 91–101. [Google Scholar] [CrossRef]
- Lõhmus, A.; Leivits, M.; Pēterhofs, E.; Zizas, R.; Hofmanis, H.; Ojaste, I.; Kurlavičius, P. The Capercaillie (Tetrao urogallus): An iconic focal species for knowledge-based integrative management and conservation of Baltic forests. Biodivers. Conserv. 2017, 26, 1–21. [Google Scholar] [CrossRef]
- Ward, M.; Rhodes, J.R.; Watson, J.E.; Lefevre, J.; Atkinson, S.; Possingham, H.P. Use of surrogate species to cost-effectively prioritize conservation actions. Conserv. Biol. 2020, 34, 600–610. [Google Scholar] [CrossRef]
- Peterson, A.T. Ecological niche conservatism: A time-structured review of evidence. J. Biogeogr. 2011, 38, 817–827. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models with Applications in R; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Hanski, I. Extinction debt and species credit in boreal forests: Modelling the consequences of different approaches to biodiversity conservation. Ann. Zool. Fenn. 2000, 37, 271–280. [Google Scholar]
- Vellend, M.; Verheyen, K.; Jacquemyn, H.; Kolb, A.; Van Calster, H.; Peterken, G.; Hermy, M. Extinction debt of forest plants persists for more than a century following habitat fragmentation. Ecology 2006, 87, 542–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinclair, S.J.; White, M.D.; Newell, G.R. How useful are species distribution models for managing biodiversity under future climates? Ecol. Soc. 2010, 15, 8. [Google Scholar] [CrossRef]
- Raphael, M.G.; Wisdom, M.J.; Rowland, M.M.; Holthausen, R.S.; Wales, B.C.; Marcot, B.G.; Rich, T.D. Status and trends of habitats of terrestrial vertebrates in relation to land management in the interior Columbia river basin. For. Ecol. Manag. 2001, 153, 63–87. [Google Scholar] [CrossRef]
- Reza, M.I.H.; Abdullah, S.A.; Nor, S.B.M.; Ismail, M.H. Integrating GIS and expert judgment in a multi-criteria analysis to map and develop a habitat suitability index: A case study of large mammals on the Malayan Peninsula. Ecol. Indic. 2013, 34, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Lindenmayer, D.B.; Manning, A.D.; Smith, P.L.; Possingham, H.P.; Fischer, J.; Oliver, I.; McCarthy, M.A. The focal-species approach and landscape restoration: A critique. Conserv. Biol. 2002, 16, 338–345. [Google Scholar] [CrossRef] [Green Version]
- Sætersdal, M.; Gjerde, I. Prioritising conservation areas using species surrogate measures: Consistent with ecological theory? J. Appl. Ecol. 2011, 48, 1236–1240. [Google Scholar] [CrossRef]
- Beresford, A.E.; Donald, P.F.; Buchanan, G.M. Repeatable and standardised monitoring of threats to Key Biodiversity Areas in Africa using Google Earth Engine. Ecol. Indic. 2020, 109, 105763. [Google Scholar] [CrossRef]
- Ortega Adarme, M.; Feitosa, R.Q.; Happ, N.P.; De Almeida, C.A.; Rodrigues Gomes, A. Evaluation of deep learning techniques for deforestation detection in the Brazilian Amazon and cerrado biomes from remote sensing imagery. Remote Sens. 2020, 12, 910. [Google Scholar] [CrossRef] [Green Version]
- Loveridge, A.J.; Sousa, L.L.; Seymour-Smith, J.; Hunt, J.; Coals, P.; O’Donnell, H.; Lindsey, P.A.; Mandisodza-Chikerema, R.; Macdonald, D.W. Evaluating the spatial intensity and demographic impacts of wire-snare bush-meat poaching on large carnivores. Biol. Conserv. 2020, 244, 108504. [Google Scholar] [CrossRef]
- Tulloch, V.J.D.; Tulloch, A.I.T.; Visconti, P.; Halpern, B.S.; Watson, J.E.M.; Evans, M.C.; Auerbach, N.A.; Barnes, M.; Beger, M.; Chadès, I.; et al. Why do we map threats? Linking threat mapping with actions to make better conservation decisions. Front. Ecol. Environ. 2015, 13, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Schamberger, M.; Krohn, W.B. Status of the habitat evaluation procedures. In Transactions of the 47th North Americal Wildlife and Natural Resources Conference; Sabol, K., Ed.; Wildlife Management Institute: Washington, DC, USA, 1982; pp. 154–164. [Google Scholar]
- Burley, J.B. Habitat suitability models: A tool for designing landscape for wildlife. Landsc. Res. 1989, 14, 23–26. [Google Scholar] [CrossRef]
- Noon, B.R.; McKelvey, K.S. Management of the spotted owl: A case history in conservation biology. Annu. Rev. Ecol. Syst. 1996, 27, 135–162. [Google Scholar] [CrossRef]
- Graul, W.D.; Torres, J.; Denney, R. A species-ecosystem approach for nongame programs. Wildl. Soc. Bull. 1976, 4, 79–80. [Google Scholar]
- Graul, W.D.; Miller, G.C. Strengthening ecosystem management approaches. Wildl. Soc. Bull. 1984, 12, 282–289. [Google Scholar]
- Mealy, S.P.; Horn, J.R. Integrating wildlife habitat objectives into the forest plan. In Transactions of the 46th North Americal Wildlife and Natural Resources Conference; Wildlife Management Institute: Washington, DC, USA, 1981; pp. 488–500. [Google Scholar]
- Edenius, L.; Mikusiński, G. Utility of habitat suitability models as biodiversity assessment tools in forest management. Scand. J. For. Res. 2006, S7, 62–72. [Google Scholar] [CrossRef]
- Halme, P.; Holec, J.; Heilmann-Clausen, J. The history and future of fungi as biodiversity surrogates in forests. Fungal Ecol. 2017, 27, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Lõhmus, A.; Vunk, E.; Runnel, K. Conservation management for forest fungi in Estonia: The case of polypores. Folia Cryptog. Estonica 2018, 55, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Runnel, K.; Lõhmus, A. Deadwood-rich managed forests provide insights into the old-forest association of wood-inhabiting fungi. Fungal Ecol. 2017, 27, 155–167. [Google Scholar] [CrossRef]
- Lõhmus, P.; Lõhmus, A. The potential of production forests for sustaining lichen diversity: A perspective on sustainable forest management. Forests 2019, 10, 1063. [Google Scholar] [CrossRef] [Green Version]
- Uliczka, H.; Angelstam, P.; Roberge, J.-M. Indicator species and biodiversity monitoring systems for non-industrial private forest owners - is there a communication problem? Ecol. Bull. 2004, 51, 379–384. [Google Scholar]
- Henrikson, L.; Arvidsson, B.; Österling, M. Aquatic Conservation with Focus on Margaritifera Margaritifera; Karlstads University: Karlstad, Sweden, 2012. [Google Scholar]
- Angelstam, P.; Törnblom, J.; Degerman, E.; Henrikson, L.; Jougda, L.; Lazdinis, M.; Malmgren, J.C.; Myhrman, L. From forest patches to functional habitat. In Farming, Forestry and the Natural Heritage: Towards a More Integrated Future; Davison, R., Garblraith, C.A., Eds.; Scottish Natural Heritage: Edinburgh, UK, 2006; pp. 193–209. [Google Scholar]
- Suislepp, K.; Rannap, R.; Lõhmus, A. Impacts of artificial drainage on amphibian breeding sites in hemiboreal forests. For. Ecol. Manag. 2011, 262, 1078–1083. [Google Scholar] [CrossRef]
- Mattsson, B.J.; Cooper, R.J. Louisiana waterthrushes (Seiurus motacilla) and habitat assessments as cost-effective indicators of instream biotic integrity. Freshw. Biol. 2006, 51, 1941–1958. [Google Scholar] [CrossRef]
- Dobler, A.H.; Geist, J.; Stoeckl, K.; Inoue, K. A spatially explicit approach to prioritize protection areas for endangered freshwater mussels. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Spies, T.A.; McComb, B.C.; Kennedy, R.S.H.; McGrath, M.T.; Olsen, K.; Pabst, R.J. Potential effects of forest policies on terrestrial biodiversity in a multi-ownership province. Ecol. Appl. 2007, 17, 48–65. [Google Scholar] [CrossRef]
- McComb, B.C.; Spies, T.A.; Olsen, K.A. Sustaining biodiversity in the Oregon Coast Range: Potential effects of forest policies in a multi-ownership province. Ecol. Soc. 2007, 12, 29. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, J.; Ray, D.; Brown, T.; Stone, D.; Watts, K.; Anderson, R. Using focal species modelling to evaluate the impact of land use change on forest and other habitat networks in western oceanic landscapes. Forestry 2009, 82, 119–134. [Google Scholar] [CrossRef] [Green Version]
- Wilhere, G.F.; Linders, M.J.; Cosentino, B.L. Defining alternative futures and projecting their effects on the spatial distribution of wildlife habitats. Landsc. Urban Plan. 2007, 79, 385–400. [Google Scholar] [CrossRef]
- Tarr, N.M.; Rubino, M.J.; Costanza, J.K.; McKerrow, A.J.; Collazo, J.A.; Abt, R.C. Projected gains and losses of wildlife habitat from bioenergy-induced landscape change. GCB Bioenergy 2017, 9, 909–923. [Google Scholar] [CrossRef]
- Marzluff, J.M.; Millspaugh, J.J.; Ceder, K.R.; Oliver, C.D.; Withey, J.; McCarter, J.B.; Mason, C.L.; Comnick, J. Modeling changes in wildlife habitat and timber revenues in response to forest management. For. Sci. 2002, 48, 191–202. [Google Scholar]
- Tikkanen, O.P.; Heinonen, T.; Kouki, J.; Matero, J. Habitat suitability models of saproxylic red-listed boreal forest species in long-term matrix management: Cost-effective measures for multi-species conservation. Biol. Conserv. 2007, 140, 359–372. [Google Scholar] [CrossRef]
- Kliskey, A.D.; Lofroth, E.C.; Thompson, W.A.; Brown, S.; Schreier, H. Simulating and evaluating alternative resource-use strategies using GIS-based habitat suitability indices. Landsc. Urban Plan. 1999, 45, 163–175. [Google Scholar] [CrossRef]
- Tremblay, J.A.; Boulanger, Y.; Cyr, D.; Taylor, A.R.; Price, D.T.; St-Laurent, M. Harvesting interacts with climate change to affect future habitat quality of a focal species in eastern Canada’s boreal forest. PLoS ONE 2018, 13, e0191645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haché, S.; Cameron, R.; Villard, M.-A.; Bayne, E.M.; MacLean, D.A. Demographic response of a neotropical migrant songbird to forest management and climate change scenarios. For. Ecol. Manag. 2016, 359, 309–320. [Google Scholar] [CrossRef]
- Roberge, J.M.; Öhman, K.; Lämås, T.; Felton, A.; Ranius, T.; Lundmark, T.; Nordin, A. Modified forest rotation lengths: Long-term effects on landscape-scale habitat availability for specialized species. J. Environ. Manag. 2018, 210, 1–9. [Google Scholar] [CrossRef]
- Bonnot, T.W.; Thompson, F.R., III; Millspaugh, J.J.; Jones-Farrand, D.T. Landscape-based population viability models demonstrate importance of strategic conservation planning for birds. Biol. Conserv. 2013, 165, 104–114. [Google Scholar] [CrossRef]
- Akçakaya, H.R.; Radeloff, V.C.; Mladenoff, D.J.; He, H.S. Integrating landscape and metapopulation modeling approaches: Viability of the sharp-tailed grouse in a dynamic landscape. Conserv. Biol. 2004, 18, 526–537. [Google Scholar] [CrossRef]
- Johansson, V.; Ranius, T.; Snäll, T. Epiphyte metapopulation persistence after drastic habitat decline and low tree regeneration: Time-lags and effects of conservation actions. J. Appl. Ecol. 2013, 50, 414–422. [Google Scholar] [CrossRef]
- Burnett, K.M.; Reeves, G.H.; Miller, D.J.; Clarke, S.; Vance-Borland, K.; Christiansen, K. Distribution of salmon-habitat potential relative to landscape characteristics and implications for conservation. Ecol. Appl. 2007, 17, 66–80. [Google Scholar] [CrossRef]
- Carroll, C.; Johnson, D.S. The importance of being spatial (and reserved): Assessing northern spotted owl habitat relationships with hierarchical bayesian models. Conserv. Biol. 2008, 22, 1026–1036. [Google Scholar] [CrossRef] [PubMed]
- Carroll, C. Role of climatic niche models in focal-species-based conservation planning: Assessing potential effects of climate change on northern spotted owl in the pacific northwest, USA. Biol. Conserv. 2010, 143, 1432–1437. [Google Scholar] [CrossRef]
- Carroll, C.; Dunk, J.R.; Moilanen, A. Optimizing resiliency of reserve networks to climate change: Multispecies conservation planning in the pacific northwest, USA. Glob. Chang. Biol. 2010, 16, 891–904. [Google Scholar] [CrossRef] [Green Version]
- Trisurat, Y.; Kanchanasaka, B.; Kreft, H. Assessing potential effects of land use and climate change on mammal distributions in northern Thailand. Wildl. Res. 2014, 41, 522–536. [Google Scholar] [CrossRef]
- Colyn, R.B.; Ehlers Smith, D.A.; Ehlers Smith, Y.C.; Smit-Robinson, H.; Downs, C.T. Predicted distributions of avian specialists: A framework for conservation of endangered forests under future climates. Divers. Distrib. 2020, 26, 652–667. [Google Scholar] [CrossRef]
- Carroll, C. Interacting effects of climate change, landscape conversion, and harvest on carnivore populations at the range margin: Marten and lynx in the northern Appalachians. Conserv. Biol. 2007, 21, 1092–1104. [Google Scholar] [CrossRef]
- Spies, T.A.; Johnson, K.N.; Burnett, K.M.; Ohmann, J.L.; McComb, B.C.; Reeves, G.H.; Bettinger, P.; Kline, J.D.; Garber-Yonts, B. Cumulative ecological and socioeconomic effects of forest policies in coastal Oregon. Ecol. Appl. 2007, 17, 5–17. [Google Scholar] [CrossRef]
- Mladenoff, D.J.; Host, G.E.; Boeder, J.; Crow, T.R. LANDIS: A spatial model of forest landscape disturbance, succession, and management. In GIS and Environmental Modeling: Progress and Research Ideas; Goodchild, M.F., Steyaert, L.T., Parks, B.O., Johnston, C., Maidment, D., Crane, M., Glendinning, S., Eds.; John Wiley & Sons: New York, NY, USA, 1996; pp. 175–179. [Google Scholar]
- Thompson, J.R.; Simons-Legaard, E.; Legaard, K.; Domingo, J.B. A LANDIS-II extension for incorporating land use and other disturbances. Environ. Model. Softw. 2016, 75, 202–205. [Google Scholar] [CrossRef]
- Sotirov, M.; Blum, M.; Storch, S.; Selter, A.; Schraml, U. Do forest policy actors learn through forward-thinking? Conflict and cooperation relating to the past, present and futures of sustainable forest management in Germany. For. Policy Econ. 2017, 85, 256–268. [Google Scholar] [CrossRef]
- Johnson, K.N.; Duncan, S.; Spies, T.A. Regional policy models for forest biodiversity analysis: Lessons from coastal Oregon. Ecol. Appl. 2017, 17, 81–90. [Google Scholar] [CrossRef]
- Kröger, M.; Raitio, K. Finnish forest policy in the era of bioeconomy: A pathway to sustainability? For. Policy Econ. 2017, 77, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Eyvindson, K.; Repo, A.; Mönkkönen, M. Mitigating forest biodiversity and ecosystem service losses in the era of bio-based economy. For. Policy Econ. 2018, 92, 119–127. [Google Scholar] [CrossRef]
- Lõhmus, A.; Fridolin, H.; Leivits, A.; Tõnisson, K.; Rannap, R. Prioritizing research gaps for national conservation management and policy: The managers’ perspective in Estonia. Biodivers. Conserv. 2019, 28, 2565–2579. [Google Scholar] [CrossRef]
- Noss, R.F. High-risk ecosystems as foci for considering biodiversity and ecological integrity in ecological risk assessments. Environ. Sci. Policy 2000, 3, 321–332. [Google Scholar] [CrossRef]
- European Environment Agency. Halting the loss of biodiversity by 2010: Proposal for a first set of indicators to monitor progress in Europe. EEA Tech. Rep. 2007, 11, 1–182. [Google Scholar]
- Levrel, H.; Kerbiriou, C.; Couvet, D.; Weber, J. OECD pressure–state–response indicators for managing biodiversity: A realistic perspective for a French biosphere reserve. Biodivers. Conserv. 2009, 18, 1719–1732. [Google Scholar] [CrossRef] [Green Version]
- Sparks, T.H.; Butchart, S.H.; Balmford, A.; Bennun, L.; Stanwell-Smith, D.; Walpole, M.; Bates, N.R.; Bomhard, B.; Buchanan, G.M.; Chenery, A.M.; et al. Linked indicator sets for addressing biodiversity loss. Oryx 2011, 45, 411–419. [Google Scholar] [CrossRef] [Green Version]
- Rempel, R.S.; Naylor, B.J.; Elkie, P.C.; Baker, J.; Churcher, J.; Gluck, M.J. An indicator system to assess ecological integrity of managed forests. Ecol. Indic. 2016, 60, 860–869. [Google Scholar] [CrossRef] [Green Version]
- Shahin, A.; Mahbod, M.A. Prioritization of key performance indicators: An integration of analytical hierarchy process and goal setting. Int. J. Oper. 2007, 56, 226–240. [Google Scholar] [CrossRef]
- Mönkkönen, M.; Reunanen, P.; Kotiaho, J.S.; Juutinen, A.; Tikkanen, O.P.; Kouki, J. Cost-effective strategies to conserve boreal forest biodiversity and long-term landscape-level maintenance of habitats. Eur. J. For. Res. 2011, 130, 717–727. [Google Scholar] [CrossRef]
- Albert, C.H.; Rayfield, B.; Dumitru, M.; Gonzalez, A. Applying network theory to prioritize multispecies habitat networks that are robust to climate and land-use change. Conserv. Biol. 2017, 31, 1383–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, K.; Eycott, A.E.; Handley, P.; Ray, D.; Humphrey, J.W.; Quine, C.P. Targeting and evaluating biodiversity conservation action within fragmented landscapes: An approach based on generic focal species and least-cost networks. Landsc. Ecol. 2010, 25, 1305–1318. [Google Scholar] [CrossRef]
- Johansson, V.; Felton, A.; Ranius, T. Long-term landscape scale effects of bioenergy extraction on dead wood-dependent species. For. Ecol. Manag. 2016, 371, 103–113. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Cressie, N.; Calder, C.A.; Clark, J.S.; Hoef, J.M.V.; Wikle, C.K. Accounting for uncertainty in ecological analysis: The strengths and limitations of hierarchical statistical modeling. Ecol. Appl. 2009, 19, 553–570. [Google Scholar] [CrossRef] [PubMed]
- Schumaker, N.H.; Brookes, A. HexSim: A modeling environment for ecology and conservation. Landsc. Ecol. 2018, 33, 197–211. [Google Scholar] [CrossRef]
- Rookwood, P. Landscape planning for biodiversity. Landsc. Urban Plan. 1995, 31, 379–385. [Google Scholar] [CrossRef]
- Theobald, D.M.; Hobbs, N.T.; Bearly, T.; Zack, J.A.; Shenk, T.; Riebsame, W.E. Incorporating biological information in local land-use decision making: Designing a system for conservation planning. Landsc. Ecol. 2000, 15, 35–45. [Google Scholar] [CrossRef]
- Brooks, R.P. Improving habitat suitability index models. Wildl. Soc. Bull. 1997, 25, 163–167. [Google Scholar]
- Zhu, M.J.; Hoctor, T.S.; Volk, M.; Frank, K.I.; Zwick, P.D.; Carr, M.H.; Linhoss, A.C. Spatial conservation prioritization to conserve biodiversity in response to sea level rise and land use change in the Matanzas River Basin, Northeast Florida. Landsc. Urban Plann. 2015, 144, 103–118. [Google Scholar] [CrossRef]
- Corona, P.; Chirici, G.; McRoberts, R.E.; Winter, S.; Barbati, A. Contribution of large-scale forest inventories to biodiversity assessment and monitoring. For. Ecol. Manag. 2011, 262, 2061–2069. [Google Scholar] [CrossRef] [Green Version]
- Rondeux, J.; Sanchez, C. Review of indicators and field methods for monitoring biodiversity within national forest inventories. Core variable: Deadwood. Environ. Monit. Assess. 2010, 164, 617–630. [Google Scholar] [CrossRef]
- Corona, P.; Franceschi, S.; Pisani, C.; Portoghesi, L.; Mattioli, W.; Fattorini, L. Inference on diversity from forest inventories: A review. Biodivers. Conserv. 2017, 26, 3037–3049. [Google Scholar] [CrossRef]
- Leslie, H.M.; Goldman, E.; Mcleod, K.L.; Sievanen, L.; Balasubramanian, H.; Cudney-Bueno, R.; Feuerstein, A.; Knowlton, N.; Lee, K.; Pollnac, R.; et al. How good science and stories can go hand-in-hand. Conserv. Biol. 2013, 27, 1126–1129. [Google Scholar] [CrossRef]
- McInerny, G.J.; Chen, M.; Freeman, R.; Gavaghan, D.; Meyer, M.; Rowland, F.; Spiegelhalter, D.J.; Stefaner, M.; Tessarolo, G.; Hortal, J. Information visualisation for science and policy: Engaging users and avoiding bias. Trends Ecol. Evol. 2014, 29, 148–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garibaldi, A.; Turner, N. Cultural keystone species: Implications for ecological conservation and restoration. Ecol. Soc. 2004, 9, 1. [Google Scholar] [CrossRef]
- Paschen, J.A.; Ison, R. Narrative research in climate change adaptation—Exploring a complementary paradigm for research and governance. Res. Policy 2014, 43, 1083–1092. [Google Scholar] [CrossRef]
- Nel, J.L.; Roux, D.J.; Driver, A.; Hill, L.; Maherry, A.C.; Snaddon, K.; Petersen, C.R.; Smith-Adao, L.B.; Van Deventer, H.; Reyers, B. Knowledge co-production and boundary work to promote implementation of conservation plans. Conserv. Biol. 2016, 30, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Gaston, K.J.; Jackson, S.F.; Cantu-Salazar, L.; Cruz-Piñón, G. The ecological performance of protected areas. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 93–113. [Google Scholar] [CrossRef] [Green Version]
- Hanski, I. Habitat loss, the dynamics of biodiversity, and a perspective on conservation. Ambio 2011, 40, 248–255. [Google Scholar] [CrossRef] [Green Version]
- Haavik, A.; Dale, S. Are reserves enough? Value of protected areas for boreal forest birds in southeastern Norway. Ann. Zool. Fenn. 2012, 49, 69–80. [Google Scholar] [CrossRef]
- Jones, H.P.; Schmitz, O.J. Rapid recovery of damaged ecosystems. PLoS ONE 2009, 4, e5653. [Google Scholar] [CrossRef]
- Vandekerkhove, K.; De Keersmaeker, L.; Walleyn, R.; Köhler, F.; Crevecoeur, L.; Govaere, L.; Thomaes, A.; Verheyen, K. Reappearance of old growth elements in lowland woodlands in northern Belgium: Do the associated species follow? Silva Fenn. 2012, 45, 909–935. [Google Scholar] [CrossRef] [Green Version]
- McLachlan, S.M.; Bazely, D.R. Recovery patterns of understory herbs and their use as indicators of deciduous forest regeneration. Conserv. Biol. 2001, 15, 98–110. [Google Scholar] [CrossRef]
- Bouget, C.; Parmain, G.; Gilg, O.; Noblecourt, T.; Nusillard, B.; Paillet, Y.; Pernot, C.; Larrieu, L.; Gosselin, F. Does a set-aside conservation strategy help the restoration of old-growth forest attributes and recolonization by saproxylic beetles? Anim. Conserv. 2014, 17, 342–353. [Google Scholar] [CrossRef]
- Runnel, K.; Sell, I.; Lõhmus, A. Recovery of the Critically Endangered bracket fungus Amylocystis lapponica in the Estonian network of strictly protected forests. Oryx 2019, in press. [Google Scholar] [CrossRef] [Green Version]
- Bernes, C.; Jonsson, B.G.; Junninen, K.; Lõhmus, A.; MacDonald, E.; Müller, J.; Sandström, J. What is the impact of active management on biodiversity in boreal and temperate forests set aside for conservation or restoration? A systematic map. Environ. Evid. 2015, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Meli, P.; Holl, K.D.; Benayas, J.M.R.; Jones, H.P.; Jones, P.C.; Montoya, D.; Mateos, D.M. A global review of past land use, climate, and active vs. passive restoration effects on forest recovery. PLoS ONE 2017, 12, e0171368. [Google Scholar] [CrossRef]
- Lõhmus, A.; Kohv, K.; Palo, A.; Viilma, K. Loss of old-growth, and the minimum need for strictly protected forests in Estonia. Ecol. Bull. 2004, 51, 401–411. [Google Scholar]
- Mazziotta, A.; Pouzols, F.M.; Mönkkönen, M.; Kotiaho, J.S.; Strandman, H.; Moilanen, A. Optimal conservation resource allocation under variable economic and ecological time discounting rates in boreal forest. J. Environ. Manag. 2016, 180, 366–374. [Google Scholar] [CrossRef]
- Kotiaho, J.S.; Mönkkönen, M. From a crisis discipline towards prognostic conservation practise: An argument for setting aside degraded habitats. Ann. Zool. Fenn. 2017, 54, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Laasimer, L. Vegetation of the Estonian S.S.R.; Valgus: Tallinn, Estonian, 1965. [Google Scholar]
- Noreika, N.; Helm, A.; Öpik, M.; Jairus, T.; Vasar, M.; Reier, Ü.; Kook, E.; Riibak, K.; Kasari, L.; Tullus, H.; et al. Forest biomass, soil and biodiversity relationships originate from biogeographic affinity and direct ecological effects. Oikos 2019, 128, 1653–1665. [Google Scholar] [CrossRef]
- Shorohova, E.; Kuuluvainen, T.; Kangur, A.; Jõgiste, K. Natural stand structures, disturbance regimes and successional dynamics in the Eurasian boreal forests: A review with special reference to Russian studies. Ann. For. Sci. 2009, 66, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Lõhmus, A.; Kraut, A. Stand structure of hemiboreal old-growth forests: Characteristic features, variation among site types, and a comparison with FSC-certified mature stands in Estonia. For. Ecol. Manag. 2010, 260, 155–165. [Google Scholar] [CrossRef]
- Kuuluvainen, T. Forest management and biodiversity conservation based on natural ecosystem dynamics in northern Europe: The complexity challenge. Ambio 2009, 38, 309–315. [Google Scholar] [CrossRef]
- Lõhmus, A.; Kraut, A.; Rosenvald, R. Dead wood in clearcuts of semi-natural forests in Estonia: Site-type variation, degradation, and the influences of tree retention and slash harvest. Eur. J. For. Res. 2013, 132, 335–349. [Google Scholar] [CrossRef]
- Sepp, T.; Liira, J. Factors influencing the species composition and richness of herb layer in old boreo-nemoral forests. For. Stud. 2009, 50, 23–41. [Google Scholar] [CrossRef]
- Lõhmus, A.; Lõhmus, P. Coarse woody debris in mid-aged stands: Abandoned agricultural versus long-term forest land. Can. J. For. Res. 2005, 35, 1502–1506. [Google Scholar] [CrossRef]
- Lõhmus, A.; Lõhmus, P. Old-forest species: The importance of specific substrata vs. stand continuity in the case of calicioid fungi. Silva Fenn. 2011, 45, 1015–1039. [Google Scholar] [CrossRef] [Green Version]
- Lõhmus, K.; Paal, T.; Liira, J. Long-term colonization ecology of forest-dwelling species in a fragmented rural landscape–dispersal versus establishment. Ecol. Evol. 2014, 4, 3113–3126. [Google Scholar] [CrossRef] [PubMed]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2020. Available online: http://qgis.osgeo.org (accessed on 18 May 2020).
- Wickham, H.; François, R.; Henry, L.; Müller, K. dplyr: A Grammar of Data Manipulation, R package version 0.8.4; 2020. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 18 May 2020).
- Pebesma, E. Simple features for R: Standardized support for spatial vector data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Pigott, C.D. Tilia cordata Miller. J. Ecol. 1991, 79, 1147–1207. [Google Scholar] [CrossRef]
- De Jaegere, T.; Hein, S.; Claessens, H. A review of the characteristics of small-leaved lime (Tilia cordata Mill.) and their implications for silviculture in a changing climate. Forests 2016, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Reier, Ü.; Tuvi, E.L.; Pärtel, M.; Kalamees, R.; Zobel, M. Threatened herbaceous species dependent on moderate forest disturbances: A neglected target for ecosystem-based silviculture. Scand. J. For. Res. 2005, 20, 145–152. [Google Scholar] [CrossRef]
- Nitare, J. (Ed.) Signalarter—Indikatorer på Skyddsvärd Skog: Flora över Kryptogamer; Skogsstyrelsens Förlag: Jönköping, Sweden, 2000. [Google Scholar]
- Lõhmus, A. Aspen-inhabiting Aphyllophoroid fungi in a managed forest landscape in Estonia. Scand. J. For. Res. 2011, 26, 212–220. [Google Scholar] [CrossRef]
- Orula, E.L. Distribution and Status of Dicranum viride (Sull. & Lesq.) Lindb. in Estonia. Ph.D. Thesis, Institute of Ecology and Earth Sciences, University of Tartu, Tartu, Estonia, 2004. (In Estonian). [Google Scholar]
- Jüriado, I.; Liira, J. Distribution and habitat ecology of the threatened forest lichen Lobaria pulmonaria in Estonia. Folia Cryptog. Estonica 2009, 46, 55–65. [Google Scholar]
- Leppik, E.; Jüriado, I.; Liira, J. Changes in stand structure due to the cessation of traditional land use in wooded meadows impoverish epiphytic lichen communities. Lichenologist 2011, 43, 257–274. [Google Scholar] [CrossRef]
- Lõhmus, A.; Runnel, K. Ash dieback can rapidly eradicate isolated epiphyte populations in production forests: A case study. Biol. Conserv. 2014, 169, 185–188. [Google Scholar] [CrossRef]
- Marmor, L.; Randlane, T.; Jüriado, I.; Saag, A. Host tree preferences of red-listed epiphytic lichens in Estonia. Balt. For. 2017, 23, 364–373. [Google Scholar]
- Angelstam, P.; Roberge, J.-M.; Lõhmus, A.; Bergmanis, M.; Brazaitis, G.; Dönz-Breuss, M.; Edenius, L.; Kosinski, Z.; Kurlavicius, P.; Lārmanis, V.; et al. Habitat modelling as a tool for landscape-scale conservation—A review of parameters for focal forest birds. Ecol. Bull. 2004, 51, 427–453. [Google Scholar]
- Brazaitis, G.; Angelstam, P. Influence of edges between old deciduous forest and clearcuts on the abundance of passerine hole-nesting birds in Lithuania. Ecol. Bull. 2004, 51, 209–217. [Google Scholar]
- Brazaitis, G. Forest interior species red-breasted flycatcher Ficedula parva habitat selection and conservation in intensive management areas. In Proceedings of the fifth International Scientific Conference, Rural Development, Kaunas, Lithuania, 24–25 November 2011; pp. 26–29. [Google Scholar]
- Mitrus, C.; Kleszko, N.; Soćko, B. Habitat characteristics, age, and arrival date of male red-breasted flycatchers Ficedula parva. Ethol. Ecol. Evol. 2006, 18, 33–41. [Google Scholar] [CrossRef]
- Juřičková, L.; Horsák, M.; Horáčková, J.; Abraham, V.; Ložek, V. Patterns of land-snail succession in Central Europe over the last 15,000 years: Main changes along environmental, spatial and temporal gradients. Quat. Sci. Rev. 2014, 93, 155–166. [Google Scholar] [CrossRef]
- Remm, L.; Lõhmus, A. Semi-naturally managed forests support diverse land snail assemblages in Estonia. For. Ecol. Manag. 2016, 363, 159–168. [Google Scholar] [CrossRef]
- Omerod, S.J.; Weatherley, N.S.; Merrett, W.J. The influence of conifer plantation on the distribution of the Golden Ringed Dragonfly Cordulegaster boltoni (Odonata) in upland wales. Biol. Conserv. 1990, 54, 241–251. [Google Scholar] [CrossRef]
- Allan, J.D. Landscapes and riverscapes: The influence of land use on stream ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef] [Green Version]
- Turley, M.D.; Bilotta, G.S.; Chadd, R.P.; Extence, C.A.; Brazier, R.E.; Burnside, N.G.; Pickwell, A.G. A sediment-specific family-level biomonitoring tool to identify the impacts of fine sediment in temperate rivers and streams. Ecol. Indic. 2016, 70, 151–165. [Google Scholar] [CrossRef] [Green Version]
- Reiber, L.; Knillmann, S.; Foit, K.; Liess, M. Species occurrence relates to pesticide gradient in streams. Sci. Total Environ. 2020, 735, 138807. [Google Scholar] [CrossRef] [PubMed]
- Ek, T.; Johannesson, J. Multi-Purpose Managementof Oak Habitats: Examples of Best Practice from the County of Östergötland, Sweden; Report 16; County Administration of Östergötland: Östergötland, Sweden, 2005. [Google Scholar]
- Bütler, R.; Lachat, T.; Larrieu, L.; Paillet, Y. Habitat trees: Key elements for forest biodiversity. In Integrative Approaches as an Opportunity for the Conservation of Forest Biodiversity; Kraus, D., Krumm, F., Eds.; European Forest Institute: Joensuu, Finland, 2013; pp. 84–91. [Google Scholar]
- Rosenvald, R.; Lõhmus, P.; Rannap, R.; Remm, L.; Rosenvald, K.; Runnel, K.; Lõhmus, A. Assessing long-term effectiveness of green-tree retention. For. Ecol. Manag. 2019, 448, 543–548. [Google Scholar] [CrossRef]
- Lindenmayer, D.; Pierson, J.; Barton, P.; Beger, M.; Branquinho, C.; Calhoun, A.; Caro, T.; Greig, H.; Gross, J.; Heino, J.; et al. A new framework for selecting environmental surrogates. Sci. Total Environ. 2015, 538, 1029–1038. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Fischer, J.; Felton, A.; Montague-Drake, R.D.; Manning, A.; Simberloff, D.; Youngentob, K.; Saunders, D.; Wilson, D.M.; Felton, A.; et al. The complementarity of single-species and ecosystem-oriented research in conservation research. Oikos 2007, 116, 1220–1226. [Google Scholar] [CrossRef]
- Landres, P.B.; Verner, J.; Thomas, J.W. Ecological uses of vertebrate indicator species: A critique. Conserv. Biol. 1988, 2, 316–328. [Google Scholar] [CrossRef]
- Roberge, J.M.; Angelstam, P. Usefulness of the umbrella species concept as a conservation tool. Conserv. Biol. 2004, 18, 76–85. [Google Scholar] [CrossRef]
- Lõhmus, A.; Soon, M. The use of umbrella species in ecologically sustainable forestry: A critical review and the perspectives in Estonia. For. Stud. 2004, 41, 73–85. [Google Scholar]
- Remm, L.; Lõhmus, P.; Leis, M.; Lõhmus, A. Long-term impacts of forest ditching on non-aquatic biodiversity: Conservation perspectives for a novel ecosystem. PLoS ONE 2013, 8, e63086. [Google Scholar] [CrossRef] [Green Version]
- DeRoy, B.; Darimont, C. Biocultural indicators to support locally led environmental management and monitoring. Ecol. Soc. 2019, 24, 21. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Likens, G.E. Direct measurement versus surrogate indicator species for evaluating environmental change and biodiversity loss. Ecosystems 2011, 14, 47–59. [Google Scholar] [CrossRef]
- Gao, T.; Nielsen, A.B.; Hedblom, M. Reviewing the strength of evidence of biodiversity indicators for forest ecosystems in Europe. Ecol. Indic. 2015, 57, 420–434. [Google Scholar] [CrossRef]
- Yu, D.; Lu, N.; Fu, B. Establishment of a comprehensive indicator system for the assessment of biodiversity and ecosystem services. Landsc. Ecol. 2017, 32, 1563–1579. [Google Scholar] [CrossRef]
- Uusitalo, L.; Lehikoinen, A.; Helle, I.; Myrberg, K. An overview of methods to evaluate uncertainty of deterministic models in decision support. Environ. Model. Softw. 2015, 63, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Laurila-Pant, M.; Lehikoinen, A.; Uusitalo, L.; Venesjärvi, R. How to value biodiversity in environmental management? Ecol. Indic. 2015, 55, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rose, D.C.; Mukherjee, N.; Simmons, B.I.; Tew, E.R.; Robertson, R.J.; Vadrot, A.B.; Doubleday, R.; Sutherland, W.J. Policy windows for the environment: Tips for improving the uptake of scientific knowledge. Environ. Sci. Policy 2017. [Google Scholar] [CrossRef] [Green Version]
- Jones-Farrand, D.T.; Fearer, T.M.; Thogmartin, W.E.; Thompson, F.R., III; Nelson, M.D.; Tirpak, J.M. Comparison of statistical and theoretical habitat models for conservation planning: The benefit of ensemble prediction. Ecol. Appl. 2011, 21, 2269–2282. [Google Scholar] [CrossRef] [Green Version]
- Penaluna, B.E.; Olson, D.H.; Flitcroft, R.L.; Weber, M.A.; Bellmore, J.R.; Wondzell, S.M.; Dunham, J.B.; Johnson, S.L.; Reeves, G.H. Aquatic biodiversity in forests: A weak link in ecosystem services resilience. Biodivers. Conserv. 2017, 26, 3125–3155. [Google Scholar] [CrossRef]
- Strayer, D.L. Challenges for freshwater invertebrate conservation. J. N. Am. Benthol. Soc. 2006, 25, 271–287. [Google Scholar] [CrossRef]
- Menezes, S.; Baird, D.J.; Soares, A.M. Beyond taxonomy: A review of macroinvertebrate trait-based community descriptors as tools for freshwater biomonitoring. J. Appl. Ecol. 2010, 47, 711–719. [Google Scholar] [CrossRef]
- Schmera, D.; Heino, J.; Podani, J.; Erős, T.; Dolédec, S. Functional diversity: A review of methodology and current knowledge in freshwater macroinvertebrate research. Hydrobiologia 2017, 787, 27–44. [Google Scholar] [CrossRef] [Green Version]
- Supp, S.R.; Ernest, S.M. Species-level and community-level responses to disturbance: A cross-community analysis. Ecology 2014, 95, 1717–1723. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Dang, H.; Wang, M.; Chai, Y.; Guo, Y.; Chen, Y.; Zhang, C.; Yue, M. Is phylogeny more useful than functional traits for assessing diversity patterns under community assembly processes? Forests 2019, 10, 1159. [Google Scholar] [CrossRef] [Green Version]
- Forest Stewardship Council. FSC International Standard: FSC Principles and Criteria for Forest Stewardship. FSC-STD-01–001 V5–2. Available online: https://fsc.org/en/document-centre/documents/resource/392 (accessed on 18 May 2020).
- Eckelt, A.; Müller, J.; Bense, U.; Brustel, H.; Bußler, H.; Chittaro, Y.; Cizek, L.; Frei, A.; Holzer, E.; Kadej, M.; et al. “Primeval forest relict beetles” of Central Europe: A set of 168 umbrella species for the protection of primeval forest remnants. J. Insect Conserv. 2018, 22, 15–28. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Fischer, J.; Hobbs, R. The need for pluralism in landscape models: A reply to Dunn and Majer. Oikos 2007, 116, 1419–1421. [Google Scholar] [CrossRef]
- Bryan, B.A.; Crossman, N.D.; King, D.; Meyer, W.S. Landscape futures analysis: Assessing the impacts of environmental targets under alternative spatial policy options and future scenarios. Environ. Model. Softw. 2011, 26, 83–91. [Google Scholar] [CrossRef]
- Parrott, L.; Meyer, W.S. Future landscapes: Managing within complexity. Front. Ecol. Environ. 2012, 10, 382–389. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
A framework of targets and activities that link basic biodiversity knowledge (left triangle) and sustainable forest management (SFM) (right triangle) through the nexus of focal species habitat modeling. The activities indicated by numbers: 1, Red-listing of threatened species; 2, distinguishing focal species by listing major threats; 3, focal species habitat modeling; 4, habitat conservation; 5, landscape design.
Figure 1.
A framework of targets and activities that link basic biodiversity knowledge (left triangle) and sustainable forest management (SFM) (right triangle) through the nexus of focal species habitat modeling. The activities indicated by numbers: 1, Red-listing of threatened species; 2, distinguishing focal species by listing major threats; 3, focal species habitat modeling; 4, habitat conservation; 5, landscape design.

Figure 2.
Decision trees for spatial modeling of the focal taxon groups D1–D3 and D5 in meso-eutrophic and eutrophic forests in Estonia (cf. Table 2): (a) poorly dispersing perennial plant of eutrophic gap-dynamic forests; (b) saprophytic fungus inhabiting continuous supply of large downed trunks of Picea abies; (c) rare fungus inhabiting senescent or dead Populus tremula in mid-succession; (d) area-sensitive vertebrate of vertically structured stands. The decision order is from left to right, and from top to bottom; the bottom row comprises habitat-quality scores (0...10; colors referring to broad classes). The parameters marked with asterisk (*) were modeled as dynamic in the 10-year future scenario. If not specified, the tree variables refer to the 1st layer. The site type codes in (a): ND, Aegopodium; SL, Hepatica; kSJ, drained Dryopteris; JK, Oxalis; AN, Filipendula.
Figure 2.
Decision trees for spatial modeling of the focal taxon groups D1–D3 and D5 in meso-eutrophic and eutrophic forests in Estonia (cf. Table 2): (a) poorly dispersing perennial plant of eutrophic gap-dynamic forests; (b) saprophytic fungus inhabiting continuous supply of large downed trunks of Picea abies; (c) rare fungus inhabiting senescent or dead Populus tremula in mid-succession; (d) area-sensitive vertebrate of vertically structured stands. The decision order is from left to right, and from top to bottom; the bottom row comprises habitat-quality scores (0...10; colors referring to broad classes). The parameters marked with asterisk (*) were modeled as dynamic in the 10-year future scenario. If not specified, the tree variables refer to the 1st layer. The site type codes in (a): ND, Aegopodium; SL, Hepatica; kSJ, drained Dryopteris; JK, Oxalis; AN, Filipendula.

Figure 3.
Predictive habitat modeling of new reserves for eutrophic and meso-eutrophic forests in Estonia. (a) Locations of the reserves. (b–d) Predicted distribution of quality habitats (scores 7–10) in 2029 for three focal taxa, zoomed in for a selected reserve (a: red box). The colors refer to 2019 habitat quality (cf. Figure 2) and reveal: (b) moderately favorable, but only slowly improving situation for the perennial plant (D1); (c) poor, but rapidly improving, situation for the Picea-inhabiting old-forest fungus (D2); (d) favorable and further improving situation for the old-aspen inhabiting fungus (D3). (e) Shrinking of non-habitat (score 0–2) for any terrestrial focal species (D1–D6) by 2029 (black) from its current distribution (red).
Figure 3.
Predictive habitat modeling of new reserves for eutrophic and meso-eutrophic forests in Estonia. (a) Locations of the reserves. (b–d) Predicted distribution of quality habitats (scores 7–10) in 2029 for three focal taxa, zoomed in for a selected reserve (a: red box). The colors refer to 2019 habitat quality (cf. Figure 2) and reveal: (b) moderately favorable, but only slowly improving situation for the perennial plant (D1); (c) poor, but rapidly improving, situation for the Picea-inhabiting old-forest fungus (D2); (d) favorable and further improving situation for the old-aspen inhabiting fungus (D3). (e) Shrinking of non-habitat (score 0–2) for any terrestrial focal species (D1–D6) by 2029 (black) from its current distribution (red).


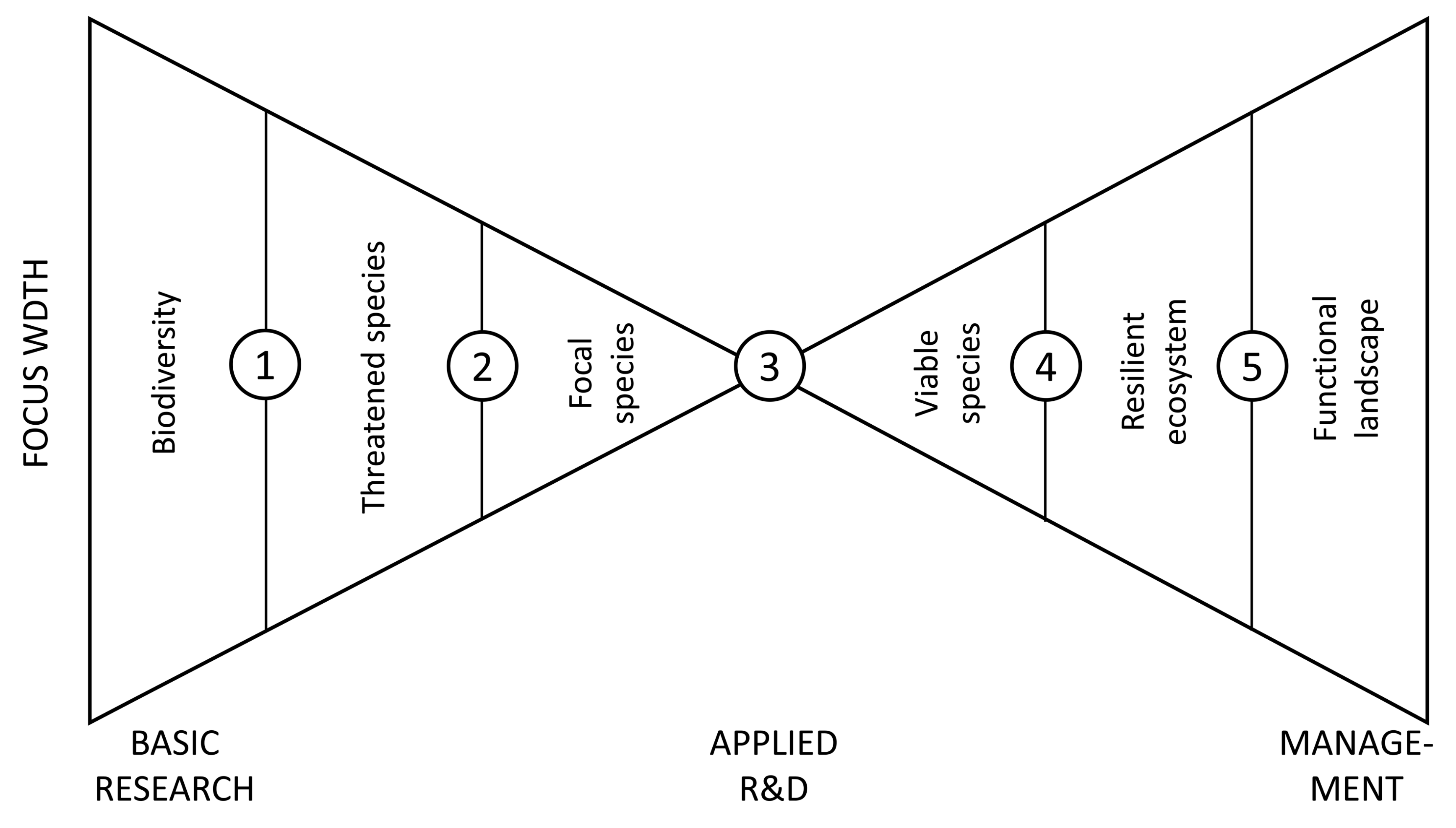{kind=link}
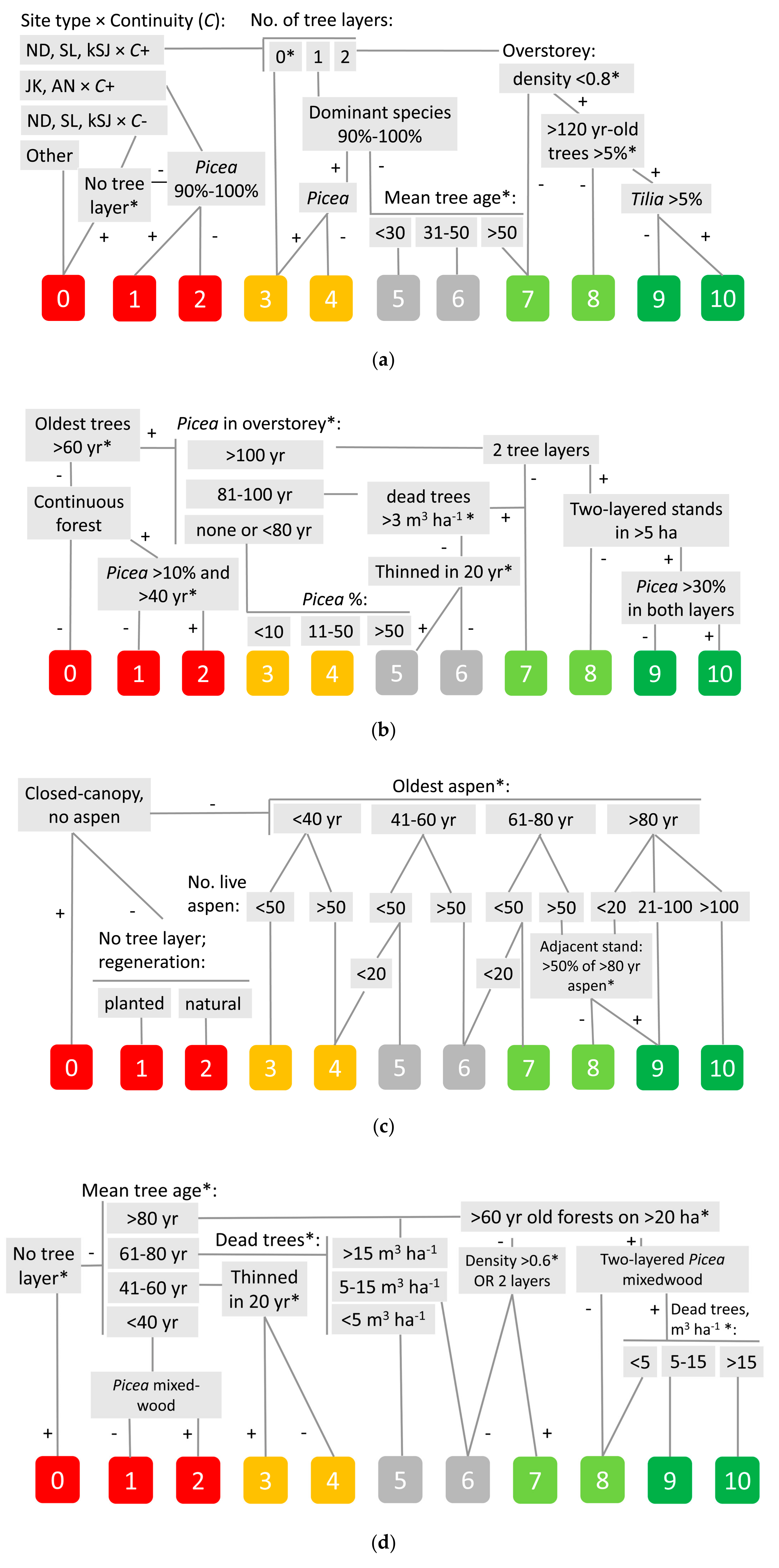{kind=link}
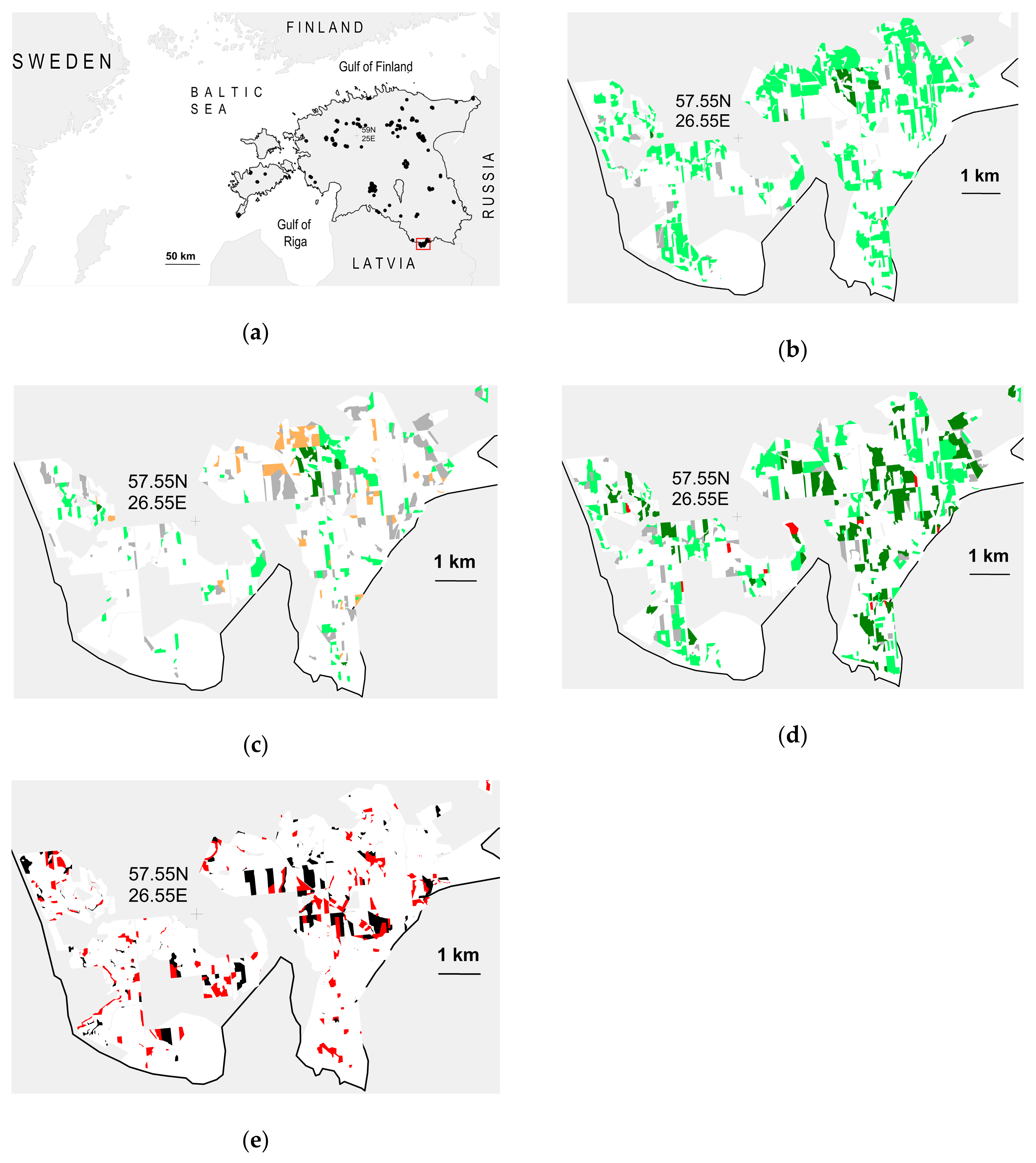{kind=link}
Table 1.
Published habitat modeling studies of focal forest species that link the distribution and dynamics of biodiversity threats with implications to management planning and forest policy.
Table 1.
Published habitat modeling studies of focal forest species that link the distribution and dynamics of biodiversity threats with implications to management planning and forest policy.
| Focal Taxon, Study System and Focus 1 | Summary of the Scenario Results | Reference 2 |
|---|---|---|
| I. Twenty-eight vertebrates sensitive to diverse habitat changes in western US | In 100 years, landscape management for ecosystem health and services would improve habitat of old-forest species to >80% and of a snag-dependent bird to ⅔ of the 19th-century levels. Fine-scale planning can increase high-quality habitat at a stable average habitat quality level. | [75] |
| I. Ten taxa (birds; mammals; macrolichens) in Oregon | Projecting the 1990s forestry policies for 100 years shows increased contrasts in habitat distribution by ownership. Public lands support an increase in old forest. Expanding retention forestry to private lands is needed to mitigate the loss of semi-open forests. Loss of hardwood habitats remains to be addressed. | [100,101] |
| I. A lichen, a bird and a butterfly in Scotland | Restoring a part of conifer plantations to native woodland and open land supports specialist species and has no apparent detrimental influence on generalist species on the landscape. | [102] |
| I. Nine vertebrates sensitive to diverse habitat changes near Seattle, US | Suburbanization generally reduces forest habitats, but some mature-forest specialists may also benefit from reduced logging if human settlers tend not to clear forests near houses. | [103]* |
| II. Sixteen habitat specialist birds and amphibians in North Carolina, US | Wood bioenergy use scenarios predict habitat gains for shrub-associated species and habitat loss for mature forest species in 40 years; the species negatively affected tend to be threatened by other processes as well. | [104] |
| II. Three mature-forest vertebrates in Washington, US | In 80-year projections, moderate thinnings to accelerate forest growth appear as the best silvicultural strategy that does not reduce the habitat of any species while producing substantial timber revenues (39% of intensive forestry). | [105] |
| II. Twenty-seven saproxylic insects, fungi, lichens in Finland | In a 60-year perspective, a cost-effective strategy to increase habitat quality of production forests is to reduce the area that is conventionally thinned. | [106]# |
| II. Woodland caribou and Martes americana in British Columbia | In a landscape with production and protected forests, a management strategy that keeps the total area of caribou winter habitat at a stable level through time optimizes the trade-offs between old-growth protection and timber harvest. | [107]# |
| II. Picoides arcticus in Canadian conifer forests | In 100 years, current-level harvesting would much reduce recruitment of this old-growth bird. Wildfire intensification due to climate change aggravates the decline. Reduced harvesting and promoting conifers mitigate these impacts. | [108] |
| II. Seiurus aurocapillus in Canadian hardwoods | In 80 years, immigration to intensively managed districts retains a sink population of this hardwood specialist at only 25% lower densities than without harvest. Replacing 10%-20% of selection cuttings with shelterwood would add little stress, but climate change would much accelerate reduction. | [109] |
| II. Three birds and a beetle specific to forest successional stages in Sweden | Extended or shortened rotations affect the species positively or negatively depending on habitat requirements. However, even favorable scenarios can cause temporary reductions in 150 years due to uneven distribution of stand-ages. | [110] |
| III. Two passerines with distinct niches in the central U.S. | Restoring forest area (afforestation) supported population increase better than restoring existing forest habitats, but it was effective only when targeted non-randomly to key areas to reduce fragmentation. | [111]# |
| III. Tympanuchus phasianellus in clearcuts in Wisconsin | Clearcutting greatly affects this early-successional species even in the presence of stable open habitat. Yet the harvest regimes creating the largest clearcut areas are not necessarily best for population viability. | [112] |
| III. Five epiphytic lichens on old oaks in Sweden | Promoting host tree availability (regeneration or clearing brushwood around shaded oaks) may effectively support the metapopulations in areas with high densities of trees still present, but not in impoverished landscapes. | [113]# |
| IV. Oncorhynchus spp. in forest streams in Oregon | Projecting the 1990s forestry policies for 100 years increases suitable stream habitat with large trees on river banks for one salmon species, while another species cannot recover without additional policies on private lands. | [114]* |
| IV. Strix occidentalis in the Pacific Northwest | Old-forest reserves are efficient in capturing current owl habitat, but official 2007 proposals would have reduced that efficiency, and it will, nevertheless, decline due to climate change. The performance of the network and its value for 130 accompanying species can be enhanced by prioritizing connectivity of current and future habitat. | [115,116,117]* |
| IV. Seventeen flagship mammals in Thailand | Along with forest cover decline from 57% to 50% by 2050, most species lose habitat despite proposed additional reserves. The vulnerability of the reserves to isolation is much increased due to climate-change caused habitat turnover. | [118] |
| V. Three birds of vulnerable forest ecosystems in South Africa | Based on the species’ habitat connectivity mapping and climate-change scenarios, the study maps and prioritizes potential extensions of the current protected area network in the region. | [119] |
| V. Martes americana in the Appalachians | For this old-forest species, the reduction in logging can mitigate population declines that are expected due to climate change in this vulnerable hotspot region. | [120] |
1 Main focus of the study: I, mapping forest biodiversity dimensions of landscape change; II, stand-scale effects of intensive timber harvesting in forests; III, metapopulation viability in dynamic woodlands; IV, forest set-asides to protect flagship species; V, biodiversity assessment of forest futures in biodiversity hotspots. 2 *Studies for official programs for biodiversity conservation or mitigating the environmental impact; #studies not captured with the formal search string
Table 2.
Seven generalized ecological groups of focal-taxa (‘ecological profiles’), their focal species, variables for the spatial models, and predicted habitat areas in strict reserves in meso-eutrophic and eutrophic forests in Estonia.
Table 2.
Seven generalized ecological groups of focal-taxa (‘ecological profiles’), their focal species, variables for the spatial models, and predicted habitat areas in strict reserves in meso-eutrophic and eutrophic forests in Estonia.
| Ecological Profile | |||||||
|---|---|---|---|---|---|---|---|
| Characteristic | D1 Gap Dynamics | D2 Picea, Saproxylic | D3 Populus, Succession | D4 Nemoral Tree Species, Epiphyte | D5 Vert. Struct., Interior | D6 Soil and Litter | D7 Streams |
| Focal taxa | |||||||
| Taxon group | Vascular plants | Fungi | Old-aspen specialists | Lichens, bryophytes | Verteb- rates | Snails | Aquatic insects |
| Frequent, sensitive species | Tilia cordata | Phellinus ferrugineo- fuscus | Megalaria grossa | Chrysothrix candelaris | Ficedula parva | Acanthinu-la aculeata | Plectrocne- mia conspersa |
| Rare, threatened species | Bromus benekenii | Antrodia piceata | Junghuhnia pseudo- zilingiana | Dicranum viride, Lobaria pulmonaria | - | Bulgarica cana | Cordulegas- ter boltonii |
| Model variables1 | |||||||
| Soil type | *** | *** | |||||
| Stand age | *** | *** | *** | *** | *** | *** | ** |
| Tree species | *** | *** | *** | *** | *** | ** | * |
| Tree layers | ** | * | * | * | |||
| Stand density | * | * | * | ||||
| Dead wood | ** | ** | |||||
| Continuity | *** | *** | *** | ||||
| Thinning | ** | ** | * | ||||
| Draining | *** | ||||||
| Landscape | * | ** | |||||
| Habitat extent (%)2 | |||||||
| Unsuitable 2019 | 45 | 54 | 56 | 81 | 42 | 35 | 96 |
| High: 2019 | 26 | 11 | 28 | 4 | 20 | 5 | 1 |
| HighC 2009-19 | -3 | +4 | +4 | +2 | +5 | 0 | 0 |
| HighC 2019-29 | +2 | +12 | +4 | +2 | +11 | 0 | 0 |
| Key ref. | [167,180,181,182] | [91,183] | [91,183,184] | [185,186,187,188,189] | [190,191,192,193] | [194,195] | [196,197,198,199] |
Note: 1 Variable priority on decision tree (cf. Figure 2): *** distinguishes non-habitat (scores 0–2); ** organizes poor habitats (3–6) or * quality habitats (7–10). Land-use history variables refer to activities carried out in the last 20 years (Thinning) or depicted on topographic maps since 1900 (Continuity of forest use; Draining). 2 Refers to the frequent species: Unsuitable, scores 0–2; High, scores 7–10. Changes in High (HighC) are percentage points relative to the 2019 level. References are for regional justification of the species and the model variables; the process also used unpublished expert knowledge.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lõhmus, A.; Kont, R.; Runnel, K.; Vaikre, M.; Remm, L. Habitat Models of Focal Species Can Link Ecology and Decision-Making in Sustainable Forest Management. Forests 2020, 11, 721. https://doi.org/10.3390/f11070721
AMA Style
Lõhmus A, Kont R, Runnel K, Vaikre M, Remm L. Habitat Models of Focal Species Can Link Ecology and Decision-Making in Sustainable Forest Management. Forests. 2020; 11(7):721. https://doi.org/10.3390/f11070721
Chicago/Turabian StyleLõhmus, Asko, Raido Kont, Kadri Runnel, Maarja Vaikre, and Liina Remm. 2020. "Habitat Models of Focal Species Can Link Ecology and Decision-Making in Sustainable Forest Management" Forests 11, no. 7: 721. https://doi.org/10.3390/f11070721
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.