Metabolic Profiling Provides Unique Insights to Accumulation and Biosynthesis of Key Secondary Metabolites in Annual Pasture Legumes of Mediterranean Origin


 , and
, and
Abstract
:
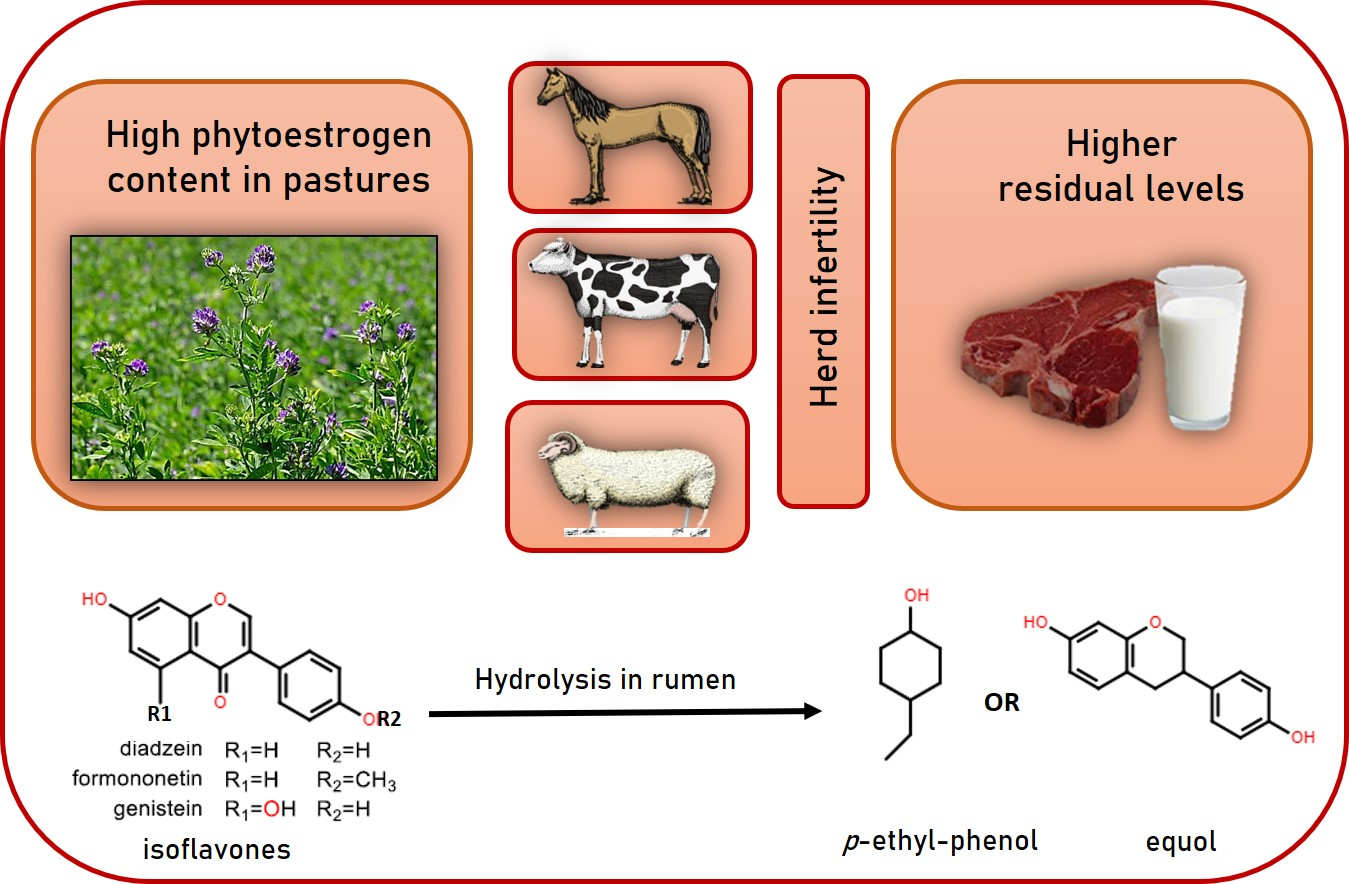
1. Introduction
2. Results
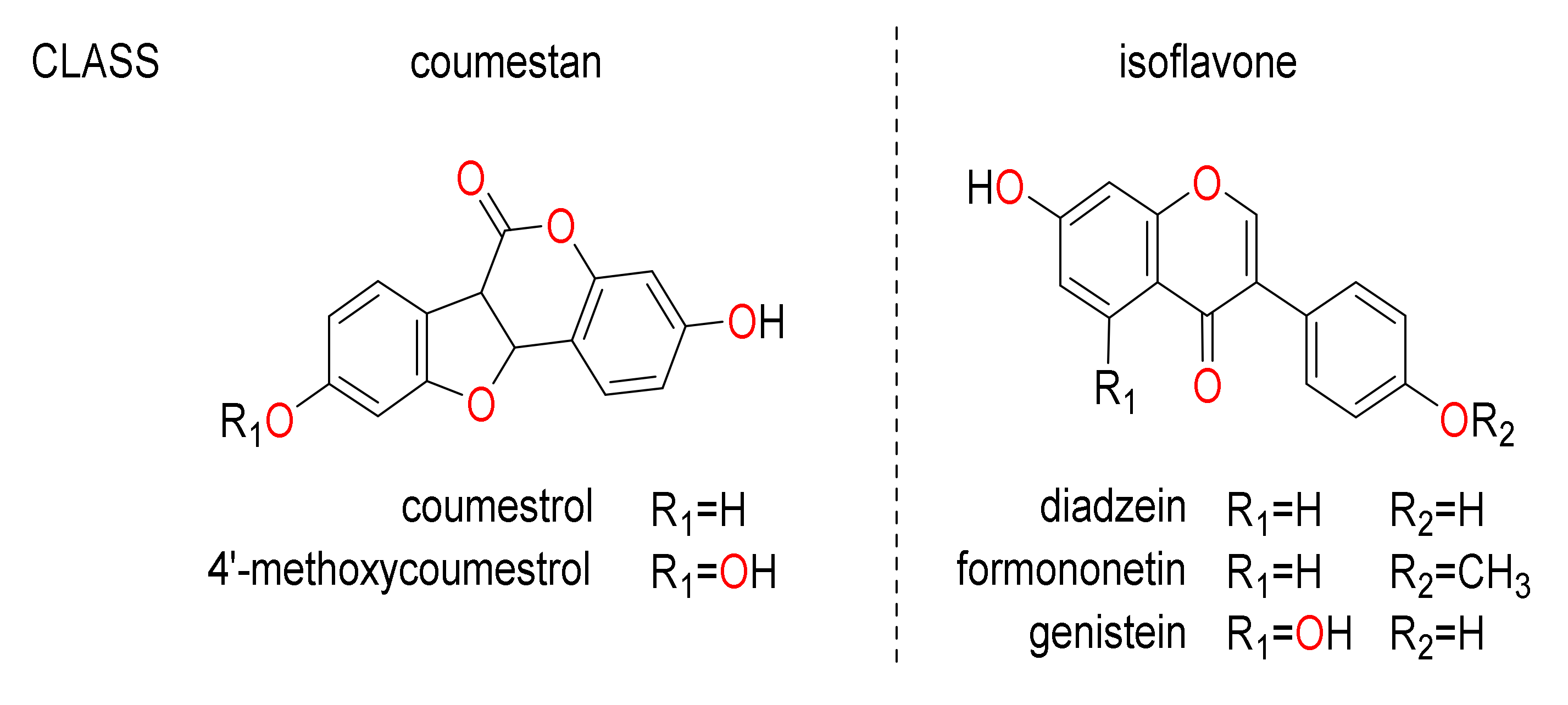
2.1. Quantification of Phytoestrogens
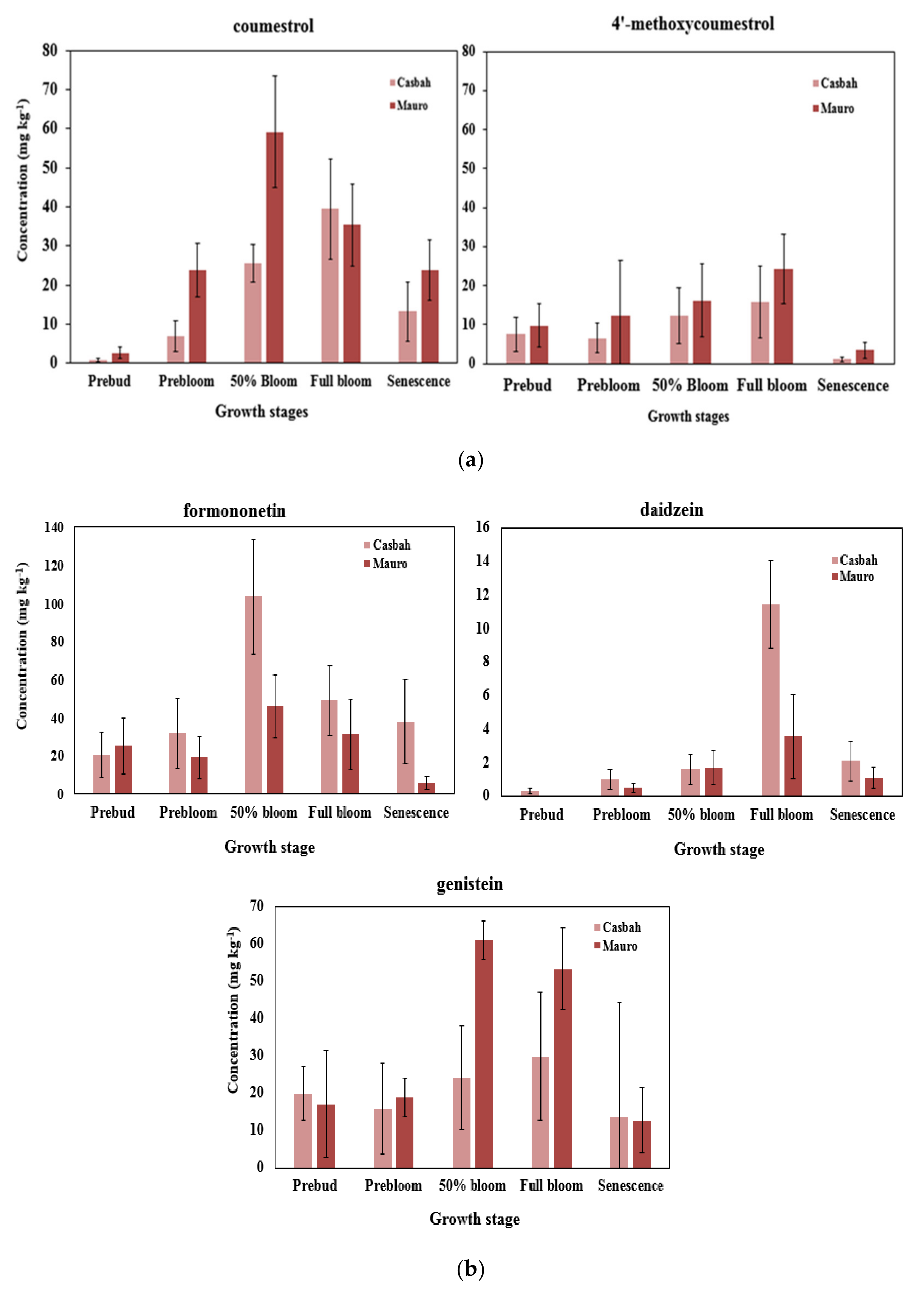
2.2. Effect of Biserrula Cultivar and Growth Stage on Phytoestrogen Levels
2.3. Quantification of Total Polyphenols and Proanthocyanidins
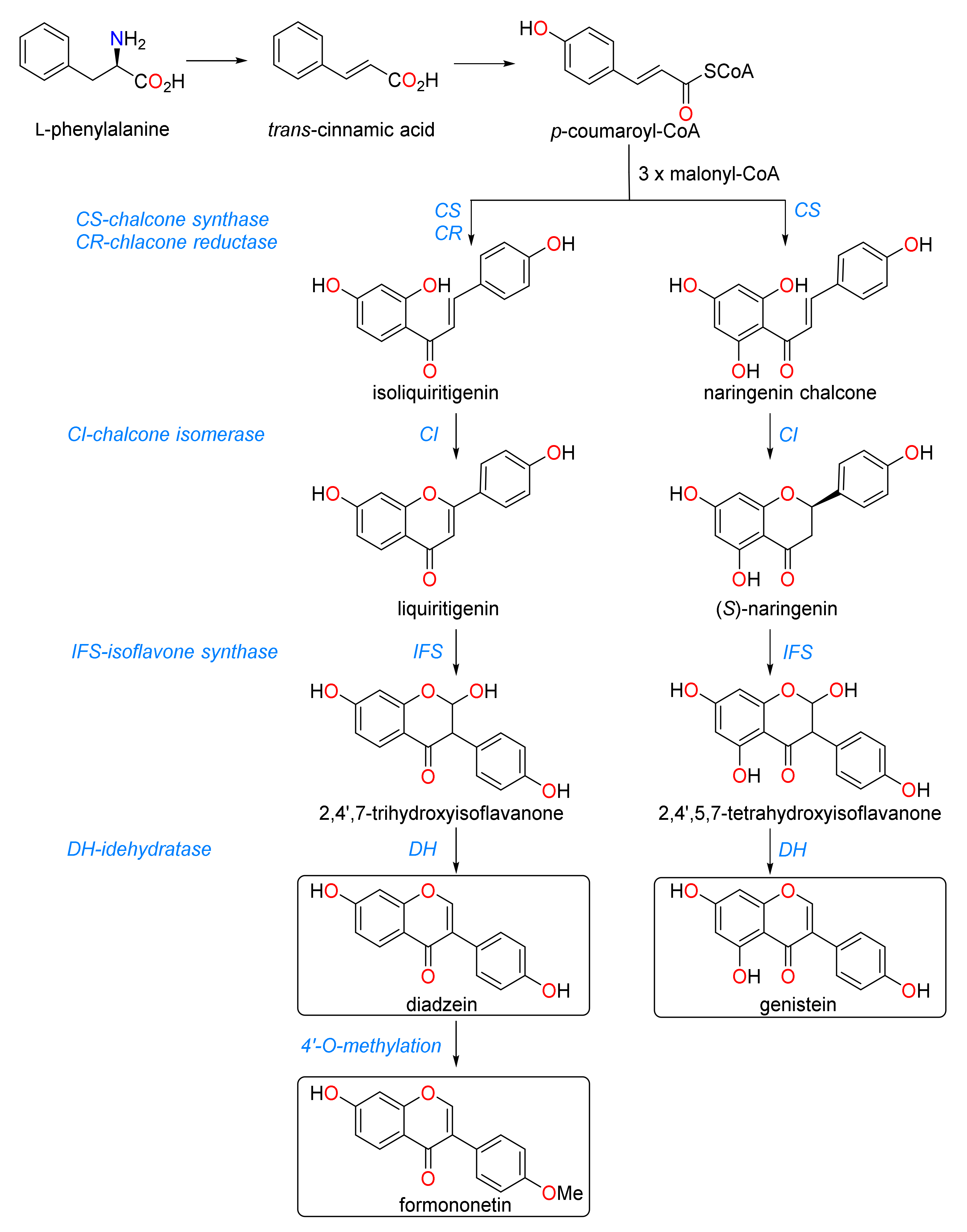
2.4. Abundance of Flavonoids and Their Glycosides
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material
4.3. LC-QToF-MS Analysis
4.4. Extraction of Polyphenols
4.5. Quantification of Total Polyphenol Content
4.6. Quantification of Total Proanthocyanidin Content (TPAC)
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Whitbread, A.M.; Hall, C.A.; Pengelly, B.C. A novel approach to planting grasslegume pastures in the mixed farming zone of southern inland Queensland, Australia. Crop Pasture Sci. 2009, 60, 1147–1155. [Google Scholar] [CrossRef]
- Hackney, B.; Dear, B.; Li, G.; Rodham, C.; Tidd, J. Current and future use of pasture legumes in central and southern NSW—Results of a farmer and advisor survey. In Proceedings of the 14th Australian Society of Agronomy Conference, Adelaide, Australia, 21–25 September 2008. [Google Scholar]
- Loi, A.; Howieson, J.G.; Nutt, B.J.; Carr, S.J. A second generation of annual pasture legumes and their potential for inclusion in Mediterranean-type farming systems. Aust. J. Exp. Agric. 2005, 45, 289–299. [Google Scholar] [CrossRef]
- Burggraaf, V.; Waghorn, G.; Woodward, S.; Thom, E. Effects of condensed tannins in white clover flowers on their digestion in vitro. Anim. Feed Sci. Technol. 2008, 142, 44–58. [Google Scholar] [CrossRef]
- Hall, D.; Wolfe, E.; Cullis, B. Performance of breeding ewes on lucerne-subterranean clover pastures. Aust. J. Exp. Agric. 1985, 25, 758–765. [Google Scholar] [CrossRef]
- Humphries, A. Future applications of lucerne for efficient livestock production in southern Australia. Crop Pasture Sci. 2013, 63, 909–917. [Google Scholar] [CrossRef]
- Mulholland, J. Animal production from lucerne-based pastures. In Realising the Potential of Pastures, the Proceedings of the 16th Riverina Outlook Conference, Wagga Wagga, Australia, 15 July, 1987; Riverina-Murray Institute of Higher Education: Wagga Wagga, Australia, 1987; pp. 57–66. [Google Scholar]
- Loi, A.; Howieson, J.H.; Carr, S.J. Register of Australian herbage plant cultivars. Biserrula pelecinus L. (biserrula) cv. Casbah. Aust. J. Exp. Agric. 2001, 41, 841–842. [Google Scholar] [CrossRef]
- Hackney, B.; Nutt, B.; Loi, A.; Yates, R.; Quinn, J.; Piltz, J.; Jenkins, J.; Weston, L.; O’Hare, M.; Butcher, A. “On-demand” hardseeded pasture legumes–a paradigm shift in crop-pasture rotations for southern Australian mixed farming systems. In Building Productive, Diverse and Sustainable Landscapes, Proceedings of the 17th Australian Agronomy Conference, Hobart, Australia, 20–24 September 2015; Acuña, T., Moeller, C., Parsons, D., Harrison, M., Eds.; Australian Agronomy society: Hobart, Australia, 2015. [Google Scholar]
- Dear, B.; Ewing, M.A. The search for new pasture plants to achieve more sustainable production systems in southern Australia. Aust. J. Exp. Agric. 2008, 48, 387–396. [Google Scholar] [CrossRef]
- Latif, S.; Gurusinghe, S.; Weston, P.A.; Brown, W.B.; Quinn, J.C.; Piltz, J.W.; Weston, L.A. Performance and weed-suppressive potential of selected pasture legumes against annual weeds in south-eastern Australia. Crop Pasture Sci. 2019, 70, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Mathesius, U. Flavonoid Functions in Plants and Their Interactions with Other Organisms. Plants 2018, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Weston, L.A.; Mathesius, U. Flavonoids: Their structure, biosynthesis and role in the rhizosphere, including allelopathy. J. Chem. Ecol. 2013, 39, 283–297. [Google Scholar] [CrossRef]
- Hassan, S.; Mathesius, U. The role of flavonoids in root–rhizosphere signalling: Opportunities and challenges for improving plant–microbe interactions. J. Exp. Bot. 2012, 63, 3429–3444. [Google Scholar] [CrossRef] [Green Version]
- Reed, K.F.M. Fertility of herbivores consuming phytoestrogen-containing Medicago and Trifolium species. Agriculture 2016, 6, 35. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, B.K.; Brunton, N.P.; Brennan, C. Handbook of Plant Food Phytochemicals: Sources, Stability and Extraction; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Wocławek-Potocka, I.; Mannelli, C.; Boruszewska, D.; Kowalczyk-Zieba, I.; Waśniewski, T.; Skarżyński, D.J. Diverse effects of phytoestrogens on the reproductive performance: Cow as a model. Int. J. Endocrinol. 2013, 2013, 650984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashem, N.; El-Azrak, K.; Sallam, S. Hormonal concentrations and reproductive performance of Holstein heifers fed Trifolium alexandrinum as a phytoestrogenic roughage. Anim. Reprod. Sci. 2016, 170, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Hloucalová, P.; Skládanka, J.; Horký, P.; Klejdus, B.; Pelikán, J.; Knotová, D. Determination of Phytoestrogen Content in Fresh-Cut Legume Forage. Animals 2016, 6, 43. [Google Scholar] [CrossRef] [Green Version]
- Burton, J.; Wells, M. The effect of phytoestrogens on the female genital tract. J. Clin. Pathol. 2002, 55, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Cvejić, J.; Bursać, M.; Atanacković, M. Phytoestrogens: “Estrogene-Like” Phytochemicals. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2012; Volume 38, pp. 1–35. [Google Scholar]
- Bickoff, E.; Booth, A.; Lyman, R.; Livingston, A.; Thompson, C.; Deeds, F. Coumestrol, a new estrogen isolated from forage crops. Science 1957, 126, 969–970. [Google Scholar] [CrossRef] [PubMed]
- Mostrom, M.; Evans, T.J. Phytoestrogens; Elsevier: Amsterdam, The Netherlands, 2011; pp. 707–722. [Google Scholar]
- Woclawek-Potocka, I.; Okuda, K.; Acosta, T.; Korzekwa, A.; Pilawski, W.; Skarzynski, D. Phytoestrogen metabolites are much more active than phytoestrogens themselves in increasing prostaglandin F2α synthesis via prostaglanin F2α synthase-like 2 stimulation in bovine endometrium. Prostaglandins Other Lipid Mediat. 2005, 78, 202–217. [Google Scholar] [CrossRef]
- Lookhart, G. Analysis of coumestrol, a plant estrogen, in animal feeds by high-performance liquid chromatography. J. Agric. Food Chem. 1980, 28, 666–667. [Google Scholar] [CrossRef]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.J.; Reddy, M.S.; Wang, L. The phenylpropanoid pathway and plant defence—A genomics perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef]
- Yu, O.; Jung, W.; Shi, J.; Croes, R.A.; Fader, G.M.; McGonigle, B.; Odell, J.T. Production of the Isoflavones Genistein and Daidzein in Non-Legume Dicot and Monocot Tissues. Plant Physiol. 2000, 124, 781–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, O.P.; Dahuja, A.; Sachdev, A.; Jain, P.K.; Kumari, S.; Praveen, S. Cytosine Methylation of Isoflavone Synthase Gene in the Genic Region Positively Regulates Its Expression and Isoflavone Biosynthesis in Soybean Seeds. DNA Cell Biol. 2019, 38, 510–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, R.A.; Harrison, M.J.; Paiva, N.L. The isoflavonoid phytoalexin pathway from enzymes to genes to transcription factors. Physiol. Plant. 1995, 93, 385–392. [Google Scholar] [CrossRef] [Green Version]
- Piluzza, G.; Sulas, L.; Bullitta, S. Tannins in forage plants and their role in animal husbandry and environmental sustainability: A review. Grass Forage Sci. 2014, 69, 32–48. [Google Scholar] [CrossRef]
- Salami, S.A.; Luciano, G.; O’Grady, M.N.; Biondi, L.; Newbold, C.J.; Kerry, J.P.; Priolo, A. Sustainability of feeding plant by-products: A review of the implications for ruminant meat production. Anim. Feed Sci. Technol. 2019, 251, 37–55. [Google Scholar] [CrossRef]
- Aerts, R.J.; Barry, T.N.; McNabb, W.C. Polyphenols and agriculture: Beneficial effects of proanthocyanidins in forages. Agric. Ecosyst. Environ. 1999, 75, 1–12. [Google Scholar] [CrossRef]
- Douglas, G.; Stienezen, M.; Waghorn, G.; Foote, A.; Purchas, R. Effect of condensed tannins in birdsfoot trefoil (Lotus corniculatus) and sulla (Hedysarum coronarium) on body weight, carcass fat depth, and wool growth of lambs in New Zealand. N. Z. J. Agric. Res. 1999, 42, 55–64. [Google Scholar] [CrossRef]
- Ramírez-Restrepo, C.; Barry, T. Alternative temperate forages containing secondary compounds for improving sustainable productivity in grazing ruminants. Anim. Feed Sci. Technol. 2005, 120, 179–201. [Google Scholar] [CrossRef]
- Kingston-Smith, A.H.; Edwards, J.E.; Huws, S.A.; Kim, E.J.; Abberton, M. Plant-based strategies towards minimising ‘livestock’s long shadow’. Proc. Nutr. Soc. 2010, 69, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Tzamaloukas, O.; Athanasiadou, S.; Kyriazakis, I.; Huntley, J.; Jackson, F. The effect of chicory (Cichorium intybus) and sulla (Hedysarum coronarium) on larval development and mucosal cell responses of growing lambs challenged with Teladorsagia circumcincta. Parasitology 2006, 132, 419–426. [Google Scholar] [CrossRef]
- Hoste, H.; Torres-Acosta, J.; Quijada, J.; Chan-Perez, I.; Dakheel, M.; Kommuru, D.; Mueller-Harvey, I.; Terrill, T. Interactions between nutrition and infections with Haemonchus contortus and related gastrointestinal nematodes in small ruminants. In Advances in Parasitology; Elsevier: Amsterdam, The Netherlands, 2016; Volume 93, pp. 239–351. [Google Scholar]
- Min, B.R.; Barry, T.N.; Attwood, G.T.; McNabb, W.C. The effect of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: A review. Anim. Feed Sci. Technol. 2003, 106, 3–19. [Google Scholar] [CrossRef]
- Mueller-Harvey, I.; Bee, G.; Dohme-Meier, F.; Hoste, H.; Karonen, M.; Kölliker, R.; Lüscher, A.; Niderkorn, V.; Pellikaan, W.F.; Salminen, J.-P.; et al. Benefits of condensed tannins in forage legumes fed to ruminants: Importance of structure, concentration, and diet composition. Crop. Sci. 2019, 59, 861. [Google Scholar] [CrossRef] [Green Version]
- Weston, L.A.; Skoneczny, D.; Weston, P.A.; Weidenhamer, J.D. Metabolic profiling: An overview—New approaches for the detection and functional analysis of biologically active secondary plant products. J. Allelochem. Interact. 2015, 1, 15–27. [Google Scholar]
- Williamson, L.N.; Bartlett, M.G. Quantitative liquid chromatography/time-of-flight mass spectrometry. Biomed. Chromatogr. 2007, 21, 567–576. [Google Scholar] [CrossRef]
- Nehybova, T.; Smarda, J.; Benes, P. Plant coumestans: Recent advances and future perspectives in cancer therapy. Anti Cancer Agents Med. Chem. 2014, 14, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Christ, B.; Hauenstein, M.; Hörtensteiner, S. A liquid chromatography–mass spectrometry platform for the analysis of phyllobilins, the major degradation products of chlorophyll in Arabidopsis thaliana. Plant. J. 2016, 88, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Liao, X.; Wood, C.M.; Xiao, C.-W.; Feng, Y.-L. A robust analytical method for measurement of phytoestrogens and related metabolites in serum with liquid chromatography tandem mass spectrometry. J. Chromatogr. B 2016, 1012–1013, 106–112. [Google Scholar] [CrossRef]
- Smith, J.; Jagusch, K.; Brunswick, L.; Kelly, R. Coumestans in lucerne and ovulation in ewes. N. Z. J. Agric. Res. 1979, 22, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Molyneux, R.J.; Ralphs, M.H. Plant toxins and palatability to herbivores. Rangel. Ecol. Manag. J. Range Manag. Arch. 1992, 45, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Oleszek, W.; Stochmal, A.; Janda, B. Concentration of isoflavones and other phenolics in the aerial parts of Trifolium species. J. Agric. Food Chem. 2007, 55, 8095–8100. [Google Scholar] [CrossRef]
- Sivesind, E.; Seguin, P. Effects of the environment, cultivar, maturity, and preservation method on red clover isoflavone concentration. J. Agric. Food Chem. 2005, 53, 6397–6402. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, K.; Khan, M.B.; Song, Y.Y.; Ijaz, M.; Luo, S.M.; Zeng, R.S. UV-irradiation enhances rice allelopathic potential in rhizosphere soil. Plant Growth Regul. 2013, 71, 21–29. [Google Scholar] [CrossRef]
- Iannucci, A.; Fragasso, M.; Platani, C.; Papa, R. Plant growth and phenolic compounds in the rhizosphere soil of wild oat (Avena fatua L.). Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Einhellig, F.A. Allelopathy—A natural protection, allelochemicals. In Handbook of Natural Pesticides: Methods; CRC Press: Boca Raton, FL, USA, 2018; pp. 161–200. [Google Scholar]
- Visnevschi-Necrasov, T.; Barreira, J.C.; Cunha, S.C.; Pereira, G.; Nunes, E.; Oliveira, M.B.P. Phylogenetic insights on the isoflavone profile variations in Fabaceae spp.: Assessment through PCA and LDA. Food Res. Int. 2015, 76, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Butkutė, B.; Padarauskas, A.; Cesevičienė, J.; Taujenis, L.; Norkevičienė, E. Phytochemical composition of temperate perennial legumes. Crop. Pasture Sci. 2018, 69, 1020–1030. [Google Scholar] [CrossRef]
- Tsao, R.; Papadopoulos, Y.; Yang, R.; Young, J.C.; McRae, K. Isoflavone profiles of red clovers and their distribution in different parts harvested at different growing stages. J. Agric. Food Chem. 2006, 54, 5797–5805. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, M.; Simon, J.E. Determination of isoflavones in red clover and related species by high-performance liquid chromatography combined with ultraviolet and mass spectrometric detection. J. Chromatogr. 2003, 1016, 195–209. [Google Scholar] [CrossRef]
- Butkutė, B.; Lemežienė, N.; Dabkevičienė, G.; Jakštas, V.; Vilčinskas, E.; Janulis, V. Source of variation of isoflavone concentrations in perennial clover species. Pharmacogn. Mag. 2014, 10, S181. [Google Scholar] [CrossRef] [Green Version]
- Zgórka, G. Ultrasound-assisted solid-phase extraction coupled with photodiode-array and fluorescence detection for chemotaxonomy of isoflavone phytoestrogens in Trifolium L. (Clover) species. J. Sep. Sci. 2009, 32, 965–972. [Google Scholar]
- Ramos, G.P.; Dias, P.M.; Morais, C.B.; Fröehlich, P.E.; Dall’Agnol, M.; Zuanazzi, J.A. LC determination of four isoflavone aglycones in red clover (Trifolium pratense L.). Chromatographia 2008, 67, 125–129. [Google Scholar] [CrossRef]
- Saviranta, N.M.; Anttonen, M.J.; von Wright, A.; Karjalainen, R.O. Red clover (Trifolium pratense L.) isoflavones: Determination of concentrations by plant stage, flower colour, plant part and cultivar. J. Sci. Food Agric. 2008, 88, 125–132. [Google Scholar] [CrossRef]
- Bennetau-Pelissero, C. Risks and benefits of phytoestrogens: Where are we now? Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Dubery, I.A.; Mienie, C. Tissue-specific expression of the chalcone synthase multigene family in Phaseolus vulgaris: Development of a RT-PCR method for the expression profiling of the chs isogenes. J. Plant Physiol. 2001, 158, 115–120. [Google Scholar] [CrossRef]
- Martin, C. Structure, function, and regulation of the chalcone synthase. In International Review of Cytology; Elsevier: Amsterdam, The Netherlands, 1993; Volume 147, pp. 233–284. [Google Scholar]
- Jung, W.; Yu, O.; Lau, S.-M.C.; O’Keefe, D.P.; Odell, J.; Fader, G.; McGonigle, B. Identification and expression of isoflavone synthase, the key enzyme for biosynthesis of isoflavones in legumes. Nat. Biotechnol. 2000, 18, 208. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Liu, C.; Jun, J.H. Metabolic engineering of anthocyanins and condensed tannins in plants. Curr. Opin. Biotechnol. 2013, 24, 329–335. [Google Scholar] [CrossRef]
- McMurray, C.H.; Laidlaw, A.S.; McElroy, M. The effect of plant development and environment on formononetin concentration in red clover (Trifolium pratense L.). J. Sci. Food Agric. 1986, 37, 333–340. [Google Scholar] [CrossRef]
- Kim, H.-K.; Jang, Y.-H.; Baek, I.-S.; Lee, J.-H.; Park, M.J.; Chung, Y.-S.; Chung, J.-I.; Kim, J.-K. Polymorphism and expression of isoflavone synthase genes from soybean cultivars. Mol. Cells 2005, 19, 67–73. [Google Scholar]
- Jonker, A.; Peiqiang, Y. The Role of Proanthocyanidins Complex in Structure and Nutrition Interaction in Alfalfa Forage. Int. J. Mol. Sci. 2016, 17, 793. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; McAllister, T.A.; Acharya, S. Condensed tannins in sainfoin: Composition, concentration, and effects on nutritive and feeding value of sainfoin forage. Crop. Sci. 2015, 55, 13–22. [Google Scholar] [CrossRef]
- Azuhnwi, B.N.; Boller, B.; Dohme-Meier, F.; Hess, H.D.; Kreuzer, M.; Stringano, E.; Mueller-Harvey, I. Exploring variation in proanthocyanidin composition and content of sainfoin (Onobrychis viciifolia). J. Sci. Food Agric. 2013, 93, 2102–2109. [Google Scholar] [CrossRef]
- Terrill, T.H.; Rowan, A.M.; Douglas, G.B.; Barry, T.N. Determination of extractable and bound condensed tannin concentrations in forage plants, protein concentrate meals and cereal grains. J. Sci. Food Agric. 1992, 58, 321–329. [Google Scholar] [CrossRef]
- Jackson, F.S.; McNabb, W.C.; Barry, T.N.; Foo, Y.L.; Peters, J.S. The Condensed Tannin Content of a Range of Subtropical and Temperate Forages and the Reactivity of Condensed Tannin with Ribulose- 1,5-bis-phosphate Carboxylase (Rubisco) Protein. J. Sci. Food Agric. 1996, 72, 483–492. [Google Scholar] [CrossRef]
- Johnson, M.T.; Smith, S.D.; Rausher, M.D. Plant sex and the evolution of plant defenses against herbivores. Proc. Natl. Acad. Sci. USA 2009, 106, 18079–18084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumner, L.; Paiva, N.; Dixon, R.; Geno, P. HPLC-Continuous-flow liquid secondary ion mass spectrometry of flavonoid glycosides in leguminous plant extracts. J. Mass Spectrom. 1996, 31, 472–485. [Google Scholar] [CrossRef]
- Neilson, E.H.; Goodger, J.Q.D.; Woodrow, I.E.; Møller, B.L. Plant chemical defense: At what cost? Trends Plant Sci. 2013, 18, 250–258. [Google Scholar] [CrossRef]
- Latif, S.; Gurusinghe, S.; Weston, P.A.; Quinn, J.C.; Piltz, J.W.; Weston, L.A. Metabolomic approaches for the identification of flavonoids associated with weed suppression in selected Hardseeded annual pasture legumes. Plant Soil 2019, 447, 199–218. [Google Scholar] [CrossRef]
- Dedio, W.; Clark, K. Biochanin A and formononetin content in red clover varieties at several maturity stages. Can. J. Plant Sci. 1968, 48, 175–181. [Google Scholar] [CrossRef]
- Zhou, Z.; Chen, X.; Zhang, M.; Blanchard, C. Phenolics, flavonoids, proanthocyanidin and antioxidant activity of brown rice with different pericarp colors following storage. J. Stored Prod. Res. 2014, 59, 120–125. [Google Scholar] [CrossRef]
- Qiu, Y.; Liu, Q.; Beta, T. Antioxidant properties of commercial wild rice and analysis of soluble and insoluble phenolic acids. Food Chem. 2010, 121, 140–147. [Google Scholar] [CrossRef]
- Min, B.; McClung, A.M.; Chen, M.H. Phytochemicals and antioxidant capacities in rice brans of different color. J. Food Sci. 2011, 76, C117–C126. [Google Scholar] [CrossRef]
- Sun, B.; Ricardo-da-Silva, J.M.; Spranger, I. Critical factors of vanillin assay for catechins and proanthocyanidins. J. Agric. Food Chem. 1998, 46, 4267–4274. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue Type | Pasture Species | Coumestrol | 4′-Methoxycoumestrol | Total Coumestans |
|---|---|---|---|---|
| Leaf | Arrowleaf clover | 14.7 ± 2.2 | 5.9 ± 1.5 | 20.6 ± 5.1 |
| Biserrula cv.* Casbah | 13.1 ± 2.7 | 5.2 ± 1.7 | 18.2 ± 6.5 | |
| Biserrula cv. Mauro | 14.6 ± 2.4 | 7.4 ± 5.5 | 22.0 ± 4.2 | |
| Bladder clover | 48.4 ± 10.6 | 24.8 ± 17.7 | 73.2 ± 13.6 | |
| French serradella | ND | ND | ND | |
| Gland clover | 27.1 ± 11.1 | 21.1 ± 5.6 | 48.2 ± 3.5 | |
| Lucerne | 31.8 ± 14.4 | 20.2 ± 7.5 | 52.0 ± 6.7 | |
| Subterranean clover | 25.1 ± 8.6 | 12.5 ± 4.5 | 37.6 ± 7.3 | |
| Yellow serradella | ND | ND | ND | |
| LSD (p < 0.05) | 9.3 | 5.4 | 13.1 | |
| Stem | Arrowleaf clover | 24.2 ± 5.3 | 6.1 ± 4.6 | 30.3 ± 10.5 |
| Biserrula cv. Casbah | ND | ND | ND | |
| Biserrula cv. Mauro | ND | ND | ND | |
| Bladder clover | 39.6 ± 7.7 | 18.7 ± 3.6 | 58.3 ± 12.1 | |
| French serradella | ND | ND | ND | |
| Gland clover | 26.2 ± 14.4 | 3.6 ± 1.9 | 29.8 ± 13.0 | |
| Lucerne | 36.6 ± 16.5 | 27.7 ± 12.8 | 64.3 ± 5.1 | |
| Subterranean clover | 25.4 ± 7.5 | 7.1 ± 1.6 | 32.5 ± 10.6 | |
| Yellow serradella | ND | ND | ND | |
| LSD (p < 0.05) | 9.4 | 14.5 | 17.5 | |
| Inflorescence | Arrowleaf clover | ND | 0.1 ± 0.1 | <0.1 |
| Biserrula cv. Casbah | ND | ND | ND | |
| Biserrula cv. Mauro | ND | 0.2 ± 0.1 | <0.1 | |
| Bladder clover | 0.2 ± 0.2 | <0.1 | <0.1 | |
| French serradella | ND | ND | ND | |
| Gland clover | 0.1 | ND | <0.1 | |
| Lucerne | 0.5 ± 0.1 | 0.2 ± 0.1 | 0.7 ± 0.1 | |
| Subterranean clover | 0.3 ± 0.1 | ND | 0.3 ± 0.1 | |
| Yellow serradella | ND | ND | ND | |
| LSD (p < 0.05) | 0.2 | 0.1 | 0.2 |
| Tissue Type | Pasture Species | Daidzein | Formononetin | Genistein | Total |
|---|---|---|---|---|---|
| Leaf | Arrowleaf clover | 55.1 ± 26.4 | 222.9 ± 84.4 | 28.3 ± 6.8 | 306.3 ± 94.3 |
| Biserrula cv. Casbah | 11.5 ± 3.5 | 73.6 ± 51.3 | 116.8 ± 2.8 | 192.5 ± 18.7 | |
| Biserrula cv. Mauro | 6.1 ± 0.7 | 58.9 ± 17.3 | 124.7 ± 12.5 | 189.7 ± 24.0 | |
| Bladder clover | 95.0 ± 2.1 | 152.3 ± 18.9 | 184.1 ± 26.3 | 431.4 ± 32.8 | |
| French serradella | 47.7 ± 12.5 | 35.5 ± 48.6 | 130.1 ± 7.6 | 113.3 ± 8.1 | |
| Gland clover | 120.3 ± 16.1 | 226.1 ± 20.5 | 97.5 ± 10.8 | 443.9 ± 61.4 | |
| Lucerne | 48.2 ± 13.3 | 329.4 ± 192.7 | 61.5 ± 10.1 | 439.1 ± 141.9 | |
| Subterranean clover | 109.1 ± 14.8 | 157.6 ± 47.3 | 72.1 ± 244.3 | 318.8 ± 40.5 | |
| Yellow serradella | 89.0 ± 13. | 91.6 ± 16.6 | 118.4 ± 5.4 | 219.1 ± 43.0 | |
| LSD (p < 0.05) | 24.9 | 37.5 | 34.8 | --- | |
| Stem | Arrowleaf clover | 9.8 ± 3.2 | 147.7 ± 58.2 | 27.6 ± 5.2 | 185.1 ± 67.1 |
| Biserrula cv. Casbah | 7.5 ± 0.9 | 23.9 ± 12.2 | 6.1 ± 1.9 | 37.5 ± 8.9 | |
| Biserrula cv. Mauro | 5.1 ± 1.4 | 60.4 ± 18.4 | 9.9 ± 3.7 | 75.4 ± 27.4 | |
| Bladder clover | 112.0 ± 33.5 | 829.8 ± 144.9 | 126.1 ± 5.4 | 968.1 ± 394.7 | |
| French serradella | 75.2 ± 23.3 | 96.7 ± 34.4 | 104.1 ± 18.9 | 276.0 ± 13.4 | |
| Gland clover | 3.4 ± 0.5 | 239.5 ± 18.4 | 88.1 ± 37.4 | 331.0 ± 107.0 | |
| Lucerne | 7.5 ± 2.3 | 120.6 ± 35.3 | 15.9 ± 5.1 | 144.0 ± 56.4 | |
| Subterranean clover | 107.8 ± 18.9 | 36.2 ± 6.4 | 18.2 ± 8.3 | 162.2 ± 42.4 | |
| Yellow serradella | 42.5 ± 7.2 | 77.7 ± 14.2 | 20.3 ± 6.9 | 140.5 ± 25.9 | |
| LSD (p < 0.05) | 20.8 | 47.9 | 19.8 | --- | |
| Inflorescence | Arrowleaf clover | 1.3 ± 0.3 | 98.2 ± 24.5 | 18.4 ± 4.6 | 117.9 ± 45.3 |
| Biserrula cv. Casbah | 2.2 ± 0.5 | 39.6 ± 9.9 | 10.2 ± 2.5 | 52.0 ± 17.6 | |
| Biserrula cv. Mauro | 1.7 ± 0.4 | 18.5 ± 4.6 | 9.6 ± 4.1 | 29.8 ± 7.5 | |
| Bladder clover | 19.3 ± 4.8 | 435.8 ± 108.9 | 158.9 ± 37.2 | 614.1 ± 190.6 | |
| French serradella | 10.5 ± 2.6 | 98.8 ± 24.7 | 19.9 ± 4.9 | 129.2 ± 43.4 | |
| Gland clover | 68.3 ± 17.1 | 184.8 ± 46.2 | 68.3 ± 17.1 | 321.0 ± 60.1 | |
| Lucerne | 25.5 ± 6.4 | 182.6 ± 45.6 | 25.7 ± 6.4 | 233.8 ± 81.1 | |
| Subterranean clover | 128.9 ± 32.2 | 187.6 ± 46.9 | 148.7 ± 39.8 | 465.2 ± 26.3 | |
| Yellow serradella | 14.6 ± 3.6 | 17.6 ± 4.4 | 25.8 ± 6.6 | 58.00 ± 5.2 | |
| LSD (p < 0.05) | 16.8 | 39.2 | 24.5 | --- |
| Pasture Species | TPC | TPAC | ||
|---|---|---|---|---|
| Extractable | Bound | Extractable | Bound | |
| Arrowleaf clover | 10.50b | 0.24de | 3.71c | ND |
| Biserrula cv. Casbah | 7.69cde | 0.43cd | 6.49a | 8.87a |
| Biserrula cv. Mauro | 6.24def | 0.11e | 4.29b | 4.06b |
| Bladder clover | 6.67def | 0.45bc | 1.73c | ND |
| French serradella | 8.29bcd | 0.35cd | 1.94c | ND |
| Gland clover | 13.84a | 0.57bc | 2.12c | ND |
| Lucerne | 4.40f | 0.11de | 2.05c | 0.33c |
| Subterranean clover | 9.39bc | 0.62bc | 4.52ab | ND |
| Yellow serradella | 5.17ef | 0.72b | 1.94c | ND |
| LSD | 2.62 | 0.28 | 2.16 | 1.95 |
| Scientific Name | Common Name | Cultivar |
|---|---|---|
| Trifolium vesiculosum Savi. | Arrowleaf clover | Cefalu |
| Biserrula pelecinus L. | Biserrula | Casbah, Mauro |
| Trifolium spumosum L. | Bladder clover | Bartolo |
| Ornithopus sativus Brot. | French serradella | Margarita |
| Trifolium glanduliferum Boiss. | Gland clover | Prima |
| Trifolium subterraneum L. | Subterranean clover | Seaton Park |
| Ornithopus compressus L. | Yellow serradella | Santorini |
| Medicago sativa L. | Lucerne | Aurora |
| Name | Molecular Formula | M + H | Basis for Identification a |
|---|---|---|---|
| Isoflavones | |||
| Daidzein | C15H10O4 | 255.0652 | STD |
| Formononetin | C16H12O4 | 269.0808 | STD |
| Genistein | C15H10O5 | 271.0601 | STD |
| Coumestans | |||
| Coumestrol | C15H8O5 | 269.0444 | STD |
| 4′-methoxycoumestrol | C16H10O5 | 283.0579 | AM |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latif, S.; Weston, P.A.; Barrow, R.A.; Gurusinghe, S.; Piltz, J.W.; Weston, L.A. Metabolic Profiling Provides Unique Insights to Accumulation and Biosynthesis of Key Secondary Metabolites in Annual Pasture Legumes of Mediterranean Origin. Metabolites 2020, 10, 267. https://doi.org/10.3390/metabo10070267
Latif S, Weston PA, Barrow RA, Gurusinghe S, Piltz JW, Weston LA. Metabolic Profiling Provides Unique Insights to Accumulation and Biosynthesis of Key Secondary Metabolites in Annual Pasture Legumes of Mediterranean Origin. Metabolites. 2020; 10(7):267. https://doi.org/10.3390/metabo10070267
Chicago/Turabian StyleLatif, Sajid, Paul A. Weston, Russell A. Barrow, Saliya Gurusinghe, John W. Piltz, and Leslie A. Weston. 2020. "Metabolic Profiling Provides Unique Insights to Accumulation and Biosynthesis of Key Secondary Metabolites in Annual Pasture Legumes of Mediterranean Origin" Metabolites 10, no. 7: 267. https://doi.org/10.3390/metabo10070267