Feeding Preference of Crapemyrtle Bark Scale (Acanthococcus lagerstroemiae) on Different Species
by

Runshi Xie
1, 
Bin Wu
1,
Haijie Dou
1,2,
Cuiyu Liu
3,
Gary W. Knox
4,
Hongmin Qin
5,* and
Mengmeng Gu
6,*
1
Department of Horticultural Sciences, Texas A&M University, College Station, TX 77843, USA
2
Department of Science & Technology Development, Beijing Industrial Technology Research Institute, No.5, Xingguang 4th Ave, Tongzhou District, Beijing 101111, China
3
College of Forestry, Nanjing Forestry University, 159 Longpan Rd., Nanjing 210037, China
4
Department of Environmental Horticulture, University of Florida/IFAS North Florida Research and Education Center, Quincy, FL 32351, USA
5
Department of Biology, Texas A&M University, College Station, TX 77840, USA
6
Department of Horticultural Sciences, Texas A&M AgriLife Extension Service, College Station, TX 77843, USA
*
Authors to whom correspondence should be addressed.
Insects 2020, 11(7), 399; https://doi.org/10.3390/insects11070399
Submission received: 2 June 2020
/
Revised: 23 June 2020
/
Accepted: 25 June 2020
/
Published: 28 June 2020
Abstract
:Crapemyrtle bark scale (CMBS; Acanthococcus lagerstroemiae) is an exotic pest species that causes aesthetic and economic damage to crapemyrtles and poses potential threats to other horticultural crops in the United States. Although previous studies reported the infestation of CMBS on several alternative hosts across multiple families in Asia, its potential threats to other documented alternative hosts remain elusive and yet to be confirmed. In this study, feeding preference studies of CMBS were conducted on forty-nine plant species and cultivars in 2016 and 2019, in order to gain insight into the expansion of CMBS distribution in the United States, as well as other regions of the world. The infestations of CMBS were confirmed on apple (Malus domestica), Chaenomeles speciosa, Disopyros rhombifolia, Heimia salicifolia, Lagerstroemia ‘Spiced Plum’, M. angustifolia, and twelve out of thirty-five pomegranate cultivars. However, the levels of CMBS infestation on these test plant hosts in this study is very low compared to Lagerstroemia, and may not cause significant damage. No sign of CMBS infestation was observed on Rubus ‘Arapaho’, R. ‘Navaho’, R. idaeus ‘Dorman Red’, R. fruticosus, B. microphylla var. koreana × B. sempervirens, B. harlandii, or D. virginiana.
1. Introduction
Crapemyrtle bark scale (CMBS; Acanthococcus lagerstroemiae Borchsenius, 1960) originates from East Asia, and has been reported to infest important horticultural crops, such as crapemyrtles and pomegranates, in Japan, Korea, India, and China [1,2]. Since its first discovery in the United States in 2004 in Richardson, TX, the infestation of this scale insect has been confirmed in Alabama, Arkansas, Georgia, Kansas, Louisiana, Maryland, Mississippi, New Mexico, North Carolina, Oklahoma, Tennessee, Virginia [3], and Washington [4]. The most common host of CMBS, crapemyrtles (Lagerstroemia spp.), is an important landscape tree in the southern United States, which generates a combined market value (wholesale) of around USD 67 million per year [5]. The wide use of crapemyrtle could be due to its various growth patterns and plant architectures, flower color and duration, attractive bark features, as well as disease and pest tolerance [6]. However, infestation by CMBS can greatly affect the performance of crapemyrtles, causing aesthetic and economic damage. In addition, the widespread distribution of this exotic pest species also poses threats to other potential alternative host crops, especially native plant species such as American beautyberry (Callicarpa americana) in the United States [4].
Crapemyrtle bark scale is a phloem-feeding hemipteran. The development of CMBS is categorized as of incomplete metamorphosis, with a life cycle of 56 to 83 days [7]. There are between two to four generations of CMBS per year, depending on the climate zones [2,8]. In the United States, up to four generations were observed in the field in Dallas, TX [2]. Under greenhouse conditions, the development of males involves five nymphal stages, but only three for females [9]. The relatively small size of CMBS (ranging from 0.3 mm to 3 mm, depending on the developmental stages) renders difficulties for detection of the insect in the early infestation.
The damage caused by CMBS includes slow and weakened plant growth, leaf abscission, absence of flowering, death of branches, and, in severe cases, the death of the juvenile seedling or young crapemyrtle trees [10]. Another major issue with CMBS infestation is the accumulation of black sooty mold, which is caused by the honeydew secretion when CMBS nymphs actively feed on the plant [2,11]. Common treatments to control CMBS include the application of horticultural oil on the bark surface and root injection or drench of systemic insecticides [8,12]; however, the lack of information on insect biology of CMBS and the population dynamics related to its adaptation in the United States is an obstacle in managing this pest insect, as it continues to spread among southeastern United States [13]. No host resistance to CMBS has been reported among Lagerstroemia species and cultivars, though different levels of infestations of CMBS can often be observed in the field, which can be caused by many environmental factors, such as temperature, shade, humidity, and the presence of natural enemies [11,14,15].
Abiotic and biotic factors are known to have impacts on insect biology in terms of altering development duration and sex ratio. For example, green pea aphid (Myzus persicae) showed varied development durations under different temperature conditions [16]. Similarly, altered development duration and sex ratio were observed on parasitoid Campoletis chlorideae at different temperatures [17]. On the other hand, the development of CMBS could be greatly affected by the direct interaction between the insect and plant host, including the CMBS feeding preference and the potential insect tolerance from the plant [18], which determines the severity of infestation.
Alternative host selection and feeding preference, as important aspects of insect biology, are valuable information in terms of understanding insect population dynamics, and how the insect disperses geographically [19,20,21]. While many insect herbivores are considered specialists, others are known to maintain the ability to feed on a wide range of plant hosts, or switch hosts, as an adaptive strategy when facing adverse environments [22,23]. For example, hemipteran Thaumastocoris peregrinus specialized in infesting host plants within Eucalyptus [24]. Although most species of aphids (Aphididae) are known to be specialized on one certain host plant [25], many are still capable of feeding on multiple plant hosts [26,27,28]. Economically important aphid species, including Aphis fabae [29], Aphis nasturtii [30], Aphis aurantia [31], Aulacorthum solani [32], Brachycaudus helichrysi [33], Macrosiphum euphorbiae [34], Myzus ornatus [35], Myzus persicae [36] are considered polyphagous. Some members of Aphididae further exhibit complex life histories involving alternation of feeding between unrelated plant taxa [37]. Thus, alternative hosts remain important for the survival and perpetuation of many insect species [38,39].
In the case of CMBS, previous reports from Asia have indicated that its host range includes apple (Malus pumila), axlewood (Anogeissus latifolia), border privet (Ligustrum obtusifolium), boxwood (Buxus microphylla), Chinese hackberry (Celtis sinensis), Chinese-quince (Pseudocydonia sinensis), common fig (Ficus carica), food wrapper plant (Mallotus japonicus), Indian rosewood (Dalbergia eremicola), needlebush (Glochidion puberum), persimmon (Diospyros kaki), pomegranate (Punica granatum), and soybean (Glycine max) [4]. In addition, as documented for many insect herbivores, including scale insects [40,41], the feeding preference of CMBS also has the potential for being altered once the infestation has been successfully established on a novel host. Therefore, the study of feeding preferences and host range is crucial in the prediction and prevention of the expansion of CMBS population.
Naturally occurring CMBS infestation has been reported on American beautyberry (C. americana) [4], crapemyrtle, Hypericum [42], and spirea (Spiraea) [43] in the United States. Wang, et al. (2019) reported four more alternative hosts, including henna (Lawsonia inermis), pomegranate (‘Wonderful’), sinicuichi (Heimia salicifolia Link), and winged loosestrife (Lythrum alatum Pursh), from a host range test in the United States. However, many other horticultural crops of great economic importance, such as Chaenomeles and additional species of Buxus, Diospyros, Malus, and Rubus, have yet to be tested as potential hosts in the United States. In a preliminary test, gravid females were found to develop on ‘Fuji’ apple seedlings when being tied with a CMBS-infested crapemyrtle branch. In addition, as a primary host of CMBS with high market value, pomegranates have been reported to suffer significant level of damage in terms of CMBS infestation in China [44], yet many other common pomegranate cultivars remain untested for their susceptibilities to CMBS.
Buxus, Chaenomeles, Diospyros, Malus, Punica, and Rubus are valuable fruit or ornamental crops in the United States, as well as other parts of the world. Boxwood, or Buxus, is one of the best-selling woody plants in the United States [5]. Diospyros, commonly known as persimmon, is the largest genus in Ebenaceae, with more than 500 species of valuable crops with versatile uses with utilizations as edible fruit, timber, ornamental plants, and medicinal usage in east Asia [45,46]. In the United States, D. virginiana is known as a native species and is cultivated as a fruit crop [47]. Malus domestica, commonly known as apple, is considered one of the most valuable fruit crops in the United States [48]. The United States is one of the largest producers of apple, second to China, in the world [49]. Blackberries and raspberries, commonly referred to as brambles, are also highly valued fruit crops in the genus Rubus. In the United States, California, Oregon, and Washington are three of the leading producers of blackberries and raspberries [50].
Finally, Punica, or pomegranate, is a species of fruit crop that has been highly valued as food and for medicinal purposes throughout human history [51]. Some cultivars of pomegranates have been developed as ornamental plants [52,53]. Pomegranates are known to be native to central Asia, while years of cultivation has spread their distribution to a wide range of geographical regions including Europe and the United States [51]. According to latest data, the United States National Clonal Germplasm Repository in Davis, California, is currently maintaining 194 accessions of pomegranates, 82 of which were cultivars originated from the United States [54].
Potential pitfalls and challenges in pomegranate production involve damages and diseases related to insects, fungi, and bacteria [51]. Arthropod pests such as the black borer Apate monachus [55] and aphids [51], are known to cause major damage on pomegranate production. Scale insects, including CMBS, have been reported to be an issue and caused severe damage in pomegranate production in China [44]. In fact, CMBS is also referred to as pomegranate felt scale in China [56]. The infestations of CMBS were found on different cultivars of pomegranate, with various sizes ranging from container plants [57], to 8–10-year-old pomegranate trees at a nursery [44]. Wang et al. 2019 have confirmed P. granatum ‘Wonderful’, one of the most popular cultivars in the United States [58], as a suitable host for CMBS, however, the threat of CMBS to other pomegranates in the U.S. remains undetermined.
Plant species and cultivars within the genus of Buxus, Diospyros, Malus, Punica, and Rubus have been previously reported as the potential alternative hosts of CMBS outside of the United States [4,59,60,61], which leads to concerns for their production and utilization in the United States, as well as other regions of the world. Therefore, it was hypothesized that plant species from these genera are potentially susceptible to CMBS attack, and thus selected for evaluation in CMBS feeding trails. Three independent feeding studies were designed to investigate the ability of CMBS to infest plant species within selected genera, along with its feeding preference.
In 2016 and 2019, two CMBS feeding trials were conducted to evaluate CMBS preference among 35 available pomegranate cultivars in the United States. Another feeding study was conducted in 2019 to evaluate the host preference of CMBS on plant species and cultivars within seven genera. In summary, a total of 49 plant species or cultivars from Buxus, Chaenomeles, Diospyros, Heimia, Malus, Punica, and Rubus were evaluated for their susceptibility to CMBS. This study aims to expand the current knowledge on the host range of CMBS in the United States, and help the development of an effective integrated pest management program for controlling this pest insect.
2. Materials and Methods
2.1. Insect Source and Plant Material
Insect samples used in three feeding studies were collected prior to inoculation from June to September in 2016, and from May to June in 2019. The branches/twigs infested with CMBS were collected from crapemyrtle trees on campus (Texas A&M University, College Station, TX), and stored in zip-lock bags at room temperature (25 °C). The CMBS collected from infested plants were used for the experiments immediately, or within one or two days after the collection.
For pomegranate feeding preference trials, 30 cultivars were obtained from University of Florida and Texas A&M AgriLife Research & Extension Center at Uvalde, TX, and evaluated in 2016 (Table 1). In 2019, cuttings from 14 pomegranate (P. granatum) cultivars (collected from Texas A&M AgriLife Research & Extension Center, Uvalde, TX, USA) were rooted in tree tubes and transplanted in containers (3.79 L) for the CMBS feeding preference trial (Table 1).
In 2019, a separate host range confirmation study was conducted to evaluate the feeding preference of CMBS on seven plant genera. Candidate plant species were selected from previously reported CMBS hosts outside of the United States. Commercially available crapemyrtle hybrid cultivar Lagerstroemia ‘Spiced Plum’, which could manifest a typical heavy infestation when under CMBS attack, was thus utilized as a control. Fourteen plant species and cultivars from seven genera, including Buxus harlandii, B. microphylla var. koreana × B. sempervirens ‘Green Gem’, Chaenomeles speciosa ‘Texas Scarlet’, Diospyros rhombifolia, D. virginiana, Heimia salicifolia, L. ‘Spiced Plum’, Malus angustifolia, M. domestica ‘Fuji’, M. domestica ‘Red Delicious’, Rubus ‘Arapaho’, R. fruticosus ‘Prime Ark Freedom’, R. idaeus ‘Dorman Red’, and R. ‘Navaho’, grown in containers (3.79 L), were used in the experiment (Table 2).
2.2. Pomegranate Feeding Trials
Two independent feeding trials were conducted at an outdoor container nursery in 2016 and in a greenhouse (25 ± 5 °C, 50 ± 10% relative humidity) in 2019, respectively, to evaluate the cultivar preferences of CMBS among a total of 35 pomegranate cultivars. Both trials were located on Texas A&M University campus (30°36′31.9″ N, 96°21′01.7″ W).
For the 2016 trial, 30 cultivars with 5 replications were tested (Table 2). Three rounds of inoculations were conducted on 6 June 2016, 22 July 2016, and 30 September in 2016, respectively. For the first and second inoculations, CMBS ovisacs were removed from infested plants and placed onto all test cultivars. For the third inoculation, One CMBS-infested branch (10-cm in length) per plant was used to inoculate all tested cultivars. Crapemyrtle bark scale infestation was recorded around one month after the third inoculation. For the 2019 trial, 14 cultivars with 5 replications were evaluated (Table 2). Two rounds of inoculation using one CMBS infested branch (10 cm in length) were conducted on 15 May 2019 and 25 June 2019, respectively. The number of male pupae and gravid females were recorded on 10 August 2019.
Male pupae and gravid females, recognized as white felt-like coverings or ovisacs, were distinguished by their size and shape. Male pupae were identified as white sacs with an elongated shape (around 1 mm long, and 0.5 wide), while gravid females were identified as white ovisacs with a much larger size (around 2 mm long, and 1 mm wide) and rounded shape [4].
2.3. Host Range Confirmation on Seven Genera
Plant chambers (127 × 127 × 127 cm) were constructed using ½ inch PVC pipes as a frame, and enclosed by Chiffon mesh netting (Fabric Wholesale Direct, Farmingdale, NY 11735). Zippers were sewn onto the mesh fabric to form openings for watering and moving the plants. All plant chambers were placed inside a greenhouse (25 ± 5 °C, 50 ± 10% relative humidity), located on Texas A&M University campus (30°36′31.9″ N, 96°21′01.7″ W).
Plants were placed inside the plant chambers, before inoculating with CMBS with one set of fourteen plant species and cultivars per chamber. Each set of plants was replicated three times. One infested crapemyrtle branch with all but five fresh ovisacs removed was attached to each test plant, on 13 May and 15 June, 2019, respectively, to ensure the successful inoculation and survival of CMBS on test plants. The numbers of male pupae and gravid females per plant were recorded biweekly from June to September, and monthly from October to December 2019.
2.4. Data Analysis and Statistics
Data on the 2016 pomegranate trial was not statistically analyzed, because ovisacs were only found on two plants. In 2019, one-way analysis of variance (ANOVA) was used to analyze the number of male pupae and gravid females found on 14 pomegranate cultivars. Sex ratio (male to female ratio) was calculated using the mean number of male pupae divided by gravid females for each plant species and cultivars. Log transformed of the original data (No. of CMBS +1) was performed to improve normality of data distribution. Data for the number of male pupae and gravid females on seven genera from June to December 2019 were analyzed using multivariate analysis of variance (MANOVA) for repeated measures of all infested plant species and cultivars. Univariate split-plot approach ANOVA was also conducted for the numbers of male pupae and gravid females recorded on all infested plant species and cultivars, and multiple means were compared using All Pair, Tukey HSD with JMP software (JMP Pro14, Statistical Analysis System, Cary, NC, USA).
3. Results
3.1. Pomegranate Feeding Trials
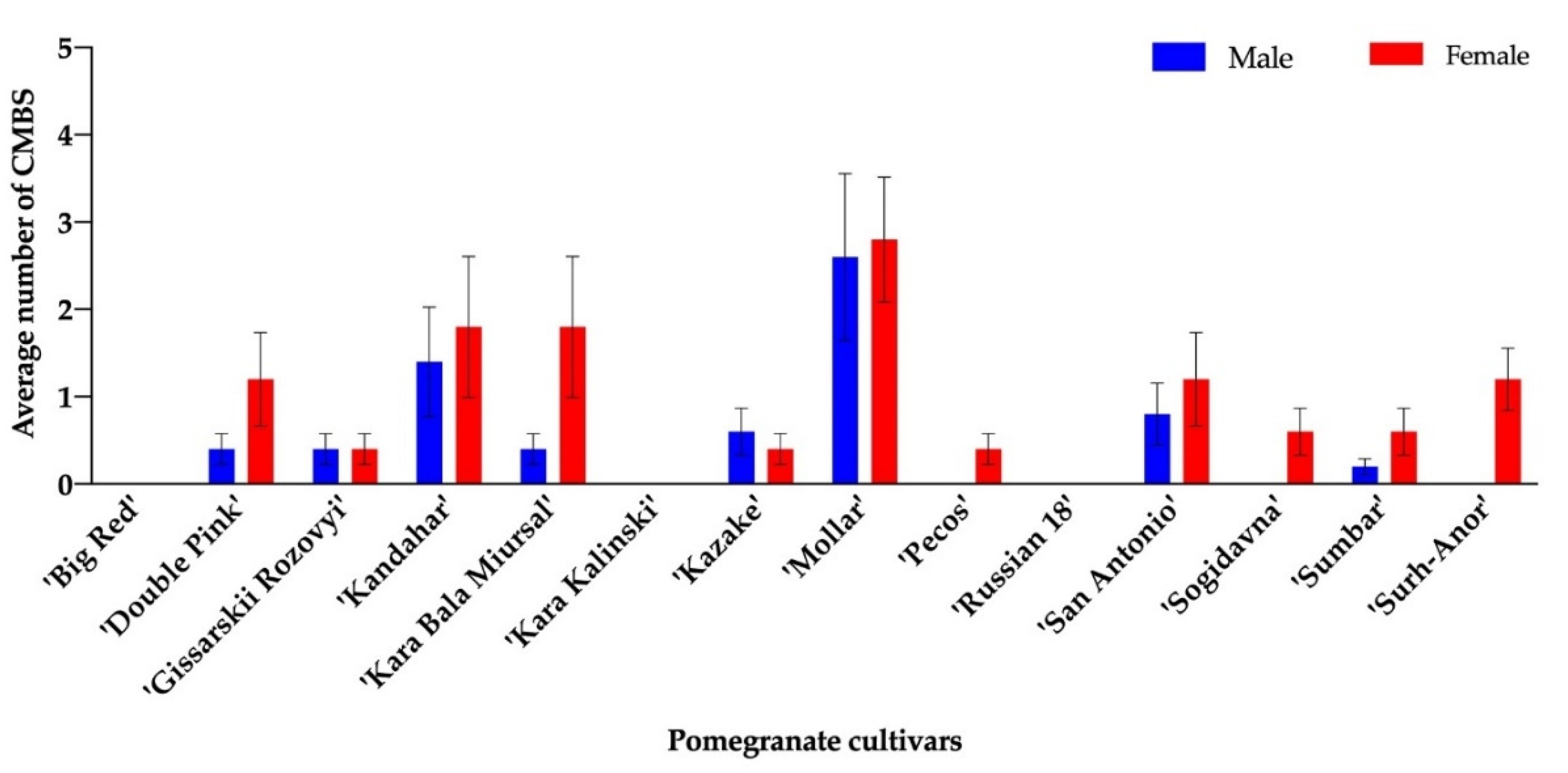
For the 2016 trial, only one gravid female was found on ‘Angel Red’ and one on ‘Sumbar’. No male pupa was found on any cultivar evaluated. For the 2019 trial, male pupae were found on ‘Double Pink’, ‘Gissarskii Rozovyi’, ‘Kandahar’, ‘Kara Bala Miursal’, ‘Kazake’, ‘Mollar’, ‘San Antonio’, and ‘Sumbar’, and gravid females were found on ‘Double Pink’, ‘Gissarskii Rozovyi’, ‘Kandahar’, ‘Kara Bala Miursal’, ‘Kazake’, ‘Mollar’, ‘Pecos’, ‘San Antonio’, ‘Sogidavna’, ‘Sumbar’, and ‘Surh-Anor’. No sign of CMBS infestation was observed on ‘Big Red’, ‘Kara Kalinski’, and ‘Russian 18’ in 2019 (Figure 1). Although the majority of cultivars included in this trial had CMBS infestations, the number of gravid females were less than 10 on all tested cultivars. The highest number of gravid females were observed for ‘Kandahar’, ‘Kara Bala Miursal’, and ‘Mollar’ (Figure 1). No significant difference in terms of CMBS infestation was found among all test cultivars in 2019.
3.2. Host Range Confirmation on Seven Genera
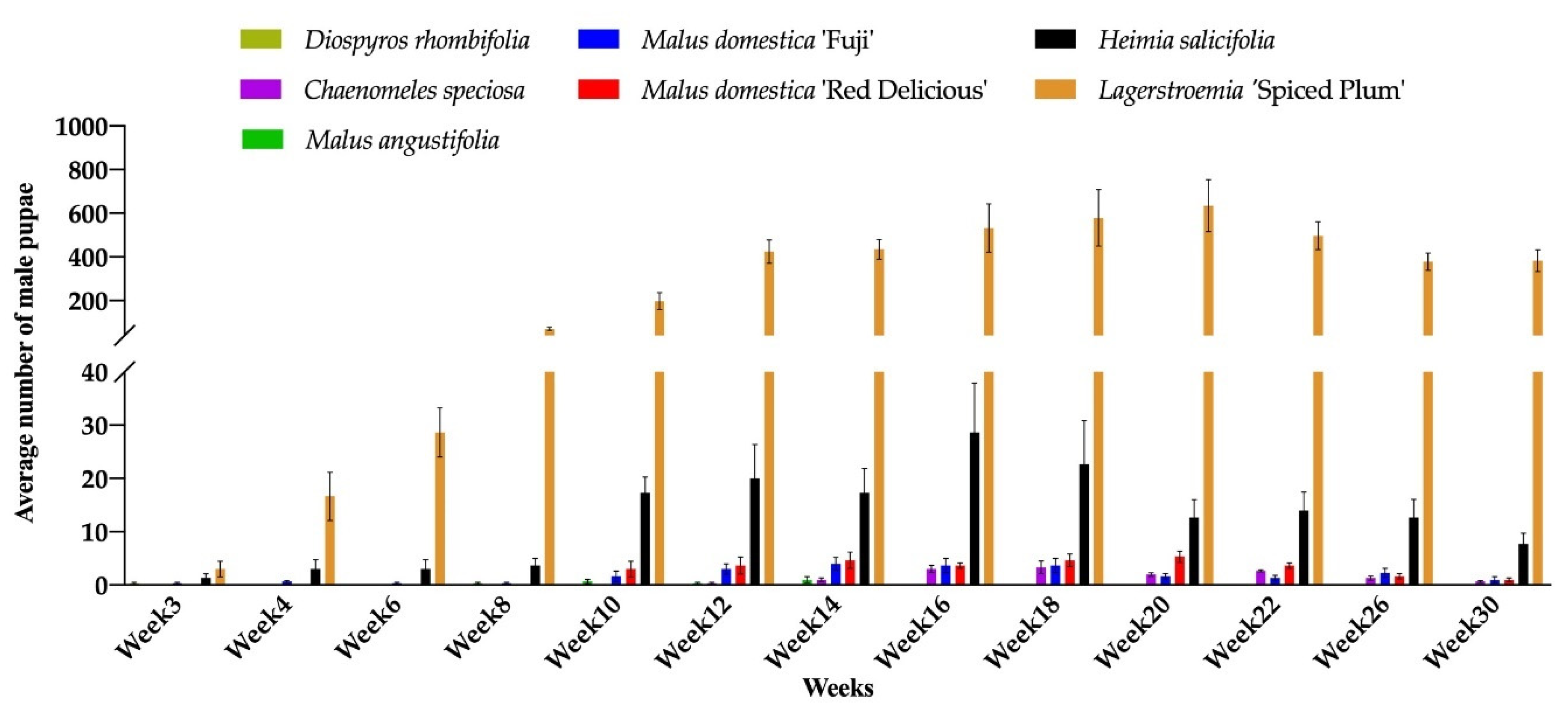
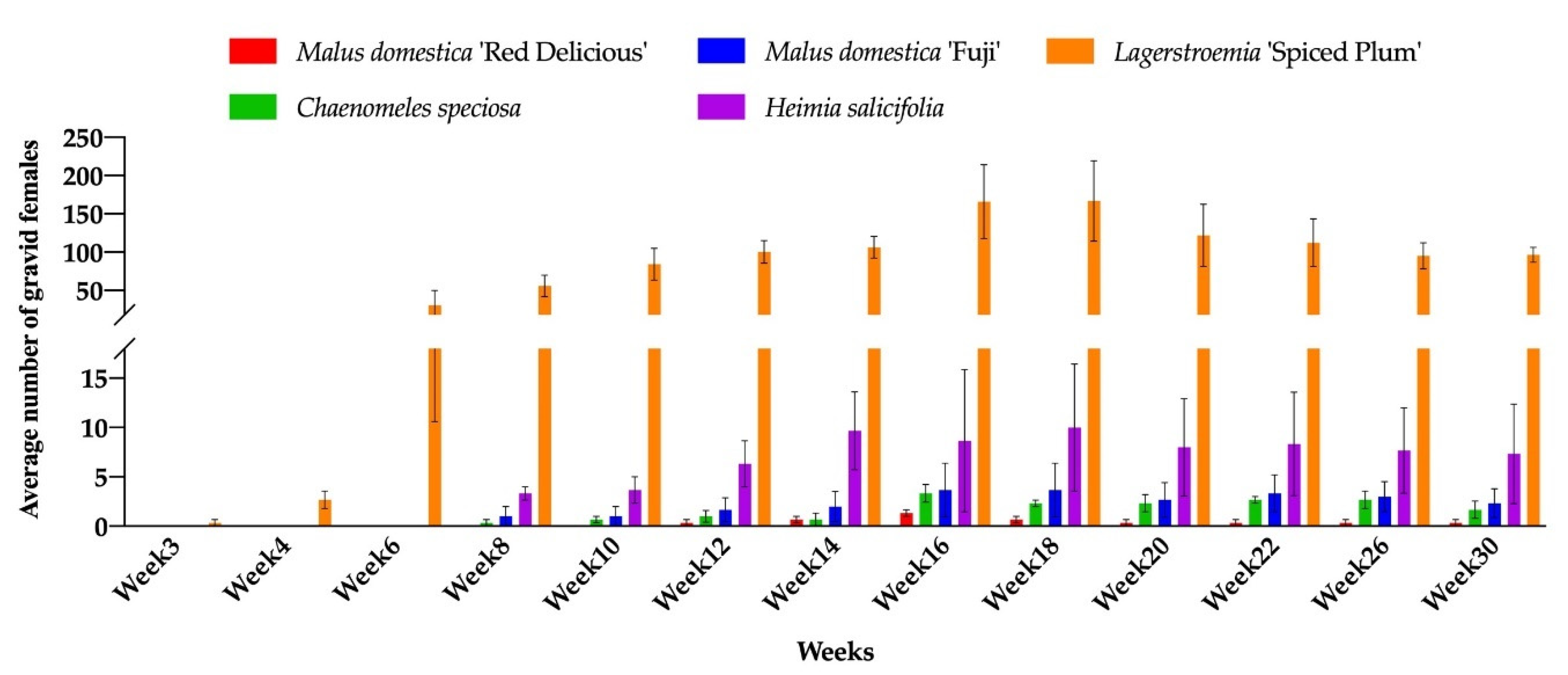
During the 30 weeks (May to December 2019) of the experiment, CMBS male pupae or gravid females were observed on seven out of 14 test plant species (C. speciosa, D. rhombifolia, H. salicifolia, L. ‘Spiced Plum’, M. angustifolia, M. domestica ‘Fuji’, and M. domestica ‘Red Delicious’). Only male pupae and no gravid female were found on D. rhombifolia and M. angustifolia (Figure 2 and Figure 3). The number of male pupae and gravid females varied significantly across the entire experiment, and the effect of data collection time (Time, p < 0.001), plant species and cultivars (Plant species, p < 0.001), and the Time by Plant species interactions (p < 0.001) were identified for the population dynamics of CMBS. The CMBS populations on all infested plant species and cultivars showed peaks between week 16 to 20 (September 2019) after inoculation (Figure 2 and Figure 3). The number of CMBS was highest on the control plant (L. ‘Spiced Plum’) in September (Table 3), and throughout the entire experiment (Table 4), compared to the other infested plant species and cultivars. The number of male pupae on ‘Spiced Plum’ after week 10 showed a significant increase, and remained the highest for the remainder of the experiment (Table S1). Similarly, the number of gravid females on ‘Spiced Plum’ after week 10 were higher than all the other test plants for the remainder of the experiment (Table S2).
3.3. The Effects of Plant Hosts on Insect Development
No male pupae and gravid females were observed on B. harlandii, B. microphylla var. koreana × B. sempervirens, D. virginiana, Rubus ‘Arapaho’, R. fruticosus, R. idaeus ‘Dorman Red’, and R. ‘Navaho’, The male pupae were first observed on D. rhombifolia, H. salicifolia, L. ‘Spiced Plum’, and M. domestica ‘Fuji’ in week 3. The gravid females were first observed on L. ‘Spiced Plum’ in week 3. The delayed onset of infestation was observed for C. speciosa, H. salicifolia, M. domestica ‘Fuji’, and M. domestica ‘Red Delicious’. The first detection of gravid females on C. speciosa, H. salicifolia, and M. domestica ‘Fuji’ was 8 weeks after inoculation (WAI), and 12 WAI on M. domestica ‘Red Delicious’, compared to 3 WAI of the first detection on L. ‘Spiced Plum’ (Figure 3). The different onset of infestation could be due to the varied development durations of nymphal stages, which shows the different level of adaptive behavior of CMBS towards novel hosts.
The highest mean number of male pupae and gravid females on L. ‘Spiced Plum’ was 634.3 ± 206.4 (mean ± SE) male pupae at week 20 and 167.0 ± 52.5 (mean ± SE) gravid females at week 18, respectively. Both male pupae and gravid females were observed on C. speciosa, H. salicifolia, M. domestica ‘Fuji’, and M. domestica ‘Red Delicious’, with lower levels of infestation compared to Lagerstroemia. In addition to L. ‘Spiced Plum’, the highest mean number of male pupae recorded during the experiment was 3.3 ± 2.0 (mean ± SE) on C. speciosa, 28.7 ± 15.9 (mean ± SE) on H. salicifolia, 4 ± 2.1 (mean ± SE) on M. domestica ‘Fuji’, and 5.3 ± 1.9 (mean ± SE) on M. domestica ‘Red Delicious’, respectively. The highest mean number of gravid females recorded during experiment was 3.3 ± 0.9 (mean ± SE) on C. speciosa, 10.0 ± 6.4 (mean ± SE) on H. salicifolia, 3.7 ± 2.7 (mean ± SE) on M. domestica ‘Fuji’, and 1.3 ± 0.3 (mean ± SE) on M. domestica ‘Red Delicious’, respectively. Only a few male pupae were observed on D. rhombifolia (0.3 at week 3) and M. angustifolia (1 at week 14) (Figure 2). The effect of host plant in altering insect sex differentiation was observed in this study, as the male to female ratios on C. speciosa, H. salicifolia, L. ‘Spiced Plum’, M. domestica ‘Fuji’, and M. domestica ‘Red Delicious’ was 0.81:1, 2.25:1, 3.67:1, 0.99:1, and 7.23:1 respectively, throughout the experiment.
4. Discussion
The pomegranate feeding trials conducted in 2016 and 2019 showed very low levels of CMBS infestation on all tested pomegranate cultivars. Lack of infestation on almost all pomegranate plants could be due to the outdoor conditions. The results from 2019 feeding trial was similar to results from Wang et al. 2019, who showed a low level of infestation (around 10 gravid females found) on ‘Wonderful’ pomegranate. Although CMBS has been reported to cause serious damage to pomegranates in China [44], our results did not indicate a level of damage on the tested cultivars comparable to crapemyrtles. While our result did confirm that pomegranates serve as alternative hosts for CMBS, it is also not uncommon that the severity of damages caused by the same pest insect varies on different genotypes [51].
The geographical origin of the pomegranate cultivars might play a role in determining the susceptibility of host plants to CMBS. According to online database [54,62], the 35 tested pomegranate cultivars in 2016 and 2019 have origins in either Georgia, India, Iran, the Soviet Union or Russia, the United States, or Turkmenistan. No tested cultivar was known to be from China, which suggests that the reported heavy CMBS infestations on pomegranates may be confined to certain pomegranate species or cultivars developed in China.
As an indicator for successful reproduction, gravid female and oviposition are often used to evaluate the performance of an insect on novel hosts [63,64,65]. The varied WAI for the first gravid female found on C. speciosa, H. salicifolia, M. domestica ‘Fuji’, and M. domestica ‘Red Delicious’, suggest that the developmental times from nymph to gravid female were at least twice as long compared to L. ‘Spiced Plum’. However, the possibility remains for CMBS to adapt and alter feeding preference on novel hosts after several generations or through mutations, as similar phenomena have been reported on other insect species [66,67,68]. The number of gravid females found on H. salicifolia was similar to the number reported by Wang et al., 2019 (less than 50 gravid females), while the sex ratio was unknown previously.
No gravid female was observed on B. harlandii, B. microphylla var. koreana × B. sempervirens, D. rhombifolia, D. virginiana, M. angustifolia, R. ‘Arapaho’, R. fruticosus, R. idaeus ‘Dorman Red’, or R. ‘Navaho’ in our current study, which might conclude that these species are not suitable hosts for CMBS. However, previous reports [59,61] reported that several plant species in Buxus, Diospyros, and Rubus were CMBS hosts. Similar phenomenon was observed in our study, where gravid females were found on M. domestica (originated from Central Asia) [69], but not on M. angustifolia (native to the North America) [70]. This suggests that CMBS have different feeding preference toward plants within the same genus, and higher host acceptance might be found especially on certain plant species or cultivars developed outside of the United States.
Crapemyrtle bark scale infestation differed greatly on different crapemyrtle species and cultivars. According to Wang et al. (2019), the highest number of gravid females on L. indica × fauriei ‘Natchez’ was higher (482 ± 92) than the highest number found on L. ‘Spiced Plum’ in this study. Based on current observations, the highest number of male pupae can reach near 1000 on L. limii and L. subcostata grown in 3.79 L containers, while, under similar conditions, the number of pupae were found to range from 11 to around 400 on L. fauriei ‘Kiowa’, L. caudata, L. indica ‘Dynamite’, and L. speciosa. For gravid females, the highest number was found to be around 600 on L. limii grown in 3.79 L containers, while, under similar conditions, the highest number was between 200 and 400 on L. subcostata and L. fauriei ‘Kiowa’, and the highest number was below 100 on L. caudata, L. indica ‘Dynamite’, and L. speciosa [71]. Thus, we found that the highest number of male pupae and gravid females were 634 and 167, respectively, on L. ‘Spiced Plum’ grown in 3.79 L containers, which suggests that the level of possible CMBS infestation of ‘Spiced Plum’ would be considered ‘medium’, among other tested crapemyrtle species and cultivars.
The different infestation among crapemyrtle species and cultivars could be related to several factors, including the predation from nature enemy, or certain plant physical or biochemical properties. In this study, the influence of a natural enemy was mostly eliminated, except for the pomegranate feeding trial conducted in 2016, where the experiment was conducted in an outdoor condition. Interestingly, the 2016 experiment showed very limited presence of CMBS on tested cultivars, which suggests that conducting experiment in either a greenhouse or enclosed plant chamber could be effective in excluding the factor of natural enemies. Without the influence of natural enemies, the factors influencing CMBS population or host selection could be refined to plant properties.
Plant physical characteristics, such as plant size and plant physical structure, are known to contribute to the insect acceptance of host plant [72]. The plant size may influence the development of infestation by increasing population density and selection pressure [73]. As CMBS primarily settle and feed on of stem or trunk of plants, a larger size plant would have bigger surface area to host a higher level of infestation, without creating extra ‘pressure’ for the CMBS population. ‘Natchez’ is a large crapemyrtle tree that grows up to 30 feet in height [74], while ‘Spiced Plum’ is expected to mature at a smaller size [75]. However, this could not explain the different infestation levels exhibited among plants of similar size. For example, ‘Spiced Plum’ (grown in 3.79 L container) had a higher level of infestation compared to a similar size crapemyrtle, ‘Dynamite’ (grown in 3.79 L container), from a different feeding experiment [71].
Crapemyrtle bark scale host selection may also involve the development of insects on hosts with a certain plant structure. According our observation and previous reports, CMBS tend to feed on the cracks and crevice, or the wounded area of plant [2]. This suggests that CMBS might find it harder to accept certain plants with a smooth stem surface structure as a host, such as several Rubus species included in this study.
Another major factor that affects CMBS host selection and the level of infestation may be plant biochemical properties, such as the composition of saccharides and amino acids [76], which leads to the variation of population dynamics, such as altered development duration and sex ratio. The altered sex ratio of insect populations was found when the insect feeds on different hosts [77,78], or under environmental factors such as temperature [79]. The sex ratio of CMBS on ‘Spiced Plum’ was 3.67:1 (male:female) in this study (Table 4), whereas the sex ratio on ‘Natchez’ remains unknown from previous studies.
The larger number of male pupae and gravid females on Lagerstroemia indicates that CMBS showed less feeding preference on other tested plants. Compared to Lagerstroemia, there is lower risk in terms of economic damage for B. harlandii, B. microphylla var. koreana × B. sempervirens, C. speciosa, D. rhombifolia, D. virginiana, M. domestica ‘Fuji’, M. domestica ‘Red Delicious’, M. angustifolia, R. ‘Arapaho’, R. fruticosus, R. idaeus ‘Dorman Red’, and R. ‘Navaho’, as well as pomegranates, when being exposed to CMBS, since no infestation was recorded, or the recorded infestation might not reach the threshold for aesthetic and economic damages.
Economic thresholds (ET) of plant damages incurred by pest insects are widely used as advisory tools to determine action points for effective treatments [80,81,82]. One study showed that soybean aphid has an ET of 250 insects per plant [82], however, the ET could be lower for CMBS infesting ornamental crops, considering the negative effect on the aesthetic value resulting from defoliation, loss of flowering, and the accumulation of sooty mold [2,4,7].
The ET of CMBS has not yet been well categorized, and could be subject to change according to the specific crops. For example, ornamental crops, such as impatiens [83], tend to have lower tolerance compared to other economically important food crops, such as strawberry [84], when exposed to similar type of pest damage. Therefore, crapemyrtles and C. speciosa present in this study might have a low ET, since the presence of small numbers of CMBS could greatly decrease the ornamental values [2].
The infestation of CMBS had showed the level of damage well above ET for the control plant (L. ‘Spiced Plum’) in terms of aesthetic value (Figure S1). The infestations found on all alternative hosts in this study should be considered under the ET, due to the low number of insects presented (Figure S1). However, the source of CMBS inoculant in this experimental setting was relatively low, as only 10 CMBS ovisacs were used to inoculate each test plant. Under a natural setting, it is possible that plants infested with CMBS would produce large, and continuous influxes of insects to the nearby healthy plants. Therefore, the effect of heavier CMBS inoculation might lead to different levels of infestations on alternative plant hosts, which could be further investigated in future studies.
5. Conclusions
Crapemyrtle bark scale feeding preference studies on eight genera reported infestation on apple (M. domestica), C. speciosa, D. rhombifolia, H. salicifolia, L. ‘Spiced Plum’, M. angustifolia, and pomegranate cultivars including ‘Double Pink’, ‘Gissarskii Rozovyi’, ‘Kandahar’, ‘Kara Bala Miursal’, ‘Kazake’, ‘Mollar’, ‘Pecos’, ‘Angel Red’, ‘San Antonio’, ‘Sogidavna’, ‘Sumbar’, and ‘Surh-Anor’. No sign of CMBS infestation was observed on R. ‘Arapaho’, R. ‘Navaho’, R. idaeus ‘Dorman Red’, R. fruticosus, B. microphylla var. koreana × B. sempervirens, B. harlandii, and D. virginiana. No sign of CMBS infestation was observed on pomegranate cultivars including ‘Al-Sirin-Nar’, ‘Apseronski Krasnyi’, ‘Austin’, ‘Azadi’, ‘Bala Miursal’, ‘Christina’, ‘Desertnyi’, ‘Elf’, ‘Entek Habi Saveh’, ‘Girkanets’, ‘Gissarskii Rozovyi’, ‘JD’, ‘Kandahar’, ‘Kara Kalinski’, ‘Kazake’, ‘Larkin’, ‘Molla Nepes’, ‘Mollar’, ‘Mridula’, ‘Russian 18’, ‘Salavatsk’, ‘Sirenevyi’, ‘Sogidavna’, ‘Spanish Sweet’, ‘Sweet’, ‘Vkusanyi’, and ‘Wonderful’ in 2016 and 2019.
This study expands the current host range for CMBS and confirmed the abilities of CMBS to infest alternative plant species, especially plants within genera of Chaenomeles, Heimia, Malus, and Punica, with different feeding preferences. The damage caused by CMBS on alternative plant hosts in this study is relatively low compared to Lagerstroemia, however, the effect and potential threats of CMBS to its current list of alternative hosts, as well as other plant species, should be evaluated under more experimental conditions.
Supplementary Materials
The following are available online at https://www.mdpi.com/2075-4450/11/7/399/s1, Figure S1: Infestation of Acanthococcus lagerstromiae (red arrows) found on Punica ‘Angel Red’ (A), P. ‘Sumbar’ (B), P. ‘Mollar’ (C), P. ‘Kandahar’ (D), Malus domestica ‘Fuji’ (E-F), M. domestica ‘Red Delicious’ (G), M. angustifolia (H), Chaenomeles speciosa (I-K), Heimia salicifolia (L-M), and Lagerstroemia ‘Spiced Plum’ (N-O) recorded from feeding preference studies in 2016 and 2019. Table S1: The number of male pupae on fourteen plant species and cultivars by time from June to December 2019, Table S2: The number of gravid females on fourteen plant species and cultivars by time from June to December 2019.
Author Contributions
Conceptualization, M.G. and H.Q.; methodology, M.G.; software, R.X.; validation, R.X., H.Q. and M.G.; formal analysis, R.X.; investigation, R.X.; resources, G.W.K. and M.G.; data curation, R.X., B.W., H.D. and C.L.; writing—original draft preparation, R.X.; writing—review and editing, G.W.K., H.Q. and M.G.; visualization, B.W. and H.Q.; supervision, M.G. and H.Q.; project administration, M.G.; funding acquisition, M.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work is partially supported by Crop Protection and Pest Management project ‘Integrated pest management strategies for crape myrtle bark scale, a new exotic pest’ [grant no. 2014-70006-22632/project accession no. 10004888] and Specialty Crop Research Initiative project ‘Systematic Strategies to Manage Crapemyrtle Bark Scale, An Emerging Exotic Pest’ [grant no. 2017-51181-26831/project accession no. 1013059] from the U.S. Department of Agriculture (USDA) National Institute of Food and Agriculture. Any opinions, findings, conclusions, or recommendations expressed in this publication are those of the author(s) and do not necessarily reflect the view of the USDA. Additional financial support was from AmericanHort Horticultural Research Institute and Center for Applied Nursery Research.
Acknowledgments
The authors would like to acknowledge technical support from Jingru Lai and Qiansheng Li, and Mack Thetford from University of Florida and Larry Stein from Texas A&M AgriLife Extension Service for providing pomegranate propagation materials. We would like to thank the College of the Agriculture Women Excited to Share Opinions, Mentoring and Experiences (AWESOME) faculty group of the College of Agriculture and Life Sciences at Texas A&M University for assistance with editing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Egolf, D.R.; Andrick, A.O. The Lagerstroemia Handbook-Checklist: A Guide to Crapemyrtle Cultivars; American Association of Botanical Gardens and Arboreta, Inc.: Chester County, PA, USA, 1978; Volume 2, p. 7. [Google Scholar]
- Gu, M.; Merchant, M.; Robbins, J.; Hopkins, J. Crape Myrtle Bark Scale: A New Exotic Pest. Texas A M AgriLife Ext. 2014. Available online: https://agrilifecdn.tamu.edu/citybugstest/files/2010/05/EHT-049-Crape-myrtle-bark-scale.pdf (accessed on 18 October 2018).
- EDDMapS. Early Detection and Distribution Mapping System. The University of Georgia-Center for Invasive Species and Ecosystem Health. Available online: http://www.eddmaps.org/cmbs/distribution.cfm?map=distribution (accessed on 8 August 2019).
- Wang, Z.; Chen, Y.; Gu, M.; Vafaie, E.; Merchant, M.; Diaz, R. Crapemyrtle Bark Scale: A New Threat for Crapemyrtles, a Popular Landscape Plant in the U.S. Insects 2016, 7, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- USDA. Census of Horticultural Specialties (2014); USDA: Washington, DC, USA, 2014.
- Knox, G.W. Crape Myrtle in Florida; University of Florida Cooperative Extension Service: Gainesville, FL, USA, 2000. [Google Scholar]
- Jiang, N.; Xu, H. Observertion on Eriococcus lagerostroemiae Kuwana. J. Anhui Agric. Coll. 1998, 25, 142–144. [Google Scholar]
- He, D.; Cheng, J.; Zhao, H.; Chen, S. Biological characteristic and control efficacy of Eriococcus lagerstroemiae. Chin. Bull. Entomol. 2008, 45, 812–814. [Google Scholar]
- Wang, Z.; Chen, Y.; Knox, G.; Ring, D.; Diaz, R. Crape Myrtle Bark Scale. Available online: http://www.lsuagcenter.com/~/media/system/7/8/d/1/78d165df43ac0d4767607d88dadfb841/pub3440bugbizcrapemyrtlebarkscale_final.pdf (accessed on 16 May 2016).
- Luo, Q.; Xie, X.; Zhou, L.; Wang, S.; Xu, Z. A study on the dynamics and biological characteristics of Eriococcus lagerstroemiae Kuwanae population in Guiyang. Acta Entomol. Sin. 2000, 43, 35–42. [Google Scholar]
- Wang, Z.; Chen, Y.; Diaz, R.; Laine, R.A. Physiology of crapemyrtle bark scale, Acanthococcus lagerstroemiae (Kuwana), associated with seasonally altered cold tolerance. J. Insect Physiol. 2019, 112, 1–8. [Google Scholar] [CrossRef]
- Robbins, J.A.; Hopkins, J.; Merchant, M.; Gu, M. Crapemyrtle Bark Scale: A New Insect Pest; Cooperative Extension Service, University of Arkansas: Little Rock, AR, USA, 2014; Available online: https://www.uaex.edu/publications/PDF/fsa-7086.pdf (accessed on 19 September 2015).
- Chappell, M.R.; Braman, S.K.; Williams-Woodward, J.; Knox, G.J. Optimizing plant health and pest management of Lagerstroemia spp. in commercial production and landscape situations in the southeastern United States: A review. HortScience. 2012, 30, 161–172. [Google Scholar]
- Wang, Z.; Chen, Y.; Diaz, R. Thermal Tolerance and Prediction of Northern Distribution of the Crapemyrtle Bark Scale (Hemiptera: Eriococcidae). Environ. Entomol. 2019, 48, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.-J. Notes on some parasitoids (Hymenoptera: Chalcidoidea) associated with Acanthococcus lagerstroemiae (Kuwana)(Hemiptera: Eriococcidae) in the Republic of Korea. Insecta Mundi 2019, 0690, 1–5. [Google Scholar]
- Liu, S.S.; Meng, X.D. Modelling development time of Myzus persicae (Hemiptera: Aphididae) at constant and natural temperatures. Bull. Entomol. Res. 1999, 89, 53–63. [Google Scholar] [CrossRef]
- Pandey, A.K.; Tripathi, C.J.B. Effect of temperature on the development, fecundity, progeny sex ratio and life-table of Campoletis chlorideae, an endolarval parasitoid of the pod borer, Helicoverpa armigera. BioControl 2008, 53, 461. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Y.; Diaz, R. Temperature-dependent development and host range of crapemyrtle bark scale, Acanthococcus lagerstroemiae (Kuwana)(Hemiptera: Eriococcidae). Fla. Entomol. 2019, 102, 181–186. [Google Scholar]
- Brodbeck, B.V.; Andersen, P.C.; Oden, S.; Mizell, R.F.; McKamey, S.H.; Zapata, M. The distribution of Cicadellinae leafhoppers and other Auchenorrhyncha on coffee and citrus in Puerto Rico. Environ. Entomol. 2017, 46, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Yu, X.; Wu, G.; Tao, L.; Chen, J.; Zheng, X. The virulence change and damage characteristics of various geographic populations of brown planthopper. Insect Sci. 1999, 6, 146–154. [Google Scholar] [CrossRef]
- Truzi, C.C.; Vieira, N.F.; de Laurentis, V.L.; Vacari, A.M.; De Bortoli, S.A. Development and feeding behavior of Helicoverpa armigera (Hübner)(Lepidoptera: Noctuidae) on different sunflower genotypes under laboratory conditions. Antrhopod Plant Inetract. 2017, 11, 797–805. [Google Scholar] [CrossRef] [Green Version]
- Prasad, A.K.; Roy, S. Role of host switching in the development of pesticide tolerance in Helopeltis theivora (Hemiptera: Miridae), the major pest of tea in India. In Annales de la Société Entomologique de France (NS); Taylor & Francis: Milton Park, UK, 2017; Volume 53, pp. 428–433. [Google Scholar]
- Morrison, C.R.; Aubert, C.; Windsor, D.M. Variation in Host Plant Usage and Diet Breadth Predict Sibling Preference and Performance in the Neotropical Tortoise Beetle Chelymorpha alternans (Coleoptera: Chrysomelidae: Cassidinae). Environ. Èntomol. 2019, 48, 382–394. [Google Scholar] [CrossRef]
- Castiglione, E.; Manti, F.; Bonsignore, C.P. First record from Calabria (southern Italy) of the “bronze bug” Thaumastocoris peregrinus Carpintero & Dellapé, 2006, alien Eucalyptus pest native to Australia (Hemiptera: Heteroptera: Thaumastocoridae). J. Entomol. Acarol. Res. 2020, 52. [Google Scholar] [CrossRef]
- Nibouche, S.; Mississipi, S.; Fartek, B.; Delatte, H.; Reynaud, B.; Costet, L. Host plant specialization in the sugarcane aphid Melanaphis sacchari. PLoS ONE 2015, 10, e0143704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troncoso, A.J.; Vargas, R.; Tapia, D.; Olivares-Donoso, R.; Niemeyer, H.M. Host selection by the generalist aphid Myzus persicae (Hemiptera: Aphididae) and its subspecies specialized on tobacco, after being reared on the same host. Bull. Entomol. Res. 2005, 95, 23–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaud, J.P. Assessment of cotton as an alternative host plant for the brown citrus aphid, Toxoptera citricida (Homoptera: Aphididae). Fla. Entomol. 2004, 87, 105–112. [Google Scholar] [CrossRef]
- Tapia, D.H.; Silva, A.X.; Ballesteros, G.I.; Figueroa, C.C.; Niemeyer, H.M.; Ramírez, C.C. Differences in learning and memory of host plant features between specialist and generalist phytophagous insects. Anim. Behav. 2015, 106, 1–10. [Google Scholar] [CrossRef]
- Wilkinson, T.L.; Douglas, A.E. Phloem amino acids and the host plant range of the polyphagous aphid, Aphis fabae. Entomol. Exp. Appl. 2003, 106, 103–113. [Google Scholar] [CrossRef]
- Adachi, S.; Matsumoto, Y.; Yoshitomi, H.; Sasaki, D.; Tokuda, M.J. New Distribution Records of Aphis nasturtii (Hemiptera: Aphididae) in Japan. Jpn. J. Syst. Entomol. 2017, 23, 191–193. [Google Scholar]
- Guidolin, A.S. Multipartite Interactions of Aphis (Toxoptera) and Their Associated Symbionts. Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2016. [Google Scholar] [CrossRef]
- Miller, G.L.; Favret, C.; Carmichael, A.; Voegtlin, D.J. Is there a cryptic species within Aulacorthum solani (Hemiptera: Aphididae)? J. Econ. Entomol. 2009, 102, 398–400. [Google Scholar] [CrossRef] [PubMed]
- Madjdzadeh, S.M.; Mehrparvar, M.; Abolhasanzadeh, F.J.R. Morphometric discrimination of host-adapted populations of Brachycaudus helichrysi (Kaltenbach)(Hemiptera Aphididae). Redia 2009, 92, 143–145. [Google Scholar]
- Raboudi, F.; Chavigny, P.; Marrakchi, M.; Makni, H.; Makni, M.; Vanlerberghe-Masutti, F. Characterization of polymorphic microsatellite loci in the aphid species Macrosiphum euphorbiae (Hemiptera: Aphididae). Mol. Ecol. Notes 2005, 5, 490–492. [Google Scholar] [CrossRef]
- Sylvester, E.S.; Richardson, J. Consecutive serial passage of strawberry crinkle virus in Myzus ornatus by injection and its occasional transmission to Fragaria vesca. Phytopathology 1986, 76, 1161–1164. [Google Scholar] [CrossRef]
- Francis, F.; Haubruge, E.; Gaspar, C. Influence of host plants on specialist/generalist aphids and on the development of Adalia bipunctata (Coleoptera: Coccinellidae). Eur. J. Entomol. 2000, 97, 481–486. [Google Scholar] [CrossRef] [Green Version]
- Moran, N.A. A 48-million-year-old aphid—Host plant association and complex life cycle: Biogeographic evidence. Science 1989, 245, 173–175. [Google Scholar] [CrossRef]
- Ocimati, W.; Were, E.; Groot, J.C.; Tittonell, P.; Nakato, G.V.; Blomme, G. Risks posed by intercrops and weeds as alternative hosts to Xanthomonas campestris pv. musacearum in banana fields. Front. Plant Sci. 2018, 9, 1471. [Google Scholar] [CrossRef]
- Bawin, T.; Dujeu, D.; De Backer, L.; Francis, F.; Verheggen, F.J. Ability of Tuta absoluta (Lepidoptera: Gelechiidae) to develop on alternative host plant species. Can. Entomol. 2016, 148, 434–442. [Google Scholar] [CrossRef]
- Unruh, T.R.; Luck, R.F. Deme formation in scale insects: A test with the pinyon needle scale and a review of other evidence. Ecol. Entomol. 1987, 12, 439–449. [Google Scholar] [CrossRef]
- Spitzer, B. Local maladaptation in the soft scale insect Saissetia coffeae (Hemiptera: Coccidae). Evolution 2006, 60, 1859–1867. [Google Scholar] [CrossRef] [PubMed]
- Schultz, P.B.; Szalanski, A.L. Hypericum kalmianum (St. Johnswort) Confirmed as a New Host of the Crapemyrtle Bark Scale in Virginia, USA. J. Agric. Urban Entomol. 2019, 35, 12–15. [Google Scholar] [CrossRef]
- Gregory, E.; USDA Systematic Entomology Laboratory, Beltsville, MD, USA; Gu, M.; Department of Horticultural Sciences, Texas A&M AgriLife Extension Service, College Station, TX, USA. Personal Communication, 2020.
- Ma, J.-H. Occurrence and biological characteristics of Eriococcus lagerostroemiae Kuwana in Panxi district. Biology 2011, 5, 3. [Google Scholar]
- Guo, D.L.; Luo, Z.R. Genetic relationships of the Japanese persimmon Diospyros kaki (Ebenaceae) and related species revealed by SSR analysis. Genet. Mol. Res. 2011, 10, 1060–1068. [Google Scholar] [CrossRef]
- Rauf, A.; Uddin, G.; Patel, S.; Khan, A.; Halim, S.A.; Bawazeer, S.; Ahmad, K.; Muhammad, N.; Mubarak, M.S. Diospyros, an under-utilized, multi-purpose plant genus: A review. Biomed. Pharm. 2017, 91, 714–730. [Google Scholar] [CrossRef]
- Celik, A.; Ercisli, S. Persimmon cv. Hachiya (Diospyros kaki Thunb.) fruit: Some physical, chemical and nutritional properties. Int. J. Food Sci. Nutr. 2008, 59, 599–606. [Google Scholar] [CrossRef]
- United States Department of Agriculture. Noncitrus Fruits and Nuts 2018 Summary; US Department of Agriculture, National Agricultural Statistics Service: Washington, DC, USA, 2019.
- Brown, S. Apple. In Fruit Breeding; Springer: Berlin/Heidelberg, Germany, 2012; pp. 329–367. [Google Scholar]
- United States Department of Agriculture. Census of Agriculture; US Department of Agriculture, National Agricultural Statistics Service: Washington, DC, USA, 2017; Volume 1.
- Holland, D.; Hatib, K.; Bar-Ya’akov, I. Pomegranate: Botany, Horticulture, Breeding. Hortic. Rev. 2009, 35, 127–191. [Google Scholar]
- Iskenderova, Z.D. Double forms of pomegranate and their uses in ornamental horticulture. Byulleten’Glavnogo Bot. Sada 1980, 115, 58–62. [Google Scholar]
- Jalikop, S.H. Pomegranate breeding. Fruit Veg. Cereal Sci. Biothec 2010, 4, 26–34. [Google Scholar]
- USDA. USA National Plant Germplasm System. Available online: https://npgsweb.ars-grin.gov/gringlobal/site.aspx?id=4 (accessed on 23 April 2020).
- Bonsignore, C.P. Apate monachus (Fabricius, 1775), a bostrichid pest of pomegranate and carob trees in nurseries-Short Communication. Plant Prot. Sci. 2012, 48, 94–97. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Dan, J.; Tang, L.; Cao, S. Fruit scientific research in New China in the past 70 years: Pomegranate. J. Fruit Sci. 2019, 36, 1389–1398. [Google Scholar]
- Deng, Q. Control Eriococcus Lagerstroemiae Kuwana for Pomegranate Bonsai. China Flower Penjing 2001, 3, 33. [Google Scholar]
- Stover, E.; Mercure, E.W. The pomegranate: A new look at the fruit of paradise. HortScience 2007, 42, 1088–1092. [Google Scholar] [CrossRef] [Green Version]
- Park, J.D.; Kim, Y.H.; Kim, S.S.; Park, I.S.; Kim, K.C. Seasonal occurrence, host preference and hatching behavior of Eriococcus lagerstroemiae. Korean J. Appl. Entomol. 1993, 32, 83–89. [Google Scholar]
- Hua, L.-Z. List of Chinese Insects; Zhongshan University Press: Guangzhou, China, 2000; Volume I, p. 448. [Google Scholar]
- Kozár, F. Catalogue of Palaearctic Coccoidea; Plant Protection Institute: Budapest, Hungary, 1998. [Google Scholar]
- Castle, B. Cultivar Selection Guide for Florida-Grown Pomegranates: Horticultural Traits. Available online: https://crec.ifas.ufl.edu/extension/pomegranates/pom_guide.html (accessed on 21 May 2020).
- Hanks, L.M.; Millar, J.G.; Paine, T.D. Biological constraints on host-range expansion by the wood-boring beetle Phoracantha semipunctata (Coleoptera: Cerambycidae). Ann. Entomol. Soc. Am. 1995, 88, 183–188. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, Y.; Zhou, D.-S.; Gao, S.-X.; Zhao, X.-C.; Tang, Q.-B.; Wang, C.-Z.; van Loon, J.J.A. Higher plasticity in feeding preference of a generalist than a specialist: Experiments with two closely related Helicoverpa species. Sci. Rep. 2017, 7, 17876. [Google Scholar] [CrossRef]
- Aidoo, O.F.; Tanga, C.M.; Khamis, F.M.; Rasowo, B.A.; Mohamed, S.A.; Badii, B.K.; Salifu, D.; Sétamou, M.; Ekesi, S.; Borgemeister, C. Host suitability and feeding preference of the African citrus triozid Trioza erytreae Del Guercio (Hemiptera: Triozidae), natural vector of “Candidatus Liberibacter africanus”. J. Appl. Entomol. 2019, 143, 262–270. [Google Scholar] [CrossRef]
- Karowe, D.N. Facultative monophagy as a consequence of prior feeding experience: Behavioral and physiological specialization in Colias philodice larvae. Oecologia 1989, 78, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-J.; Zhang, S.-S.; Niu, B.-L.; Ji, D.-F.; Liu, X.-J.; Li, M.-W.; Bai, H.; Palli, S.R.; Wang, C.-Z.; Tan, A. A determining factor for insect feeding preference in the silkworm, Bombyx mori. PLoS Biol. 2019, 17, e3000162. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, S.; Blanchet, E.; Egas, M.; Olivieri, I.J.B.E.B. Are adaptation costs necessary to build up a local adaptation pattern? BMC Evol. Biol. 2009, 9, 182. [Google Scholar]
- Kellerhals, M. Introduction to apple (Malus × domestica). In Genetics and Genomics of Rosaceae; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2009; pp. 73–84. [Google Scholar]
- Hokanson, S.; Lamboy, W.; Szewc-McFadden, A.; McFerson, J. Microsatellite (SSR) variation in a collection of Malus (apple) species and hybrids. Euphytica 2001, 118, 281–294. [Google Scholar] [CrossRef]
- Wu, B.; Xie, R.; Knox, G.W.; Qin, H.; Gu, M. Crapemyrtle bark scale (Hemiptera: Eriococcidae) showed different acceptance among crapemyrtle species significantly. 2020; unpublished; manuscript in preparation. [Google Scholar]
- Denno, R.; Roderick, G. Influence of patch size, vegetation texture, and host plant architecture on the diversity, abundance, and life history styles of sapfeeding herbivores. In Habitat Structure; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1991; pp. 169–196. [Google Scholar]
- Thompson, J.N. Selection pressures on phytophagous insects feeding on small host plants. Oikos 1983, 438–444. [Google Scholar] [CrossRef]
- Pecot, H.C. Influence of Planting Depth and Mulch on the Growth of Nine Species of Ornamental Plants in Landscape and Container Settings. Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 2004. [Google Scholar]
- Hansen, H.A. Lagerstroemia Plant Named ‘Spiced Plum’. U.S. Patent 15/530,535, 16 July 2018. [Google Scholar]
- Thorsteinson, A.J. Host selection in phytophagous insects. Annu. Rev. Entomol. 1960, 5, 193–218. [Google Scholar] [CrossRef]
- Kumar, H.; Bhattacharya, S. Biology of Spodoptera litura (Fabricius) on different crop plants. J. Entomol. Res. 2019, 43, 165–168. [Google Scholar] [CrossRef]
- Mani, C.; Lawrence, L.; Ranjith, M.J.E. Biology and morphometry of Paracoccus marginatus Williams and Granara de Willink (Hemiptera: Pseudococcidae). Entomon 2013, 38, 97–110. [Google Scholar]
- Raja, S.; Gillani, W.; Copland, M.J.B.S.-P. Effect of Different Temperatures and Host Plants on the Biology of the Long-Tailed Mealy Bug Pseudococcus longispinus (Targioni and Tozzetti)(Homoptera: Pseudococcidae). Pak. J. Sci. Ind. Res. Ser. B Biol. Sci. 2011, 54, 142–151. [Google Scholar]
- Chaudhari, V.; Desai, H.; Patel, N. Determination of Economic Threshold Level of Cotton Leafhopper (Amarasca biguttula biguttula Ishida) on Cotton under South Gujarat Condition. Trends Biosci. 2018, 11, 52–57. [Google Scholar]
- Soliman, A.; Hendawy, A.; El-hefny, A.S.; Sherif, M. Determination of Economic Threshold Level for Chilo agamemnon Bles. Infestation In Rice Plants, Based on Simulated Dead Hearts. Egypt. J. Agric. Res. 2016, 94. [Google Scholar]
- McCarville, M.T.; Kanobe, C.; Macintosh, G.C.; O’Neal, M. What is the economic threshold of soybean aphids (Hemiptera: Aphididae) in enemy-free space? J. Econ. Entomol. 2011, 104, 845–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alatawi, F.; Margolies, D.; Nechols, J. Aesthetic damage thresholds for twospotted spider mites (Acari: Tetranychidae) on impatiens: Effect of plant age and level of infestation. J. Econ. Entomol. 2007, 100, 1904–1909. [Google Scholar] [CrossRef] [PubMed]
- Short, B.D.; Janisiewicz, W.; Takeda, F.; Leskey, T.C. UV-C irradiation as a management tool for Tetranychus urticae on strawberries. Pest. Manag. Sci. 2018, 74, 2419–2423. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Number of male pupae and gravid females of Acanthococcus lagerstromiae per cultivar of pomegranates, including ‘Big Red’, ‘Double Pink’, ‘Gissarskii Rozovyi’, ‘Kandahar’, ‘Kara Bala Miursal’, ‘Kara Kalinski’, ‘Kazake’, ‘Mollar’, ‘Pecos’, ‘Russian 18’, ‘San Antonio’, ‘Sogidavna’, ‘Sumbar’, and ‘Surh-Anor’, recorded on August 10th 2019. No male pupa or gravid female was observed on ‘Big Red’, ‘Kara Kalinski’, or ‘Russian 18’.
Figure 1.
Number of male pupae and gravid females of Acanthococcus lagerstromiae per cultivar of pomegranates, including ‘Big Red’, ‘Double Pink’, ‘Gissarskii Rozovyi’, ‘Kandahar’, ‘Kara Bala Miursal’, ‘Kara Kalinski’, ‘Kazake’, ‘Mollar’, ‘Pecos’, ‘Russian 18’, ‘San Antonio’, ‘Sogidavna’, ‘Sumbar’, and ‘Surh-Anor’, recorded on August 10th 2019. No male pupa or gravid female was observed on ‘Big Red’, ‘Kara Kalinski’, or ‘Russian 18’.

Figure 2.
Number of male pupae of Acanthococcus lagerstromiae on Malus domestica ‘Fuji’, M. domestica ‘Red Delicious’, M. angustifolia, Chaenomeles speciosa, Diospyros rhombifolia, Heimia salicifolia, and Lagerstroemia ‘Spiced Plum’, recorded from June to December 2019.
Figure 2.
Number of male pupae of Acanthococcus lagerstromiae on Malus domestica ‘Fuji’, M. domestica ‘Red Delicious’, M. angustifolia, Chaenomeles speciosa, Diospyros rhombifolia, Heimia salicifolia, and Lagerstroemia ‘Spiced Plum’, recorded from June to December 2019.

Figure 3.
Number of gravid females of Acanthococcus lagerstromiae on Malus domestica ‘Fuji’, M. domestica ‘Red Delicious’, Chaenomeles speciosa, Heimia salicifolia, and Lagerstroemia ‘Spiced Plum’, recorded from June to December 2019.
Figure 3.
Number of gravid females of Acanthococcus lagerstromiae on Malus domestica ‘Fuji’, M. domestica ‘Red Delicious’, Chaenomeles speciosa, Heimia salicifolia, and Lagerstroemia ‘Spiced Plum’, recorded from June to December 2019.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Pomegranate (Punica granatum) cultivars as host candidates of Acanthococcus lagerstroemiae used in feeding trials in 2016 and 2019, respectively.
Table 1.
Pomegranate (Punica granatum) cultivars as host candidates of Acanthococcus lagerstroemiae used in feeding trials in 2016 and 2019, respectively.
| 2016 | 2019 | |
|---|---|---|
| ‘Al-Sirin-Nar’ | ‘Kazake’ | ‘Big Red’ |
| ‘Angel Red’ | ‘Larkin’ | ‘Double Pink’ |
| ‘Apseronski Krasnyi’ | ‘Molla Nepes’ | ‘Gissarskii Rozovyi’ |
| ‘Austin’ | ‘Mollar’ | ‘Kandahar’ |
| ‘Azadi’ | ‘Mridula’ | ‘Kara Bala Miursal’ |
| ‘Bala Miursal’ | ‘Russian 18’ | ‘Kara Kalinski’ |
| ‘Christina’ | ‘Salavatski’ | ‘Kazake’ |
| ‘Desertnyi’ | ‘Sirenevyi’ | ‘Mollar’ |
| ‘Elf’ | ‘Sogidavna’ | ‘Pecos’ |
| ‘Entek Habi Saveh’ | ‘Spanish Sweet’ | ‘Russian 18’ |
| ‘Girkanets’ | ‘Sumbar’ | ‘San Antonio’ |
| ‘Gissarskii Rozovyi’ | ‘Surh-Anor’ | ‘Sogidavna’ |
| ‘JD’ | ‘Sweet’ | ‘Sumbar’ |
| ‘Kandahar’ | ‘Vkusanyi’ | ‘Surh-Anor’ |
| ‘Kara Kalinski’ | ‘Wonderful’ | |
Table 2.
Plant species as host candidates of Acanthococcus lagerstroemiae used in feeding preference study in 2019.
Table 2.
Plant species as host candidates of Acanthococcus lagerstroemiae used in feeding preference study in 2019.
| Scientific Name | Common Name | Family | USDA Cold Hardiness Zone |
|---|---|---|---|
| Buxus harlandii | Harland boxwood | Buxaceae | 7–9 |
| Buxus microphylla var. koreana × B. sempervirens ‘Green Gem’ | Boxwood | Buxaceae | 7–9 |
| Chaenomeles speciosa ‘Texas Scarlet’ | Common quince | Rosaceae | 4–8 |
| Diospyrosrhombifolia | Diamond-leaf Persimmon | Ebenaceae | 7–11 |
| Diospyros virginiana | Common persimmon | Ebenaceae | 4–9 |
| Heimia salicifolia | Sinicuichi | Lythraceae | 9–11 |
| Lagerstroemia ‘Spiced Plum’ | Crapemyrtle | Lythraceae | 6–10 |
| Malus angustifolia | Southern crabapple | Rosaceae | 3–8 |
| Malus domestica ‘Fuji’ | Apple | Rosaceae | 3–8 |
| Malus domestica ‘Red Delicious’ | Apple | Rosaceae | 3–8 |
| Rubus ‘Arapaho’ | Blackberry | Rosaceae | 4–9 |
| Rubus ‘Navaho’ | Raspberry | Rosaceae | 6–10 |
| Rubus fruticosus ‘Prime Ark Freedom’ | Blackberry | Rosaceae | 6–9 |
| Rubus idaeus ‘Dorman Red’ | Raspberry | Rosaceae | 5–9 |
Table 3.
The number of male pupae and gravid females, and sex ratio (male to female ratio) on five plant species and cultivars infested with Acanthococcus lagerstromiae in September 2019.
Table 3.
The number of male pupae and gravid females, and sex ratio (male to female ratio) on five plant species and cultivars infested with Acanthococcus lagerstromiae in September 2019.
| Plant Species | No. Male Pupae | No. Gravid Females | Sex Ratio |
|---|---|---|---|
| Lagerstroemia ‘Spiced Plum’ | 555.7a Z | 166.5a | 3.3:1 |
| Heimia salicifolia | 25.7b | 9.3b | 2.8:1 |
| Malus domestica ‘Red Delicious’ | 4.2b | 1.0b | 4.2:1 |
| M. domestica ‘Fuji’ | 3.7b | 3.7b | 1.0:1 |
| Chaenomeles speciosa | 3.2b | 2.8b | 1.1:1 |
Z Means within each column followed by the same letter are not significantly different according to All Pairs, Tukey Honestly Significant Difference at 0.05 confidence level.
Table 4.
The number of male pupae and gravid females, and sex ratio (male to female ratio) on seven plant species and cultivars infested with Acanthococcus lagerstromiae from June to December 2019.
Table 4.
The number of male pupae and gravid females, and sex ratio (male to female ratio) on seven plant species and cultivars infested with Acanthococcus lagerstromiae from June to December 2019.
| Plant Species | No. Male Pupae | No. Gravid Females | Sex Ratio |
|---|---|---|---|
| Lagerstroemia ‘Spiced Plum’ | 321.5a Z | 87.7a | 3.7:1 |
| Heimia salicifolia | 12.6b | 5.6b | 2.3:1 |
| Malus domestica ‘Red Delicious’ | 2.4b | 0.3b | 7.2:1 |
| M. domestica ‘Fuji’ | 1.9b | 1.9b | 1.0:1 |
| Chaenomeles speciosa | 1.1b | 1.4b | 0.8:1 |
| M. angustifolia | 0.2b | - | - |
| Diospyrosrhombifolia | 0.03b | - | - |
Z Means within each column followed by the same letter are not significantly different according to All Pairs, Tukey Honestly Significant Difference at 0.05 confidence level.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xie, R.; Wu, B.; Dou, H.; Liu, C.; Knox, G.W.; Qin, H.; Gu, M. Feeding Preference of Crapemyrtle Bark Scale (Acanthococcus lagerstroemiae) on Different Species. Insects 2020, 11, 399. https://doi.org/10.3390/insects11070399
AMA Style
Xie R, Wu B, Dou H, Liu C, Knox GW, Qin H, Gu M. Feeding Preference of Crapemyrtle Bark Scale (Acanthococcus lagerstroemiae) on Different Species. Insects. 2020; 11(7):399. https://doi.org/10.3390/insects11070399
Chicago/Turabian StyleXie, Runshi, Bin Wu, Haijie Dou, Cuiyu Liu, Gary W. Knox, Hongmin Qin, and Mengmeng Gu. 2020. "Feeding Preference of Crapemyrtle Bark Scale (Acanthococcus lagerstroemiae) on Different Species" Insects 11, no. 7: 399. https://doi.org/10.3390/insects11070399
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.