Overview of Folic Acid Supplementation Alone or in Combination with Vitamin B12 in Dairy Cattle during Periparturient Period

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
1.1. Immunity and Periparturient Period
1.2. Folic Acid and Vitamin B12 Functions
2. The Research Progress of Folic Acid and Vitamin B12 in Dairy Cattle
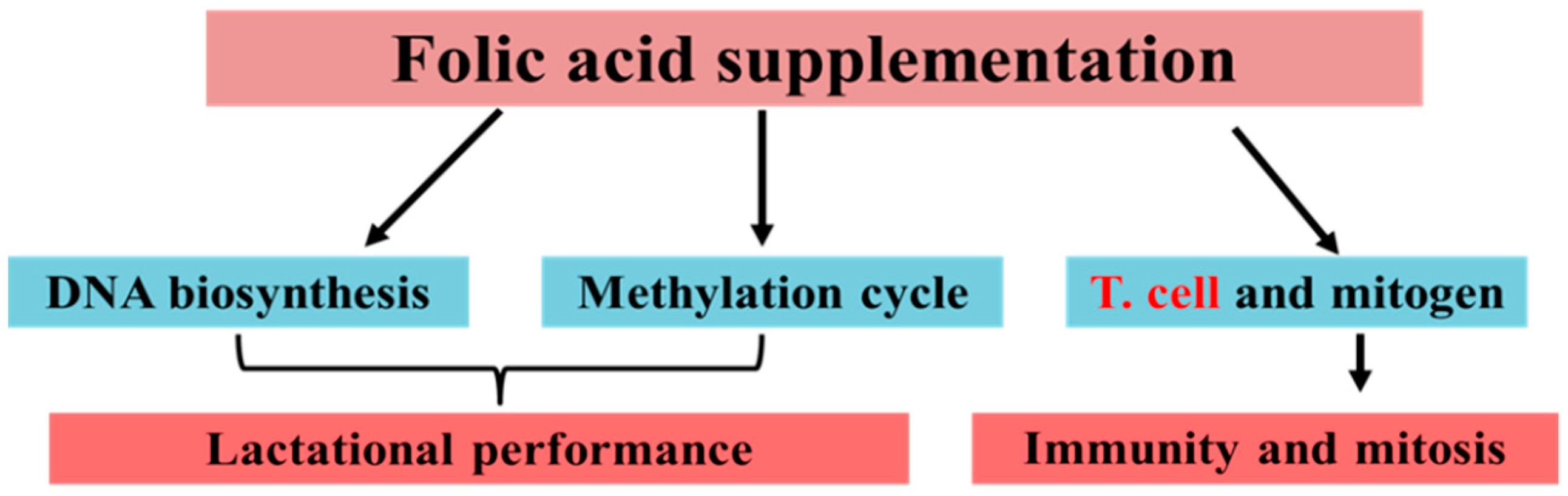
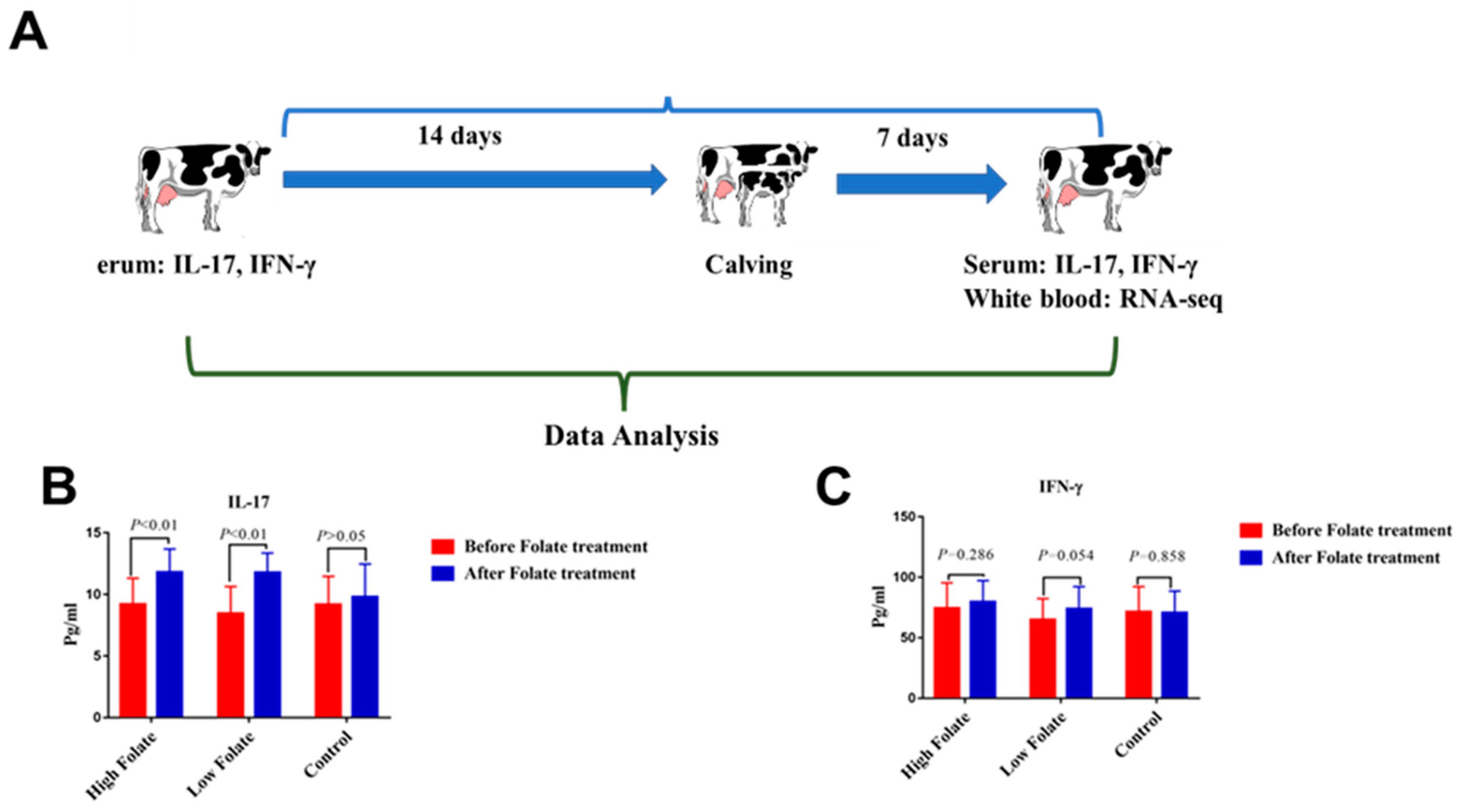
2.1. Folic Acid Significantly Regulates Immunity in Periparturient Dairy Cattle
2.2. Folic Acid Regulates Mastitis Resistance and Down-Regulates the Genes Involved in Mastitis Development
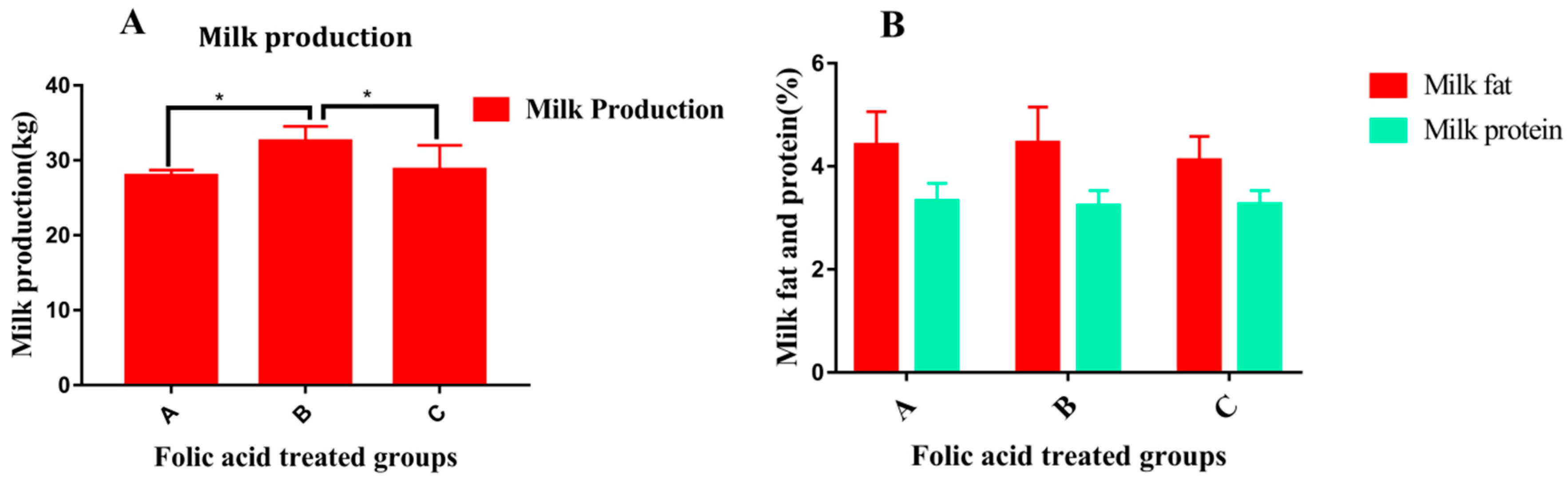
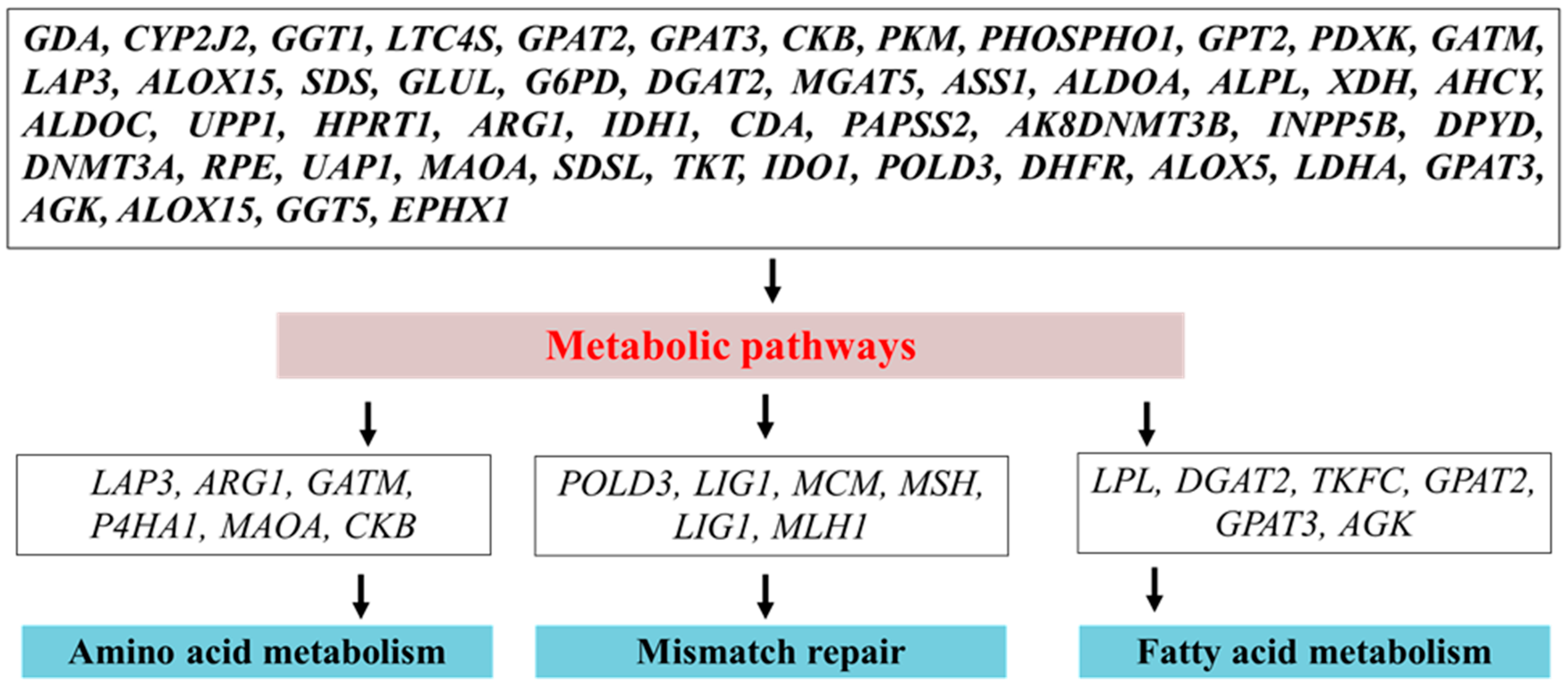
2.3. Folic Acid Alone or in Combination with Vitamin B12 Improves Milk Production and Metabolism in Periparturient Dairy Cattle
3. Future Direction for Research of Folic Acid and Vitamin B12 in Transition Dairy Cattle
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grummer, R.R.; Wiltbank, M.C.; Fricke, P.M.; Watters, R.D.; Silva-Del-Rio, N. Management of dry and transition cows to improve energy balance and reproduction. J. Reprod. Dev. 2010, 56, S22–S28. [Google Scholar] [CrossRef] [Green Version]
- Esposito, G.; Irons, P.C.; Webb, E.C.; Chapwanya, A. Interactions between negative energy balance, metabolic diseases, uterine health and immune response in transition dairy cows. Anim. Reprod. Sci. 2014, 144, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Lacasse, P.; Vanacker, N.; Ollier, S.; Ster, C. Innovative dairy cow management to improve resistance to metabolic and infectious diseases during the transition period. Res. Vet. Sci. 2018, 116, 40–46. [Google Scholar] [CrossRef]
- Bell, A.W. Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation. J. Anim. Sci. 1995, 73, 2804–2819. [Google Scholar] [CrossRef]
- Janovick, N.A.; Drackley, J.K. Prepartum dietary management of energy intake affects postpartum intake and lactation performance by primiparous and multiparous Holstein cows. J. Dairy Sci. 2010, 93, 3086–3102. [Google Scholar] [CrossRef]
- Roche, J.R.; Bell, A.W.; Overton, T.R.; Loor, J.J. Nutritional management of the transition cow in the 21st century-a paradigm shift in thinking. Anim. Prod. Sci. 2013, 53, 1000–1023. [Google Scholar] [CrossRef]
- Grummer, R.R. Impact of changes in organic nutrient metabolism on feeding the transition dairy cow. J. Anim. Sci. 1995, 73, 2820–2833. [Google Scholar] [CrossRef]
- Gonzalez, M.; Yabuta, A.K.; Galindo, F. Behaviour and adrenal activity of first parturition and multiparous cows under a competitive situation. Appl. Anim. Behav. Sci. 2003, 83, 259–266. [Google Scholar] [CrossRef]
- Meglia, G.E.; Johannisson, A.; Agenas, S.; Holtenius, K.; Waller, K.P. Effects of feeding intensity during the dry period on leukocyte and lymphocyte sub-populations, neutrophil function and health in periparturient dairy cows. Vet. J. 2005, 169, 376–384. [Google Scholar] [CrossRef]
- Kulberg, S.; Storset, A.K.; Heringstad, B.; Larsen, H.J.S. Reduced levels of total leukocytes and neutrophils in Norwegian cattle selected for decreased mastitis incidence. J. Dairy Sci. 2002, 85, 3470–3475. [Google Scholar] [CrossRef] [Green Version]
- Mulligan, F.J.; Doherty, M.L. Production diseases of the transition cow. Vet. J. 2008, 176, 3–9. [Google Scholar] [CrossRef]
- Mallard, B.A.; Dekkers, J.C.; Ireland, M.J.; Leslie, K.E.; Sharif, S.; Vankampen, C.L.; Wagter, L.; Wilkie, B.N. Alteration in Immune Responsiveness during the Peripartum Period and Its Ramification on Dairy Cow and Calf Health. J. Anim. Sci. 1998, 81, 585–595. [Google Scholar] [CrossRef]
- Dänicke, S.; Meyer, U.; Kersten, S.; Frahm, J. Animal models to study the impact of nutrition on the immune system of the transition cow. Res. Vet. Sci. 2018, 116, 15–27. [Google Scholar] [CrossRef]
- Trevisi, E.; Minuti, A. Assessment of the innate immune response in the periparturient cow. Res. Vet. Sci. 2018, 116, 47–54. [Google Scholar] [CrossRef]
- Jacob, S.K.; Ramnath, V.; Philomina, P.T.; Raghunandhanan, K.V.; Kannan, A. Assessment of physiological stress in periparturient cows and neonatal calves. Indian J. Physiol. Pharmacol. 2001, 45, 233–238. [Google Scholar]
- Lacetera, N.; Franci, O.; Scalia, D.; Bernabucci, U.; Ronchi, B.; Nardone, A. Effects of nonesterified fatty acids and β-hydroxybutyrate on functions of mononuclear cells obtained from ewes. Am. J. Vet. Res. 2002, 63, 414–418. [Google Scholar] [CrossRef]
- Bai, H.; Talukder, M.S.; Hiroki, K.; Tsukino, I.; Manabu, K.; Masashi, T. Evaluation of the immune status of peripheral blood monocytes from dairy cows during the periparturition period. J. Reprod. Dev. 2019, 65, 313–318. [Google Scholar] [CrossRef] [Green Version]
- Hammon, D.S.; Evjen, I.M.; Dhiman, T.R.; Goff, J.P.; Walters, J.L. Neutrophil function and energy status in Holstein cows with uterine health disorders. Vet. Immunol. Immunopathol. 2006, 113, 21–29. [Google Scholar] [CrossRef]
- Kehrli, M.E.; Nonnecke, B.J.; Roth, J.A. Alterations in bovine neutrophil function during the periparturient period. Am. J. Vet. Res. 1989, 50, 207–214. [Google Scholar]
- Sheldon, I.M.; Cronin, J.; Goetze, L.; Donofrio, G.; Schuberth, H.J. Defining postpartum uterine disease and the mechanisms of infection and immunity in the female reproductive tract in cattle. Biol. Reprod. 2009, 81, 1025–1032. [Google Scholar] [CrossRef]
- Crookenden, M.A.; Heiser, A.; Murray, A.; Dukkipati, V.S.R.; Kay, J.K.; Loor, J.J.; Meier, S.; Mitchell, M.D.; Moyes, K.M.; Walker, C.G.; et al. Parturition in dairy cows temporarily alters the expression of genes in circulating neutrophils. J. Dairy Sci. 2016, 99, 6470–6483. [Google Scholar] [CrossRef] [Green Version]
- Nonnecke, B.J.; Kimura, K.; Goff, J.P.; Kehrli, M.E. Effects of the mammary gland on functional capacities of blood mononuclear leukocyte populations from periparturient cows 1. J. Dairy Sci. 2003, 86, 2359–2368. [Google Scholar] [CrossRef]
- Itle, A.J.; Huzzey, J.M.; Weary, D.M.; Von Keyserlingk, M.A.G. Clinical ketosis and standing behavior in transition cows. J. Dairy Sci. 2015, 98, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Bertoni, G.; Trevisi, E.; Han, X.; Bionaz, M. Effects of inflammatory conditions on liver activity in puerperium period and consequences for performance in dairy cows. J. Dairy Sci. 2008, 91, 3300–3310. [Google Scholar] [CrossRef] [Green Version]
- McArt, J.A.A.; Nydam, D.V.; Oetzel, G.R. Epidemiology of subclinical ketosis in early lactation dairy cattle. J. Dairy Sci. 2012, 95, 5056–5066. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, E.S.; Lima, F.S.; Greco, L.F.; Bisinotto, R.S.; Monteiro, A.P.A.; Favoreto, M.; Ayres, H.; Marsola, R.S.; Martinez, N.; Thatcher, W.W.; et al. Prevalence of periparturient diseases and effects on fertility of seasonally calving grazing dairy cows supplemented with concentrates. J. Dairy Sci. 2013, 96, 5682–5697. [Google Scholar] [CrossRef]
- Choi, S.-W.; Mason, J.B. Folate and carcinogenesis: An integrated scheme. J. Nutr. 2000, 130, 129–132. [Google Scholar] [CrossRef] [Green Version]
- Ghoshal, K.; Li, X.; Datta, J.; Bai, S.; Pogribny, I.; Pogribny, M.; Huang, Y.; Young, D.; Jacob, S.T. A folate- and methyl-deficient diet alters the expression of DNA methyltransferases and methyl CpG binding proteins involved in epigenetic gene silencing in livers of F344 rats. J. Nutr. 2006, 136, 1522–1527. [Google Scholar] [CrossRef]
- Pogribny, I.P.; Karpf, A.R.; James, S.R.; Melnyk, S.; Han, T.; Tryndyak, V.P. Epigenetic alterations in the brains of Fisher 344 rats induced by long-term administration of folate/methyl-deficient diet. Brain Res. 2008, 1237, 25–34. [Google Scholar] [CrossRef]
- Mikkelsen, K.; Apostolopoulos, V. Vitamin B12, Folic Acid, and the Immune System. In Nutrition and Immunity; Springer: Cham, Switzerland, 2019; pp. 103–114. [Google Scholar]
- Kunisawa, J.; Eri, H.; Izumi, I.; Hiroshi, K.A. Pivotal role of vitamin B9 in the maintenance of regulatory T cells in vitro and in vivo. PLoS ONE 2012, 7, e32094. [Google Scholar] [CrossRef] [Green Version]
- Abe, I.; Shirato, K.; Hashizume, Y.; Mitsuhashi, R.; Kobayashi, A.; Shiono, C.; Sato, S.; Tachiyashiki, K.; Imaizumi, K. Folate-deficiency induced cell-specific changes in the distribution of lymphocytes and granulocytes in rats. Environ. Health Prev. Med. 2013, 18, 78–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metz, J. A high prevalence of biochemical evidence of vitamin B12 or folate deficiency does not translate into a comparable prevalence of anemia. Food Nutr. Bull. 2008, 29, S74–S85. [Google Scholar] [CrossRef] [PubMed]
- Antony, A.C. In utero physiology: Role of folic acid in nutrient delivery and fetal development. Am. J. Clin. Nutr. 2007, 85, 598S–603S. [Google Scholar] [CrossRef] [Green Version]
- Hollingsworth, J.W.; Maruoka, S.; Boon, K.; Garantziotis, S.; Li, Z.; Tomfohr, J.; Bailey, N.; Potts, E.N.; Whitehead, G.; Brass, D.M.; et al. In utero supplementation with methyl donors enhances allergic airway disease in mice. J. Clin. Investig. 2008, 118, 3462–3469. [Google Scholar] [CrossRef] [Green Version]
- Preynat, A.; Lapierre, H.; Thivierge, C.M.; Palin, F.M.; Cardinault, N.; Matte, J.J.; Desrochers, A.; Girard, L.C. Effects of supplementary folic acid and vitamin B12 on hepatic metabolism of dairy cows according to methionine supply. J. Dairy Sci. 2010, 93, 2130–2142. [Google Scholar] [CrossRef] [Green Version]
- Duplessis, M.; Girard, C.L.; Santschi, D.E.; Lefebvre, D.M.; Pellerin, D. Folic acid and vitamin B12 supplement enhances energy metabolism of dairy cows in early lactation. J. Dairy Sci. 2012, 95, 118. [Google Scholar]
- Froese, S.D.; Brian, F.; Matthias, R.B. Vitamin B12, folate, and the methionine remethylation cycle—Biochemistry, pathways, and regulation. J. Inherit. Metab. Dis. 2019, 42, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.W.; Nadea, M.R.; Smith, J.; Smith, D.; Selhub, J. Folate-deficiency-induced homocysteinaemia in rats: Disruption of S-adenosylmethionine’s co-ordinate regulation of homocysteine metabolism. Biochem. J. 1994, 298, 415–419. [Google Scholar] [CrossRef] [Green Version]
- James, S.J.; Miller, B.J.; Mcgarrity, L.J.; Morris, S.M. The effect of folic-acid and/or methionine deficiency on deoxyribonucleotide pools and cell cycle cell cycle distribution in mitogen-stimulated rat lymphocytes. Cell Prolif. 1994, 27, 395–406. [Google Scholar] [CrossRef]
- Girard, C.L.; Matte, J.J. Folic acid and vitamin B12 requirements of dairy cows: A concept to be revised. Livest. Prod. Sci. 2005, 98, 123–133. [Google Scholar] [CrossRef]
- Bailey, L.B.; Gregory, J.F. Folate metabolism and requirements. J. Nutr. 1999, 129, 779–782. [Google Scholar] [CrossRef] [Green Version]
- Girard, C.L.; Matte, J.J. İmpact of B-vitamin supply on major metabolic pathways of lactating dairy cows. Can. J. Anim. Sci. 2006, 86, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Bechdel, S.I.; Honeywell, H.E.; Dutcher, R.A.; Knutsen, M.H. Synthesis of vitamin B in the rumen of the cow. J. Biol. Chem. 1928, 80, 231–238. [Google Scholar]
- Matte, J.J.; Girard, C.L.; Tremblay, G.F. Effect of long-term addition of folic acid on folate status, growth performance, puberty attainment, and reproductive capacity of gilts. J. Anim. Sci. 1993, 71, 151–157. [Google Scholar] [CrossRef]
- Girard, C.L.; Matte, J.J.; Tremblay, G.F. Serum folates in gestating and lactating dairy cows. J. Dairy. Sci. 1989, 72, 3240–3246. [Google Scholar] [CrossRef]
- Girard, C.L.; Lapierre, H.; Matte, J.J.; Lobley, G.E. Effects of dietary supplements of folic acid and rumen-protected methionine on lactational performance and folate metabolism of dairy cows. J. Dairy Sci. 2005, 88, 660–670. [Google Scholar] [CrossRef]
- Ster, C.; Loiselle, M.C.; Lacasse, P. Effect of postcalving serum nonesterified fatty acids concentration on the functionality of bovine immune cells. J. Dairy Sci. 2012, 95, 708–717. [Google Scholar] [CrossRef]
- Khan, M.Z.; Zhang, Z.; Liu, L.; Wang, D.; Mi, S.; Liu, X.; Liu, G.; Guo, G.; Li, X.; Wang, Y.; et al. Folic acid supplementation regulates key immunity-associated genes and pathways during the periparturient period in dairy cows. Asian-Australas. J. Anim. Sci. 2020, 00, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ouattara, B.; Bissonnette, N.; Duplessis, M.; Girard, C.L. Supplements of vitamins B9 and B12 affect hepatic and mammary gland gene expression profiles in lactating dairy cows. BMC. Genom. 2016, 17, 640. [Google Scholar] [CrossRef] [Green Version]
- Shirato, K.; Motohashi, N.; Tanihata, J.; Tachiyashiki, K.; Tomoda, A.; Imaizumi, K. Effects of two types of inactivity on the number of white blood cells in rats. Eur. J. Appl. Physiol. 2006, 98, 590–600. [Google Scholar] [CrossRef]
- Wang, B.; Li, H.; Zhen, L.; Jian, L.; Gao, Y.; Qu, Y.; Liu, C.; Xu, C.; Li, Y.; Diao, Z.; et al. Folic acid supplementation modulates the growth performance, muscle development and immunity of Hu sheep offspring of different litter size. J. Nutr. Biochem. 2019, 70, 194–201. [Google Scholar] [CrossRef]
- Fang, L.; Hou, Y.; An, J.; Li, B.; Song, M.; Wang, X.; Sørensen, P.; Dong, Y.; Liu, C.; Wang, Y.; et al. Genome-wide transcriptional and post-transcriptional regulation of innate immune and defense responses of bovine mammary gland to Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Kosciuczuk, E.M.; Lisowski, P.; Jarczak, J.; Majewska, A.; Rzewuska, M.; Zwierzchowski, L.; Bagnicka, E. Transcriptome profiling of Staphylococci-infected cow mammary gland parenchyma. BMC. Vet. Res. 2017, 13, 161. [Google Scholar] [CrossRef]
- He, Y.S.; Song, M.; Zhang, Y.; Li, X.; Song, J.; Zhang, Y.; Yu, Y. Whole-genome regulation analysis of histone H3 lysin 27 trimethylation in subclinical mastitis cows infected by Staphylococcus aureus. BMC Genom. 2016, 17, 565. [Google Scholar] [CrossRef] [Green Version]
- Croker, A.B.; Danielle, L.K.; Zhang, J.; Sam, W.; Tracy, A.W.; Edouard, G.S.; Lorraine, R.; Christopher, J.G.; Förster, I.; Clausen, B.E.; et al. SOCS3 negatively regulates IL-6 signaling In Vivo. Nat. Immunol. 2003, 4, 540–545. [Google Scholar] [CrossRef]
- Huang, Y.; Feng, Z.; Luo, C.; Zhang, X.; Yu, S.; Sun, Z.; Zhang, L.; Li, Q.; Xue-Jun, G. SOCS3-Mediated Blockade Reveals Major Contribution of JAK2/STAT5 Signaling Pathway to Lactation and Proliferation of Dairy Cow Mammary Epithelial Cells In Vitro. Molecules 2013, 18, 12987–13002. [Google Scholar] [CrossRef] [Green Version]
- Khan, Z.M.; Lei, L.; Zhichao, Z.; Adnan, K.; Di, W.; Siyuan, M.; Tahir, U.; Gang, L.; Gang, G.; Xizhi, L.; et al. Folic acid supplementation regulates milk production variables, metabolic associated genes and pathways in perinatal holsteins. J. Anim. Physiol. Anim. Nutr. 2020, 104, 438–492. [Google Scholar] [CrossRef]
- Graulet, B.; Matte, J.J.; Desrochers, A.; Doepel, L.; Palin, M.F.; Girard, C.L. Effects of dietary supplements of folic acid and vitamin B12 on metabolism of dairy cows in early lactation. J. Dairy Sci. 2007, 90, 3442–3455. [Google Scholar] [CrossRef] [Green Version]
- Girard, C.L.; Matte, J.J. Dietary Supplements of Folic Acid during Lactation: Effects on the Performance of Dairy Cows. J. Dairy Sci. 1998, 81, 1412–1419. [Google Scholar] [CrossRef]
- Preynat, A.; Lapierre, H.; Thivierge, M.C.; Palin, M.F.; Matte, J.J.; Desrochers, A.; Girard, C.L. Influence of methionine supply on the response of lactational performance of dairy cows to supplementary folic acid and vitamin B12. J. Dairy Sci. 2009, 92, 1685–1695. [Google Scholar] [CrossRef]
- Li, H.Q.; Liu, Q.; Wang, C.; Yang, Z.M.; Guo, G.; Huo, W.J.; Pei, C.X.; Zhang, Y.L.; Zhang, S.L.; Wang, H.; et al. Effects of dietary supplements of rumen-protected folic acid on lactation performance, energy balance, blood parameters and reproductive performance in dairy cows. Anim. Feed Sci. Technol. 2016, 213, 55–63. [Google Scholar] [CrossRef]
- Girard, C.L.; Matte, J.J. Effects of intramuscular injections of vitamin B12 on lactation performance of dairy cows fed dietary supplements of folic acid and rumen-protected methionine. J. Dairy Sci. 2005, 98, 671–676. [Google Scholar] [CrossRef]
- Girard, C.L.; Benchaar, C.; Chiquette, J.; Desrochers, A. Net flux of nutrients across the rumen wall of lactating dairy cows as influenced by dietary supplements of folic acid. J. Dairy Sci. 2009, 92, 6116–6122. [Google Scholar] [CrossRef] [Green Version]
- Preynat, A.; Lapierre, H.; Thivierge, M.C.; Palin, M.F.; Matte, J.J.; Desrochers, A.; Girard, C.L. Effects of supplements of folic acid, vitamin B12, and rumen-protected methionine on whole body metabolism of methionine and glucose in lactating dairy cows. J. Dairy Sci. 2009, 92, 677–689. [Google Scholar] [CrossRef]
- Menzies, K.K.; Lefèvre, C.; Sharp, J.A.; Macmillan, K.L.; Sheehy, P.A.; Nicholas, K.R. A novel approach identified the FOLR1 gene, a putative regulator of milk protein synthesis. Mamm. Genome 2009, 20, 498–503. [Google Scholar] [CrossRef]
- Cui, X.; Yali, H.; Shaohua, Y.; Yan, X.; Shengli, Z.; Yuan, Z.; Qin, Z.; Xuemei, L.; George, E.L.; Sun, D. Transcriptional profiling of mammary gland in Holstein cows with extremely different milk protein and fat percentage using RNA sequencing. BMC Genom. 2014, 15, 226. [Google Scholar] [CrossRef] [Green Version]
- Wickramasinghe, S.; Rincon, G.; Islas-Trejo, A.; Medrano, J.F. Transcriptional profiling of bovine milk using RNA sequencing. BMC. Genom. 2012, 13, 45. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.; Lee, H.J.; Kim, K.; Caetano-Anolles, K.; Jeong, J.Y.; Park, S.; Oh, Y.K.; Cho, S.; Kim, H. Characterizing milk production related genes in Holstein using RNA-seq. Asian-Australas. J. Anim. Sci. 2016, 29, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Lemay, D.G.; Hovey, R.C.; Hartono, S.R.; Hinde, K.; Smilowitz, J.T.; Ventimiglia, F.; Schmidt, K.A.; Lee, J.W.S.; Islas-Trejo, A.; Silva, P.I.; et al. Sequencing the transcriptome of milk production: Milk trumps mammary tissue. BMC Genom. 2013, 14, 872. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Jiang, J.; Liu, X.; Wang, H.; Guo, G.; Zhang, Q.; Jiang, L. Differential expression of genes in milk of dairy cattle during lactation. Anim. Genet. 2016, 47, 174–180. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Liu, Q.; Guo, G.; Huo, W.J.; Liang, Y.; Pei, C.X.; Zhang, S.L.; Yang, W.Z.; Wang, H. Effects of different dietary protein levels and rumen-protected folic acid on ruminal fermentation, degradability, bacterial populations and urinary excretion of purine derivatives in beef steers. J. Agric. Sci. 2017, 155, 1477–1486. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Q.; Guo, G.; Huo, W.J.; Ma, L.; Zhang, Y.L.; Pei, C.X.; Zhang, S.L.; Wang, H. Effects of dietary supplementation of rumen-protected folic acid on rumen fermentation, degradability and excretion of urinary purine derivatives in growing steers. Arch. Anim. Nutr. 2016, 70, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Ragaller, V.; Peter, L.; Wiebke, B.; Karl-Heinz, S.; Liane, H.; Gerhard, F. Effects of folic acid supplementation to rations differing in the concentrate to roughage ratio on ruminal fermentation, nutrient flow at the duodenum, and on serum and milk variables of dairy cows. Arch. Anim. Nutr. 2010, 64, 484–503. [Google Scholar] [CrossRef] [PubMed]
- Reece, J.B. Campbell Biology, 9th ed.; Benjamin Cummings: Boston, MA, USA, 2011; p. 143. ISBN 978-0-321-55823-7. [Google Scholar]
- Wagner, C. Biochemical role of folate in cellular metabolism. Clin. Res. Regul. Aff. 2001, 18, 161–180. [Google Scholar] [CrossRef]
- Nafikov, R.A.; Beitz, D.C. Carbohydrate and Lipid Metabolism in Farm Animals. J. Nutr. 2007, 137, 702–705. [Google Scholar] [CrossRef] [Green Version]
- Sawaengsri, H.; Wang, J.; Christina, R.; Josiane, S.; Wu, D.; Jacob, S.; Ligi, P. High folic acid intake reduces natural killer cell cytotoxicity in aged mice. J. Nutr. Biochem. 2016, 30, 102–107. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | S. aureus Treatment | Folic Acid-Treated Group [49] |
|---|---|---|
| PIM1, SOCS3, ATP12A, NFKBIA, DUSP4, ZC3H12, ESPNL, TNFAIP3 | Up-regulated [53] | Down-regulated |
| CX3CR1, ALOX5, KIT, LPL | Down-regulated [53] | Up-regulated |
| C1QA, C1QB, CCL5, MMP9, VNN, BLA-DQB | Up-regulated [54] | Up-regulated |
| ICAM1, CXCL10 | Up-regulated [54] | Down-regulated |
| PPARD, ARG1, PTX3, CD4 | Up-regulated [55] | Up-regulated |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.Z.; Khan, A.; Xiao, J.; Dou, J.; Liu, L.; Yu, Y. Overview of Folic Acid Supplementation Alone or in Combination with Vitamin B12 in Dairy Cattle during Periparturient Period. Metabolites 2020, 10, 263. https://doi.org/10.3390/metabo10060263
Khan MZ, Khan A, Xiao J, Dou J, Liu L, Yu Y. Overview of Folic Acid Supplementation Alone or in Combination with Vitamin B12 in Dairy Cattle during Periparturient Period. Metabolites. 2020; 10(6):263. https://doi.org/10.3390/metabo10060263
Chicago/Turabian StyleKhan, Muhammad Zahoor, Adnan Khan, Jianxin Xiao, Jinhuan Dou, Lei Liu, and Ying Yu. 2020. "Overview of Folic Acid Supplementation Alone or in Combination with Vitamin B12 in Dairy Cattle during Periparturient Period" Metabolites 10, no. 6: 263. https://doi.org/10.3390/metabo10060263