Community Structure of Aquatic Insects Adapted to Lentic Water Environments, and Fine-Scale Analyses of Local Population Structures and the Genetic Structures of an Endangered Giant Water Bug Appasus japonicus



Abstract
:1. Introduction
2. Materials and Methods
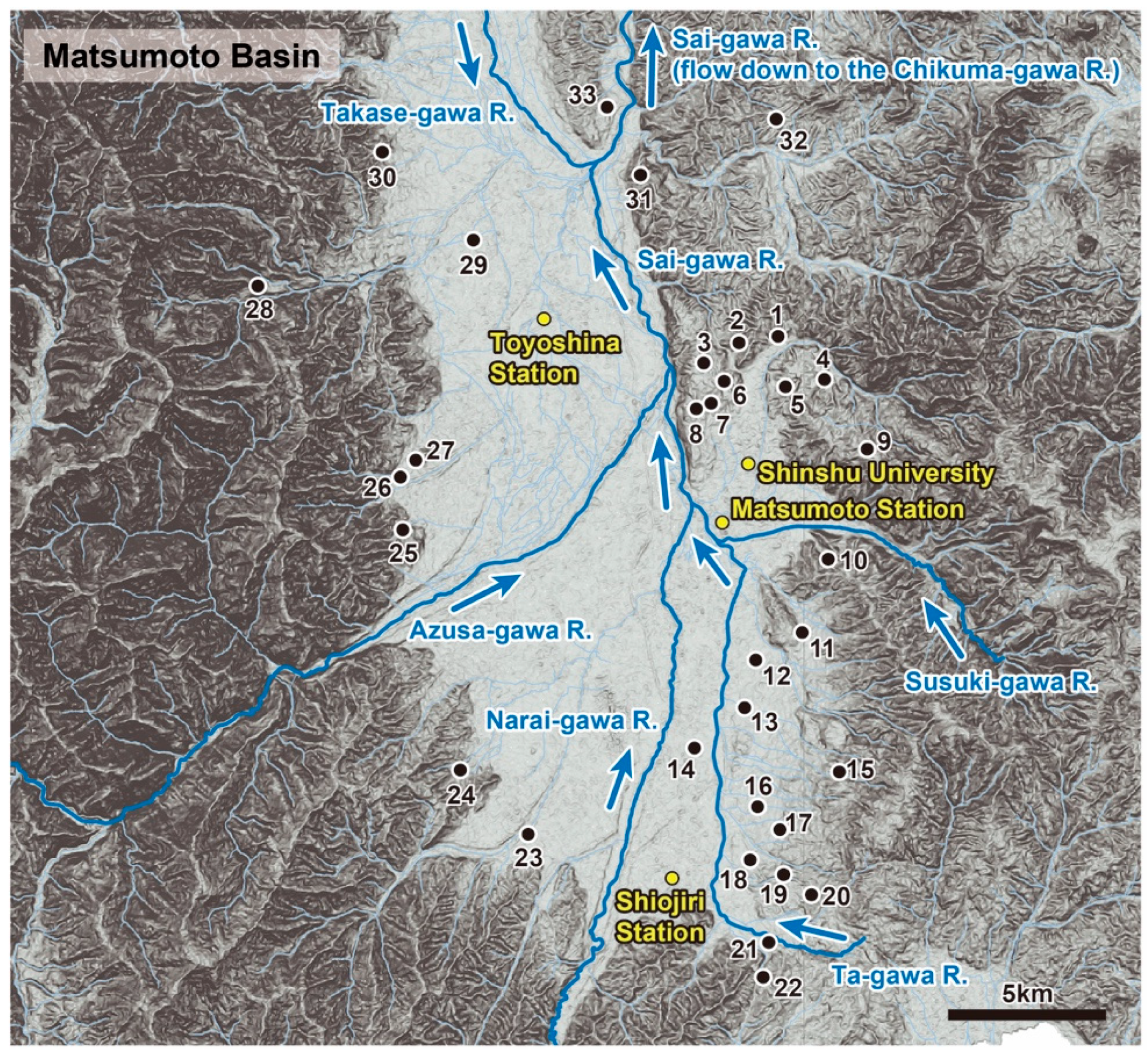
2.1. Research Sites and Target Animals
2.2. Measurement of Environmental Factors at Each Pond
2.3. Sampling for Population and Genetic Structure Analyses of A. japonicus
2.4. Survey on Overwintering Rate for A. japonicus
2.5. Redundancy Analysis (RDA)
2.6. Generalized Linear Model (GLM) Analysis
2.7. Genetic Analysis
2.8. Analysis between Pairwise Genetic and Geographic Distances
2.9. Statistical Analyses Based on the Genetic Data Obtained for A. japonicus
3. Results
3.1. Quantitative Survey of Community Structure among Lentic Water Adapted Aquatic Insect Fauna
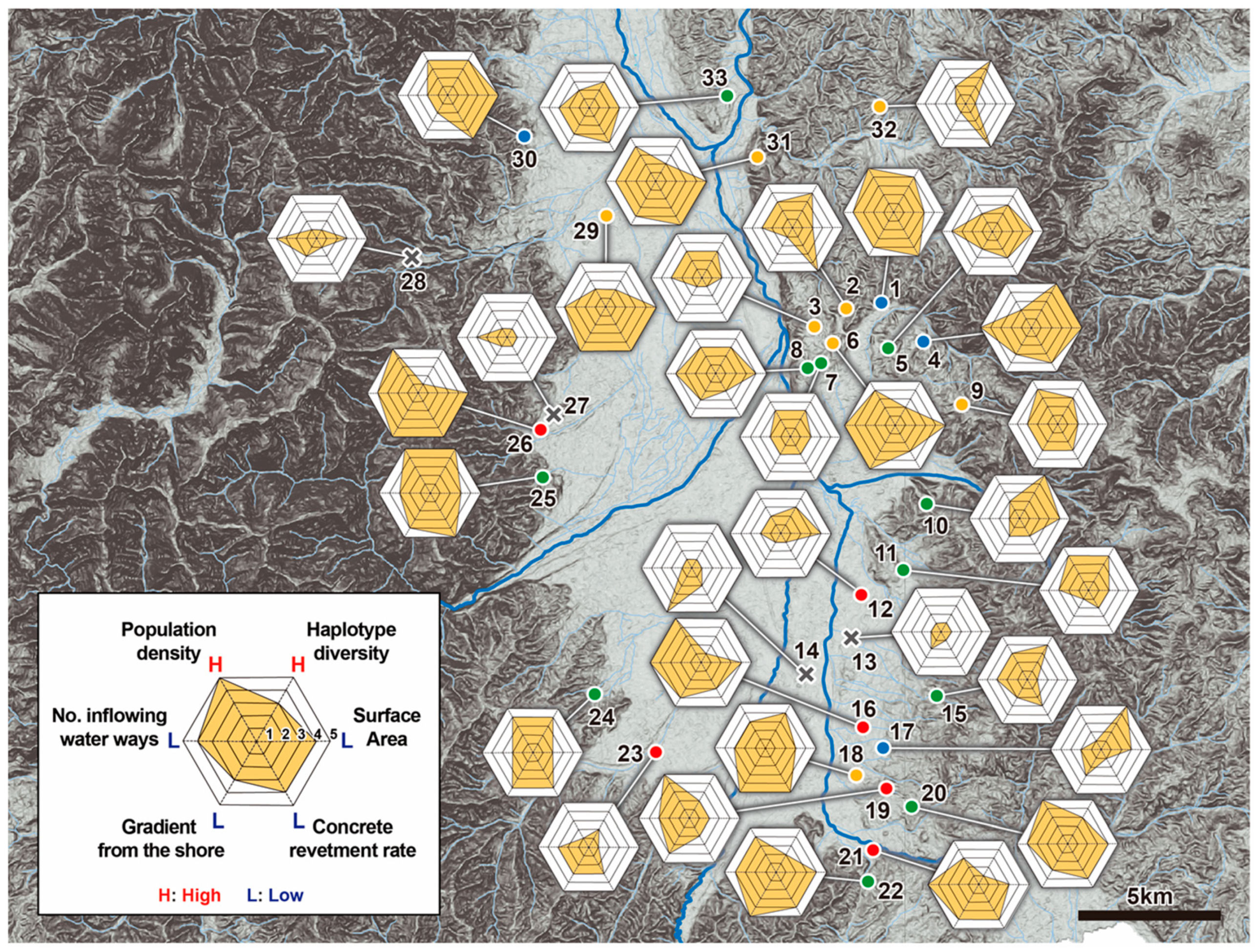
3.2. Relationships between the A. japonicus Populations and Environmental Factors for the 33 Ponds Studied within the Matsumoto Basin
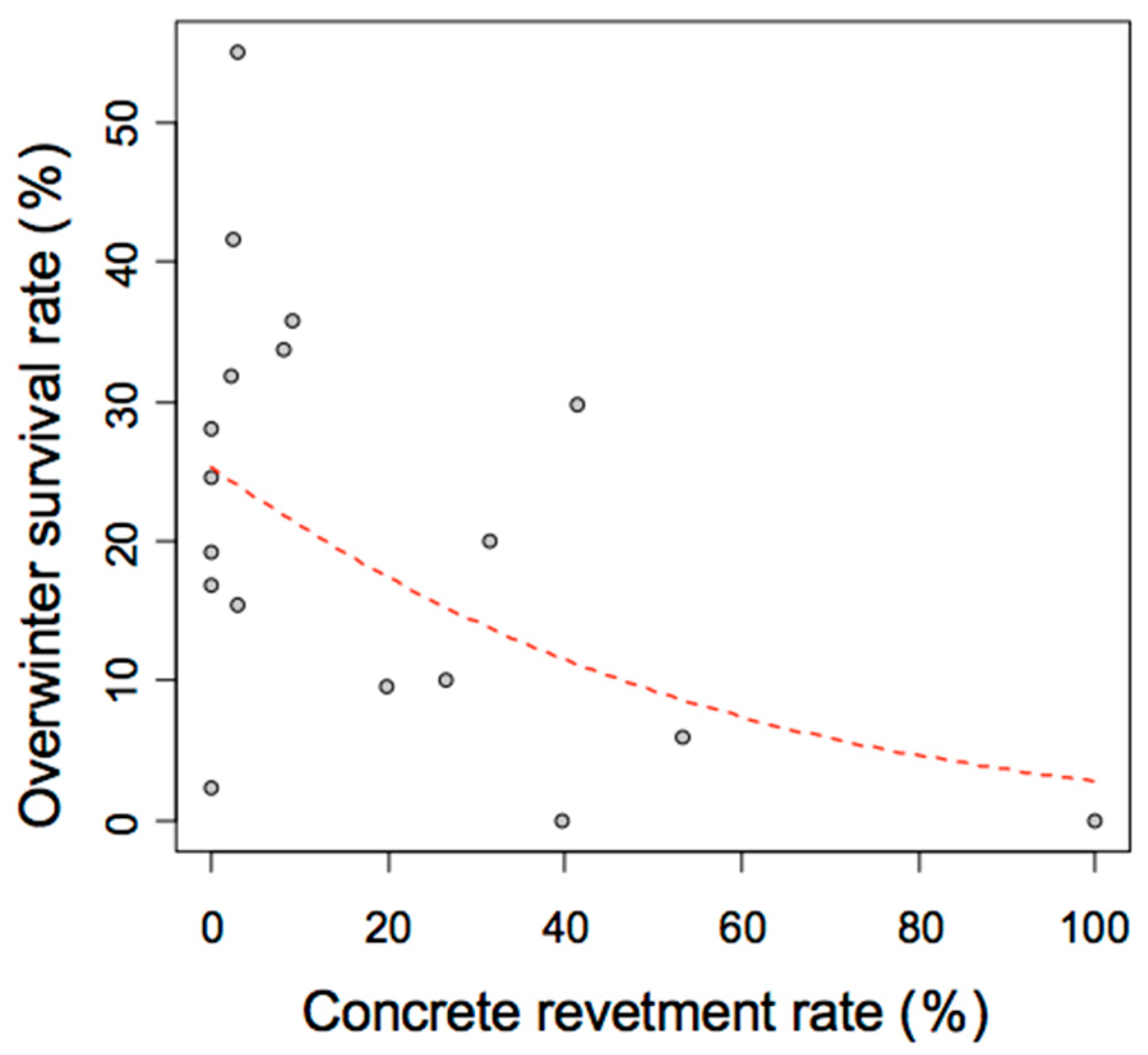
3.3. Survey on the Successful Overwintering Rate of the Giant Water Bug A. japonicus
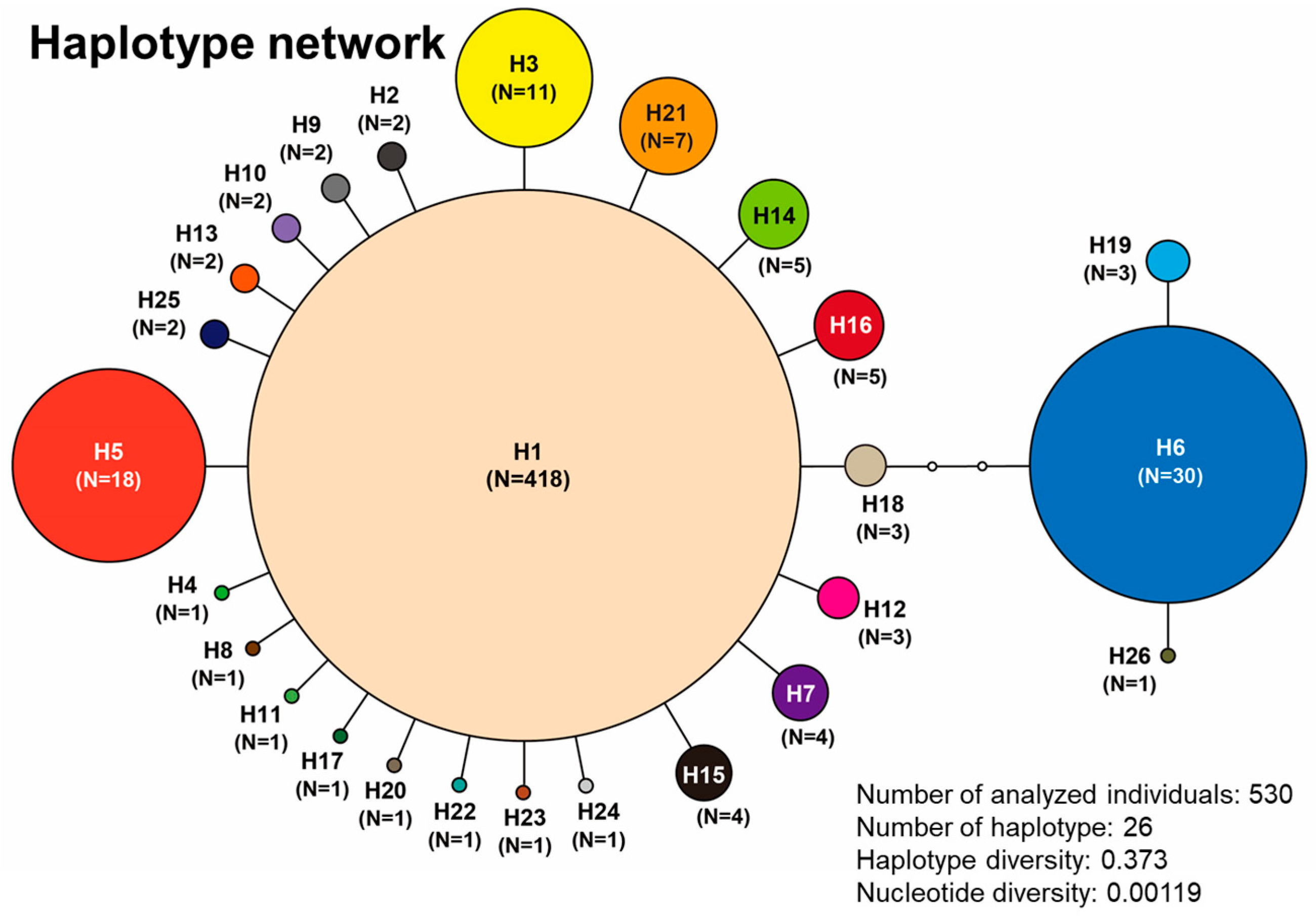
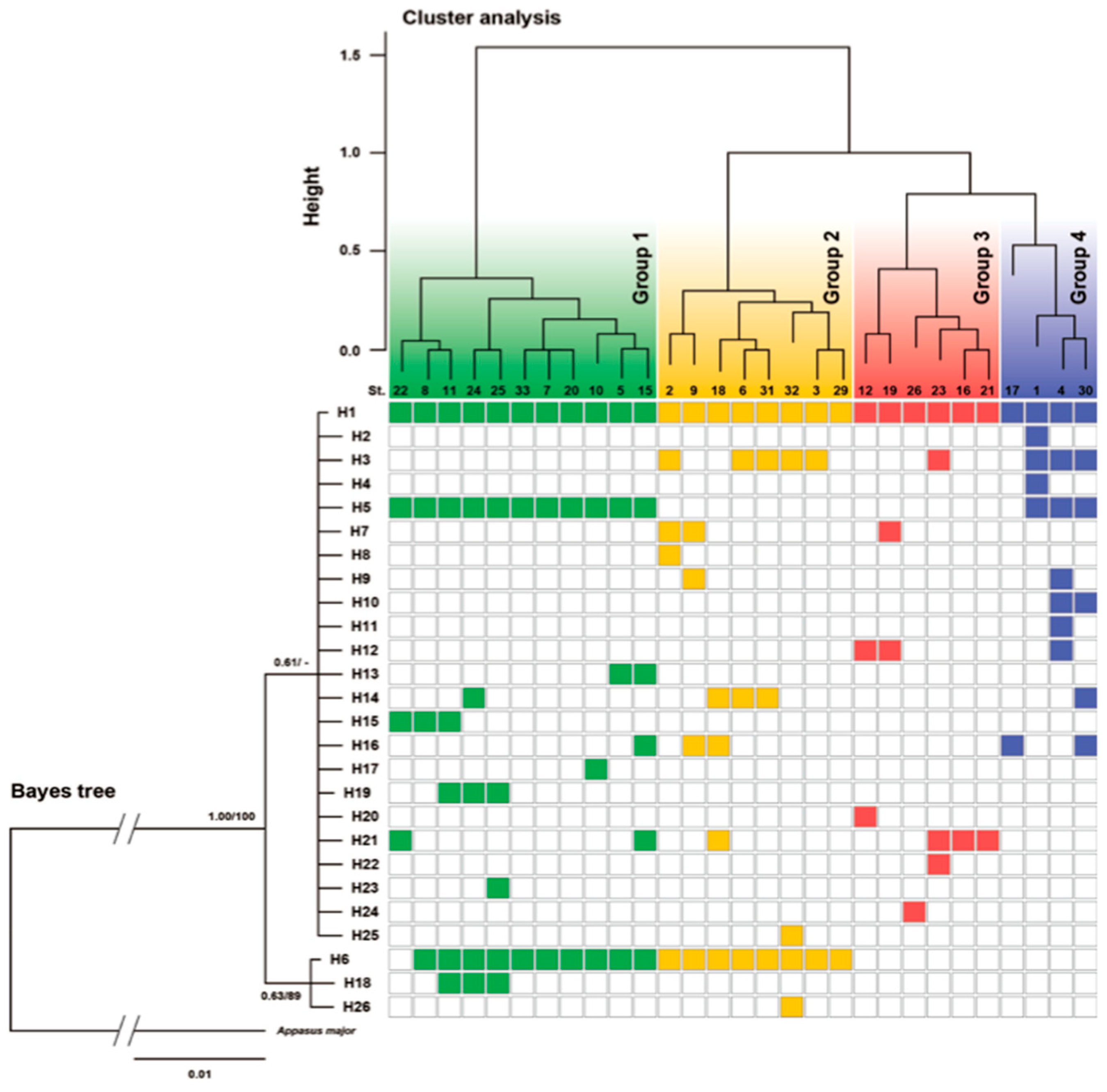
3.4. Results of Genetic Analysis of A. japonicus
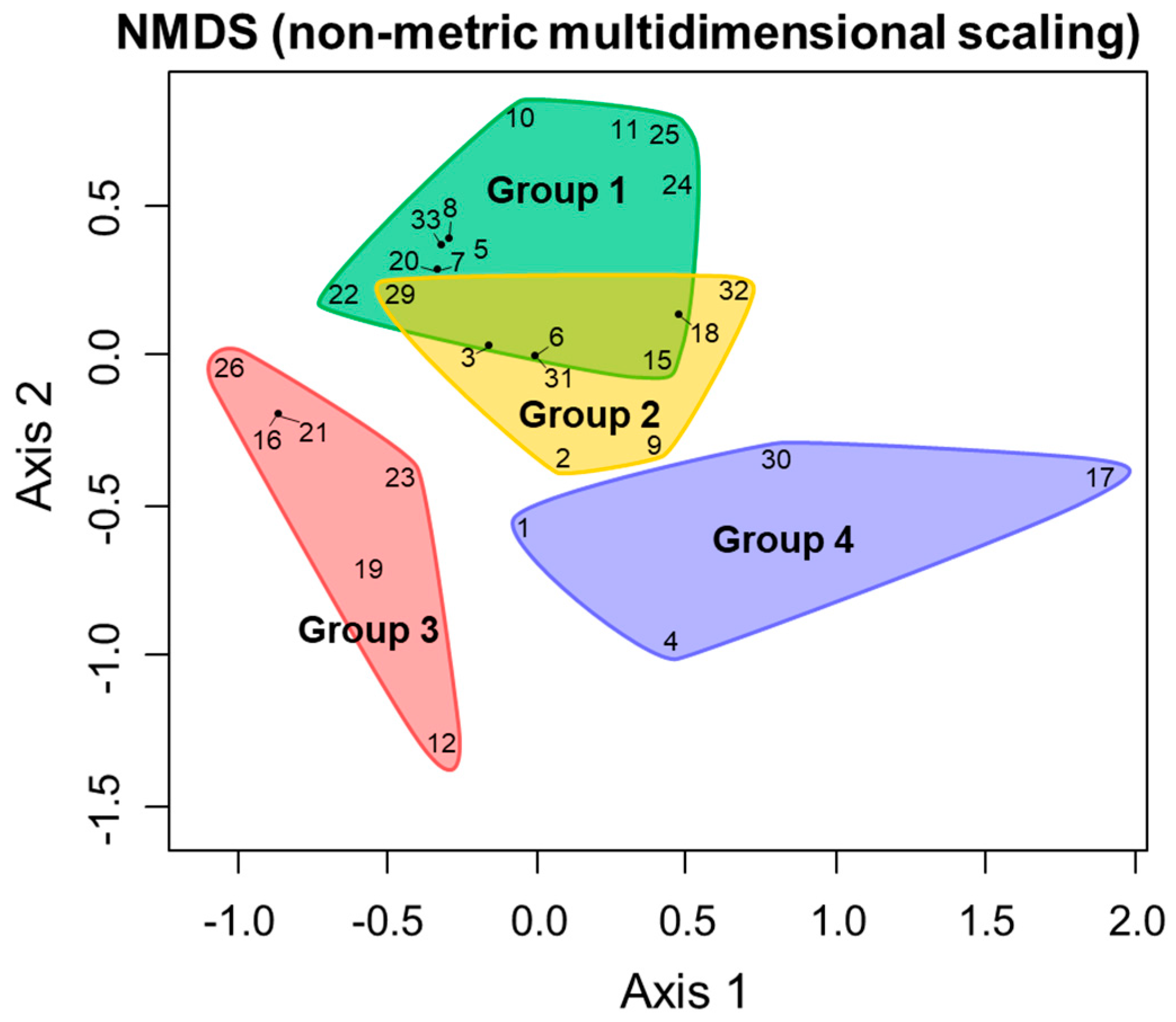
3.5. Comprehensive Evaluation of Population Structure and Genetic Structure of A. japonicus, and Environmental Factors
3.6. Survey on the Successful Overwintering Rate of the Giant Water Bug A. japonicus
4. Discussion
4.1. Environmental Factors Affecting the Community and Population Structure of Aquatic Insects
4.2. Environmental Factors That Influence the Intra-Specific Structure in Several Dominant Species of Aquatic Insects
4.3. Characteristics of Genetic Structure Analysis of A. japonicus
4.4. “Source−Sink” Relationships at the Intra-Specific Group Level
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Céréghino, R.; Boix, D.; Cauchie, H.M.; Martens, K.; Oertli, B. The ecological role of ponds in a changing world. Hydrobiologia 2014, 723, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Yano, K.; Takenaka, M.; Tojo, K. Genealogical position of Japanese populations of the globally distributed mayfly Cloeon dipterum and related species (Ephemeroptera, Baetidae): A molecular phylogeographic analysis. Zool. Sci. 2019, 36, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Yano, K.; Takenaka, M.; Mitamura, T.; Tojo, K. Identifying a “pseudogene for the mitochondrial DNA COI region of the corixid aquatic insect, Hesperocorixa distanti (Heteroptera, Corixidae). Limnology 2020, in press. [Google Scholar] [CrossRef]
- Strong, E.E.; Gargominy, O.; Ponder, W.F.; Bouchet, P. Global diversity of gastropods (Gastropoda; Mollusca) in freshwater. Hydrobiologia 2008, 595, 149–166. [Google Scholar] [CrossRef]
- Lorencová, E.; Horsák, M. Environmental drivers of mollusc assemblage diversity in a system of lowland lentic habitats. Hydrobiologia 2019, 836, 49–64. [Google Scholar] [CrossRef]
- Wiliams, D.D. Environmental constrains in temporary fresh waters and their consequences for the insect fauna. J. N. Am. Benth. Soc. 1996, 15, 634–650. [Google Scholar] [CrossRef] [Green Version]
- Kurokawa, M.; Katano, O.; Tojo, K.; Kitano, S. Environmental factors and distribution of invertebrates in riverside pools in a small stream. Jpn. J. Limnol. 2009, 70, 67–85. [Google Scholar] [CrossRef]
- Katano, O.; Hosoya, K.; Iguchi, K.; Yamaguchi, M.; Aoyama, A.; Kitano, S. Species diversity and abundance of freshwater fishes in irrigation ditches around rice fields. Environ. Biol. Fishes 2003, 66, 107–121. [Google Scholar] [CrossRef]
- Nakano, D.; Nakamura, F. Responses of macroinvertebrate communities to river restoration in a channelized segment of the Shibetsu River, Northern Japan. River Res. Appl. 2006, 22, 681–689. [Google Scholar] [CrossRef]
- Ohba, S. Impact of the invasive crayfish Procambarus charkii on the giant water bug Kirkaldyia deyrolli (Hemiptera) in a rice ecosystems. Jpn. J. Environ. Entomol. 2011, 22, 93–98. [Google Scholar]
- Katayama, N.; Baba, Y.G.; Kusumoto, Y.; Tanaka, K. A review of post-war changes in rice farming and biodiversity in Japan. Agric. Syst. 2015, 132, 73–84. [Google Scholar] [CrossRef]
- Yamazaki, M.; Hamada, Y.; Ibuka, T.; Momii, T.; Kimura, M. Seasonal variations in the community structure of aquatic organisms in a paddy field under a long-term fertilizer trial. Soil Sci. Plant Nutr. 2001, 47, 587–599. [Google Scholar] [CrossRef]
- Ministry of the Environment of Japan. The 4th Version of the Japanese Red List on 9 Taxonomic Groups. Available online: http://www.env.go.jp/press/files/jp/21555 (accessed on 20 April 2020).
- Takenaka, M.; Tokiwa, T.; Tojo, K. Concordance between molecular biogeography of Dipteromimus tipuliformis and geographical history in the local fine scale (Ephemeroptera, Dipteromimidae). Mol. Phylogenet. Evol. 2019, 139, 106547. [Google Scholar] [CrossRef] [PubMed]
- Saito, R.; Kato, S.; Kuranishi, R.B.; Nozaki, T.; Fujino, T.; Tojo, K. Phylogeographic analyses of the Stenopsyche caddisflies (Trichoptera: Stenopsychidae) of the Asian region. Freshw. Sci. 2018, 37, 562–572. [Google Scholar] [CrossRef] [Green Version]
- Tojo, K.; Sekiné, K.; Takenaka, M.; Isaka, Y.; Komaki, S.; Suzuki, T.; Schoville, S.D. Species diversity of insects in Japan: Their origins and diversification processes. Entomol. Sci. 2017, 20, 357–381. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, M.; Tojo, K. Ancient origin of a dipteromimid mayfly family endemic to the Japanese Islands and its genetic differentiation across tectonic faults. Biol. J. Linn. Soc. 2019, 126, 555–573. [Google Scholar] [CrossRef]
- Suzuki, T.; Suzuki, N.; Tojo, K. Parallel evolution of an alpine type ectomorph in a scorpionfly: Independent adaptation to high-altitude environments in multiple mountain locations. Mol. Ecol. 2019, 28, 3225–3240. [Google Scholar] [PubMed]
- Suzuki, T.; Tanizawa, T.; Sekiné, K.; Kunimi, J.; Tojo, K. Morphological and genetic relationship of two closely-related giant water bugs: Appasus japonicus Vuillefroy and Appasus major Esaki (Heteroptera: Belostomatidae). Biol. J. Linn. Soc. 2013, 110, 615–643. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Kitano, T.; Tojo, K. Constructing genetic structure of closely related giant water bugs: Phylogeography of Appasus japonicus and Appasus major (Insecta: Heteroptera, Belostomatidae). Mol. Phylogenet. Evol. 2014, 72, 7–16. [Google Scholar] [CrossRef]
- Sugimoto, R. KASHIMIR 3D for 3DCG Maps & GPS. Available online: https://www.kashmir3d.com/ (accessed on 12 May 2020).
- Ihaka, R.; Gentleman, R. R: A language for data analysis and graphics. J. Comp. Graph. Stat. 1996, 5, 299–314. Available online: http://www.R-project.org (accessed on 12 May 2020).
- Huelsenbeck, J.P.; Ronquist, F. MrBayes: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, A.S. MrBayes5D. Available online: http://www.fifthdimension.jp/ (accessed on 4 June 2020).
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood- based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, A.S. Kakusan: A computer program to automate the selection of a nucleotide substitution model and the configuration of a mixed model on multilocus data. Mol. Ecol. Notes 2007, 7, 962–964. [Google Scholar] [CrossRef]
- Schwarz, G. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Miller, M.P. Alleles in space: Computer software for the joint analysis of interindividual spatial and genetic information. J. Hered. 2005, 96, 722–724. [Google Scholar] [CrossRef] [PubMed]
- Chao, A.; Chazdon, R.L.; Colwell, R.K.; Shan, T. A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecol. Lett. 2005, 8, 148–159. [Google Scholar] [CrossRef]
- Doi, H.; Okamura, H. Similarity indices, ordination, and community analysis tests using the software R. Jpn. J. Ecol. 2011, 61, 3–20. [Google Scholar]
- Clarle, K.R.; Gorley, R.N.; Somerfield, P.J.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E: Devon, UK, 2014. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Nakanishi, K.; Nishida, T.; Kon, M.; Sawada, H. Effects of environmental factors on the species composition of aquatic insects in irrigation ponds. Entomol. Sci. 2014, 17, 251–261. [Google Scholar] [CrossRef]
- Okada, H.; Nakasuji, F. Comparative studies on the seasonal occurrence, nymphal development and food menu in two giant water bugs, Diplonychus japonicus Vuillefroy and Diplonychus major Esaki (Hemiptera: Belostomatidae). Res. Popul. Ecol. 1993, 35, 15–22. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site No. | Locality | City/Village | Latitude (N) | Longitude (E) | Altitude (m) | Surface (m2) | Perimeter Length (m) | Vegetation Rate *1 | Shoreline Vegetation Rate (%) | Concrete Revetment Rate (%) | Gradient from the Shore Mean ± SD (%) | Inflow the Number of Waterways | Outflow the Number of Waterways | Water Transparency Mean ± SD (cm) | TN Mean ± SD (mg/L) | NH4+-N Mean ± SD (mg/L) | PO43−-P Mean ± SD (mg/L) | Genbank Accession No. | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Shinagura | Matsumoto | 36.2923 | 137.9904 | 828 | 940 | 121 | III | 66.7 | 0.0 | 33.7 | ± | 15.8 | 1 | 1 | 46.4 | ± | 6.8 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.13 | ± | 4.71 × 10−2 | LC556393–LC556412 |
| 2 | Okada-ibuka | Matsumoto | 36.2897 | 137.9742 | 776 | 2470 | 206 | II | 100.0 | 0.0 | 66.2 | ± | 29.7 | 2 | 1 | 36.2 | ± | 3.9 | 1.7 | ± | 2.4 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556413–LC556432 |
| 3 | Shimauchi | Matsumoto | 36.2827 | 137.9602 | 848 | 3470 | 231 | II | 12.2 | 100.0 | 61.5 | ± | 17.0 | 2 | 1 | 14.8 | ± | 0.8 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556433–LC556452 |
| 4 | Misayama | Matsumoto | 36.2778 | 138.0091 | 846 | 180 | 55 | II | 57.4 | 5.2 | 42.4 | ± | 16.2 | 0 | 1 | 49.3 | ± | 3.5 | 5.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556453–LC556472 |
| 5 | Hora | Matsumoto | 36.2755 | 137.9929 | 802 | 590 | 95 | II | 49.2 | 10.8 | 46.3 | ± | 12.5 | 1 | 1 | 68.8 | ± | 20.2 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556473–LC556492 |
| 6 | Okada-ibuka | Matsumoto | 36.2768 | 137.9685 | 767 | 120 | 46 | II | 48.5 | 19.8 | 22.5 | ± | 13.8 | 0 | 0 | 30.7 | ± | 0.4 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556493–LC556512 |
| 7 | Okada-shimookada | Matsumoto | 36.2704 | 137.9630 | 736 | 2270 | 188 | II | 17.4 | 39.6 | 48.8 | ± | 20.3 | 3 | 1 | 32.4 | ± | 0.7 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556513–LC556532 |
| 8 | Okada-shimookada | Matsumoto | 36.2679 | 137.9578 | 737 | 650 | 98 | II | 42.5 | 31.5 | 44.5 | ± | 29.3 | 1 | 1 | 56.9 | ± | 5.8 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.13 | ± | 4.71 × 10−2 | LC556533–LC556552 |
| 9 | Satoyamabe | Matsumoto | 36.2545 | 138.0268 | 1205 | 3520 | 237 | II | 57.2 | 9.2 | 42.0 | ± | 25.0 | 2 | 1 | 53.1 | ± | 9.0 | 1.7 | ± | 2.4 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556553–LC556572 |
| 10 | Satoyamabe | Matsumoto | 36.2183 | 138.0109 | 835 | 810 | 120 | II | 31.5 | 39.8 | 54.3 | ± | 26.9 | 4 | 1 | >100.0 | 3.3 | ± | 2.4 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556573–LC556581 | ||
| 11 | Nakayama | Matsumoto | 36.1941 | 138.0004 | 726 | 2540 | 215 | V | 48.4 | 53.2 | 67.5 | ± | 26.7 | 2 | 1 | 83.1 | ± | 10.2 | 1.7 | ± | 2.4 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556582–LC556601 |
| 12 | Kotobuki-toyooka | Matsumoto | 36.1857 | 137.9816 | 646 | 670 | 106 | II | 66.2 | 72.8 | 56.8 | ± | 29.3 | 3 | 1 | 66.8 | ± | 6.6 | 6.7 | ± | 2.4 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556602–LC556611 |
| 13 | Kotobuki-koaka | Matsumoto | 36.1695 | 137.9769 | 661 | 4840 | 268 | II | 40.9 | 59.1 | 47.0 | ± | 19.9 | 4 | 1 | 37.9 | ± | 6.6 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | - |
| 14 | Hirooka-yoshida | Shiojiri | 36.1564 | 137.9558 | 654 | 5950 | 333 | III | 63.4 | 43.8 | 24.5 | ± | 14.8 | 4 | 1 | 77.3 | ± | 22.7 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | - |
| 15 | Kataoka | Shiojiri | 36.1489 | 138.0155 | 1030 | 6000 | 305 | II | 43.2 | 26.6 | 51.1 | ± | 22.6 | 2 | 1 | >100.0 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556612–LC556631 | ||
| 16 | Kataoka | Shiojiri | 36.1374 | 137.9820 | 745 | 540 | 95 | II | 47.0 | 8.1 | 27.8 | ± | 10.7 | 0 | 0 | 77.2 | ± | 15.3 | 11.7 | ± | 9.4 | 1.5 | ± | 0.7 | 0.10 | ± | 1.39 × 10−17 | LC556632–LC556651 |
| 17 | Kataoka | Shiojiri | 36.1296 | 137.9910 | 813 | 2020 | 196 | II | 47.5 | 62.3 | 42.1 | ± | 11.1 | 3 | 1 | 88.4 | ± | 0.4 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556652–LC556654 |
| 18 | Kataoka | Shiojiri | 36.1196 | 137.9787 | 734 | 860 | 134 | II | 81.7 | 0.0 | 24.3 | ± | 18.4 | 2 | 1 | >100.0 | 1.7 | ± | 2.4 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556655–LC556674 | ||
| 19 | Sajiki | Shiojiri | 36.1149 | 137.9922 | 799 | 2600 | 231 | II | 61.5 | 41.3 | 31.4 | ± | 20.7 | 2 | 1 | 83.7 | ± | 2.5 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556675–LC556694 |
| 20 | Kakizawa | Shiojiri | 36.1078 | 138.0040 | 922 | 620 | 104 | II | 71.8 | 3.0 | 43.4 | ± | 13.4 | 1 | 1 | >100.0 | 1.7 | ± | 2.4 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556695–LC556714 | ||
| 21 | Kaminishijo | Shiojiri | 36.0924 | 137.9870 | 781 | 1600 | 161 | II | 97.1 | 2.9 | 34.5 | ± | 13.1 | 0 | 1 | >100.0 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556715–LC556734 | ||
| 22 | Kyu-shiojiri | Shiojiri | 36.0810 | 137.9840 | 916 | 720 | 110 | II | 58.7 | 2.1 | 23.3 | ± | 7.9 | 1 | 1 | >100.0 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556735–LC556754 | ||
| 23 | Nishiseba | Asahi | 36.1276 | 137.8889 | 764 | 3780 | 262 | II | 100.0 | 21.0 | 46.2 | ± | 26.0 | 2 | 1 | >100.0 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556755–LC556774 | ||
| 24 | Kami-oike | Yamagata | 36.1491 | 137.8616 | 799 | 2130 | 186 | II | 65.0 | 2.3 | 40.1 | ± | 19.8 | 3 | 1 | 92.3 | ± | 5.7 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556775–LC556794 |
| 25 | Azusagawa-azusa | Matsumoto | 36.2285 | 137.8381 | 809 | 1480 | 160 | II | 30.7 | 0.0 | 38.3 | ± | 16.7 | 2 | 1 | >100.0 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556795–LC556814 | ||
| 26 | Misato-ogura | Azumino | 36.2457 | 137.8376 | 744 | 150 | 64 | III | 100.0 | 0.0 | 9.0 | ± | 3.4 | 1 | 1 | >100.0 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556815–LC556834 | ||
| 27 | Misato-ogura | Azumino | 36.2515 | 137.8433 | 742 | 4860 | 313 | II | 13.9 | 100.0 | 61.9 | ± | 34.2 | 2 | 1 | >100.0 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | - | ||
| 28 | Horigane-karasugawa | Azumino | 36.3085 | 137.7797 | 1178 | 1100 | 149 | II | 14.3 | 100.0 | 52.4 | ± | 18.9 | 1 | 1 | 97.3 | ± | 3.8 | 1.7 | ± | 2.4 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | - |
| 29 | Hotaka-kashiwabara | Azumino | 36.3235 | 137.8671 | 585 | 170 | 59 | II | 92.6 | 0.0 | 23.7 | ± | 15.8 | 1 | 1 | 17.7 | ± | 2.2 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556835–LC556842 |
| 30 | Hotaka-ariake | Azumino | 36.3525 | 137.8301 | 692 | 120 | 57 | II | 38.3 | 0.0 | 49.1 | ± | 15.3 | 3 | 1 | >100.0 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556843–LC556862 | ||
| 31 | Akashina-nakagawate | Azumino | 36.3448 | 137.9341 | 768 | 110 | 46 | II | 100.0 | 0.0 | 31.3 | ± | 8.4 | 1 | 0 | 47.0 | ± | 1.3 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556863–LC556882 |
| 32 | Aida | Matsumoto | 36.3633 | 137.9900 | 774 | 7880 | 502 | II | 96.1 | 0.0 | 64.3 | ± | 37.0 | 4 | 2 | 39.9 | ± | 1.2 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556883–LC556902 |
| 33 | Akashina-nanaki | Azumino | 36.3675 | 137.9211 | 546 | 1100 | 141 | III | 72.6 | 7.6 | 42.6 | ± | 18.9 | 2 | 0 | >100.0 | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | 0.10 | ± | 1.39 × 10−17 | LC556903–LC556922 | ||
| Site No. | n | h | Hd | Pi | Haplotype No. | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H1 | H2 | H3 | H4 | H5 | H6 | H7 | H8 | H9 | H10 | H11 | H12 | H13 | H14 | H15 | H16 | H17 | H18 | H19 | H20 | H21 | H22 | H23 | H24 | H25 | H26 | |||||
| 1 | 20 | 5 | 0.442 | 0.00074 | 15 | 2 | 1 | 1 | 1 | |||||||||||||||||||||
| 2 | 20 | 5 | 0.442 | 0.00120 | 15 | 1 | 1 | 2 | 1 | |||||||||||||||||||||
| 3 | 20 | 3 | 0.353 | 0.00178 | 16 | 1 | 3 | |||||||||||||||||||||||
| 4 | 20 | 7 | 0.521 | 0.01813 | 14 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||
| 5 | 20 | 4 | 0.363 | 0.00105 | 16 | 2 | 1 | 1 | ||||||||||||||||||||||
| 6 | 20 | 4 | 0.284 | 0.00091 | 17 | 1 | 1 | 1 | ||||||||||||||||||||||
| 7 | 20 | 3 | 0.353 | 0.00178 | 16 | 1 | 3 | |||||||||||||||||||||||
| 8 | 20 | 4 | 0.363 | 0.00105 | 16 | 2 | 1 | 1 | ||||||||||||||||||||||
| 9 | 20 | 5 | 0.368 | 0.00106 | 16 | 1 | 1 | 1 | 1 | |||||||||||||||||||||
| 10 | 9 | 4 | 0.583 | 0.00203 | 6 | 1 | 1 | 1 | ||||||||||||||||||||||
| 11 | 20 | 6 | 0.516 | 0.00186 | 14 | 1 | 1 | 2 | 1 | 1 | ||||||||||||||||||||
| 12 | 10 | 3 | 0.378 | 0.00061 | 8 | 1 | 1 | |||||||||||||||||||||||
| 15 | 20 | 6 | 0.447 | 0.00122 | 15 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||
| 16 | 20 | 2 | 0.100 | 0.00015 | 19 | 1 | ||||||||||||||||||||||||
| 17 | 3 | 2 | 0.667 | 0.00101 | 2 | 1 | ||||||||||||||||||||||||
| 18 | 20 | 5 | 0.511 | 0.00174 | 14 | 2 | 1 | 1 | 2 | |||||||||||||||||||||
| 19 | 20 | 3 | 0.195 | 0.00030 | 18 | 1 | 1 | |||||||||||||||||||||||
| 20 | 20 | 3 | 0.353 | 0.00178 | 16 | 1 | 3 | |||||||||||||||||||||||
| 21 | 20 | 2 | 0.100 | 0.00015 | 19 | 1 | ||||||||||||||||||||||||
| 22 | 20 | 4 | 0.284 | 0.00046 | 17 | 1 | 1 | 1 | ||||||||||||||||||||||
| 23 | 20 | 4 | 0.284 | 0.00046 | 17 | 1 | 1 | 1 | ||||||||||||||||||||||
| 24 | 20 | 6 | 0.516 | 0.00219 | 14 | 1 | 2 | 1 | 1 | 1 | ||||||||||||||||||||
| 25 | 20 | 6 | 0.574 | 0.00259 | 13 | 1 | 3 | 1 | 1 | 1 | ||||||||||||||||||||
| 26 | 20 | 2 | 0.100 | 0.00015 | 19 | 1 | ||||||||||||||||||||||||
| 29 | 8 | 2 | 0.250 | 0.00152 | 7 | 1 | ||||||||||||||||||||||||
| 30 | 20 | 6 | 0.516 | 0.00090 | 14 | 2 | 1 | 1 | 1 | 1 | ||||||||||||||||||||
| 31 | 20 | 4 | 0.284 | 0.00091 | 17 | 1 | 1 | 1 | ||||||||||||||||||||||
| 32 | 20 | 5 | 0.626 | 0.00278 | 12 | 2 | 3 | 2 | 1 | |||||||||||||||||||||
| 33 | 20 | 3 | 0.353 | 0.00102 | 16 | 3 | 1 | |||||||||||||||||||||||
| Total | 530 | 418 | 2 | 11 | 1 | 18 | 30 | 4 | 1 | 2 | 2 | 1 | 3 | 2 | 5 | 4 | 5 | 1 | 3 | 3 | 1 | 7 | 1 | 1 | 1 | 2 | 1 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomita, K.; Suzuki, T.; Yano, K.; Tojo, K. Community Structure of Aquatic Insects Adapted to Lentic Water Environments, and Fine-Scale Analyses of Local Population Structures and the Genetic Structures of an Endangered Giant Water Bug Appasus japonicus. Insects 2020, 11, 389. https://doi.org/10.3390/insects11060389
Tomita K, Suzuki T, Yano K, Tojo K. Community Structure of Aquatic Insects Adapted to Lentic Water Environments, and Fine-Scale Analyses of Local Population Structures and the Genetic Structures of an Endangered Giant Water Bug Appasus japonicus. Insects. 2020; 11(6):389. https://doi.org/10.3390/insects11060389
Chicago/Turabian StyleTomita, Kazuhiro, Tomoya Suzuki, Koki Yano, and Koji Tojo. 2020. "Community Structure of Aquatic Insects Adapted to Lentic Water Environments, and Fine-Scale Analyses of Local Population Structures and the Genetic Structures of an Endangered Giant Water Bug Appasus japonicus" Insects 11, no. 6: 389. https://doi.org/10.3390/insects11060389