
Abstract
Two-day-old honeybee workers (± 6 h) were placed in cages and supplied with sucrose solution (1 mol/dm3) ad libitum. Subsequently, the cages with bees were placed in an electric field (E-field) exposure system with field intensities of 5.0 kV/m, 11.5 kV/m, 23 kV/m, and 34.5 kV/m. The duration of exposure was 1 h, 3 h, and 6 h. The biochemical parameters SOD (superoxide dismutase), CAT (catalase), FRAP (ferric ion reducing antioxidant power), and also acidic, neutral, and alkaline proteases in the worker bee hemolymph were analyzed. The E-field increased activities of antioxidant systems, especially SOD, and also the proteolytic systems. In the groups: 11.5 kV/m–time 6 h, 23.0 kV/m–time 1 h, and 34.5 kV/m–time 1 h, FRAP levels were decreased in comparison with the control samples. These findings are discussed in context with possible consequences for honeybee health in urban and rural environments.
Similar content being viewed by others

1 Introduction
All living organisms are exposed to electromagnetic fields (EMF) emitted by electrical and electronic devices, power lines, and RF (radio frequency) radiation from wireless devices such as cordless telephones or the antenna of mobile network base stations (Hardell and Sage 2008). Life on earth is adapted to the natural electromagnetic field emitted by lightning, other planets, and animals (Coe et al. 1995). The constantly developing technologies and the enlarging demand for electricity contribute to the increase of artificial electromagnetic fields in the environment (reviewed in Kim et al. 2019). Bees, against negative factors, have evolved defense mechanisms at individual (immunity of 1—innate: humoral and cellular; and 2—acquired) and colony (especially behavior as a “superorganism”) levels. Both types of reactions complement each other (Simone-Finstrom 2017; reviewed in Strachecka et al. 2018). The first line of individual defense is anatomic-physiological barriers and the cascade of proteolytic enzymes that are activated in the cuticle and the body fat when the immune system detects a pathogen or non-self-substance (Bíliková et al. 2001; Evans and Lopez 2004). When the pathogen breaks down the anatomical-physiological barriers, the humoral response, which includes the antioxidant and proteolytic systems, is activated in the insect organism. Insect enzymes block pathogen enzymes by competitive and/or non-competitive inhibition and activate processes in the body fat whose effect is the translation and transformation of immune proteins (Nazzi et al. 2014; Evans et al. 2006; Nazzi and Pennacchio 2018). During these reactions, reactive oxygen species (ROS) are formed, which are inactivated and/or detoxified by the antioxidant system, especially by superoxide dismutase (SOD). SOD catalyzes the dismutation of superoxide radicals to hydrogen peroxide (H2O2) and, with the further help of catalase (CAT), to water (H2O) and molecular oxygen (O2). (Harrison and Bonning 2010; Farjan et al. 2012; Słowińska et al. 2016). Schuà (1954) showed that the honeybee avoided being exposed to a static EF (1.5 kV/m) on feeding places. Altmann (1959) established that honeybee oxygen consumption increased by 15% in a static EF (1.4–2.8 kV/m). Bindokas et al. (1988a, 1988b) proved that exposure to an intense EF (7 kV/m) caused a disturbance only if the bees were in contact with a conductive substrate. Honeybees exposed to conductive tunnels contributed to increased mortality and abnormal propolization. Honeybee exposure to 60-Hz EF > 150 kV/m caused vibrations of the wings, antennae, and body hairs (Bindokas et al. 1989). Shepherd et al. (2018) showed that exposure to an extremely low-frequency EMF impaired learning ability and flight dynamics, reducing the success of foraging and feeding by the honeybee. Odemer and Odemer (2019) proved that mobile phone radiation may modify honeybee pupal development but no further impairment is manifested in adulthood. This suggests that the increase in environmental pollution by artificial electromagnetic fields poses a further challenge to the honeybee which it has to face every day. Keep in mind that the loss of bees contributes to the impairment of fruit and vegetable productivity, resulting in economic and, above all, environmental impacts. If an EMF exerts such an effect on the behavior of bees, then we hypothesize that this factor may also contribute to lowering the sensitivity of the biochemical protective barriers of the body. Therefore, we hypothesized that an electric field (E-field) at 50 Hz has a non-negligible effect on the antioxidant and proteolytic systems of honeybee workers (hypothesis 1). The longer exposure of the bees to the E-field, the deeper changes in their organisms (hypothesis 2). The overall aim of our studies was to show the effect of a 50-Hz E-field on honeybee workers in controlled laboratory conditions.
2 Material and methods
2.1 Test organisms
The experiment was carrying out from 15 June to 15 August 2019. Twenty honeybee (Apis mellifera carnica) colonies from the Wroclaw University of Environmental and Life Sciences apiary were treated against Varroa destructor using amitraz fumigation four times at 4-day intervals, a 12.5 mg/tablet (Amitraz® Biowet Pulawy), before starting the experiment. To monitor the number of Nosema spp. spores, the hemocytometer method was used (30 bees per hive in three repetitions). Eight-day-old A. mellifera carnica L. queens originating from the same mother-queen were artificially inseminated with the semen of drones from the same father-queen colony for standardized research material. Inseminated queens were individually introduced into the queen-less colonies and kept in isolators. Each colony contained one empty comb. After 12 h of egg laying, the queens were released and isolators containing combs with eggs were left within the colonies for further worker-brood rearing. On the 19th day of apian development, the combs with the already sealed worker brood were transferred to an incubator (temperature of 34.4 °C ± 0.5 °C and relative humidity of 70% ± 5%) in which they were maintained within individual chambers for 1-day-old workers to emerge.
2.2 Experimental setup
For adaptation, 1-day-old workers were placed within 150 wooden cages (20 × 15 × 7 cm), each containing 100 workers and two inner feeders with sucrose solution (Chempur®) at a concentration of 1 mol/dm3 ad libitum. The adaptation process lasted 24 h at a temperature of 25 °C ± 0.5 °C and relative humidity of 70% ± 5%. Caged bees were maintained in the incubator in the same conditions described above until being used for the experiment. Dead bees were utilized. Bees were divided into 12 groups (Table I). Each group consisted of ten cages. Bees in the experimental groups were exposed to the following E-field intensities: 5.0 kV/m, 11.5 kV/m, 23.0 kV/m, and 34.5 kV/m for 1 h, 3 h, and 6 h. The control groups were not treated to the artificial electromagnetic field, they were under the influence of an electromagnetic field < 2.00 kV/m. The group name is the combination of the E-field intensity and the exposure time. For example, the name of group with bees exposed to a 5 kV/m E-field for 1 h is 5 kV/m1h. The control group was marked with the letter C. Bees from the control group were collected at the same time as the other groups.
2.3 E-field exposure
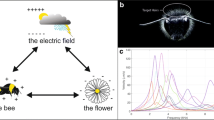
A homogeneous 50-Hz E-field was generated in the exposure system in the form of a plate capacitor with distance of 20 cm between two electrodes constructed as a squared cage made out of wire mesh (dimension 100 cm × 100 cm) with a 10 mm × 10 mm grid diameter. Electrodes were connected to a high-voltage transformer powered by an autotransformer, which allowed the voltage to be adjusted and the E-field intensity to be changed. The field intensity was fixed at 5.0 kV/m, 11.5 kV/m, 23.0 kV/m, and 34.5 kV/m. Changes in the homogeneity and stability of the E-field intensity were no higher than ± 5% in the emitter, to which the bees were exposed during the whole experiment. The E-field intensity and the homogeneity in the test area were verified by measurements made by an LWiMP accredited testing laboratory (certification AB-361 of Polish Centre for Accreditation) using a ESM-100-meter No. 972153 with calibration certificate LWiMP/W/070/2017 dated on 15/02/2017 issued by the accredited calibration laboratory PCA AP-078. The measurements were done in the points of 10 × 10 × 5 cm3 mesh inside an empty emitter (without experimental cages). The stability of the electric field was maintained by permanently monitoring the voltage applied to the exposure system using a control circuit (Figure 7).
2.4 Biochemical analyses
Hemolymph was taken from 100 live bees from each group after exposure by removing the antennae with sterile tweezers. Hemolymph was collected in sterile glass capillaries with a volume of 20 μl end-to-end without anticoagulant. The prepared capillaries were placed in 1.5-ml Eppendorf tubes filled with 150 μl of 0.6% NaCl (sodium chloride). The test tubes were placed on the cooling block during this procedure. The prepared tubes were transferred to a cryo-box and then frozen at – 80 °C (Łoś and Strachecka 2018). Determinations of the acidic, neutral, and alkaline protease activities were done according to the Anson method (1938) modified by Strachecka and Demetraki-Paleolog (2011). Superoxide dismutase (SOD) activities were determined using a commercial Sigma-Aldrich 19160 SOD determination kit. Catalase activities (CAT) were determined using a commercial kit from EnzyChromTMCatalase Assay Kit (ECAT-100). The antioxidant capacity of the hemolymph was determined by FRAP (Ferric ion reducing antioxidant power), and the determination of this parameter was carried out according to a procedure developed by Benzie and Strain (1996) and subsequently modified by Thaipong et al. (2006). Fifteen bees were taken from all groups for each parameter.
2.5 Data evaluation
The normality of the data distribution was analyzed using the Shapiro-Wilk test. The statistical significance of data within groups and between groups was determined by the Kruskal-Wallis test. For all tests, RStudio (R Core Team, 2018) and a significance level of α = 0.05 was used.
3 Results
3.1 Superoxide dismutase activity
In all groups, SOD activities were higher in comparison with the control group. The highest SOD activities were recorded in the 34.5 kV/m1h and 34.5 kV/m6h groups and they were, respectively, 4 and 3.5 times higher than in the control group (Figure 1). The lowest increase in activity, at 25%, was for the 11.5 kV/m3h group. A linear increase of superoxide dismutase activities as a function of time was observed in the 5 kV/m groups. In the case of the other groups, SOD activities changed periodically—high activities were observed for 1-h exposures, then they decreased at 3 h and increased at 6 h.

Superoxide dismutase (SOD) activity in honeybee hemolymph after exposure to the E-field (50 Hz and different intensities). The group name is the combination of the E-field intensity and the exposure time. For example, the name of the group with bees exposed to a 5 kV/m E-field for 1 h is 5 kV/m1h. The control group was marked with the letter C. The control groups were not treated in the artificial electromagnetic field, they were under the influence of an electromagnetic field lower than 2.00 kV/m. Different (capital) letters are statistically different at the P ≤ 0.05 significance level.
3.2 Catalase activity
The highest CAT activities were recorded in the 34.5 kV/m1h group and were 50.31% on average higher than the control group (Figure 2). The 5 kV/m groups had similar values to the control group. In the other groups, these values were higher. In all experimental groups, especially in the 11.5 kV/m and 23.0 kV/m groups, CAT activities changed in waves similar to the SOD activities.

Catalase (CAT) activities in honeybee hemolymph after exposure to the E-field (50 Hz and different intensities). Details as in Figure 1. Different (capital) letters are statistically different at the P ≤ 0.05 significance level.
3.3 Total antioxidant potential
FRAP levels did not show statistically significant differences between groups or within groups (Figure 3). The biggest difference (0.306 μmol/l) was observed between the 11.5 kV/m6h group and the 23 kV/m3h group, which was a fluctuation of 0.6%.

Ferric reducing antioxidant power (FRAP) in honeybee hemolymph after exposure to the E-field (50 Hz and different intensities). Details as in Figure 1. The differences among groups were not statistically significant.
3.4 Protease activity
In all experimental groups, the acidic and neutral protease activities were higher than in the control group (Figures 4 and 5). In the case of alkaline protease activities, this tendency was observed between the 23 kV/m and 34.5 kV/m groups and the control group. A linear increase of acidic protease activities as a function of time (the longer the exposure time) was observed in the 5 kV/m, 11.5 kV/m, and 23 kV/m groups and in the 5 kV/m groups in the case of alkaline protease (but not all results were statistically significant) (Figure 6).

Acidic protease activity in honeybee hemolymph after exposure to the E-field (50 Hz and different intensities). Details as in Figure 1. Different (capital) letters are statistically different at the P ≤ 0.05 significance level.

Neutral protease activity in honeybee hemolymph after exposure to the E-field (50 Hz and different intensities). Details as in Figure 1. Different (capital) letters are statistically different at the P ≤ 0.05 significance level.

Alkaline protease activity in honeybee hemolymph after exposure to the E-field (50 Hz and different intensities). Details as in Figure 1. Different (capital) letters are statistically different at the P ≤ 0.05 significance level.
4 Discussion
4.1 Superoxide dismutase activity
The effect of the electromagnetic field on the bees can be determined using the hemolymph antioxidants (Figure 1) which are the first defensive mechanism of bees (De la Rúa et al. 2001; Farjan et al. 2012; Słowińska et al. 2016). The results of our analysis show that exposure to an E-field impacts the honeybee’s antioxidant system (Figure 7). In all groups, superoxide dismutase activities were higher than in the control group. Nearly fourfold higher SOD activities in the groups treated with an electromagnetic field indicate that this factor may deregulate energy metabolism in mitochondria through interactions with complex I, similar to the acaricides (Motoba et al. 2000). Moreover, the electromagnetic field, as a stressor factor, probably intensifies processes connected with intercepting reactive oxygen forms and binding ions (Augustyniak and Skrzydlewska 2004). In addition, Felton and Summers (1995), Poljšak and Fink (2014), and Schieber and Chandel (2014) established that harmful factors (i.e., pesticides) contributed to an increase in free-radical oxidation abilities which damages the cellular membranes, lipids, proteins, carbohydrates, RNA, and DNA.

Electric field emitter with experimental cages (a) and intensity generator (b).
In our study, the values of SOD activities in the hemolymph of 2-day-old bees in the experimental groups (Figure 1) were similar to the experiment of Strachecka et al. (2014) when the workers were between 18 and 30 days old. The hemolymph of our workers had higher SOD activities than bees at the same age from other studies (Słowińska et al. 2016; Strachecka et al. 2014, 2016). Słowińska et al. (2016) suggested that old bees are less susceptible to the harmful factor and the 1-day-old workers tried to overcome the stress by increasing their antioxidative potential. Only Weirich et al. (2002) presented higher values of antioxidant (in some cases ten times higher) in different tissues (muscle, hemolymph, ventriculi, sperm, and spermatheca) of three castes (queen, worker, drone) in comparison with our research. These differences may have resulted from age and seasonal variations between the workers used in the studies. SOD activities increased in the bee trials from industrial and urbanized areas—the reference sample was bees taken from rural areas (Nikolic et al. 2015). This increase may be caused not only by environmental pollution but also by greater exposure of bees to the effect of the electromagnetic field in urbanized areas than in rural areas. In turn, Ercal et al. (2001) and Farmand et al. (2005) suggested that metals are accumulated in the tissues and increase the formation of free radicals, which are the cause of the increase in SOD and CAT activities. Therefore, the electromagnetic field, by impairing the worker antioxidative defense, consequently may make them more sensitive to this factor, which is in agreement with the suggestions of Gregorc et al. (2020) on the subject of imidacloprid influence.
4.2 Catalase activity
High catalase activities in groups exposed to the intensities of an E-field from 11.5 to 34.5 kV/m may indicate a body’s increased need for this antioxidant enzyme (Figure 2). Catalase is extremely important in animal cells because it breaks down hydrogen peroxide into water and oxygen, thus protecting it from damage. When water deficiency occurs, catalase shows peroxidase activity. Then, hydrogen donors (ethanol, methanol, etc.), thanks to which catalase removes hydrogen peroxide from the cells while oxidizing these compounds, are necessary. Our results for the control group are characteristic for 4-day-old bees. Akyol et al. (2015) presented that catalase activities change very slowly with the age of bees and, according to Orr and Sohal (1992), this may not affect their lifespan. The opposite situation was shown in the studies of Strachecka et al. (2014, 2016) and Farjan et al. (2012), where CAT activities increased with the age of the workers and affected their vitality and longevity. Thus, CAT participates in the aging and senescence of the organism (Münch et al. 2008). So, the increased catalase activities demonstrated in our studies may shorten the life of honeybee workers. Corona et al. (2005) showed that old workers, as foragers, which have the highest levels of flight activity, also have the highest level of antioxidant gene expression, thereby their life gets shorter. The relationship between flight activity, ROS production, and longevity was also similar in the case of Musca domestica (Yan and Sohal 2000). An increase in catalase activities was observed during inflammatory reactions in the organism and the action of mutagenic agents (Adachi et al. 1993). Dussaubat et al. (2012) showed similar increases in catalase and glutathione activities during Nosema spp. infections in the bee midgut. This may indicate that honeybees respond to an electromagnetic field in a similar way as they do to a pathogen.
4.3 Total antioxidant potential (FRAP)
In our studies, there were no statistically significant differences in the FRAP level between groups (Figure 3). A similar observation was in Strachecka et al.’s (2016) studies, where the FRAP levels in the hemolymph of 14- and 21-day-old workers treated with bromfenvinphos did not change. In other studies by Strachecka et al. (2017), differences in the hemolymph of Osmia rufa L. females and males were not seen during the winter months of the diapause. No changes in the FRAP levels are most likely related to the capacity of a system to reduce or break down reactive oxygen species. In our research, the model was 2-day-old workers, which probably did not suffer considerable damage from oxidative stress. In addition, not only do enzymatic antioxidants have an impact on the FRAP value but so do non-enzymatic ones. The latter have not been studied by us. Therefore, when setting the direction of future research, we believe that the concentrations of non-enzymatic antioxidants should also be determined.
4.4 Protease activity
We suppose that electromagnetic fields disturb the functioning of the proteolytic system and the processes in which it is involved: phagocytosis, melanization, cellular adhesion molecule recognition, generation of the reactive intermediates of oxygen and nitrogen, activation of pro-apoptotic molecules, synthesis of cytokines and antimicrobial peptides, enzyme activation, and molecular and hormonal signaling, as well as pathogen protein degrading (Bode et al. 1999; Grzywnowicz et al. 2008). To accurately confirm this assumption, older bees should also be examined in addition to the young ones in our study. The stronger the electromagnetic field and the longer the exposure time, alkaline protease activities were higher than in the control group (Figure 6) This tendency is similar as in the studies by Strachecka et al. (2016) where bees were treated with bromfenvinphos and the reverse was true where coenzyme Q10 and curcumine were used (Strachecka et al. 2014, 2015). According to Strachecka et al. (2010), the proteolytic system on the body surface reacts to changes in the degree of environmental pollution. Moreover, Strachecka et al. (2016) presented that the activities of these proteases increased in the hemolymph of workers treated with bromfenvinphos. Łoś and Strachecka et al. (2018) reported that during contact with unfavorable or harmful factors, protease activities increase drastically on the first of the 3 days and then decrease, and the activities in old bees were lower in the treated group than in the control group. These tendencies are also observed in our studies in young bees (Figure 4). The reverse tendencies are observed when bees are treated with biostimulators and vitamins and they have higher activities of proteases (Łoś and Strachecka 2018). Ashihara and Crozier (2001) reported that substances such as caffeine, coenzyme Q10, and others have long been considered to constitute the chemical defense against biological stressors. Kim et al. (2008) suggest that these substances stimulate the production and/or translocation of proteases and their inhibitors, and signaling molecules. The enzymes involved in the proteolytic system are essential components of insect resistance barriers and are associated with the activities of the antioxidant systems. The honeybee’s proteolytic system, like other organisms, plays an important role in the extracellular and/or intracellular digestion of proteins and as activators of biological processes. It was also shown that the honeybee is characterized by a lower activity of proteases than, for example, the fruit fly (Drosophila melanogaster L.) or mosquitoes of the Anopheles gambiae complex, and it is probably caused by a greater share of the bee’s social immunity. This dependence also applies to other social insects (Evans et al. 2006). Nevertheless, the interaction of acidic, neutral, and alkaline proteases is a very important element that creates and activates the response cascade of the bee organism to environmental changes. Their presence and activity are closely related to the expression of relevant genes (Zou et al. 2006). Our research showed a significant increase in the activities of acidic and neutral proteases in all experimental groups in relation to the control group.
The effectiveness of the processes that allow the fight against free radicals depends on the activities of non-enzymatic antioxidants (coenzyme Q10, lycopene, vitamin E, etc.) and enzymatic antioxidants (SOD, GPx, CAT, etc.) (Seung-Kwon et al. 2013). Variable activities of the antioxidant barrier under the influence of various environmental factors were observed in homogenates of the mitochondrial sperm and muscle fractions (Weirich et al. 2002), and its presence was also confirmed in the venom glands (Peiren et al. 2008) and the hemolymph of honeybee workers (Strachecka et al. 2014, 2016).
5 Conclusion
Our research showed a negative effect of all selected intensities of an E-field at 50 Hz on the organism of honeybee workers in laboratory conditions. The length of exposure to this factor contributes to changes in the activities of antioxidants and proteases. We propose that the electromagnetic field is the factor to be included as potentially threatening the honeybee. This phenomenon should be taken into account in future research.
All biochemical barriers are designed to protect the body from pathogen penetration. When a pathogen enters, the barrier should prevent its spread and destruction of the body. When the functioning of this complex defense system is disturbed, “gates” are created and various stressors can get inside the body. All biochemical mechanisms have one main goal—to maintain homeostasis of the body. Such mechanisms include, among others, antioxidant and proteolytic systems.
References
Adachi S., Nagano S., Ishimori K., Watanabe Y., Morishima I., Egawa T., Kitagawa T., Makino R. 1993. Roles of proximal ligand in heme proteins: replacement of proximal histidine of human myoglobin with cysteine and tyrosine by site-directed mutagenesis as models for P-450, chloroperoxidase, and catalase. Biochemistry. 32, 241–252.
Akyol E., Selamoğlu Z., Gülhan M.F. 2015. Determination of some biochemical parameters of worker honeybees (Apis mellifera L.) belonging to different age groups. Turk. J. Zool.. 39, 995-997.
Altmann G. 1959. Hormon-physiologische Untersuchungen des Stoffwechsels der Honigbiene. Dtsch Bienenwirtsch. 2,74-76
Anson M.L. 1938. Estimation of pepsin, trypsin papain and cathepsin with haemoglobin. J. Gen. Physiol. 22, 79-84.
Ashihara H., Crozier A. 2001. Caffeine: a well known but little mentioned compound in plant science. Trends in Plant Science. 6 (9), 407-413.
Augustyniak, A.; Skrzydlewska, E. Antioxidative abilities during aging. Postȩpy Higieny i Medycyny Doświadczalnej 2004, 58, 194–201.
Bíliková K., Wu G., Šimúth J. 2001. Isolation of a peptide fraction from honeybee royal jellyas a potential antifoulbrood factor. Apidologie. 32 (3), 275-283.
Bindokas V.P., Gauger J.R., Greenberg B. 1988a. Mechanism of biological effects observed in honey bees (Apis mellifera, L.) hived under extra-high-voltage transmission lines: Implications derived from bee exposure to simulated intense electric fields and shocks. Bioelectromagnetics. 9 (3), 285-301.
Bindokas V.P., Gauger J.R., Greenberg B. 1988b. Exposure scheme separates effects of electric shock and electric field for honey bees, Apis mellifera L. Bioelectromagnetics. 9 (3), 275-284.
Bindokas V.P., Gauger J.R., Greenberg B. 1989. Laboratory investigations of the electrical characteristics of honey bees and their exposure to intense electric fields. Bioelectromagnetics. 10 (1), 1-12.
Bode W., Fernandez-Catalan C., Nagase H., Maskos K. 1999. Endoproteinase—Protein inhibitor interaction. Acta Pathologica et Microbiologica Scandinavica.107, 3–10.
Poljšak B., Fink R. (2014) The Protective Role of Antioxidants in the Defence against ROS/RNS-Mediated Environmental Pollution. Oxidative
Coe R.S., Prévot M., Camps P. 1995. New evidence for extraordinarily rapid change of the geomagnetic field during a reversal. Nature. 374 (6524), 687–692.
Corona M., Hughes K.A., Weaver D.B., Robinson G.E. 2005. Gene expression patterns associated with queen honey bee longevity. Mech. Ageing Dev. 126, 1230–1238.
De la Rúa P., Galián J., Serrano J., Moritz R.F.A. 2001. Molecular characterization and population structure of the honeybees from the balearic islands (Spain). Apidologie. 32 (5) 417-427.
Dussaubat C., Brunet J.L., Higes M., Colbourne J.K., Lopez J., Choi J.H., Martín-Hernández R., Botías C., Cousin M., McDonnell C., Bonnet M., Belzunces L.P., Moritz R.F.A., Conte Y.L., Alaux C. 2012. Gut Pathology and Responses to the Microsporidium Nosema ceranae in the Honey Bee Apis mellifera . PLoS One 7(5), e37017.
Ercal N., Gurer-Orhan H., Aykin-Burns N. 2001. Toxic metals and oxidative stress part I: mechanismsinvolved in metal induced oxidative damage. Curr. Top. Med. Chem. 1(6), 529–539.
Evans J.D., Lopez D.L. 2004. Bacterial probiotics induce an immune response in the honey bee (Hymenoptera: Apidae).J. Econ. Entomol. 97(3), 752-6.
Evans J.D., Aronstein K., Chen Y.P., Hetru C., Imler J.L., Jiang H., Kanost M., Thompson G.J., Zou Z., Hultmark D. 2006. Immune pathways and defense mechanisms in honey bees Apis mellifera. Insect Mol. Biol. 15(5), 645-56.
Farjan M., Dmitryjuk M., Lipiński Z., Biernat-Łopieńska E., Żółtowska K. 2012. Supplementation of the honey bee diet with vitamin C: The effect on the antioxidative system of Apis mellifera carnica brood at different stages, J. Apic. Res. 51:3, 263-270, DOI: https://doi.org/10.3896/IBRA.1.51.3.07
Farmand F., Ehdaie A., Roberts C.K., Sindhu R.K. 2005. Lead-induced dysregulation of superoxidedismutases, catalase, glutathione peroxidase, and guanylate cyclase. Environ. Res. 98(1), 33–39.
Felton G.W., Summers C.B. 1955. Antioxidant systems in insects. Archives of Insect Biochemistry and Physiology. 29, 187 - 197.
Gregorc, A., Alburaki, M., Rinderer, N. et al. Effects of coumaphos and imidacloprid on honey bee (Hymenoptera: Apidae) lifespan and antioxidant gene regulations in laboratory experiments. Sci Rep 8, 15003 (2018). https://doi.org/10.1038/s41598-018-33348-4
Grzywnowicz K., Ciołek A., Tabor A., Jaszek M. 2008. Profiles of the body-surface proteolytic system of honey bee queens, workers and drones: Ontogenetic and seasonal changes in proteases and their natural inhibitors. Apidologie. 40, 1, 4–19.
Hardell L., Sage C. 2008. Biological effects from electromagnetic field exposure and public exposure standards. Biomed. Pharmacother. 62(2), 104-9.
Harrison R.L., Bonning B.C. 2010. Proteases as insecticidal agents. Toxins (Basel). 2(5), 935-53.
Iris F.F. Benzie, J.J. Strain, (1996) The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Analytical Biochemistry 239 (1):70-76
Kim J.H., Lee J.K., Kim H.G.,, Kim K.B., Kim H.R. 2019. Possible Effects of Radiofrequency Electromagnetic Field Exposure on Central Nerve System. Biomolecules and Therapeutic (Seoul). 2019;27(3):265‐275. https://doi.org/10.4062/biomolther.2018.152
Łoś, A., Strachecka, A. 2018. Fast and Cost-Effective Biochemical Spectrophotometric Analysis of Solution of Insect “Blood” and Body Surface Elution. Sensors. 18, 1494.
Medicine and Cellular Longevity 2014:1-22
Motoba K., Nishizawa T., Suzuki H., Hamaguchi M., Uchida M., Funayama S. 2000. Species-specific detoxification metabolism of fenpyroximate, a potent acaricide. Pestic. Biochem. Physiol. 67, 73-84.
Münch D., Amdam G.V., Wolschin F. 2008. Ageing in a eusocial insect: molecular and physiological characteristics of life span plasticity in the honey bee. Funct. Ecol. 22, 3, 407-421.
Nazzi F.; Pennacchio F. 2018. Honey Bee Antiviral Immune Barriers as Affected by Multiple Stress Factors: A Novel Paradigm to Interpret Colony Health Decline and Collapse. Viruses. 10, 159.
Nazzi F., Annoscia D., Caprio E., Di Prisco G., & Pennacchio F. 2014. Honeybee immunity and colony losses. Entomologia. 2(2). https://doi.org/10.4081/entomologia.2014.203
Nikolic T., Purac J., Orčić S., Kojić D., Vujanović D., Stanimirović Z., Gržetić I., Ilijević K., Šikoparija B., Blagojević D.P. 2015. Environmental Effects on Superoxide Dismutase and Catalase Activity and Expression in Honey Bee. Arch. Insect Biochem. Physiol. 90 (4), 181-94.
Odemer R., Odemer F. 2019. Effects of radiofrequency electromagnetic radiation (RF-EMF) on honey bee queen development and mating success. Sci. Total Environ. 15 (661), 553-562.
Orr W.C., Sohal R.S. 1992. The effects of catalase gene overexpression on life span and resistance to oxidative stress in transgenic Drosophila melanogaster. Arch. Biochem. Biophys. 297, 35–41.
Peiren N., Graaf D.C., Vanrobaeys F., Danneels E.L., Devreese B., van Beeumen J., Jacobs F.J. 2008. Proteomic analysis of the honey bee worker venom gland focusing on the mechanisms of protection against tissue damage. Toxicon. 52, 72–83.
Schieber M., Chandel N. S., (2014) ROS Function in Redox Signaling and Oxidative Stress. Current Biology 24 (10):R453-R462
Schuà L. 1954. Die Wirkung von luftelektrischen Feldern auf Tiere. Verhandlungen der Deutschen Zoologischen Gesellschaft. 18, 435-440.
Seung-Kwon M., Woong J., Belong C., Seung-Won O., Sang Min P., Bon-Kwon K., Byung-Joo P. 2013. Efficacy of vitamin and antioxidant supplements in prevention of cardiovascular disease: systematic review and meta-analysis of randomised controlled trials. BMJ. 346, f10.
Shepherd S., Lima M. A. P., Oliveira E. E., Sharkh S. M., Jackson C. W., Newland P. L. 2018. Extremely Low Frequency Electromagnetic Fields impair the Cognitive and Motor Abilities of Honey Bees. Nature. 8 (7932).
Simone-Finstrom M.: Social Immunity and the Superorganism: Behavioral Defenses Protecting Honey Bee Colonies from Pathogens and Parasites. Bee World 2017, 94, 21-29.
Słowińska, M., Nynca, J., Wilde, J. Bąk B., Siuda M., Ciereszko A. 2016. Total antioxidant capacity of honeybee haemolymph in relation to age and exposure to pesticide, and comparison to antioxidant capacity of seminal plasma. Apidologie. 47: 227. https://doi.org/10.1007/s13592-015-0391-9
Strachecka A., Demetraki-Paleolog J. 2011. System proteolityczny powierzchni ciała Apis mellifera w zachowaniu zdrowotności rodzin pszczelich. KOSMOS – Problemy Nauk Biologicznych. 60,1–2, 290–291, 43–51.
Strachecka A.J., Gryzińska M.M., Krauze M. 2010. Influence of Environmental Pollution on the Protective Proteolytic Barrier of the Honey Bee Apis mellifera mellifera. Pol. J. Environ. Stud. 19(4), 855–859.
Strachecka A., Olszewski K., Paleolog J., Borsuk G., Bajda M. 2014. Coenzyme Q10 treatments influence the lifespan and key biochemical resistance systems in the honeybee, Apis mellifera, Arch. Insect Biochem. Physiol. 86, 165–179.
Strachecka A.J., Olszewski K., Paleolog J., 2015. Curcumin Stimulates Biochemical Mechanisms of Apis Mellifera Resistance and Extends the Apian Life-Span.Journal of Apicultural Science.59, 1, 129-141.
Strachecka, A., Olszewski, K. and Paleolog, J. 2016. Varroa treatment with bromfenvinphos markedly suppresses honeybee biochemical defense levels. Entomol. Exp. Appl. 160(1), 57-71.
Strachecka A., Chobotow J., Paleolog J., Łoś A., Schulz M., Teper D., Kucharczyk H., Grzybek M. 2017. Insights into the biochemical defense and methylation of the solitary bee Osmia rufa L: A foundation for examining eusociality development. PLoS One 12(4), e0176539.
Strachecka A., Łoś A., Filipczuk J., Schulz M. 2018. Individual and social immune mechanisms of the honey bee (Apis mellifera). Vet. Med. 74 (7), 426-433.
Thaipong K., Boonprakob U., Crosby K., Cisneros-Zevallos L., Byrne D. 2006. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 19, 669-675.
Weirich G.F., Collins A.M., Williams V.P. 2002. Antioxidant enzymes in the honey bee, Apis mellifera. Apidology. 33, 3–14.
Yan L-J., Sohal R. S., (2000) Prevention of flight activity prolongs the life span of the housefly, Musca domestica, and attenuates the age-associated oxidative damamge to specific mitochondrial proteins. Free Radical Biology and Medicine 29 (11):1143-1150
Zou Z., Lopez D.L., Kanost M.R., Evans J.D., Jiang H. 2006. Comparative analysis of serine protease-related genes in the honey bee genome: possible involvement in embryonic development and innate immunity. Insect Mol. Biol.15 (5), 603–614.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Manuscript editor: Yves Le Conte
Modification de certains paramètres biochimiques de l’abeille sous l’influence d’un champ électrique à 50 Hz et d’intensités variables.
champ électromagnétique / abeille / paramètres biochimiques / antioxydants / système protéolytique.
Veränderungen in ausgewählten Parametern bei Honigbienen unter dem Einfluss eines elektrischen Felds von 50 Hz und variabler Intensität.
Elektromagnetisches Feld / Honigbiene / biochemische Parameters / Antioxidantien / proteolytisches System.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Migdał, P., Roman, A., Strachecka, A. et al. Changes of selected biochemical parameters of the honeybee under the influence of an electric field at 50 Hz and variable intensities. Apidologie 51, 956–967 (2020). https://doi.org/10.1007/s13592-020-00774-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13592-020-00774-1