Small RNAs as Fundamental Players in the Transference of Information During Bacterial Infectious Diseases
 Juan José González Plaza
Juan José González Plaza- Faculty of Forestry and Wood Sciences, Czech University of Life Sciences Prague, Prague, Czechia
Communication shapes life on Earth. Transference of information has played a paramount role on the evolution of all living or extinct organisms since the appearance of life. Success or failure in this process will determine the prevalence or disappearance of a certain set of genes, the basis of Darwinian paradigm. Among different molecules used for transmission or reception of information, RNA plays a key role. For instance, the early precursors of life were information molecules based in primitive RNA forms. A growing field of research has focused on the contribution of small non-coding RNA forms due to its role on infectious diseases. These are short RNA species that carry out regulatory tasks in cis or trans. Small RNAs have shown their relevance in fine tuning the expression and activity of important regulators of essential genes for bacteria. Regulation of targets occurs through a plethora of mechanisms, including mRNA stabilization/destabilization, driving target mRNAs to degradation, or direct binding to regulatory proteins. Different studies have been conducted during the interplay of pathogenic bacteria with several hosts, including humans, animals, or plants. The sRNAs help the invader to quickly adapt to the change in environmental conditions when it enters in the host, or passes to a free state. The adaptation is achieved by direct targeting of the pathogen genes, or subversion of the host immune system. Pathogens trigger also an immune response in the host, which has been shown as well to be regulated by a wide range of sRNAs. This review focuses on the most recent host-pathogen interaction studies during bacterial infectious diseases, providing the perspective of the pathogen.
Introduction
“Dizese calétura epidemia porque es común a muchos: perniciosa, porque mata a muchos quanto es de su parte, por tener mucha actividad de calor proveniente (como despues diremos) de vn podrecimiento extremo.”
“It is called an epidemic fever because it is common to many: pernicious, because it kills many on its part, due to a lot of heat activity originating (as we will say later) from extreme rotting. de Viana (1637); plague that heavily struck the port city of Málaga.”
A continuous flow of myriads of energy and matter from atomic to beyond gravitational level shapes life on Earth. Information, its structure and movement across hierarchies, has played a paramount role on the evolution of all living or extinct organisms since the appearance of life. For instance, the early precursors of life were information molecules based in primitive RNA forms (Higgs and Lehman, 2015; Taylor, 2016). The fact that all biological species live in networks, force them to interact with other living beings and the environment (Bermúdez-Barrientos et al., 2020). Interactions, different forms of sociality, transference or reception of information, determine how species perform through evolutionary history. Communication in biology or biocommunication reaches far beyond the human concept of language (Witzany, 2010), and includes a wide variety of forms: sexual deception (color display in orchids to attract pollinators) (Streinzer et al., 2009), vibroacoustic or chemical alarm signaling (alarm pheromones in termites) (Cristaldo et al., 2016), or camouflage (How et al., 2017). Additionally, it encompass hostile communication forms, such as signals to attack among the microbiome (antibiotic production for elimination of competitive bacteria) (Fischbach, 2009; Romero et al., 2011), or counterfeit the defenses of others (fungal RNAs suppressing plant defenses) (Weiberg et al., 2013). Success or failure in the transmission of these signals dramatically affects the prevalence or disappearance of a given set of genes, the basis of Darwinian paradigm (Langdon, 2016).
Infectious diseases are biological examples of competition between species for the same metabolic resources (Herrera and Nunn, 2019), relationships that have been shaped through evolution. In essence, those are evolutionary arms races between hosts and their infectious agents (Ingle et al., 2006; Kuijl and Neefjes, 2009). An infectious disease is a malady caused by a pathogenic organism, including bacteria, fungi, parasites, or viruses (Krämer et al., 2009). The process of infection comprises the change from an outer environment, to another one inside of the host, where the conditions are hostile due to the presence of immune systems. In some animal hosts, the recognition of the pathogen could unchain a series of events leading to an inflammatory response, which represent a plethora of environmental stresses for the bacteria.
Hosts have developed sophisticated mechanisms to sense invaders, and to react against them. Diving at a very deep molecular level one of the first barriers of defense to trigger innate immune responses, is the recognition of pathogen-associated molecular patterns (PAMPs) that will further activate toll-like receptors (TLRs) pathways (Ausubel, 2005; Moresco et al., 2011). Bacteria and archaea have developed during evolution primitive adaptive mechanisms to identify “pathogens” by identification and restriction of foreign genetic material through CRISPR-CAS systems (Hille et al., 2018; Ratner et al., 2019).
The key for the infection success is a quick response and efficient adaptation to a changing hostile environment within the host (Sauder and Kendall, 2018), the translation of cues from the extracellular domain into triggering a set of instructions aimed for the survival of the pathogen. Different molecules play a role in the transference of information, for example during the formation of biofilms, sub-inhibitory concentrations of antibiotics have been proposed to serve as carriers of information between bacteria (Romero et al., 2011). Among different molecules, RNA has achieved a very important role as an information mediator, because it is the molecular link between genome (DNA) and phenotype (proteins or metabolites). It can be quickly recruited during biotic or abiotic stresses. The average life of RNA molecules is short, because the response has been tuned to serve for the synthesis of proteins and to be degraded once they are not needed anymore (Gilbertson et al., 2018).
Small regulatory RNAs (sRNAs) are a subset of RNA molecules that are involved in several mechanisms that aid the pathogen in adaptation, counterfeiting, or suppressing the host immune system (González Plaza, 2018), side to side to other molecules participating in this complex process. The sRNAs are transcribed from the genome, but do not follow the canonical path toward protein translation (Waters and Storz, 2009).
One reason for the involvement sRNAs at infection processes, is the flexibility to target a number of genes or transcription factors, leading to continuous ranges of expression and responses to fluctuating environmental stresses, instead of an abrupt triggering or shutting down of the expression. In that regard, Silva and collaborators have shown that SraL sRNA is responsible for regulating the expression of the transcription termination protein Rho (Silva et al., 2019). This protein is essential for the transcription balance in Bacillus subtilis, and its impairment affects negatively cell motility, biofilm formation, and sporulation (Bidnenko et al., 2017). Thus, the case of SraL and Rho illustrates a complex system of “regulation of regulators.” However, it could be argued that many proteins can carry out similar regulatory roles. Among other reasons, it is probably the faster response of RNAs what has given them a key role during infection, because they do not require translation and it represents a lower energy consumption for the cell. Modulation of the transcriptomic levels allows for a faster response to environmental changes (Sheehan and Caswell, 2018), because it can help to correct or modulate the mRNA levels of many genes before the protein is translated, and ultimately modulate the phenotype according to external fluctuations.
The current review article presents the advances in the field regarding the involvement of sRNAs during bacterial infections, highlighting the latest contributions in the 2 years since my previous review (González Plaza, 2018). The field has expanded broadly, and the number of contributions points toward a future increase in the number of research efforts. The current review aims as well to broaden the scope from diseases affecting humans to other species. It will first cover the type of existing sRNAs and their mode of action, the molecular behavior of sRNAs from the pathogen perspective, and modulation of molecular processes when facing host immune systems. Lastly, different type of sRNAs during several infection processes in a number of species, ranging from plants to animals.
Types of sRnas
The subset of small regulatory RNAs, termed in literature as sRNAs, are a group of primarily non-coding RNA forms (Waters and Storz, 2009) often ranging from 20 to 200 nucleotides (nt) in length, even reaching up to 500 nt (Carrier et al., 2018a). They carry key roles regulating expression levels in a wide range of prokaryotic or eukaryotic genes (Waters and Storz, 2009; Brant and Budak, 2018; Carrier et al., 2018b). Their targets include important genes that are relevant either for the infection process or for the defense of the host organism (Guo et al., 2019). A notable feature is their reported participation in trans-kingdom communication (Benbow et al., 2018; Zeng et al., 2019; Bermúdez-Barrientos et al., 2020). Attending to their origin, they can be divided in prokaryotic and eukaryotic sRNAs.
Prokaryotic sRNAs
Prokaryotic regulatory RNAs can be classified in three main groups: (i) elements present in the 5′ untranslated regions (UTR), (ii) those acting in cis and termed anti-sense RNAs (Lejars et al., 2019), and (iii) those acting in trans that are expressed from other genomic regions than their targets (Chakravarty and Massé, 2019). The third type can be originated at intergenic regions, but also at 5′ or 3′ UTR regions, and are usually termed as sRNAs (Carrier et al., 2018a; Chakravarty and Massé, 2019). The trans-acting sRNAs are the main focus of the current article. The regulation occurs according to several mechanisms, which have been thoroughly reviewed by Carrier, Lalaouna, and Massé (Carrier et al., 2018a). Briefly:
(i) Binding to a regulatory protein. For example binding of sRNAs to the regulatory CsrA protein, that cannot occlude the Shine Dalgarno (SD) sequence of its target (Romeo and Babitzke, 2018; Figure 1A).
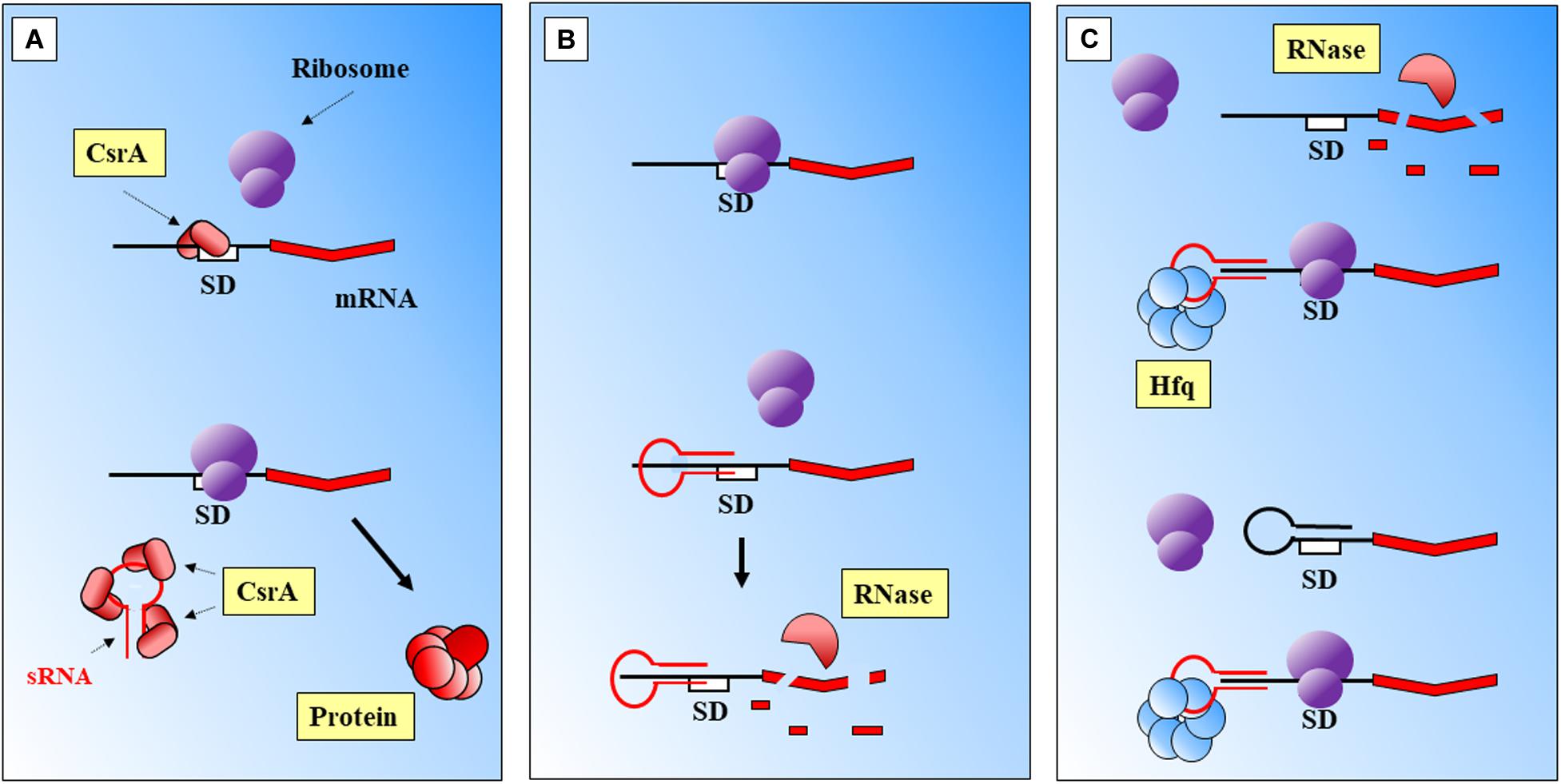
Figure 1. Main modes of action of prokaryotic sRNAs. (A) sRNA binding to a CsrA regulatory protein frees the SD region. (B) Direct sRNA-mRNA binding. The shown sRNA competes with the ribosome, which cannot bind to the SD site. The mRNA is further degraded. (C) Protein mediation. Hfq-sRNA bind to the mRNA and stabilize it, and degradation is prevented. The complex protein-sRNA can unfold secondary structures in the 5′ UTR which will prevent ribosome binding. SD, Shine-Dalgarno. Black thick arrows indicate flow of a biological process, e.g., protein translation. References: Massé et al., 2003; Waters and Storz, 2009; Faucher and Shuman, 2011; Chakravarty and Massé, 2019.
(ii) Direct sRNA-mRNA interaction (Figure 1B). Trans acting sRNAs bind to their targets with partial complementarity (Caldelari et al., 2013). There are different types of interaction, for instance the destabilization of the mRNA by pairing to upstream locations from RBS (Ross et al., 2019) or by interference with the 5′ UTR (Rübsam et al., 2018). Binding of the sRNA can also mask the ribosomal binding site (RBS), consequently the ribosome cannot bind and the translation of the gene is attenuated (Kiekens et al., 2018). The complex mRNA-sRNA can be subject of degradation by RNaseE (Lalaouna et al., 2013).
(iii) Protein mediation (Figure 1C). RNA-binding proteins (RBP) can mediate the regulatory activities of sRNAs, such as Hfq (Beisel and Storz, 2010) or ProQ (Smirnov et al., 2016). The Hfq chaperone mediates interactions between sRNAs and their targets helping to improve base-pair recognition (Massé et al., 2003; Lenz et al., 2004; Holmqvist and Vogel, 2018; Santiago-Frangos et al., 2019). This protein binds to the regulatory RNAs, helps to stabilize them, and leads the base pairing with the targets (Hu et al., 2018; Han et al., 2019). Hoekzema et al. (2019) have suggested an additional mechanism, where the action of Hfq will unfold a hairpin in the targeted mRNA. This activity would create a temporary local structure that facilitates the access of the sRNA. There are other microorganisms where Hfq protein is lacking and FinO, ProQ or RocC carry out its broad regulatory roles: sRNA protection from degradation, alteration of RNA structures to facilitate annealing, stabilization of the sRNA-mRNA complex, or modulation of ribosome binding after the complex is formed, and regulation of RNA degradation (Olejniczak and Storz, 2017).
Eukaryotic sRNAs
In eukaryotes the description of the first types of sRNAs dates back to 1990 with studies reporting silencing of gene expression in petunia plants (Napoli et al., 1990; van der Krol et al., 1990), and later in 1993 in the nematode Caenorhabditis elegans (Lee et al., 1993; Wightman et al., 1993).
The main types of described sRNAs range from 20 to 30 nt, and carry out silencing functions by mediation of Argonaute family proteins (Kim et al., 2009). These sRNAs were firstly divided in three classes: microRNAs (miRNAs), small interfering RNAs (siRNAs), and PIWI-interacting RNAs (piRNAs) (Grimson et al., 2008; Okamura and Lai, 2008; Islam et al., 2018). Long-non-coding RNAs (lncRNAs) are a new group of described regulatory molecules over 200 nt (Agliano et al., 2019).
In the case of miRNAs it has been reported that they can control up to 60% of the human transcriptome, therefore, their involvement in the response to infectious diseases is not surprising (Aguilar et al., 2019).
sRNAs Regulate Key Processes for the Establishment of Infection
Establishment of infection requires a prompt adaptation effort from a pathogenic perspective, in order to proliferate within the host. There are different type of environmental challenges faced by bacteria when entering the host, including different immune barriers to infection (Chakravarty and Massé, 2019). Regulation of transcription aids to adapt quickly to the newly encountered hostile conditions: changes in nutrient availability, pH, temperature, or presence of antimicrobials among other variables. Together, those force bacteria to behave differently during the infective process in comparison with the free form state. It is during these conditions when the diverse toolkit of RNA regulatory activities greatly help for survival. An overview of the different stresses that bacteria must adapt to when entering a host, and the role of sRNAs, have been summarized in Table 1.
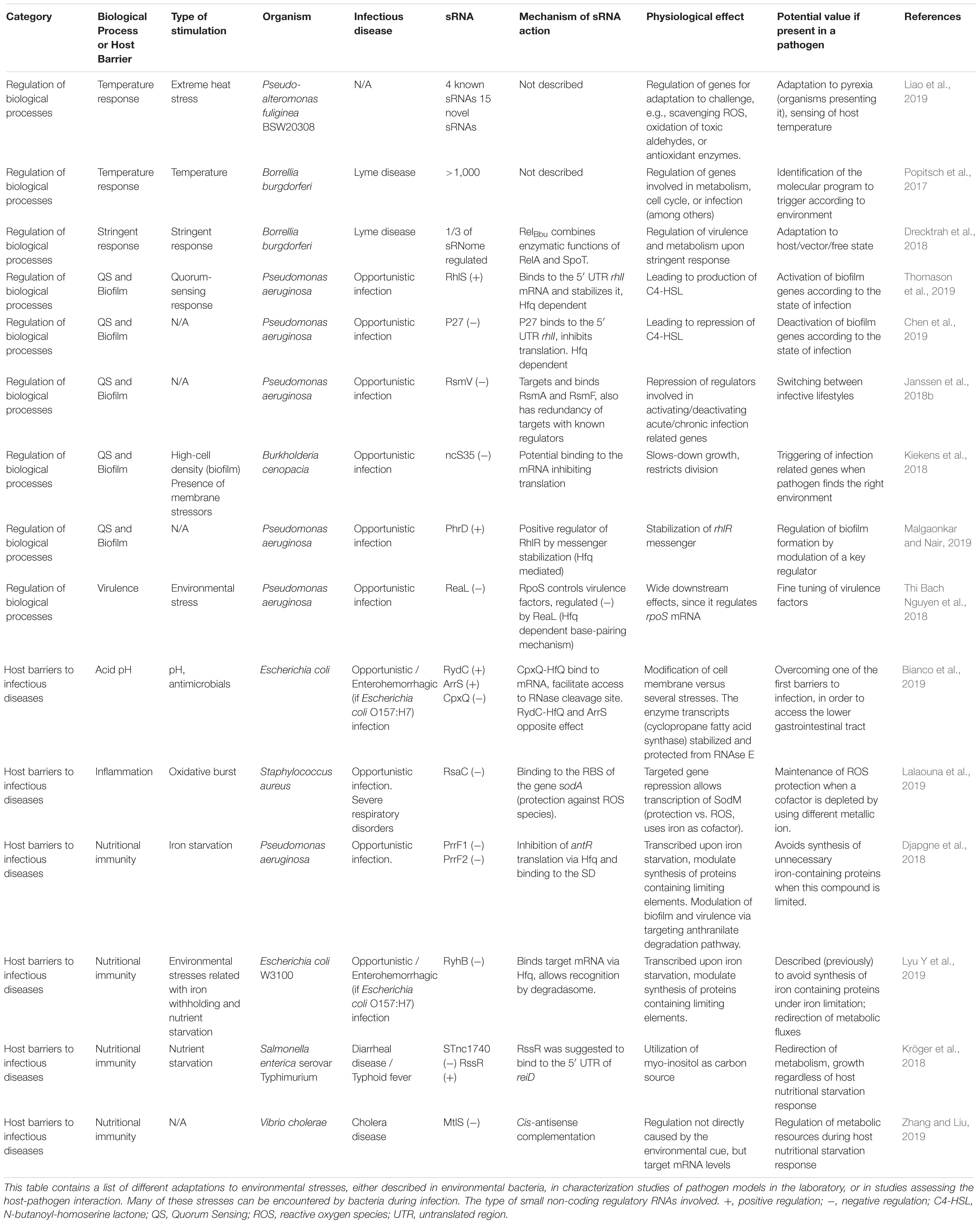
Table 1. Adaptation to environmental stresses mediated by sRNAs.
The range of adaptations mediated by sRNAs have been grouped in two main related categories: (i) regulation of key bacterial processes for the success of infection, and (ii) regulation of responses against host barriers to infection. Most of them have been described in bacteria causing diseases concerning animals, especially in mammals and mostly humans.
Regulation of Biological Processes
Key biological processes set the basis for the adaptation to the host molecular environment, including important responses such as: temperature sensing, stringent microbial response, biofilm formation and Quorum Sensing (QS), or regulation of virulence.
Temperature Response
One of the first cues helping the pathogen to perceive when it has entered the host is the change in temperature. Besides, it cannot be overlooked that a characteristic of infectious diseases is the hyperthermia response, which elevate the body temperature during inflammation (Kluger et al., 1998; Casadevall, 2016). While it is difficult to ascertain, it has been appointed that fever has beneficial effects for the protection against pathogens (Mackowiak, 1981; Casadevall, 2016), since such a high metabolic cost in higher vertebrates would have been lost during evolution if it did not present an advantage (Ostberg et al., 2000).
From a pathogen perspective, RNA presents advantages, as it has been shown to be a relevant molecular thermometer capable of controlling expression of heat shock and virulence genes (Narberhaus, 2010; Loh et al., 2018), when increasing temperatures melt secondary structures and allow access to the ribosome binding site (RBS) (Narberhaus et al., 2006). Through temperature monitorization pathogens can differentiate between free state, insect vector (if present), or hosts with regulated body temperature (González Plaza et al., 2016; Álvarez-Estrada et al., 2018), which could lead as well to develop a specific program to respond to fever (in hosts where this mechanism is present).
A recent study has evaluated the transcriptomic response of the psychrotrophic bacterium Pseudoalteromonas fuliginea BSW20308, which is adapted to Arctic environmental conditions (Liao et al., 2019). The aim was to evaluate the impact of global warming over the ecologically dominant genus, where temperature increases may trigger regulation mediated by sRNA. Authors described the whole sRNome (repertoire of sRNAs) when this microorganism grew at different temperatures, from very low ones resembling its natural environmental conditions, to higher temperatures of a global warming scenario. Results, according to authors, indicated an intense involvement of sRNAs in temperature adaptation. A 316 nt novel sRNA, termed Pf1, showed to be correlated with the expression to a wide group of 644 genes mostly annotated in the categories of catabolism, energy, translation, and intracellular transport. As previously mentioned, sRNAs have an ample range of effects over the transcriptome, helping bacteria to regulate their physiology upon environmental perturbations. Besides, since sRNAs are genetic carriers of information not translated into proteins, they can perform their regulatory roles in a faster fashion than other important regulators as heat-shock proteins. Although this characterization relies on an RNA-seq approach and further in silico data analyses, it is not surprising such a broad regulatory network as suggested in the article. Either by conservation or convergent evolution as appointed by Narberhaus (2010), this regulatory response is present in many pathogens for adaptation to the host conditions, and may be an important variable to consider during pyrexia (Mackowiak, 1981; Kluger et al., 1998; Ostberg et al., 2000; Casadevall, 2016; González Plaza et al., 2016).
In their analysis of the differential expression between infective and environmental temperatures in Borrelia burgdorferi, Popitsch et al. (2017) report a large set of sRNAs with differential expression between both conditions, and also reveal a variety of transcription origins. A brilliant conclusion of this study is that in overall, the mode of action of sRNAs could deeply impact the way we perform genetic studies. Deletion of a gene could erase as well an important regulator for a number of downstream targets. This can undoubtedly represent a confounding factor for the interpretation of results on loss-of-function phenotypes due to the interference of sRNA regulation.
What becomes clear is that monitorization of temperature changes is a central event in the life cycle of bacterial pathogens (Papenfort and Vogel, 2010), and that regulation follows an intricate pathway with several levels. In a fascinating example of interaction between kingdoms and pathogen manipulation, the spirochaetal outer surface (lipo)protein (Osp) C (OspC) from Borrelia burgdorferi binds to the SALP15 salivary protein belonging to the tick vector Ixodes scapularis. The pathogen uses the insect protein to succeed in the transmission to the mammalian host and its infection (Ramamoorthi et al., 2005). The expression of ospC is influenced by RpoS (Hübner et al., 2001), which back was suggested by Lybecker and Samuels (2007) to be regulated by a sRNA in response to temperature (Figure 2). RpoS is a key global regulator controlling virulence or response to general stresses in several pathogens (Fang et al., 1992; Suh et al., 1999; Dong and Schellhorn, 2010; Battesti et al., 2011). The regulation of temperature responses mediated by RpoS, is modulated by the sRNA DsrABb in B. burgdorferi, which is expressed upon increase in temperature (Lybecker and Samuels, 2007). Few years later, it was experimentally confirmed to occur through Hfq mediation (Lybecker et al., 2010; Figure 2).
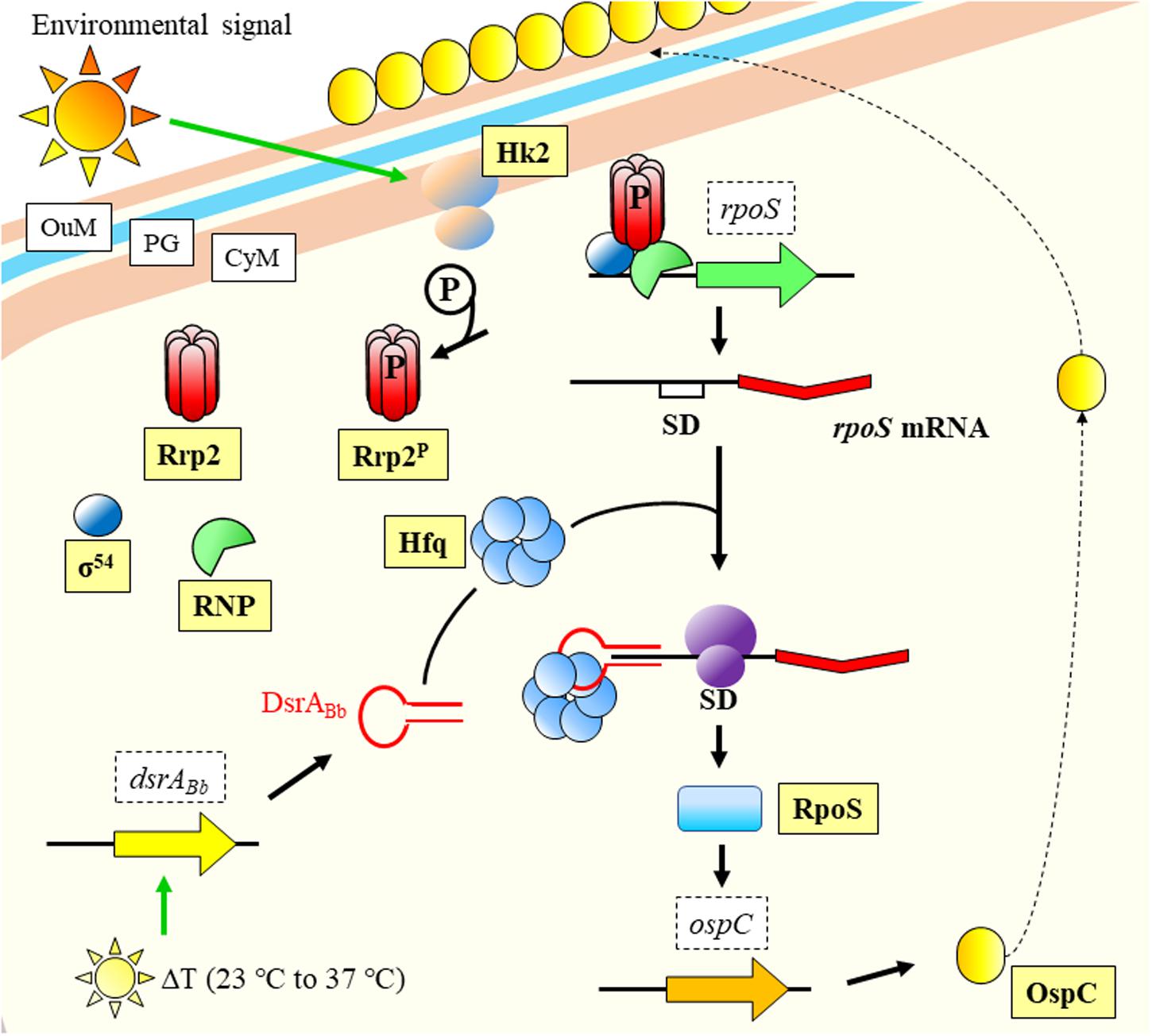
Figure 2. Temperature response model of Borrelia burgdorferi. An environmental signal activates Hk2 membrane protein that phosphorylates and activates Rrp2. Together with RNP and σ54, they facilitate transcription of rpoS. The mRNA is stabilized by the joint action of DsrABb (expressed upon temperature increase to host conditions) and Hfq, and the messenger is translated into protein. RpoS regulates the transcription of ospC, which product is displayed on the OuM. CyM, Cytoplasmic membrane; OuM, Outer membrane; PG, peptidoglycan; RNP, RNA Polymerase; SD, Shine-Dalgarno. Black thick arrows indicate flow of a biological process, e.g., protein translation. Green thick arrows indicate activation. References: Burtnick et al., 2007; Lybecker and Samuels, 2007; Lybecker et al., 2010; Radolf et al., 2012; Steere et al., 2016.
Stringent Response
Stringent response can be defined as the set of bacterial conserved mechanisms activated during nutritional environmental stresses (Poole, 2012; Drecktrah et al., 2018). It produces a general decrease on the expression of genes related with growth, involving synthesis of proteins or nucleic acids, and enhanced transcriptional levels of genes for survival (Chatterji and Ojha, 2001; Poole, 2012; Irving and Corrigan, 2018). When the bacterial stringent response is unleashed, mediation of enzymes such as RelA or SpoT lead to global transcriptomic changes (Atkinson et al., 2011; Shyp et al., 2012). A different version of RelA in Borrellia burgdorferi, RelBbu, has regulatory capabilities on a third of the sRNAs identified in this bacteria to date (Drecktrah et al., 2018). This pathogen has a life cycle that includes a vertebrate and an invertebrate host, thus, adaptation to different environments in order to regulate the behavior in such disparate conditions represent a challenge for survival. Not surprisingly Drecktrah et al. (2018) found that most of the targets of the regulated sRNAs are involved in two biological processes required during conditions of infection, virulence and metabolism. Interestingly, the sRNAs targets of RelBbu are not exclusively found within the chromosome, but could be found as well in plasmids.
Biofilm Formation and Quorum Sensing
Biofilms are bacterial community structures that provide additional protection against the host immune system, e.g., the effect of cytokines (Leid et al., 2005). Biofilm formation requires coordination of QS mechanisms of communication, which are mediated mainly by N-acyl homoserine lactones (AHLs) among other molecules and mainly controlled by the rhlR-rhlI and lasR-lasI signaling systems (Davies et al., 1998; Lee and Zhang, 2015; Figure 3). The AHL N-butanoyl-homoserine lactone (C4-HSL) is an important molecule in the QS response of Pseudomonas aeruginosa. This molecule binds to RhlR and the activated complex regulates positively the expression of rhlI, and its translation to RhlI that synthesizes C4-HSL. The synthesis of RhlR has been positively related with a 74-nucleotide sRNA, PhrD (Malgaonkar and Nair, 2019; Figure 3). The interaction between the regulator molecule and the mRNA target was predicted in silico and further demonstrated in experimental conditions (which mimicked the host during pathogenesis). While this system seems to act independently of any P. aeruginosa proteins (authors expressed it heterologously in E. coli), the system achieves better expression levels in the P. aeruginosa background, probably assisted by native proteins.
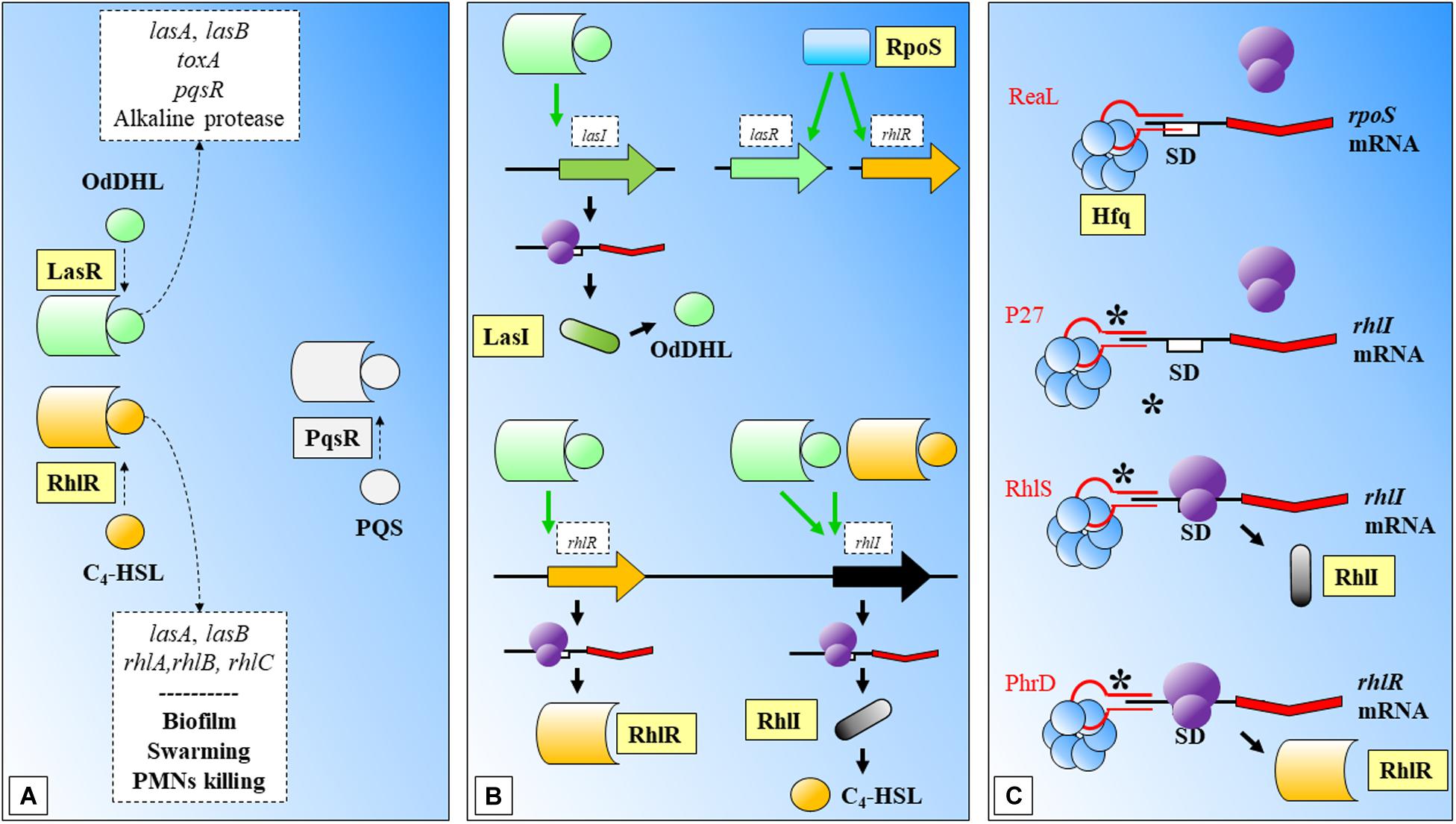
Figure 3. Biofilm formation and Quorum Sensing in Pseudomonas aeruginosa. (A) LasR and RhlR are two key regulatory molecules that need OdDHL and C4-HSL participation. LasR-OdDHL regulate the genes included in the upper box. RhlR-(C4-HSL) regulate genes or affect processes indicated in the lower box. PqrS is another important regulator, it has been shaded as it does not participate in the shown processes. (B) LasR and RhlR expression is dependent on RpoS. LasR-OdDHL regulates positively: the expression of lasI, and ultimately the synthesis of OdDHL; the expression of rhlR; and rhlI. RhlR-(C4-HSL) regulates positively the expression of rhlI. (C) Mechanisms of action of several sRNAs involved in regulation of the synthesis of regulatory proteins within these signaling systems: ReaL affects negatively the expression of rpoS; P27 is another negative regulator, in this case of rhlI; RhlS regulates positively rhlI; PhrD is a positive regulator of rhlR. C4-HSL: N-butanoyl-homoserine lactone; OdDHL: N-(3-oxododecanoyl)-L-homoserine lactone; PQS: Pseudomonas quinolone signal; SD: Shine-Dalgarno. Black thick arrows indicate flow of a biological process, e.g., protein translation. Green thick arrows indicate activation. Dashed-line text boxes: indicate downstream effects of the two signaling systems, phenotypes are indicated in bold (e.g., swarming). ∗: indicates that this mechanism of sRNA regulation has been proposed. References: Schuster et al., 2004; Nadal Jimenez et al., 2012; Brouwer et al., 2014; Pita et al., 2018; Thi Bach Nguyen et al., 2018; Chen et al., 2019; Malgaonkar and Nair, 2019.
The involvement of sRNAs in the QS response of P. aeruginosa was investigated with mutants of AHL synthesis by Thomason et al. (2019), (Figure 3). Authors found a group of sRNAs responsive to AHLs treatment, where RhlS (previously SPA0104) (Ferrara et al., 2012) showed the highest accumulation at inductive conditions. This regulator acts positively over the translation of rhlI, leading ultimately to the production of C4-HSL. P27 is another sRNA that regulates negatively the translation of rhlI bindings to the 5′ UTR aided by Hfq (Chen et al., 2019; Figure 3).
The environmental bacteria Burkholderia cenocepacia belongs to the Burkholderia cepacia complex (Bcc) and can become an opportunistic pathogen in plants (Mahenthiralingam et al., 2005), but also in patients affected by cystic fibrosis (Drevinek and Mahenthiralingam, 2010). The sRNA ncS35 is involved in growth regulation by predicted binding to mRNA targets, probably facilitating the degradation of the transcript (Kiekens et al., 2018). Authors used a deletion mutant (ΔncS35) in comparison with the wild type (WT) strain, and a complemented ΔncS35 overexpressing the sRNA under inductive conditions. The mutant phenotype displayed biofilms with larger aggregates, increased optical density or metabolic activity in comparison with the WT or the complemented mutant. Differential gene expression between the mutant and the WT showed upregulation of genes involved in metabolism both in exponential and stationary growth phases. This indicates a negative regulatory role of ncS35 over bacterial growth. Additionally, authors observed higher expression values for ncS35 when bacteria form biofilms than in free planktonic culture. Additional increases in transcription occurred during nutrient limitation after cultivation in M9 minimal media or in the presence SDS, a known membrane stressor (Flahaut et al., 1996). In overall, the effect of this sRNA is to slow-down growth. The higher expression during presence of stressors, may indicate its involvement in protection of the bacteria by restricting division when environmental conditions are detrimental for the pathogen.
Virulence
Bacterial virulence can be defined as the “relative capacity to overcome available defenses” (Sparling, 1983), or “the relative capacity of a microorganism to cause damage in a host” (Casadevall and Pirofski, 2003). This capability is mediated by virulence genes, which have to fulfill three requirements: (i) active in the interaction between pathogen and host, (ii) direct determinants of the pathogen damage, and (iii) the lack of those virulence genes in non-pathogenic strains (Wassenaar and Gaastra, 2001). Some authors also use the term “virulence factor” instead of “virulence gene” (Diard and Hardt, 2017).
The role of sRNAs over virulence has been well-characterized in Pseudomonas aeruginosa. In this organism, an important system for regulation of virulence is the carbon store regulator (Csr) or repressor of stationary-phase metabolites (Rsm), being the main regulatory protein CsrA (or RsmA) (Figure 4). This system can also control other important features of the interaction with the host, such as biofilm formation, carbon metabolism, or stress responses (Romeo and Babitzke, 2018). Central to this system is the Gac/Rsm pathway, where the GacS/GacA two component system has a fundamental role (Coggan and Wolfgang, 2012; Figure 4). GacA induces the expression of the sRNAs rsmY and rsmZ, which can bind to the central regulatory protein RsmA (CsrA) to block its regulatory functions (Kay et al., 2006; Figure 4). RsmW is another sRNA that can bind to the regulatory protein (Miller et al., 2016; Valentini et al., 2018). This sRNA mediation can allow cells to respond precisely to environmental challenges, and aid transition from different infective phenotypes in P. aeruginosa. Additionally, RsmF/RsmN (a CsrA family protein), have overlapping functions to RsmA (Marden et al., 2013; Romero et al., 2018). Janssen et al. (2018a) have identified RsmV, a new 192-nt small non-coding RNA that has binding activity to RsmA and RsmF. In vitro electrophoretic assays confirmed that both proteins bind RsmV probe with high affinity. This interaction was supported by complementation studies using a two-plasmid reporter system in a mutant with high levels of RsmA/RsmF (lacking rsmV, rsmY, and rsmZ). Complementation with a plasmid expressing RsmV antagonized the activity of RsmA/RsmF (Figure 4). The displayed redundancy of targets with previously known sRNAs illustrates the fine-tuned coordination of sRNA regulation.
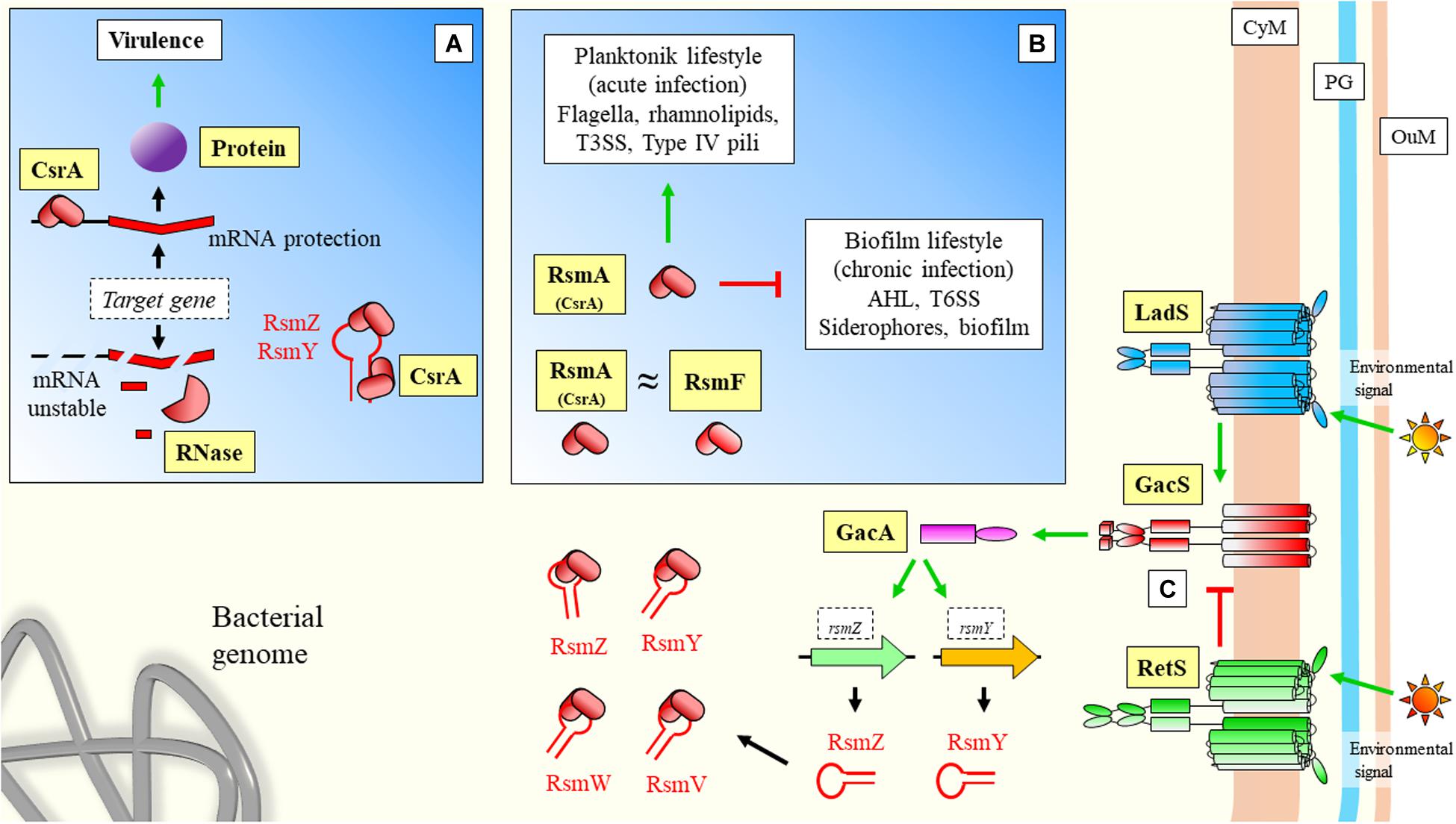
Figure 4. Virulence sRNA mediated regulation in Pseudomonas aeruginosa: Role of CsrA regulatory protein and sequestration by sRNAs. (A) CsrA has been described to have also a positive effect on target mRNAs by protecting transcripts from degradation. (B) Regulatory effects of CsrA. In the case of positive CsrA regulation of targets, when sRNAs bind and sequester this protein, they force the instability and degradation of the messenger (A). In the case of CsrA negative regulation of targets (Figure 1A), sRNAs can prevent binding of the regulatory protein to the mRNA allowing translation (where sRNAs have a positive effect over mRNA targets). (C) GacS/GacA two component system is either activated by LadS, or inhibited by RetS. These two transmembrane proteins mediate between GacS and environmental stimulation. Upon activaction, Gac promotes the transcription of sRNAs, which can further bind CsrA/RsmA and modify expression of downstream genes. CyM, Cytoplasmic membrane; PG, peptidoglycan; OuM, Outer membrane. Black thick arrows indicate flow of a biological process, e.g., protein translation. Green thick arrows indicate activation. Red thick arrows with flat cap indicate inhibition. Text boxes: indicate downstream effects of RsmA. References: Wei et al., 2001; Records and Gross, 2010; Sonnleitner et al., 2011; Coggan and Wolfgang, 2012; Nadal Jimenez et al., 2012; Marden et al., 2013; Yakhnin et al., 2013; Chambonnier et al., 2016; Janssen et al., 2018a; Romeo and Babitzke, 2018; Valentini et al., 2018.
The sigma factor RpoS controls as well a wide number of virulence related genes in Pseudomonas aeruginosa under environmental stresses. RpoS translation has been shown to be negatively regulated by the sRNA ReaL, through a Hfq dependent base pairing mechanism (Thi Bach Nguyen et al., 2018; Figure 3). As authors point out, this was the first case of a negative sRNA transcriptional regulator of rpoS.
Host Barriers to Infectious Diseases: sRNA-Mediated Bacterial Adaptation to a Hostile Environment
The second category groups those responses from the pathogen to known barriers of infection, such as acidic pH, inflammation, or nutritional immunity.
Acidic pH: One of the First Barriers to Infection
The acidic pH of the stomach represents one of the first barriers to infection set by the host. Microbial parasites can overcome it through different mechanisms, including the degradation of urea into CO2 and NH3 (Burne and Chen, 2000). This strategy increases the survival chances of enteropathogenic organisms, such as Yersinia pseudotuberculosis, a Gram-negative food-borne bacterial pathogen (Hu et al., 2010). Another mechanism involves modifying the composition of the cell membrane to withstand sudden exposure to acid pH, by incorporation of different membrane proteins, or the enzymatic modification of the pre-existing fatty acids already in the membrane (Bianco et al., 2019). Cell membrane composition changes have key roles during infective processes, as it determines the fluidity of toxic compounds or antimicrobials. The enzyme cyclopropane fatty acid synthase mediates the incorporation of a methylene group into unsaturated fatty acids. The enzyme transcripts were shown to be stabilized and protected from the degradative activity of RNAse E in Escherichia coli, by two sRNAs, RydC, and ArrS; while a third one, CpxQ, had a repressive role (Bianco et al., 2019). While RydC and ArrS mask the mRNA cleavage site that is not available to RNase E, CpxQ increases the accessibility to the same site.
Inflammation: Oxidative Stresses
When the host detect the presence of a pathogen, one of the characteristic responses is inflammation and the subsequent presence of oxidative stresses for the bacteria (Carlos et al., 2018). These stresses are common for extremophiles, such as the haloarchaeon Haloferax volcanii. A recent study reports the presence of hundreds of sRNAs in response to oxidative stress caused by hydrogen peroxide in this species (Gelsinger and DiRuggiero, 2018). Despite the potential evolutionary lineage distance between archaea and bacteria, this study shows as well how small regulatory RNAs can play a key role for regulation of biological processes in extreme conditions. In the case of infectious diseases, the extreme transient conditions relate to the stress caused by the host during oxidative burst, which induce heavily the expression of RsaC sRNA in Staphylococcus aureus (Lalaouna et al., 2019). The sRNA binds to the start codon region of the sodA mRNA, involved in protection against reactive oxygen species (ROS). The repression of the targeted gene allows the transcription of a second enzyme, SodM, involved in ROS protection but using iron as cofactor, instead of manganese (limited due to the nutritional immunity).
Nutritional Immunity
Besides oxidative stress (although related), pathogen recognition triggers other quick immune responses from the host, aimed to clear the bacteria by restriction of the available metabolic resources. The term “nutritional immunity” (Damo et al., 2013) refers to the limitation by the host of essential elements for the development of the pathogen. This strategy includes targeting of iron, manganese, or glucose (Carlos et al., 2018). Undoubtedly, the decrease in nutrients cellular levels has a direct relationship with the triggering of stringent mechanisms in bacteria. Regarding glucose metabolism, a characteristic behavior of human patients during infective diseases are patterns of transient anorexia that lead to a lower energetic intake, thus, limiting the availability of nutrients. This is a probable evolutionary response from the host, in order to create a metabolically stressful environment for the pathogen. Given that these host-pathogen relationships have developed through evolution, additional layers of regulation must have appeared in key microbial processes, in order to ensure survival of the bacterial strains, where sRNAs play a fundamental role.
In the case of manganese, the host immune system can limit its extracellular levels (Diaz-Ochoa et al., 2014), causing an impairment in the oxidative stress protection machinery from the pathogen, while increasing the oxidative burst. In that regard, the previous example of RsaC, can help S. aureus to avoid the synthesis of a non-functional enzyme (SodA) that requires manganese, and synthesize the second enzyme (SodM) that uses available iron as cofactor restoring the oxidative protection.
Besides targeting manganese, iron starvation is another innate immunity strategy of vertebrates, when detrimental bacteria are identified: the so-called “iron withholding strategy” (Ong et al., 2006). From a broader perspective, mechanisms to deal with iron deprivation have been described in environmental bacteria. For instance, the sRNA iron-stress activated RNA 1 (IsaR1) mediates the acclimation to conditions of iron starvation and high salinity in cyanobacteria (Rübsam et al., 2018). Even an additional role has been reported in the regulation of osmotic response. IsaR1 down-regulates the expression of the gene ggpS, encoding for the enzyme GG-phosphate synthase which is involved in the accumulation of heteroside glucosylglycerol (GG), and the adaptation to high saline concentrations. IsaR1 interferes with the 5′UTR of its target gene, ggpS. As it is becoming clear in the sRNAs research field, the ggpS regulation strategy does not follow an abrupt “all or nothing” scheme. It rather serves to integrate a wide range of environmental fluctuations in a continuous manner.
The previous model of cyanobacterial regulation in response to low iron, is relevant as well during infection of P. aeruginosa. This bacterium is able to detect environmental iron fluctuations, which drive to the expression of virulence genes in response to the host primary line of defense. Two of its sRNAs, PrrF1, and PrrF2, react to iron (Fur mediated) and have been shown to repress anthranilate metabolism (Djapgne et al., 2018; Figure 5). These two sRNAs have been described as functional homologs of the RyhB in E. coli (Wilderman et al., 2004). The regulatory effect is indirect, as they inhibit the translation of a transcriptional activator, antR, with downstream effects over genes for degradation of anthranilate. The metabolic pathway from anthranilate degradation ends with the synthesis of Pseudomonas quinolone signal (PQS), relevant for Quorum Sensing (QS) (Brouwer et al., 2014; Figure 5). RyhB, was previously described as a negative regulator of genes involved in the control of iron levels within the cell (Massé and Gottesman, 2002). Besides the regulatory activities toward genes related with iron homeostasis, Massé and Gottesman reported the RyhB mediated regulation of three enzymes of the tricarboxylic-acid (TCA) cycle (succinate dehydrogenase, aconitase, and fumarase) (Figure 6).
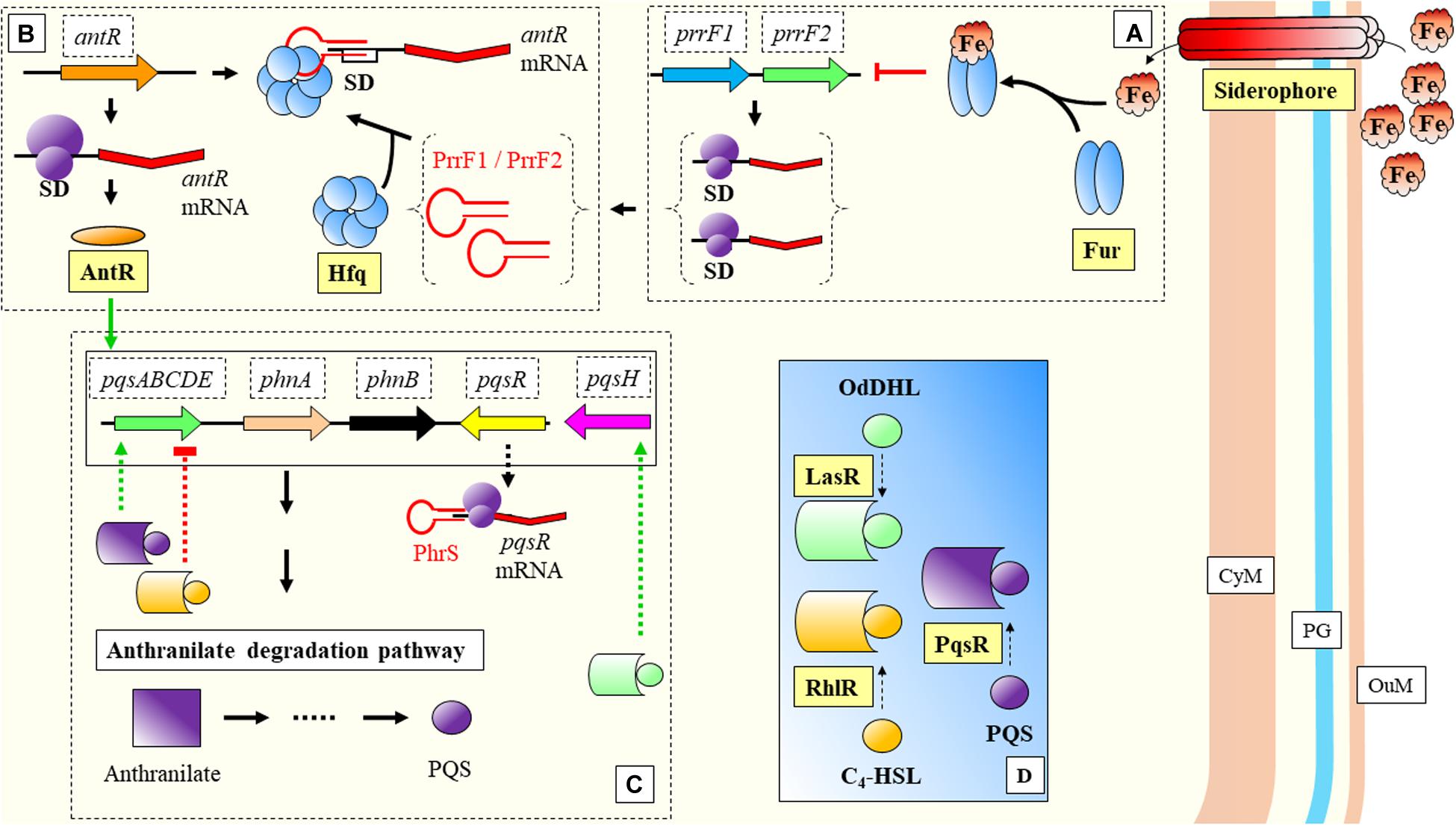
Figure 5. Nutritional immunity response in Pseudomonas aeruginosa. (A) Excess of extracellular iron is bound to Fur protein that inhibits transcription of PrrF1 and PrrF2. (B) PrrF1 and PrfF2 bind to the SD site by mediation of Hfq and block the translation of antR. (C) AntR has a positive effect over anthranilate degradation genes. Another sRNA, PhrS, allows the transcription of pqsR. (D) PqsR is an important regulator, which together with RhlR and LasR have key regulatory effects over genes belonging to the anthranilate degradation pathway (shown in panel C). CyM, Cytoplasmic membrane, OuM, Outer membrane; PG, peptidoglycan. Black thick arrows indicate flow of a biological process, e.g., protein translation. Green thick arrows indicate activation (equal for green thick dashed lines). Red thick arrows with flat cap indicate inhibition (equal for red thick dashed lines). References: Dubern and Diggle, 2008; Brouwer et al., 2014; Baker et al., 2017; Djapgne et al., 2018.
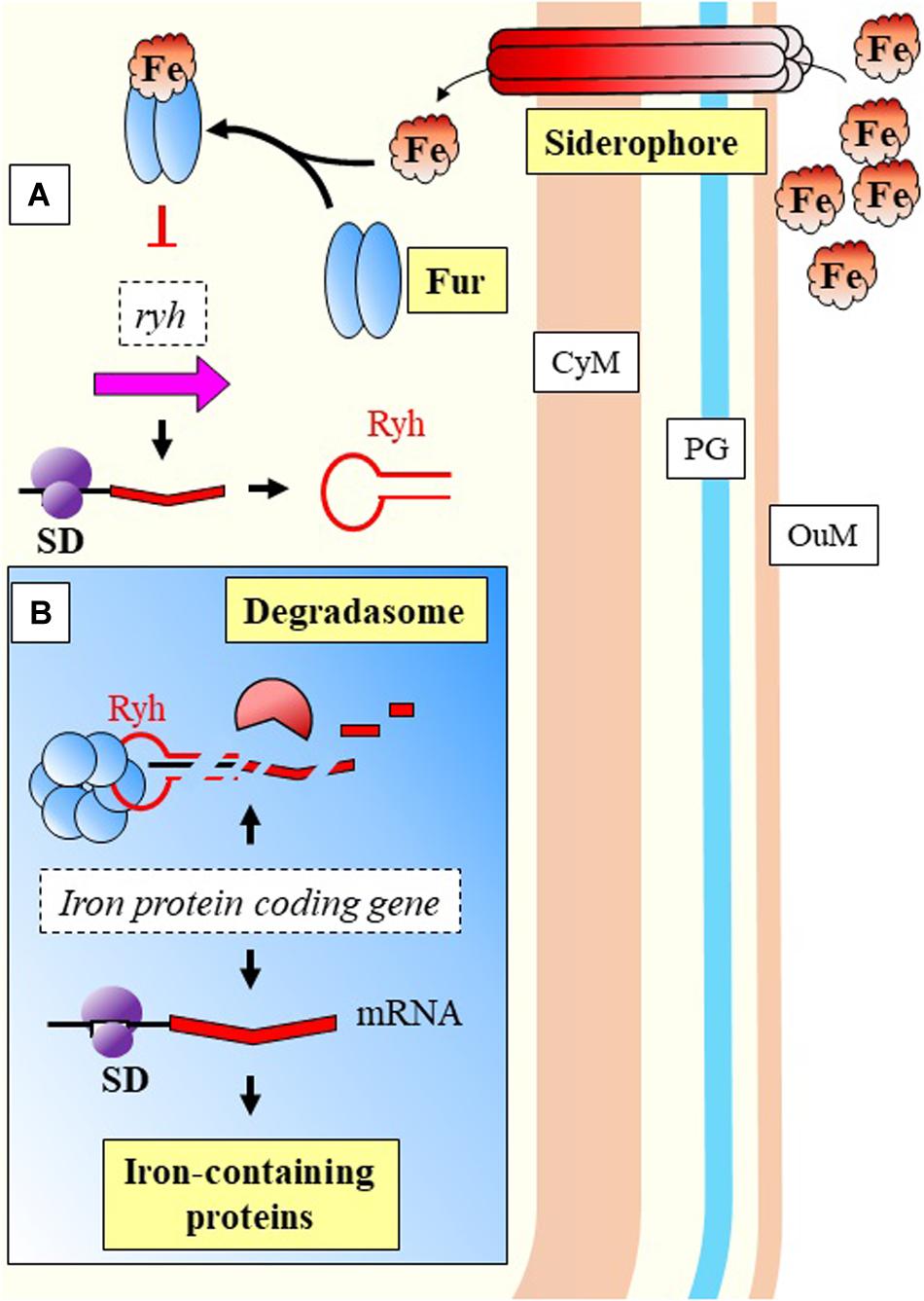
Figure 6. Nutritional immunity response in E. coli. (A) Excess of extracellular iron is bound to Fur protein that inhibits transcription of Ryh. (B) Ryh can inhibit the synthesis of proteins containing iron when this element is limited due to nutritional starvation exerted by the host. Ryh negative regulation is mediated by Hfq, binding to the target mRNAs directing the activity of the degradasome. CyM, Cytoplasmic membrane, OuM, Outer membrane; PG, peptidoglycan. Black thick arrows indicate flow of a biological process, e.g., protein translation. Red thick arrows with flat cap indicate inhibition (equal for red thick dashed lines).
Recently, the contribution of RyhB over the carbon metabolism was quantified (Lyu Y et al., 2019). A mutant was generated by recombination-mediated deletion of RyhB, in comparison with the WT and an inducible mutant. Authors report a redirection of the metabolic flux toward the pentose phosphate pathway. These results support the role of sRNAs in modification of central catabolism in order to adapt to changing environments and allow survival. According to authors, it may be due to the fact that key cellular catabolism enzymes have iron as component of their structure. Several of the enzymes involved in the TCA cycle require iron on their structure (Cornelis et al., 2011). The relationship between carbon and iron levels has been previously addressed in environmental bacteria (Kirchman et al., 2000), and similar mechanisms occur with pathogenic regimes (Andrews et al., 2003). Central carbon metabolism sRNA regulation has been also reported for Escherichia coli, according to environmental conditions (Shimizu, 2013; Lyu Y et al., 2019). That is of especial relevance during nutrient starvation, where alternative metabolic resources must be sought.
Another example of adaptation during nutrient limitation can be found in the Gram-negative bacteria Vibrio cholerae, responsible for cholera disease. A set of sRNAs (CsrB, CsrC, and CsrD) (functional homologs of RsmA protein and RsmB/C/D sRNAs in P. aeruginosa) bind to this protein through a region that resembles the SD of the CsrA mRNA targets (Butz et al., 2019; Figure 7). This bacterium adapts to the clear differences and limitations innutrient content between the human host and the external environment. The mannitol operon is related to the adaptation to aquatic environment and to biofilmformation, and is regulated by the non-coding 120 nt RNA MtlS (Mustachio et al., 2012; Zhang and Liu, 2019). Zhang and Liu (2019) have shown what are the causes determining MtlSregulation. Very interestingly, it is not directly the environmental cue that triggers alterations in the expression level of the sRNA, but the mRNA levels of thetarget gene, mtlA. Another sRNA recently described, CoaR, binds to the mRNA of tcpI to block its translation (Xi et al., 2020; Figure 7). TcpI is a negative regulator of tcpA, a gene encoding the structural major pilin subunit of TCP (Harkey et al., 1994).
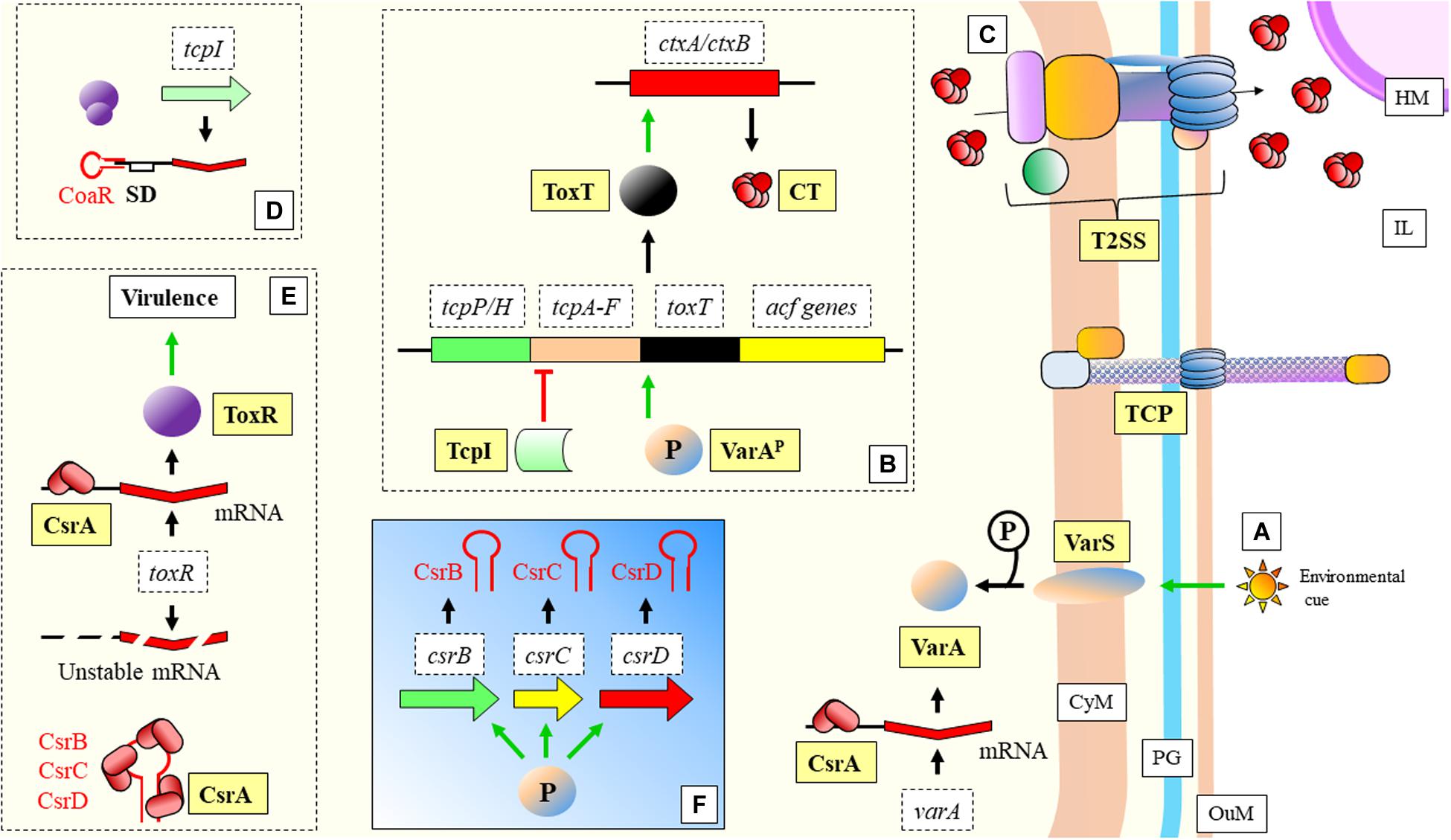
Figure 7. Vibrio cholerae: mechanisms of virulence and regulation. (A) Environmental cues trigger the signalization in the two-component system VarS/VarA (functional homologs to the GacS/GacA two-component system of P. aeruginosa). Phosporilation of VarA activates this protein. Transcription of varA is facilitaded by CsrA. (B) VarAP (activated) acts as a transcriptional activator of toxT. ToxT activates as well ctxA/ctxB encoding the two subunits of the CT protein, which is extruded to the IL through a T2SS (C). The tcpA gene, required for the assembly of the TCP, is located in the same operon as toxT. TcpI is a transcriptional inhibitor of tcpA. TCP structure is necessary for the progression of the disease (bacterial aggregation and microcolony formation). (D) CoaR sRNA blocks binding of the ribosome to the tcpI mRNA. (E) CsrA stabilizes toxR mRNA, which is necessary for positive regulation of virulence genes. (F) VarAP activates transcription of CsrB, CsrC, and CsrC sRNAs, which sequester CsrA, yielding an unstable toxR transcript. CT, Cholera toxin; CyM, Cytoplasmic membrane; HM, Host cell membrane; IL, Gastrointestinal lumen; OuM, Outer membrane; PG, peptidoglycan; T2SS, Type II secretion system; TCP, Toxin coregulated pilus. Black thick arrows indicate flow of a biological process, e.g., protein translation. Green thick arrows indicate activation. Red thick arrows with flat cap indicate inhibition. References: Hammer and Bassler, 2007; Jang et al., 2011; Cobaxin et al., 2014; Mey et al., 2015; Ng et al., 2016; Dorman and Dorman, 2018; Jemielita et al., 2018; Butz et al., 2019; Xi et al., 2020.
Infectious Diseases in Plants
Most of the studies involving plants have been conducted in relevant crop species. Understanding the role of sRNAs has interest for breeders and companies, because advances in regulation of the disease could allow to control bacterial pathogens and prevent economic losses. In a world with continuous population growth, the improvement in agriculture efficiency among other measures could help to avoid shortages in food supply and help to mitigate the carbon footprint of agricultural practices (Beebe et al., 2013). Additionally, higher efficiency means that less agricultural land is required, and bigger efforts and extensions could be directed toward maintaining biodiversity (Trewavas, 2001). One of the most interesting aspects of the war between plants and bacteria, is the sessile nature of plants while bacteria have the advantage of being mobile. This fact has forced the evolutionary development in plants of sophisticated mechanisms of defense (Dangl and Jones, 2001). However, the immune response both from plants and animals, can cause a fitness disadvantage if it is held through time and not regulated. The eukaryotic sRNAs aid in the modulation of that response. But in the case of prokaryotes, sRNAs are relevant to fight back, involved in the regulation of different important pathogenic features during infection, such as secretion systems or mobility.
In this never-ending war biofilms help bacteria to endure different stresses, but plant immune systems have “learned” to target key molecules for its formation. Thus, mechanisms to evade the host recognition are necessary for the survival of the pathogen and establishment of a successful infection. In that regard, Nakatsu et al. (2019) have shown that not all isolates of Pseudomonas syringae produce AHLs. Many of the reported isolates carry mutations in two key genes: the AHL synthase psyI, or the AHL transcription factor psyR. Most probably the production of AHLs could have represented a biological burden for the infective processs. Strikingly, these strains still show responses of two sRNAs that should be related with AHLs, RsmY, and RsmX. Furthermore, their expression is enhanced at high cell densities, suggesting the existence of alternative routes for QS signaling. Although further research is needed, the loss of AHLs and the increased levels of these two sRNAs during situations with high number of cells, may reflect the involvement of these regulatory RNAs in mediating the coordination of a QS response.
In another brilliant event of the antagonism, plants have developed the capability to synthesize compounds that interfere with QS signalization. Rosmarinic acid (RA) was reported to bind with RhlR (instead of C4-HSL), inducing abnormal premature behaviors related with biofilm formation and virulence. When cultures of Pseudomonas aeruginosa PAO1 were challenged with RA, a group of sRNAs showed differential expression in response to the presence of RA, including the induction of RsmY (Fernández et al., 2018). This study highlights the relevance of sRNAs in the communication process between host and pathogen. If the host detects the presence of the bacteria, it can produce certain defensive compounds that will impair the virulence response of the pathogen through modification of important regulators and alteration of sRNA levels.
Plant-pathogen interaction it is not limited to detection and secretion, but it can reach up to a complex alteration of behaviors through genetic manipulation. Hijacking of the host metabolic machinery is not exclusive of viruses, it is a fine strategy of the gram-negative soil phytopathogen Agrobacterium tumefaciens (Gelvin, 2003; Hwang et al., 2017). The evolutionary counter-attack from plants has been the development of mechanisms to disrupt communication among pathogen cells, through the production of γ-aminobutyric acid (GABA) (Sheehan and Caswell, 2018). Agrobacterium tumefaciens and other Rhizobiales have an additional offensive strategy consisting in the production of AbcR1, a sRNA that regulates the plant transporter responsible for importing GABA molecules into the cell and its deleterious effects (Wilms et al., 2011). AbcR1 binds to the SD sequence and decreases the stability of the target mRNA.
A different sRNA, the highly conserved PmaR, can be found in Agrobacterium species and a specific strain of Rhizobium sp. It has been reported to be related to positive regulation of genes involved with peptidoglycan biosynthesis, motility, and virulence according to the study by Borgmann et al. (2018). Additionally, PmaR has a very important role in mediating ampicillin resistance, as observed after studying deletion mutants with impaired survival capabilities in growing concentrations of this antibiotic. The control of the antibiotic resistance gene was proposed by the authors as a means to obtain a biological advantage in the highly competitive rhizosphere environment. PmaR is a positive regulator binding to the 5′UTR, leading to the stabilization of the Shine-Dalgarno region, which would be otherwise prone to form structures preventing ribosome binding or even mRNAs destabilization.
Flowers are sensitive structures of plants, which can be used as an entry point by pathogens. Such an example is Erwinia amylovora, a Gram-negative bacteria responsible for fire blight disease in apple or pear trees belonging to Rosaceae (Oh and Beer, 2005). Once the bacterium enters the host, it spreads using flagella through the vascular system to continue infection. Three sRNAs dependent on Hfq (ArcZ, OmrAB, and RmaA) have been linked to the maintenance of the swimming and motility levels of this bacterium (Zeng and Sundin, 2014). However, the specific molecular regulation mechanisms were not understood. Given that motility is related with flagella, a potential target of regulation mediated by sRNAs is the dual system composed by the proteins FlhD and FlhC regulating the expression of the rest of the genes in the flagellar regulon (Liu and Matsumura, 1994; Frye et al., 2006). Schachterle et al. (2019) researched the involvement of ArcZ, OmrAB, and RmaA in flagellar regulation. First, they found reduced expression levels for regulatory and flagellar structural genes in lack-of-function mutants (ΔarcZ, Δhfq, ΔomrAB, and ΔrmaA) in comparison with the WT. Besides, they observed similar flhD mRNA levels in double and triple deletion mutants compared to the single deletion mutants. While ArcZ and RmaA regulate the transcription of flhD, the same ArcZ and OmrAB affect post-transcriptionally flhD master regulator mRNA.
The broad host range plant pathogen Pantoea ananatis is responsible for yield losses in many important crops. The roles and targets of Hfq were unknown, and a recent study measured the molecular and phenotypical differences between a WT strain versus the Δhfq deletion and the hfq complementing mutant (Shin et al., 2019). The phenotype of Δhfq strains showed a range of phenotypical impairments such as slower growth rate, or loss of virulence when infecting onion. Within virulence traits, the mutants were affected in swimming motility, had a reduced AHL production, and a decreased ability to form biofilms. The complementing mutant resembled the WT strain phenotype. Authors identified the affected sRNAs by comparison of loss-of-function mutant with WT, both in low-density and high-density conditions. After data analysis, expression levels for 9 sRNAs were assayed through RT-qPCRs (arcZ, fnrS, glmZ, rprA, ryeB, ryhB2, pPAR237, pPAR238, and pPAR395), with reduced transcript levels for all them in the mutant. That was not the case for glmZ and ryhB2, and authors conclude that it is due to a negative regulation of those two targets by Hfq in WT strains of P. ananatis.
The study of Yuan et al. (2019) used Dickeya dadantii as a model bacterial pathogen, which depend on swimming motility for migration to the entry structures of the plant. RsmA (CsrA) and RsmB (CsrB) comprise a protein-sRNA system of regulators of the Type III Secretion System (T3SS) and other virulence phenotypes. While RsmA controls negatively hrpL, the master regulator of the T3SS, RsmB can sequester the protein. Besides RsmB, AcrZ (Hfq mediation) is as well involved in the regulation of motility and virulence in D. dadantii.
Infectious Diseases in Animals
As previously stated, infection represents a competition relationship between pathogenic bacteria and a variety of hosts (ranging from plants to animals) for the same metabolic resources (Rohmer et al., 2011). The development of the field is much broader in humans due to the relevance of infectious diseases from an anthropogenic perspective. For other species, the research interest has been driven by economic interest in farming, because the studied species can potentially act as reservoirs for zoonoses (Slingenbergh et al., 2004), or their potential as model organisms.
Non-vertebrates
Several non-vertebrate metazoans have attracted attention of the research community for different reasons, among them their similarity at innate immune responses with mammals (Tanji and Ip, 2005), because they constitute a vector for infectious diseases (Slingenbergh et al., 2004), or their importance as crop pests (Li et al., 2019).
The model organism Caenorhabditis elegans grazes on bacteria in the soil environment. Both bacteria and host establish a relationship which constitutes one of the most clear examples of interkingdom communication (Legüe and Calixto, 2019). It has been reported that E. coli sRNAs (OxyS and DsrA) can impact the expression of genes in the host (Liu et al., 2012). The uptake of non-self RNA molecules can happen via RNA transporters that have as well homologs in humans (Legüe and Calixto, 2019), or through membrane vesicle transport (Dauros-Singorenko et al., 2018).
Mutualistic endosymbionts are subjected to genome erosion, e.g., Buchnera in aphids (Wernegreen, 2002; Bennett and Moran, 2015). This reduction leads to losses in genes encoding for transcription factors, and highlights the importance of sRNAs as alternative regulatory elements. An early study by Hansen and Degnan (2014) found an interesting lack of mRNA expression in different life stages of Buchnera, while it was clear that proteins were differentially expressed. They predicted the involvement of a group of more than 600 sRNAs in protein regulation. Thairu et al. (2018) studied if these sRNAs were to be involved in post-transcriptional regulation events, by evaluation of two life stages of the endosymbiont, the extracellular proliferating stage in aphid embryos of Acyrthosiphon pisum, and the intracellular non-proliferating state in bacteriocytes of the same host. After RNA isolation (size ≤ 200 nt) and library preparation, authors performed sequencing, where data analysis provided a first indication of a group of 90 differentially expressed putative sRNAs. Authors evaluated in vitro the regulatory properties of one of these sRNAs, which putative target is carB, by cloning the regulatory element and its predicted coding sequence (CDS) target into two separate plasmids. The CDS was fused with the green fluorescent protein CDS (GFP), and indication in stabilization of mRNA would be suggested by enhanced fluorescence in the dual-plasmid system, in comparison with the control (where the vector that should contain the sRNA was empty). Results indicated that carB sRNA stabilizes the messenger of the target, and would compensate for the evolutionary loss of regulatory genes. But also, the changes in the insect diet can exert an effect over the endosymbiont. Thairu and Hansen (2019) measured through transcriptomic approaches if changes in the host diet would affect the sRNA regulatory pathways. Authors found that most of the potential targets among two conditions, when the aphid fed on two plants with notable differences in secondary metabolites, were related with amino-acid biosynthesis. The results are probably explained by the intimate relationship between the host insect and the endosymbiont, where changing in the host will drive a different transcriptional program to the cohabiting bacterium.
Wolbachia pipientis is an endosymbiont of insects that can affect the host behavior for ensuring its own vertical transmission (Werren et al., 2008). In experiments of infection of Drosophila melanogaster specimens or Aedes albopictus C6/36 cell lines with W. pipientis, Woolfit et al. (2015) examined the transcriptome response of both host and microorganism. Their results yielded a set of reads corresponding to intergenic regions of the bacteria that together with in silico candidates, allowed authors to identify two putative sRNAs. While further research is required for a better characterization of Wolbachia small regulatory RNAs, authors hypothesize that there could be an involvement in host manipulation.
Vertebrates
High-throughput approaches evaluating the transcriptomic profile of both host and pathogen have become very important for the discovery of new sRNAs and their effect during the infection process (Saliba et al., 2017). The molecular behavior of pathogens is different when cultivated at in vitro conditions than when they are interacting with the host (Westermann et al., 2012). This method is known as “dual RNA-seq” (reviewed in Saliba et al., 2017; Westermann et al., 2017) and was used in bacterial infections models first by Westermann et al. (2016), studying the interaction between Salmonella enterica serovar Typhimurium and HeLa cells. Authors identified a highly expressed sRNA, PinT, which interacts with mRNAs by Hfq mediation. Not only this approach allows to characterize the bacterial sRNA profile, but also the study revealed a correlation of this sRNA with effects over the host immune pathways.
Non-human Models
A great fraction of the development on the field has been driven by economic interest, in order to understand the life cycle of causative agents for bacterial diseases in aquaculture farming industry. These settings concentrate in a short spatial range a significant number of specimens that could become up to 1,000 times higher than natural populations (Sundberg et al., 2016). Intensive fish farming has led to use of antibiotics, which together with the abnormal number of individuals, turn these exploitations into a hot-spot for the selection and spread of antibiotic resistance in the environment (McPhearson et al., 1991). Furthermore, the selection pressure within the microbiome due to interference and competition, directs toward quick changes in pathogen virulence that could be fixed in the genome (Sundberg et al., 2016). A necessary first step for an accurate understanding of the role of sRNAs at infection processes, is to survey the core repertoire of non-coding RNAs, as reported in a recent study (Segovia et al., 2018). Authors analyzed eleven P. salmonis genomes, having described more than 2,000 sRNAs (referred as non-coding RNAs, ncRNAs), from which more than 1,300 formed the ncRNA core group. Analyses of these RNAs have shown that many of the targeted genes in the bacterial genome show similarities to those described in section “sRNAs Regulate Key Processes for the Establishment of Infection.” of the current review. Among these similarities: manganese sensing response, membrane transport, components of the Type I and II Secretion Systems, expression of enzymes necessary for tissue colonization and acid resistance, or regulation of carbon flux.
Besides aquaculture, research in other vertebrates has been driven by biomedical interest in species that could serve as an alternative experimental model for human infectious diseases. In comparison with insect species, murine models show additional advantages, such as the presence of adaptive immunity or a wide range of available genetic resources. A murine macrophage line has been used to evaluate the interaction with Brucella abortus, a pathogen causing infectious diseases in cattle but also in humans (Golding et al., 2001; Budnick et al., 2018). Several sRNAs have been identified in Brucella spp., some of which have been associated with the pathogen virulence (Dong et al., 2018). When the sRNA BASI74 was overexpressed, the virulence of Brucella was negatively affected, while deletion had no effect. This is potentially explained by the presence of different copies of the sRNA in the bacterial genome, due to an important regulatory role. It means that while overexpression produces an excessive regulatory response, loss of an sRNA is compensated by different versions or alternative sRNAs. From an evolutionary perspective, it will help the pathogen to maintain regulation of important genes even when some sRNAs are lost from the genome.
Human as a Host
Recognition of bacterial infective agents relies on the innate immunity and a series of germline-encoded pattern-recognition receptors (PRRs) (Sellge and Kufer, 2015; González Plaza, 2018). Among others, TLRs strongly trigger systemic inflammation via macrophages and neutrophils (Moresco et al., 2011). These receptors can sense as well RNA molecules (Kawai and Akira, 2009), especially through TLR3, TLR7, and TLR8 (Hornung et al., 2008; Moresco et al., 2011). These receptors recognize molecules that are essential for the proper biological function of the bacteria, and thus, will be subjected to less evolutionary modifications than others (Akira et al., 2006).
But not in all cases bacterial RNA unleash an inflammatory response. Milillo et al. (2019) have reported down-modulation of MHC-II surface proteins in human monocytes/macrophages in the presence of Brucella abortus RNA. Because RNA is a molecule with a short lifespan and is subject to rapid degradation, it is present in alive bacteria rather than in dead cells. Authors argue that their findings suggest how B. abortus may thrive undetected within macrophages, due to the impairment of the MHC-II antigen presentation by inhibition of the gene expression. Nevertheless, the effect was also observed with partially degraded RNA. This molecular evasion could happen as well for many other pathogens, that could multiply and cause a serious disease requiring clinical treatment with antimicrobials. However, due to the current antibiotic crisis, the treatment of bacterial infections is becoming increasingly difficult due to the appearance of resistance (Bogaert et al., 2004). Given the role of sRNAs as regulatory molecules, it is clear that understanding their role during infection could lead to improved therapies to control the expression of resistance genes (Dersch et al., 2017). Focusing on sRNAs as therapeutic targets could be done in two main ways, targeting the mechanisms that regulate the virulence and infection related biological processes in the pathogen, or targeting the expression and regulation of antibiotic resistance genes. The latter group of genes are commonly shared among clinical pathogens in matter of years or months (O’Neill, 2014). Understanding interaction of one gene with its sRNA or set of sRNAs would likely be a universal therapy for different bacteria harboring the same resistance factor and become a promising research area for new treatments where bacteria will not presumably develop resistance (Ghaly and Gillings, 2018).
Respiratory diseases
One of the biggest issues during the onset of viral respiratory tract infections, is the frequent co-occurrence of bacterial infections that can dramatically aggravate the symptoms and the number of deaths (Mallia and Johnston, 2007). One of the explanations is that viral infections weaken host defense mechanisms, such as the clearance of bacteria by ciliated epithelial cells. Viral induced impairment of these processes create a favorable environment for the development of side bacterial infections by opportunistic pathogens (Hendaus et al., 2015).
Among those, Staphylococcus aureus can colonize different body surfaces and cause serious respiratory disorders (Lowy, 1998). The pool of more than 600 regulatory sRNAs present in this bacterium remains largely uncharacterized (Tomasini et al., 2014; Sassi et al., 2015; Carroll et al., 2016), but also provides an idea of the potential for adaptation to changing environments and success in the establishment of infection (Bayer et al., 1996). The global regulator sarA locus encodes a protein that regulate directly and indirectly genes involved in virulence (Dunman et al., 2001). This locus has three promoters (P2, P3, and P1 according to their genomic order) in a region of 850 bp upstream of the sarA coding sequence, and a 196 nucleotide sRNA (teg49) located within two of the promoters (Kim et al., 2014). Three overlapping transcripts including sarA ORF are produced. Mutagenesis assays in several regulatory sRNAs and the three promoters, indicated that teg49 is probably generated at the promoter P3 mRNA, likely through cleavage (Manna et al., 2018). Deletion of the P3 promoter resulted in the disappearance of both sarA P3 mRNA and teg49, besides lower SarA protein levels. Additional transcriptomics assays in the teg49 mutant, disclosed a group of genes which were up- and down-regulated in the absence of this sRNA. Those genes were involved in regulation, metabolism, and virulence. Teg41 is another sRNA involved in virulence regulation in S. aureus. It has been further characterized by Zapf et al. (2019; Table 2). This sRNA of approximately 200 nucleotides is located downstream of the transcript region of the potent phenol soluble modulin (PSMs) toxin type α. Authors propose that this sRNA positively regulates the toxin production by mRNA stabilization. Among different assays, they showed how the deletion of the 3′ sRNA region induced decreased levels of the αPSM transcripts, while higher levels of the toxin transcript were reported when Teg41 was overexpressed. The metabolic adaptation to nutritional starvation is also present in this pathogen, with the participation of sRNAs for the regulation of metabolism, as shown for RsaE (Bohn et al., 2010). A recent study on this regulator compared the transcriptomic response of two RsaE mutants, one by deletion, and the other by addition of an inducible promoter, with a wild type strain (Rochat et al., 2018). This sRNA has an effect over genes encoding enzymes of the TCA cycle, but also in the regulation of arginine catabolism, which was newly reported in this study.
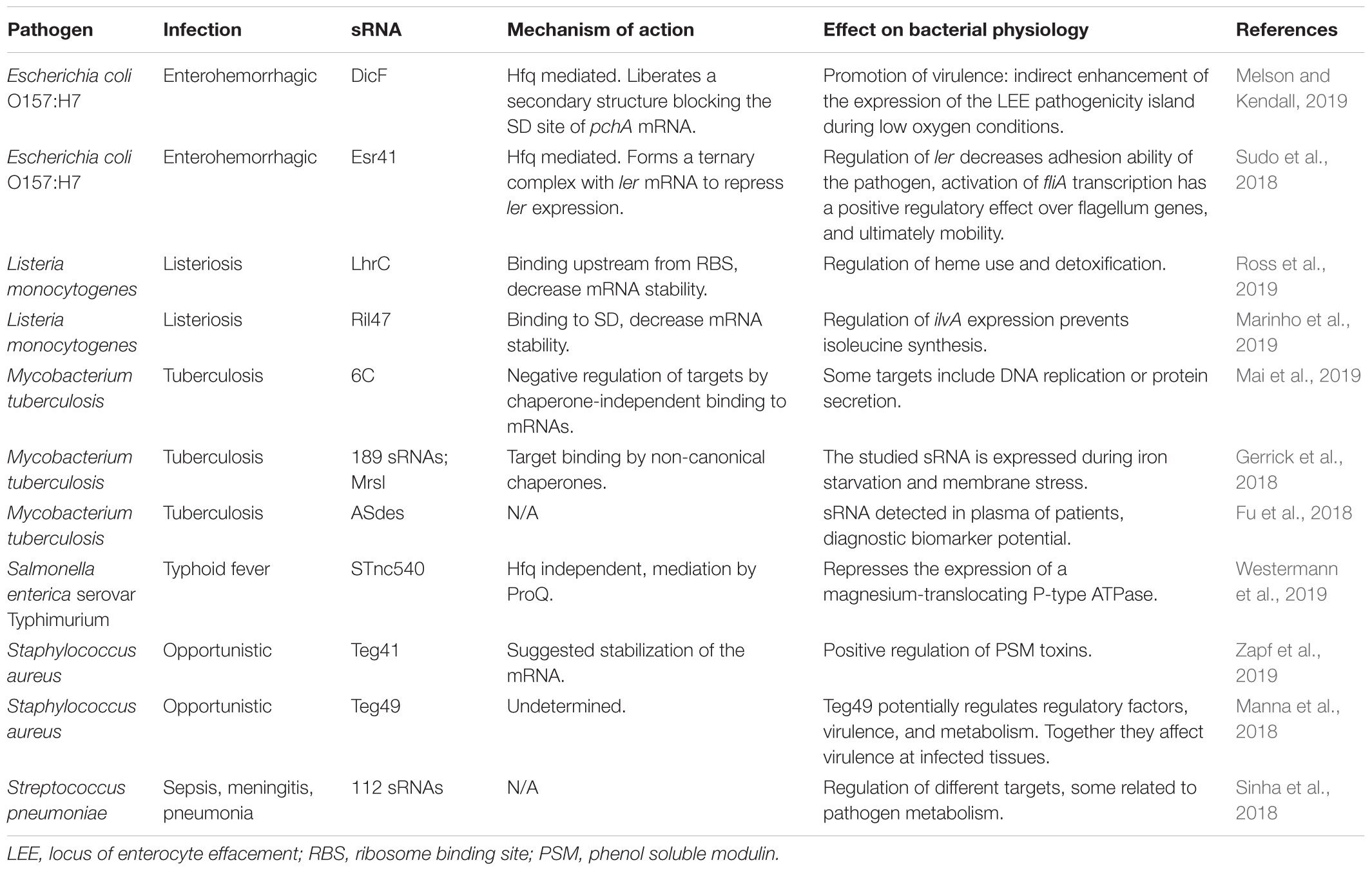
Table 2. Relevant sRNAs of several infectious diseases having humans as a host.
Streptococcus pneumoniae is a Gram-positive bacterium that causes a wide range of complications, including sepsis, meningitis, or pneumonia. It has been suggested that it was a magnifying cause of the high death rates during the 1918 influenza pandemic (Mallia and Johnston, 2007). Sinha et al. (2018) studied through a massive RNA sequencing S. pneumoniae strain D39W grown in laboratory conditions (Table 2). The first list yielded a total of 57 sRNAs, and authors raised the question if those were primarily expressed, or further cleaved by RNase to get to its final form. In that regard, differential RNAseq was carried out, and authors report a group of 44 novel sRNA candidates. These regulatory RNAs fall in three categories: antisense RNAs, short-antisense RNAs, or long-antisense RNAs. Some of these regulatory molecules have been proposed to regulate metabolic responses of the pathogen.
Tuberculosis is one of the most studied infectious diseases due to its incidence. It is estimated to affect 23% of the world population in its latent stage (Lyu L et al., 2019). Although previously the disease was projected to be eradicated by 2010, it has been continuously re-emerging due to a complex combination of factors (Cohen, 2000). The role of sRNAs has been long established, and there is a growing body of knowledge on the topic. However, as reported by Mai et al. (2019), there are considerable gaps of knowledge regarding small regulatory RNAs. The field has discovered until now a wide set of these molecules, but bigger efforts to elucidate their molecular mechanisms of action are needed. The sRNA 6C (six cytosine residues) was hypothesized, by the authors of this study, to play an important role in regulation of cell division of Mycobacterium tuberculosis (M. tb) (Mai et al., 2019). In their experiments, the authors used a vector to overexpress 6C sRNA of M. tb, and transformed Mycobacterium smegmatis. Analyses of expression through RNAseq indicate that the potential targets of 6C could be under negative regulation, through base pairing to the mRNA targets at the C-rich loops. While in Gram-negative bacteria the interaction must be mediated by Hfq, in high GC Gram-positive bacteria has been hypothesized the presence of putative chaperones, or as in this study by direct binding mechanisms independent of chaperones (Mai et al., 2019; Table 2).
Additional regulation of Mycobacterium tuberculosis by sRNAs was provided in the study of Gerrick et al. (2018), who tested the landscape of small regulatory RNAs in the presence of five stresses relevant for the pathogen, having found 189 sRNAs (Table 2). The direct interaction of one of those sRNAs, MrsI, with the target mRNA is potentially mediated by non-canonical chaperones, given that there are no described HfQ or ProQ homologs in mycobacteria. MrsI is expressed in conditions of iron starvation and membrane stress, when the host immune system triggers an inflammatory response including an iron withholding strategy, especially when entering macrophages. This response seems to be activated when the bacteria starts to suffer membrane and oxidative stresses, as the previous state of iron deprivation. This anticipatory response is not exclusive for M. tuberculosis, but according to authors has been observed in different prokaryotic organisms and it may have been developed as an evolutionary response to the inflammatory response by the host.
There is interest in developing sRNAs as stable markers for early diagnosis of the disease (Fu et al., 2018). Bacterial cell cultures of tuberculosis were studied by Fu and collaborators (Fu et al., 2018), who report four types of sRNAs (ASdes, ASpks, AS1726, and AS1890) (Table 2). On a first phase of this study, the authors tested whether these sRNAs would be detectable in the supernatant of cultures of Mycobacterium bovis BCG. Cultures of BCG were selected because the regulatory RNAs display 100% identity in silico between both strains. Once it was clear that the methodology was robust for identification of sRNAs from supernatant, authors tested the plasma of infected patients (Mycobacterium tuberculosis) in comparison with healthy controls. Among the four above mentioned regulatory RNAs, only ASdes was traced to the plasma of most tuberculosis patients, with statistical significance. These results are promising for the development of quick diagnostic assays based on the detection in blood or saliva of sRNAs derived from pathogens.
Gastrointestinal pathogenic diseases
When enterohemorrhagic Escherichia coli O157:H7 (EHEC), a food-borne pathogen, enters the digestive tract, it must adapt from oxygen rich environments to an increasingly oxygen-limiting environment toward the colon, where pathogens sense the differential gas concentrations and start to express their virulence factors (Melson and Kendall, 2019). As mentioned above, this is a key behavior of bacterial strains in order to set the basis of the infection onset, which seems to have evolved to start from a very low number of input cells (Van Elsas et al., 2011). DicF is a sRNA dependent on Hfq directly targeting the SD sequence of the transcriptional activator pchA, to promote its expression under low oxygen conditions (Melson and Kendall, 2019; Table 2). Ultimately, DicF has an effect over virulence by indirect increase of the expression of the locus of enterocyte effacement (LEE) pathogenicity island, but only when Hfq is present and oxygen concentration is very low (Figure 8). This means that during the disease establishment and progression, the pathogen has a mechanism to sense when it has reached the colon so that the virulence factors should be expressed at the right time. One of the most relevant and novel aspects of the previous study, is the reporting on a sRNA that has a positive expression effect by liberating a transcriptional activator virulence gene from its own mRNA repressive activities (Figure 8). Another sRNA was recently shown to be involved in regulation of LEE and flagellar genes (Sudo et al., 2018). Esr41 requires Hfq mediation, and has regulatory functions over cell motility (Sudo et al., 2014) by induced activation of fliA, which encodes the transcription factor σ28 that regulates the expression of the class 3 flagellin subunit genes (Aldridge et al., 2006). Esr41 forms a ternary complex with the ler mRNA and Hfq for repression of the gene (Figure 9), leading to consequent repression of the LEE and a decrease in the adhesion ability of the pathogen (Sudo et al., 2018).
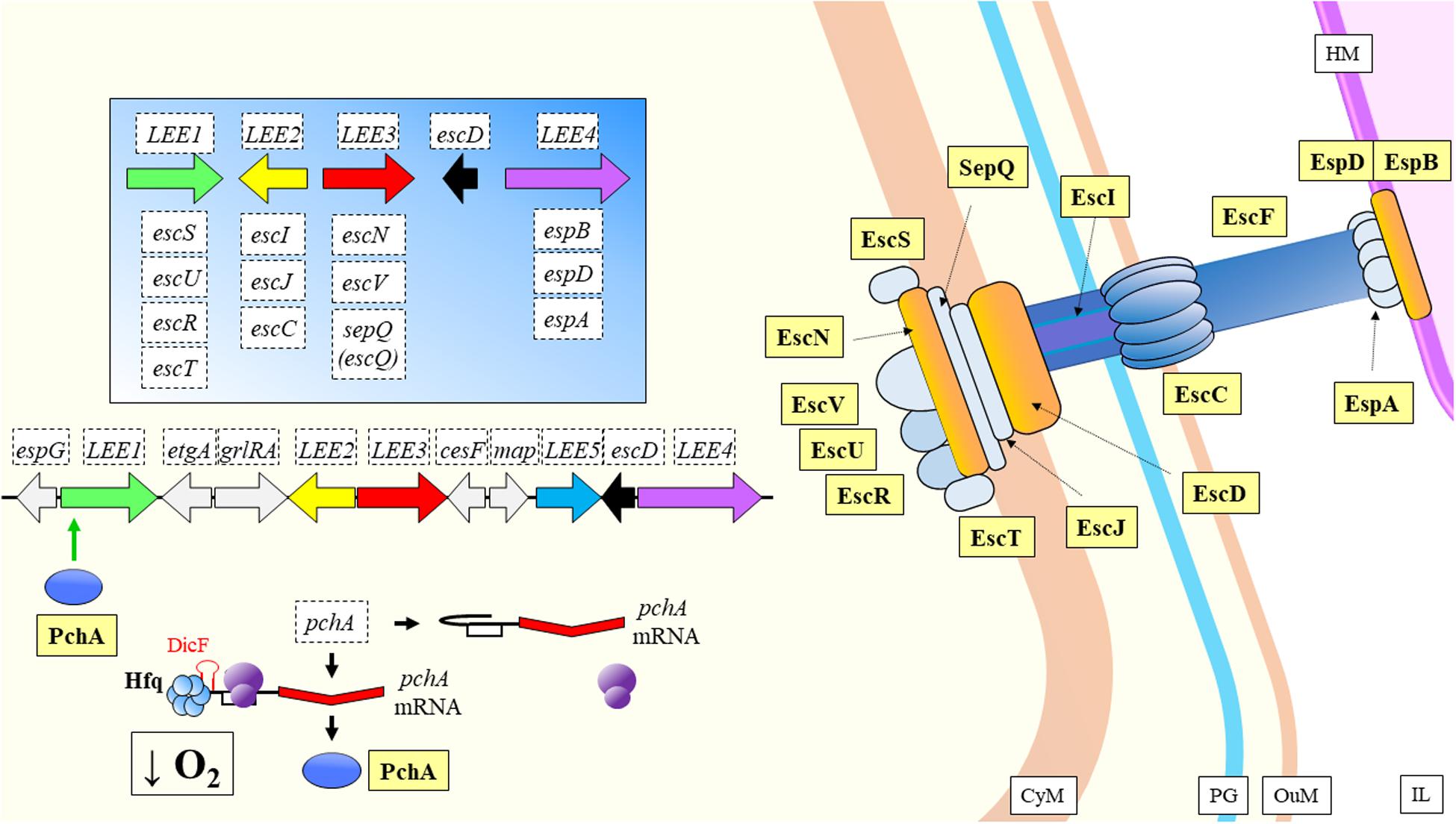
Figure 8. Type III secretion system in EHEC. The pchA transcript has autoinhibitory activity, as it forms a secondary structure covering the SD site. When concentration of oxygen is low, DicF by mediation of Hfq frees the SD of pchA, allowing translation. PchA activates transcription of LEE genes involved in the synthesis of components of the T3SS. CyM, Cytoplasmic membrane; HM, Host cell membrane; IL, Gastrointestinal lumen; OuM, Outer membrane; PG, peptidoglycan; T3SS, Type III secretion system. Black thick arrows indicate flow of a biological process, e.g., protein translation. Green thick arrows indicate activation. References: Deng et al., 2004; Pallen et al., 2005; Izoré et al., 2011; Ruano-Gallego et al., 2015; Furniss and Clements, 2018; Pena et al., 2019.
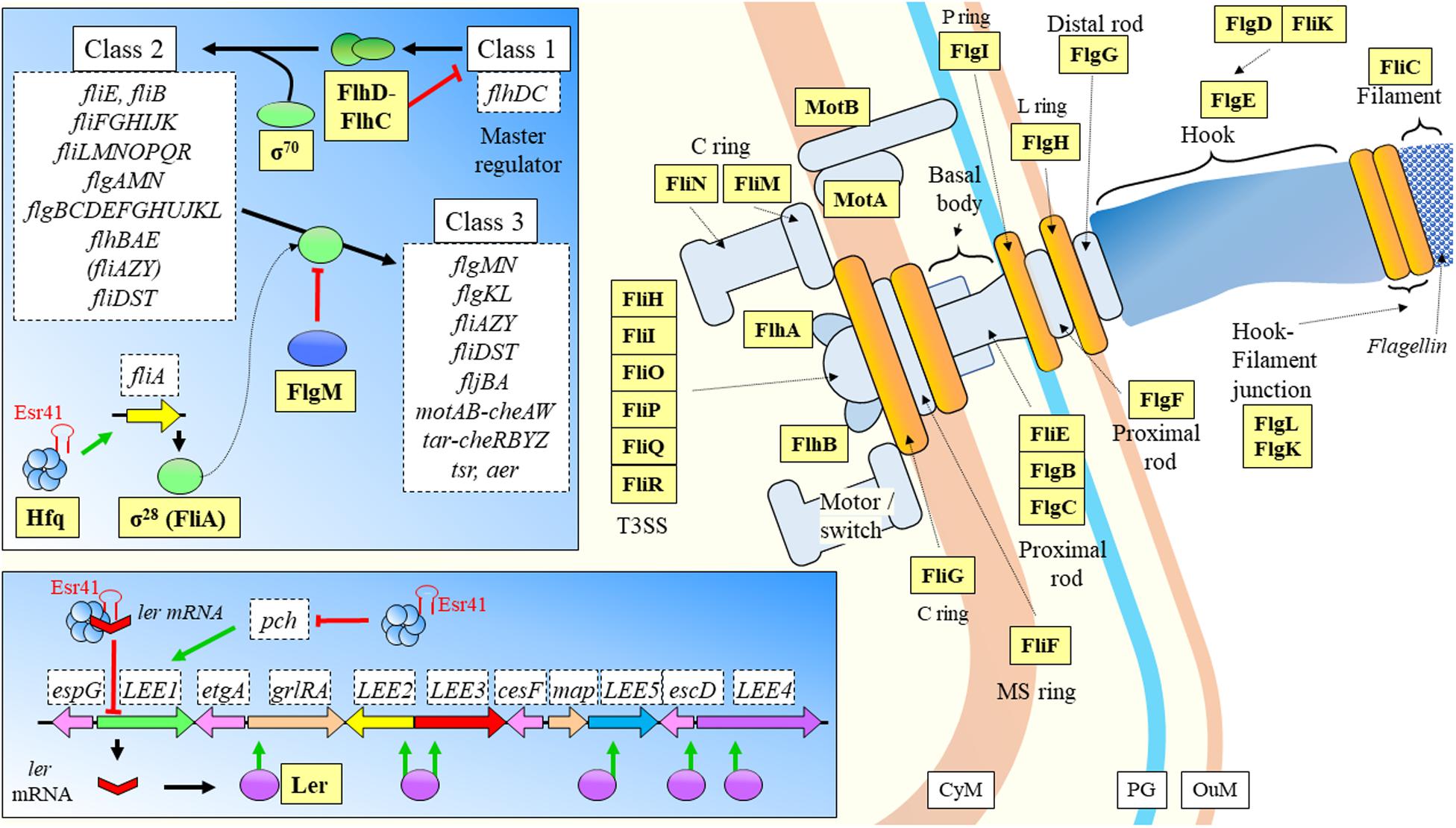
Figure 9. EHEC flagellar system. The fliA encodes for a transcription factor controlling expression of class 3 genes. The sRNA Esr41 regulates positively via Hfq mediation fliA. The flagellar system is regulated by sRNAs at a key point. Esr41 has as well repressing activities over LEE1 operon, when it forms a ternary complex with Hfq and ler mRNA. This inhibition compromises coding of Ler protein, which is a transcriptional activator of different genes at the LEE. Esr41 has as well inhibitory activities over pch that facilitate LEE1 gene expression through promoter binding. CyM, Cytoplasmic membrane; HM, Host cell membrane; IL, Gastrointestinal lumen; OuM, Outer membrane; PG, peptidoglycan; T3SS, Type III secretion system. Black thick arrows indicate flow of a biological process, e.g., protein translation. Green thick arrows indicate activation. Red thick arrows with flat cap indicate inhibition. References: Kalir et al., 2001; Chevance et al., 2006; Frye et al., 2006; Liu and Ochman, 2007; Chevance and Hughes, 2008; Sudo et al., 2014, 2018; Bhatt et al., 2016 and KEGG (https://www.genome.jp/kegg/pathway/eco/eco02040.html).
Listeria monocytogenes is the causative agent of listeriosis (Ágoston et al., 2009), characterized by different episodes of fever, diarrhea, or meningoencephalitis among other symptoms (Swaminathan and Gerner-Smidt, 2007). There are mainly two forms of this disease, the non-invasive gastrointestinal febrile manifestation, and the invasive life-threatening form causing for instance meningoencephalitis (Allerberger and Wagner, 2010). In this organism, the seven sRNAs comprising the LhrC family act by regulating the target tcsA that encodes a virulence protein, CD4+ T cell-stimulating antigen (Sanderson et al., 1995). The regulatory mechanism of LhrCs was recently elucidated, and follow a non-canonical pathway, where sRNAs bind far upstream of the RBS, impairing the stability of the mRNA through the participation of an unknown RNase decreasing the messenger half-life (Ross et al., 2019; Table 2). During the nutritional starvation confronted by the pathogen in the host, the bacteria can make use of the available iron at the host iron-containing proteins, as the heme group in hemoglobin (Cassat and Skaar, 2013). LhrC1-5, belonging to the LhrC family of sRNAs, has been related to the regulation of heme usage in this pathogen. dos Santos et al. (2018) have shown the involvement of these five sRNAs in the regulation of three genes that control heme toxicity (tcsA, oppA, and lapB). Besides, authors found indications of regulation through these sRNAs of three genes related with the uptake and use of heme. Host induced starvation of amino acids is one of the strategies of nutritional immunity used by mammalian hosts to limit the infection (Zhang and Rubin, 2013). In L. monocytogenes Ril47 is an sRNA under control of the stress activated sigma factor sigma B (σB), which is involved in a plethora of environmental stresses affecting the pathogen (Dorey et al., 2019). Ril47 binds to the SD sequence of the ilvA mRNA decreasing its stability, as suggested by a recent study (Marinho et al., 2019). The consequence is a negative regulation of isoleucine biosynthesis by prevention of threonine deaminase synthesis (which is encoded by ilvA).
Bacteria belonging to the genus Salmonella cause mainly two affections: diarrheal disease and typhoid fever. The latter has a mortality rate of 10−20% if it is not treated with antibiotics, being its causative agent is Salmonella enterica serovar Typhimurium (Fàbrega and Vila, 2013). An example of influence of sRNAs over the nutritional adaptation of this pathogen was provided in the study of Kröger et al. (2018), where they report two novel sRNAs involved in positive (STnc1740) and negative (RssR) modulation of the growth of S. enterica serovar Typhimurium, through utilization of myo-inositol as carbon source. While the role of Hfq has been widely studied in this pathogen, Westermann et al. (2019) evaluated the contribution of ProQ as alternative RBP. Authors followed a previous study where ProQ was reported to display novel posttranscriptional regulatory activities (Smirnov et al., 2016). The study from Westermann sought to shed light over the role of ProQ in virulence by altering the phenotype (deletion of the RBP) and used a dual RNA-seq approach. The invasion capabilities of the single mutant were decreased (2-fold vs. WT), but no dramatic differences were found when the RBP was overexpressed. While these phenotypes were observed when infecting HeLa cells, very mild virulence was reported during infection of human phagocytic cells. Further evaluation of the dual RNA-seq results (HeLa cells as host) revealed alteration in metabolism of the host and immune signaling pathways. The mutant pathogen had a range of transcriptomic alterations over genes involved in chemotaxis and motility (downregulation), or in ribosomal and invasion genes (upregulation). In their analysis authors identified STnc540 as an sRNA independent of Hfq and regulated by ProQ. The interplay between sRNA and RBP was further shown to affect the expression of a magnesium-translocating P-type ATPase.
Antibiotic resistance
Antibiotic resistance is a serious concern for society that threatens health systems worldwide (O’Neill, 2014). One the biggest problems is the continuous appearance of novel mechanisms of resistance often fuelled by the natural presence of those mechanisms in the environment, quick global spread of antibiotic resistance genes (ARGs), uptake by pathogenic bacteria that can become resistant to several antibiotics, and lastly the lack of alternatives for treatment of infections caused by multidrug resistant bacteria (MDR) (O’Neill, 2014). The emergence of novel antibiotic resistance genes has dramatic consequences when they reach the clinical practice as they will be irreversibly fixed (Böhm et al., 2020). This brings to memory ideas of bacterial caused plagues in the pre-antibiotic era, when there were no means of controlling pathogens. Resistance to antimicrobials is additionally shaped by environmental stresses (in many cases of anthropogenic origin) faced by bacteria (Poole, 2012), but also interestingly coincide with those highlighted in section “sRNAs Regulate Key Processes for the Establishment of Infection.”
Genes conferring resistance to antibiotics are essentially subjected to genetic regulation, and thus, their expression can be as well tuned by sRNAs (Lalaouna et al., 2014). This fact makes sRNAs a potentially invaluable tool to treat emerging infections mediated by antibiotic resistant bacteria (ARBs). An increasing number of studies point toward that direction, and have been excellently reviewed by Lalaouna et al. (2014), Dersch et al. (2017), Felden and Cattoira (2018). The antibiotic susceptibility of multidrug-resistant (MDR) Pseudomonas aeruginosa was dramatically altered when AS1974 sRNA was overexpressed (Law et al., 2019). Authors of the study carried out a first screening on the sRNAs repertoire on three MDR clinical isolates, having found three small-regulatory RNAs that were significantly downregulated. Targets of AS1974 are involved mostly in pathways related with drug-resistance, but also regulation over oxidative stress, pilus biogenesis, and iron acquisition.
Escherichia coli can activate a stress-response mechanism mediated by RpoS (Poole, 2012). However, this mechanism is under regulation of a non-coding RNA, rprA, which induced ampicillin resistance when overexpressed (Sahni et al., 2019). The mRNA levels of this sRNA had been previously shown to increase in cells when treated with sub-inhibitory concentrations of ampicillin (Gutierrez et al., 2013).
Shigella strains are gaining attention due to the accumulation of resistances toward antibiotics (Puzari et al., 2018), for instance using efflux pumps to extrude fluoroquinolones from their intracellular space (Roy et al., 2020). Gan and Tan (2019) evaluated whether the sRNA SdsR, a transcriptomic regulator of the efflux pump tolC in E. coli and Salmonella (Choi et al., 2018), plays as well a regulatory role in Shigella sonnei or not. Although in previous reports SdsR lowers the levels of TolC mRNA and increases its sensitivity to fluoroquinolones, in their study the authors found that at decreasing levels of TolC mRNA the strain was less sensitive to antibiotics, such as norfloxacin.
Discussion
This review focuses on the latest advances on small non-coding regulatory RNAs involved in bacterial infections, a field that has grown substantially in the last 2 years since the appearance of a previous review article (González Plaza, 2018). The importance of studying infectious diseases lies within the fact that global warming can lead to changing patterns in the distribution of bacteria causing diseases, or in the vectors harboring those microorganisms (Cohen, 2000; Epstein, 2001; Rodó et al., 2002; Khasnis and Nettleman, 2005). But not only from an anthropogenic perspective, these changing patterns in the distribution of infectious diseases can even cause the extinction of whole species (Harvell et al., 2002; Pounds et al., 2006).
Transference of information is a key process for quick adaptation to dynamic environments or species interactions, being one clear example the host-pathogen relationship. Infectious diseases have been shaped by co-evolution between pathogen and animal or plant hosts, both competing for the same metabolic resources. Due to the nature of RNA as a molecule with a short life-span, its role for modulation of environmental signals is of paramount importance for success of infection. This process represents a challenge for the survival of the bacteria, because the host sets a wide range of measures to limit the growth of the microbial strains, eliminate them, and recover homeostasis. The overall role of sRNAs is to fine-tune the bacterial response to different stresses, with an impressive amount of mechanisms of induction/repression of transcription. Not only they affect specific genes involved at synthesis/degradation pathways, but in many cases, they regulate the activity of transcription factors and regulators. Out of several studies it is clear that sRNAs can have multiple targets, and even achieve regulatory redundancy with other non-coding regulatory RNAs.
Innate immune system response can limit nutrient (e.g. iron and manganese) availability to induce death of the pathogen, and a series of sRNAs help bacteria to quickly adapt to immune system challenges, such as the iron or nutrient withholding strategy, where the host limits the available resources in order to limit the survival chances of the invading bacteria. Bacteria encounter some other challenges in the host, for instance acid pH in the stomach of animals, or inflammation leading to oxidative stress. Many of these microbial responsive strategies seem to be conserved from environmental bacteria to pathogens (e.g., involvement of sRNAs in oxidative stress response due to salinity or inflammation), although it could have appeared owing to convergent evolution since sRNAs are very flexible molecules in their activities.
As presented in this review, a very useful approach, is to study both mRNA and sRNAs in the same samples, because it can lead to elucidate regulatory networks.
A number of the studies reviewed here are focused on surveying the sRNome repertoire, or also on the regulation of targets. There is a need to carry on with these studies, since this basic research is fundamental for being able to understand e.g., which genes could be directly modulated by the pathogen. This brings to memory the Spanish Presidential Elections in 2019, where as always, political parties kept promising the rise in research funds to 2% of GDP. Although every Spanish Scientist understands the magnitude of this false assertion, it was concerning how politicians have made theirs the mantra of “Applied Science” as the sole generator of progress and development. For instance, Pfizer was reported to have abandoned recently their program of applied research in neurodegenerative diseases (Trullàs, 2019), which should be an example of “how could we apply what we do not fully understand?”. This seems to be a trend in every scientific system around the world (Lindsley, 2016), where the application paragraph is requested at every grant proposal, and does not seem possible to carry out a research project where the main aim is to obtain a “general understanding of nature and its laws” (Bush, 1945). It becomes clear why these type of basic studies are needed, because control of infections through drugs that can target pathogen sRNA regulation are not possible without a deep understanding of their structure, modes of regulation, behavior patterns during infection, or what genes alter their expression during infection. Small non-coding regulatory RNAs are a promising therapeutic target in the fight against multidrug-resistant bacteria, and since these molecules are less prone to changes, we may have a great opportunity to tackle antibiotic resistance through gene-based therapies.
Having said that, what is still largely missing in the field are more studies focusing on the inter-kingdom exchange of sRNAs, either as counterfeit to impair the activity of the bacteria, or as a pathogenic mechanism to hijack the metabolic machinery of the host. Many protein effectors, such as those translocated by the T3SS can modify the response from the host. In the case of sRNAs, membrane vesicles (MVs) can carry the genetic material to their targets protecting it from degradation (Toyofuku et al., 2019), and ultimately subvert the molecular behavior of the host (Ghosal, 2017; González Plaza, 2018).
Lastly, the wide regulatory effects of sRNAs over the transcriptome must be taken in account due to their potential to impact genetic studies where a loss-of-function phenotype is considered as suggested by Popitsch et al. (2017). Deletion may affect expression of unannotated sRNAs and have undesired effects over the phenotype because regulation of a number of targets could be lost. It could as well potentially affect the expression of targets if some coding genetic regions carrying sRNAs are overexpressed. These possibilities invite to consider carefully the presence of small regulatory RNAs in genetic studies, and maybe even revisit some of the studies to date.
Author Contributions
JG reviewed the literature, constructed all tables and figures according to published literature, and wrote the manuscript.
Funding
The author has been funded by projects IGA No. B_19_04 from the Faculty of Forestry and Wood Sciences (Czech University of Life Sciences, Prague); and “Advanced research supporting the forestry and wood-processing sector’s adaptation to global change and the 4th industrial revolution,” OP RDE, Ministry of Education Youth and Sports of Czechia, Grant No. CZ.02.1.01/0.0/0.0/16_019/0000803.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
I want to thank Assoc. Prof. J. Šobotník, for comments regarding the introduction and Ph.D. candidate P. Stiblík for technical support. I want to acknowledge the constructive comments of the reviewers.
References
Agliano, F., Rathinam, V. A., Medvedev, A. E., Vanaja, S. K., and Vella, A. T. (2019). Long noncoding RNAs in host-pathogen interactions. Trends Immunol. 40, 492–510. doi: 10.1016/j.it.2019.04.001
Ágoston, R., Soni, K., Jesudhasan, P. R., Russell, W. K., Mohácsi-Farkas, C., and Pillai, S. D. (2009). Differential expression of proteins in Listeria monocytogenes under thermotolerance-inducing, heat shock, and prolonged heat shock conditions. Foodborne Pathog. Dis. 6, 1133–1140. doi: 10.1089/fpd.2009.0286
Aguilar, C., Mano, M., and Eulalio, A. (2019). MicroRNAs at the host-bacteria interface: host defense or bacterial offense. Trends Microbiol. 27, 206–218. doi: 10.1016/j.tim.2018.10.011
Akira, S., Uematsu, S., and Takeuchi, O. (2006). Pathogen recognition and innate immunity. Cell 124, 783–801. doi: 10.1016/j.cell.2006.02.015
Aldridge, P. D., Karlinsey, J. E., Aldridge, C., Birchall, C., Thompson, D., Yagasaki, J., et al. (2006). The flagellar-specific transcription factor, σ28, is the type III secretion chaperone for the flagellar-specific anti- σ28 factor FlgM. Genes Dev. 20, 2315–2326. doi: 10.1101/gad.380406
Allerberger, F., and Wagner, M. (2010). Listeriosis: a resurgent foodborne infection. Clin. Microbiol. Infect. 16, 16–23. doi: 10.1111/j.1469-0691.2009.03109.x
Álvarez-Estrada, Á, Gutiérrez-Martín, C. B., Rodríguez-Ferri, E. F., and Martínez-Martínez, S. (2018). Transcriptomics of Haemophilus (Glässerella) Parasuis serovar 5 subjected to culture conditions partially mimetic to natural infection for the search of new vaccine antigens. BMC Vet. Res. 14:326. doi: 10.1186/s12917-018-1647-1641
Andrews, S. C., Robinson, A. K., and Rodríguez-Quiñones, F. (2003). Bacterial iron homeostasis. FEMS Microbiol. Rev. 27, 215–237. doi: 10.1016/S0168-6445(03)00055-X
Atkinson, G. C., Tenson, T., and Hauryliuk, V. (2011). The RelA/SpoT homolog (RSH) superfamily: distribution and functional evolution of ppGpp synthetases and hydrolases across the tree of life. PLoS One 6:e23479. doi: 10.1371/journal.pone.0023479
Ausubel, F. F. M. (2005). Are innate immune signaling pathways in plants and animals conserved? Nat. Immunol. 6, 973–979. doi: 10.1038/ni1253
Baker, Y. R., Hodgkinson, J. T., Florea, B. I., Alza, E., Galloway, W. R. J. D., Grimm, L., et al. (2017). Identification of new quorum sensing autoinducer binding partners in: Pseudomonas aeruginosa using photoaffinity probes. Chem. Sci. 8, 7403–7411. doi: 10.1039/c7sc01270e
Battesti, A., Majdalani, N., and Gottesman, S. (2011). The RpoS-Mediated General Stress Response in Escherichia coli. Annu. Rev. Microbiol. 65, 189–213. doi: 10.1146/annurev-micro-090110-102946
Bayer, M. G., Heinrichs, J. H., and Cheung, A. L. (1996). The molecular architecture of the sar locus in Staphylococcus aureus. J. Bacteriol. 178, 4563–4570. doi: 10.1128/jb.178.15.4563-4570.1996
Beebe, S., Rao, I., Mukankusi, C., and Buruchara, R. (2013). Improving Resource Use Efficiency And Reducing Risk Of Common Bean Production In Africa, Latin America, And The Caribbean, In Eco-Efficiency: From Vision To Reality, 117-134. Available online at: http://ciat.cgiar.org/wp-content/uploads/2013/04/eco-efficiency_book.pdf (accessed December 17, 2019).
Beisel, C. L., and Storz, G. (2010). Base pairing small RNAs and their roles in global regulatory networks. FEMS Microbiol. Rev. 34, 866–882. doi: 10.1111/j.1574-6976.2010.00241.x
Benbow, M. E., Pechal, J. L., Tomberlin, J. K., and Jordan, H. R. (2018). “Interkingdom community interactions in disease ecology,” in The Connections Between Ecology and Infectious Disease. Advances in Environmental Microbiology, Vol. 5, ed. C. Hurst, (Cham: Springer).
Bennett, G. M., and Moran, N. A. (2015). Heritable symbiosis: the advantages and perils of an evolutionary rabbit hole. Proc. Natl. Acad. Sci. U.S.A. 112, 10169–10176. doi: 10.1073/pnas.1421388112
Bermúdez-Barrientos, J. R., Ramírez-Sánchez, O., Chow, F. W.-N., Buck, A. H., and Abreu-Goodger, C. (2020). Disentangling sRNA-Seq data to study RNA communication between species. Nucleic Acids Res. 48:e21. doi: 10.1093/nar/gkz1198
Bhatt, S., Egan, M., Jenkins, V., Muche, S., and El-Fenej, J. (2016). The tip of the iceberg: on the roles of regulatory small RNAs in the virulence of enterohemorrhagic and enteropathogenic Escherichia coli. Front. Cell. Infect. Microbiol. 6:105. doi: 10.3389/fcimb.2016.00105
Bianco, C. M., Fröhlich, K. S., and Vanderpool, C. K. (2019). Bacterial cyclopropane fatty acid synthase mRNA is targeted by activating and repressing small RNAs. bioRxiv [Preprint], doi: 10.1128/jb.00461-419
Bidnenko, V., Nicolas, P., Grylak-Mielnicka, A., Delumeau, O., Auger, S., Aucouturier, A., et al. (2017). Termination factor Rho: from the control of pervasive transcription to cell fate determination in Bacillus subtilis. PLoS Genet. 13:e1006909. doi: 10.1371/journal.pgen.1006909
Bogaert, D., De Groot, R., and Hermans, P. W. M. (2004). Streptococcus pneumoniae colonisation: the key to pneumococcal disease. Lancet Infect. Dis. 4, 144–154. doi: 10.1016/S1473-3099(04)00938-937
Böhm, M.-E., Razavi, M., Marathe, N. P., Flach, C.-F., and Larsson, D. G. J. (2020). Discovery of a novel integron-borne aminoglycoside resistance gene present in clinical pathogens by screening environmental bacterial communities. Microbiome 8:41. doi: 10.1186/s40168-020-00814-z
Bohn, C., Rigoulay, C., Chabelskaya, S., Sharma, C. M., Marchais, A., Skorski, P., et al. (2010). Experimental discovery of small RNAs in Staphylococcus aureus reveals a riboregulator of central metabolism. Nucleic Acids Res. 38, 6620–6636. doi: 10.1093/nar/gkq462
Borgmann, J., Schäkermann, S., Bandow, J. E., and Narberhaus, F. (2018). A small regulatory RNA controls cell wall biosynthesis and antibiotic resistance. mBio 9:e002100-18. doi: 10.1128/mBio.02100-2118
Brant, E. J., and Budak, H. (2018). Plant small non-coding RNAs and their roles in biotic stresses. Front. Plant Sci. 9:1038. doi: 10.3389/fpls.2018.01038
Brouwer, S., Pustelny, C., Ritter, C., Klinkert, B., Narberhaus, F., and Häussler, S. (2014). The PqsR and RhlR transcriptional regulators determine the level of Pseudomonas quinolone signal synthesis in Pseudomonas aeruginosa by producing two different pqsABCDE mRNA Isoforms. J. Bacteriol. 196, 4163–4171. doi: 10.1128/JB.02000-2014
Budnick, J. A., Sheehan, L. M., Kang, L., Michalak, P., and Caswell, C. C. (2018). Characterization of three small proteins in Brucella abortus linked to fucose utilization. J. Bacteriol. 200:e00127-18. doi: 10.1128/JB.00127-118
Burne, R. A., and Chen, Y. Y. M. (2000). Bacterial ureases in infectious diseases. Microbes Infect. 2, 533–542. doi: 10.1016/S1286-4579(00)00312-319
Burtnick, M. N., Downey, J. S., Brett, P. J., Boylan, J. A., Frye, J. G., Hoover, T. R., et al. (2007). Insights into the complex regulation of rpoS in Borrelia burgdorferi. Mol. Microbiol. 65, 277–293. doi: 10.1111/j.1365-2958.2007.05813.x
Butz, H. A., Mey, A. R., Ciosek, A. L., and Payne, S. M. (2019). Vibrio cholerae CsrA directly regulates varA to increase expression of the three nonredundant Csr small RNAs. mBio 10:e001042-19. doi: 10.1128/mBio.01042-1019
Caldelari, I., Chao, Y., Romby, P., and Vogel, J. (2013). RNA-mediated regulation in pathogenic bacteria. Cold Spring Harb. Perspect. Med. 3:10298. doi: 10.1101/cshperspect.a010298
Carlos, A. R., Weis, S., and Soares, M. P. (2018). Cross-talk between iron and glucose metabolism in the establishment of disease tolerance. Front. Immunol. 9:2498. doi: 10.3389/fimmu.2018.02498
Carrier, M.-C., Lalaouna, D., and Massé, E. (2018a). Broadening the definition of bacterial small RNAs: characteristics and mechanisms of action. Annu. Rev. Microbiol. 72, 141–161. doi: 10.1146/annurev-micro-090817-62607
Carrier, M. C., Laliberté, G., and Massé, E. (2018b). Identification of new bacterial small RNA targets using MS2 affinity purification coupled to RNA sequencing. Methods Mol. Biol. 1737, 77–88. doi: 10.1007/978-1-4939-7634-8_5
Carroll, R. K., Weiss, A., Broach, W. H., Wiemels, R. E., Mogen, A. B., Rice, K. C., et al. (2016). Genome-wide annotation, identification, and global transcriptomic analysis of regulatory or small RNA gene expression in Staphylococcus aureus. mBio 7:1990. doi: 10.1128/mBio.01990-1915
Casadevall, A. (2016). thermal restriction as an antimicrobial function of fever. PLoS Pathog. 12:5577. doi: 10.1371/journal.ppat.1005577
Casadevall, A., and Pirofski, L. (2003). The damage-response framework of microbial pathogenesis. Nat. Rev. Microbiol. 1, 17–24. doi: 10.1038/nrmicro732
Cassat, J. E., and Skaar, E. P. (2013). Iron in infection and immunity. Cell Host Microbe 13, 509–519. doi: 10.1016/j.chom.2013.04.010
Chakravarty, S., and Massé, E. (2019). RNA-dependent regulation of virulence in pathogenic bacteria. Front. Cell. Infect. Microbiol. 9:337. doi: 10.3389/fcimb.2019.00337
Chambonnier, G., Roux, L., Redelberger, D., Fadel, F., Filloux, A., Sivaneson, M., et al. (2016). The hybrid histidine kinase LadS forms a multicomponent signal transduction system with the GacS/GacA two-component system in Pseudomonas aeruginosa. PLoS Genet. 12:e1006032. doi: 10.1371/journal.pgen.1006032
Chatterji, D., and Ojha, A. K. (2001). Revisiting the stringent response, ppGpp and starvation signaling. Curr. Opin. Microbiol. 4, 160–165. doi: 10.1016/S1369-5274(00)00182-X
Chen, R., Wei, X., Li, Z., Weng, Y., Xia, Y., Ren, W. R., et al. (2019). Identification of a small RNA that directly controls the translation of the quorum sensing signal synthase gene rhlI in Pseudomonas aeruginosa. Environ. Microbiol. 21, 2933–2947. doi: 10.1111/1462-2920.14686
Chevance, F. F. V., and Hughes, K. T. (2008). Coordinating assembly of a bacterial macromolecular machine. Nat. Rev. Microbiol. 6, 455–465. doi: 10.1038/nrmicro1887
Chevance, F. F. V., Karlinsey, J. E., Wozniak, C. E., and Hughes, K. T. (2006). A little gene with big effects: a serT mutant is defective in flgM gene translation. J. Bacteriol. 188, 297–304. doi: 10.1128/JB.188.1.297-304.2006
Choi, J. S., Kim, W., Suk, S., Park, H., Bak, G., Yoon, J., et al. (2018). The small RNA, SdsR, acts as a novel type of toxin in Escherichia coli. RNA Biol. 15, 1319–1335. doi: 10.1080/15476286.2018.1532252
Cobaxin, M., Martínez, H., Ayala, G., Holmgren, J., Sjoling, A., and Sánchez, J. (2014). Cholera toxin expression by El Tor Vibrio cholerae in shallow culture growth conditions. Microb. Pathog. 66, 5–13. doi: 10.1016/j.micpath.2013.11.002
Coggan, K. A., and Wolfgang, M. C. (2012). Global regulatory pathways and cross-talk control Pseudomonas aeruginosa environmental lifestyle and virulence phenotype. Curr. Issues Mol. Biol. 14, 47–70. doi: 10.21775/cimb.014.047
Cohen, M. L. (2000). Changing patterns of infectious disease. Nature 406, 762–767. doi: 10.1038/35021206
Cornelis, P., Wei, Q., Andrews, S. C., and Vinckx, T. (2011). Iron homeostasis and management of oxidative stress response in bacteria. Metallomics 3, 540–549. doi: 10.1039/c1mt00022e
Cristaldo, P. F., Jandák, V., Kutalová, K., Rodrigues, V. B., Brothánek, M., Jiøíèek, O., et al. (2016). The nature of alarm communication in Constrictotermes cyphergaster (Blattodea: Termitoidea: Termitidae): the integration of chemical and vibroacoustic signals. Biol. Open 4, 1649–1659. doi: 10.1242/bio.014084
Damo, S. M., Kehl-Fie, T. E., Sugitani, N., Holt, M. E., Rathi, S., Murphy, W. J., et al. (2013). Molecular basis for manganese sequestration by calprotectin and roles in the innate immune response to invading bacterial pathogens. Proc. Natl. Acad. Sci. U.S.A. 110, 3841–3846. doi: 10.1073/pnas.1220341110
Dangl, J. L., and Jones, J. D. G. (2001). Plant pathogens and integrated defence responses to infection. Nature 411, 826–833. doi: 10.1038/35081161
Dauros-Singorenko, P., Blenkiron, C., Phillips, A., and Swift, S. (2018). The functional RNA cargo of bacterial membrane vesicles. FEMS Microbiol. Lett. 365:fny02. doi: 10.1093/femsle/fny023
Davies, D. G., Parsek, M. R., Pearson, J. P., Iglewski, B. H., Costerton, J. W., and Greenberg, E. (1998). The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 280, 295–298. doi: 10.1126/science.280.5361.295
de Viana, J. (1637). Tratado de Peste, sus Causas y curacion, y el Modo Que se ha tenido de curar las Secas y Carbuncos Pestilentes que Han Oprimido A Esta Ciudad de Málaga este año de 1637., ed. J. Serrano de Vargas y Ureña Málaga. Available online at: http://bdh.bne.es/bnesearch/biblioteca/Tratado de peste, sus causas y curacion, y el modo que se ha tenido de curar las secas y carbuncos pestilentes que han oprimido a esta ciudad de Malaga este año de 1637. /qls/Viana, Juan de/qls/bdh0000089890;jses (accessed April 5, 2020).
Deng, W., Puente, J. L., Gruenheid, S., Li, Y., Vallance, B. A., Vázquez, A., et al. (2004). Dissecting virulence: systematic and functional analyses of a pathogenicity island. Proc. Natl. Acad. Sci. U.S.A. 101, 3597–3602. doi: 10.1073/pnas.0400326101
Dersch, P., Khan, M. A., Mühlen, S., and Görke, B. (2017). Roles of regulatory RNAs for antibiotic resistance in bacteria and their potential value as novel drug targets. Front. Microbiol. 8:803. doi: 10.3389/fmicb.2017.00803
Diard, M., and Hardt, W.-D. (2017). Evolution of bacterial virulence. FEMS Microbiol. Rev. 41, 679–697. doi: 10.1093/femsre/fux023
Diaz-Ochoa, V. E., Jellbauer, S., Klaus, S., and Raffatellu, M. (2014). Transition metal ions at the crossroads of mucosal immunity and microbial pathogenesis. Front. Cell. Infect. Microbiol. 4:2. doi: 10.3389/fcimb.2014.00002
Djapgne, L., Panja, S., Brewer, L. K., Gans, J. H., Kane, M. A., Woodson, S. A., et al. (2018). The Pseudomonas aeruginosa PrrF1 and PrrF2 small regulatory RNAs promote 2-alkyl-4- quinolone production through redundant regulation of the antR mRNA. J. Bacteriol. 200:e0704-17. doi: 10.1128/JB.00704-717
Dong, H., Peng, X., Liu, Y., Wu, T., Wang, X., De, Y., et al. (2018). BASI74, a virulence-related sRNA in Brucella abortus. Front. Microbiol. 9:2173. doi: 10.3389/fmicb.2018.02173
Dong, T., and Schellhorn, H. E. (2010). Role of RpoS in virulence of pathogens. Infect. Immun. 78, 887–897. doi: 10.1128/IAI.00882-889
Dorey, A., Marinho, C., Piveteau, P., and O’Byrne, C. (2019). Role and regulation of the stress activated sigma factor sigma B (σ B) in the saprophytic and host-associated life stages of Listeria monocytogenes. Adv. Appl. Microbiol. 106, 1–48. doi: 10.1016/bs.aambs.2018.11.001
Dorman, M. J., and Dorman, C. J. (2018). Regulatory hierarchies controlling virulence gene expression in Shigella flexneri and Vibrio cholerae. Front. Microbiol. 9:2686. doi: 10.3389/fmicb.2018.02686
dos Santos, P. T., Menendez-Gil, P., Sabharwal, D., Christensen, J.-H., Brunhede, M. Z., Lillebæk, E. M. S., et al. (2018). The small regulatory RNAs LhrC1-5 contribute to the response of Listeria monocytogenes to heme toxicity. Front. Microbiol. 9:599. doi: 10.3389/fmicb.2018.00599
Drecktrah, D., Hall, L. S., Rescheneder, P., Lybecker, M., and Scott Samuels, D. (2018). The stringent response-regulated sRNA transcriptome of Borrelia burgdorferi. Front. Cell. Infect. Microbiol. 8:231. doi: 10.3389/fcimb.2018.00231
Drevinek, P., and Mahenthiralingam, E. (2010). Burkholderia cenocepacia in cystic fibrosis: epidemiology and molecular mechanisms of virulence. Clin. Microbiol. Infect. 16, 821–830. doi: 10.1111/j.1469-0691.2010.03237.x
Dubern, J. F., and Diggle, S. P. (2008). Quorum sensing by 2-alkyl-4-quinolones in Pseudomonas aeruginosa and other bacterial species. Mol. Biosyst. 4, 882–888. doi: 10.1039/b803796p
Dunman, P. M., Murphy, E., Haney, S., Palacios, D., Tucker-Kellogg, G., Wu, S., et al. (2001). Transcription profiling-based identification of Staphylococcus aureus genes regulated by the agr and/or sarA loci. J. Bacteriol. 183, 7341–7353. doi: 10.1128/JB.183.24.7341-7353.2001
Epstein, P. R. (2001). Climate change and emerging infectious diseases. Microb. Infect. 3, 747–754. doi: 10.1016/S1286-4579(01)01429-1420
Fàbrega, A., and Vila, J. (2013). Salmonella enterica serovar Typhimurium skills to succeed in the host: virulence and regulation. Clin. Microbiol. Rev. 26, 308–341. doi: 10.1128/CMR.00066-12
Fang, F. C., Libby, S. J., Buchmeier, N. A., Loewen, P. C., Switala, J., Harwood, J., et al. (1992). The alternative σ factor KatF (RpoS) regulates Salmonella virulence. Proc. Natl. Acad. Sci. U.S.A. 89, 11978–11982. doi: 10.1073/pnas.89.24.11978
Faucher, S. P., and Shuman, H. A. (2011). Small regulatory RNA and Legionella pneumophila. Front. Microbiol. 2:98. doi: 10.3389/fmicb.2011.00098
Felden, B., and Cattoira, V. (2018). Bacterial adaptation to antibiotics through regulatory RNAs. Antimicrob. Agents Chemother. 62:e002503-17. doi: 10.1128/AAC.02503-2517
Fernández, M., Corral-Lugo, A., and Krell, T. (2018). The plant compound rosmarinic acid induces a broad quorum sensing response in Pseudomonas aeruginosa PAO1. Environ. Microbiol. 20, 4230–4244. doi: 10.1111/1462-2920.14301
Ferrara, S., Brugnoli, M., de Bonis, A., Righetti, F., Delvillani, F., Dehò, G., et al. (2012). Comparative profiling of Pseudomonas aeruginosa strains reveals differential expression of novel unique and conserved small RNAs. PLoS One 7:36553. doi: 10.1371/journal.pone.0036553
Fischbach, M. A. (2009). Antibiotics from microbes: converging to kill. Curr. Opin. Microbiol. 12, 520–527. doi: 10.1016/j.mib.2009.07.002
Flahaut, S., Frere, J., Boutibonnes, P., and Auffray, Y. (1996). Comparison of the bile salts and sodium dodecyl sulfate stress responses in Enterococcus faecalis. Appl. Environ. Microbiol. 62, 2416–2420. doi: 10.1128/aem.62.7.2416-2420.1996
Frye, J., Karlinsey, J. E., Felise, H. R., Marzolf, B., Dowidar, N., McClelland, M., et al. (2006). Identification of new flagellar genes of Salmonella enterica serovar typhimurium. J. Bacteriol. 188, 2233–2243. doi: 10.1128/JB.188.6.2233-2243.2006
Fu, Y., Li, W., Wu, Z., Tao, Y., Wang, X., Wei, J., et al. (2018). Detection of mycobacterial small RNA in the bacterial culture supernatant and plasma of patients with active tuberculosis. Biochem. Biophys. Res. Commun. 503, 490–494. doi: 10.1016/j.bbrc.2018.04.165
Furniss, R. C. D., and Clements, A. (2018). Regulation of the locus of enterocyte effacement in attaching and effacing pathogens. J. Bacteriol. 200:e00336-17. doi: 10.1128/JB.00336-317
Gan, I.-N., and Tan, H. S. (2019). A small RNA decreases the sensitivity of Shigella sonnei to norfloxacin. BMC Res. Notes 12:97. doi: 10.1186/s13104-019-4124-4124
Gelsinger, D. R., and DiRuggiero, J. (2018). Transcriptional landscape and regulatory roles of small noncoding RNAs in the oxidative stress response of the haloarchaeon Haloferax volcanii. J. Bacteriol. 200:e00779-17. doi: 10.1128/JB.00779-717
Gelvin, S. B. (2003). Agrobacterium-mediated plant transformation: the biology behind the “gene-jockeying”. Tool. Microbiol. Mol. Biol. Rev. 67, 16–37. doi: 10.1128/mmbr.67.1.16-37.2003
Gerrick, E. R., Barbier, T., Chase, M. R., Xu, R., François, J., Lin, V. H., et al. (2018). Small RNA profiling in Mycobacterium tuberculosis identifies MrsI as necessary for an anticipatory iron sparing response. Proc. Natl. Acad. Sci. U.S.A. 115, 6464–6469. doi: 10.1073/pnas.1718003115
Ghaly, T. M., and Gillings, M. R. (2018). Mobile DNAs as ecologically and evolutionarily independent units of life. Trends Microbiol. 26, 904–912. doi: 10.1016/j.tim.2018.05.008
Ghosal, A. (2017). Importance of secreted bacterial RNA in bacterial-host interactions in the gut. Microb. Pathog. 104, 161–163. doi: 10.1016/j.micpath.2017.01.032
Gilbertson, S., Federspiel, J. D., Hartenian, E., Cristea, I. M., and Glaunsinger, B. (2018). Changes in mRNA abundance drive shuttling of RNA binding proteins, linking cytoplasmic RNA degradation to transcription. eLife 7:37663. doi: 10.7554/eLife.37663
Golding, B., Scott, D. E., Scharf, O., Huang, L. Y., Zaitseva, M., Lapham, C., et al. (2001). Immunity and protection against Brucella abortus. Microb. Infect. 3, 43–48. doi: 10.1016/S1286-4579(00)01350-1352
González Plaza, J. J. (2018). Small RNAs in cell-to-cell communications during bacterial infection. FEMS Microbiol. Lett. 365:fny024. doi: 10.1093/femsle/fny024
González Plaza, J. J., Hulak, N., Zhumadilov, Z., Akilzhanova, A., Plaza, J. J. G., Jj, G. P., et al. (2016). Fever as an important resource for infectious diseases research. Intractable Rare Dis. Res. 5, 97–102. doi: 10.5582/irdr.2016.01009
Grimson, A., Srivastava, M., Fahey, B., Woodcroft, B. J., Chiang, H. R., King, N., et al. (2008). Early origins and evolution of microRNAs and Piwi-interacting RNAs in animals. Nature 455, 1193–1198.
Guo, Z., Li, Y., and Ding, S.-W. (2019). Small RNA-based antimicrobial immunity. Nat. Rev. Immunol. 19, 31–44. doi: 10.1038/s41577-018-0071-x
Gutierrez, A., Laureti, L., Crussard, S., Abida, H., Rodríguez-Rojas, A., Blázquez, J., et al. (2013). β-lactam antibiotics promote bacterial mutagenesis via an RpoS-mediated reduction in replication fidelity. Nat. Commun. 4:2607. doi: 10.1038/ncomms2607
Hammer, B. K., and Bassler, B. L. (2007). Regulatory small RNAs circumvent the conventional quorum sensing pathway in pandemic Vibrio cholerae. Proc. Natl. Acad. Sci. U.S.A. 104, 11145–11149. doi: 10.1073/pnas.0703860104
Han, Y., Chen, D., Yan, Y., Gao, X., Liu, Z., Xue, Y., et al. (2019). Hfq globally binds and destabilizes sRNAs and mRNAs in Yersinia pestis. mSystems 4:245. doi: 10.1128/msystems.00245-219
Hansen, A. K., and Degnan, P. H. (2014). Widespread expression of conserved small RNAs in small symbiont genomes. ISME J. 8, 2490–2502. doi: 10.1038/ismej.2014.121
Harkey, C. W., Everiss, K. D., and Peterson, K. M. (1994). The Vibrio cholerae toxin-coregulated-pilus gene tcpI encodes a homolog of methyl-accepting chemotaxis proteins. Infect. Immun. 62, 2669–2678. doi: 10.1128/IAI.62.7.2669-2678.1994
Harvell, C. D., Mitchell, C. E., Ward, J. R., Altizer, S., Dobson, A. P., Ostfeld, R. S., et al. (2002). Climate warming and disease risks for terrestrial and marine biota. Science 296, 2158–2162. doi: 10.1126/science.1063699
Hendaus, M. A., Jomha, F. A., and Alhammadi, A. H. (2015). Virus-induced secondary bacterial infection: a concise review. Ther. Clin. Risk Manag. 11, 1265–1271. doi: 10.2147/TCRM.S87789
Herrera, J., and Nunn, C. L. (2019). Behavioural ecology and infectious disease: implications for conservation of biodiversity. Philos. Trans. R. Soc. B Biol. Sci. 374:54. doi: 10.1098/rstb.2018.0054
Higgs, P. G., and Lehman, N. (2015). The RNA world: molecular cooperation at the origins of life. Nat. Rev. Genet. 16, 7–17. doi: 10.1038/nrg3841
Hille, F., Richter, H., Wong, S. P., Bratoviè, M., Ressel, S., and Charpentier, E. (2018). The biology of CRISPR-Cas: backward and forward. Cell 172, 1239–1259. doi: 10.1016/j.cell.2017.11.032
Hoekzema, M., Romilly, C., Holmqvist, E., and Wagner, E. G. H. (2019). Hfq-dependent mRNA unfolding promotes sRNA -based inhibition of translation. EMBO J. 38:e101199. doi: 10.15252/embj.2018101199
Holmqvist, E., and Vogel, J. (2018). RNA-binding proteins in bacteria. Nat. Rev. Microbiol. 16, 601–615. doi: 10.1038/s41579-018-0049-45
Hornung, V., Barchet, W., Schlee, M., and Hartmann, G. (2008). “RNA Recognition via TLR7 and TLR8,” in Toll-Like Receptors (TLRs) and Innate Immunity. Handbook of Experimental Pharmacology, Vol. 183, eds S. Bauer and G. Hartmann, (Berlin: Springer), 71–86. doi: 10.1007/978-3-540-72167-3_4
How, M. J., Norman, M. D., Finn, J., Chung, W. S., and Marshall, N. J. (2017). Dynamic skin patterns in cephalopods. Front. Physiol. 8:393. doi: 10.3389/fphys.2017.00393
Hu, Y., Lu, P., Zhang, Y., Li, L., and Chen, S. (2010). Characterization of an aspartate-dependent acid survival system in Yersinia pseudotuberculosis. FEBS Lett. 584, 2311–2314. doi: 10.1016/j.febslet.2010.03.045
Hu, Y., Zhang, L., Wang, X., Sun, F., Kong, X., Dong, H., et al. (2018). Two virulent sRNAs identified by genomic sequencing target the type III secretion system in rice bacterial blight pathogen. BMC Plant Biol. 18:1477. doi: 10.1186/s12870-018-1470-1477
Hübner, A., Yang, X., Nolen, D. M., Popova, T. G., Cabello, F. C., and Norgard, M. V. (2001). Expression of Borrelia burgdorferi OspC and DbpA is controlled by a RpoN-RpoS regulatory pathway. Proc. Natl. Acad. Sci. U.S.A. 98, 12724–12729. doi: 10.1073/pnas.231442498
Hwang, H.-H., Yu, M., and Lai, E.-M. (2017). Agrobacterium -mediated plant transformation: biology and applications. Arab. B 15:e0186. doi: 10.1199/tab.0186
Ingle, R. A., Carstens, M., and Denby, K. J. (2006). PAMP recognition and the plant-pathogen arms race. Bioessays 28, 880–889. doi: 10.1002/bies.20457
Irving, S. E., and Corrigan, R. M. (2018). Triggering the stringent response: signals responsible for activating (p)ppGpp synthesis in bacteria. Microbiology 164, 268–276. doi: 10.1099/mic.0.000621
Islam, W., Noman, A., Qasim, M., and Wang, L. (2018). Plant responses to pathogen attack: small rnas in focus. Int. J. Mol. Sci. 19:515. doi: 10.3390/ijms19020515
Izoré, T., Job, V., and Dessen, A. (2011). Biogenesis, regulation, and targeting of the type III secretion system. Structure 19, 603–612. doi: 10.1016/j.str.2011.03.015
Jang, J., Jung, K. T., Park, J., Yoo, C. K., and Rhie, G. E. (2011). The Vibrio cholerae VarS/VarA two-component system controls the expression of virulence proteins through ToxT regulation. Microbiology 157, 1466–1473. doi: 10.1099/mic.0.043737-43730
Janssen, K. H., Diaz, M. R., Gode, C. J., Wolfgang, M. C., and Yahr, T. L. (2018a). RsmV, a small noncoding regulatory RNA in Pseudomonas aeruginosa that sequesters RsmA and RsmF from target mRNAs. J. Bacteriol. 200:e00277-18. doi: 10.1128/JB.00277-218
Janssen, K. H., Diaz, M. R., Golden, M., Graham, J. W., Sanders, W., Wolfgang, M. C., et al. (2018b). Functional analyses of the RsmY and RsmZ small noncoding regulatory RNAs in Pseudomonas aeruginosa. J. Bacteriol. 200:e0736-17. doi: 10.1128/JB.00736-17
Jemielita, M., Wingreen, N. S., and Bassler, B. L. (2018). Quorum sensing controls Vibrio cholerae multicellular aggregate formation. eLife 7:42057. doi: 10.7554/eLife.42057
Kalir, S., McClure, J., Pabbaraju, K., Southward, C., Ronen, M., Leibler, S., et al. (2001). Ordering genes in a flagella pathway by analysis of expression kinetics from living bacteria. Science 292, 2080–2083. doi: 10.1126/science.1058758
Kawai, T., and Akira, S. (2009). The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 21, 317–337. doi: 10.1093/intimm/dxp017
Kay, E., Humair, B., Dénervaud, V., Riedel, K., Spahr, S., Eberl, L., et al. (2006). Two GacA-dependent small RNAs modulate the quorum-sensing response in Pseudomonas aeruginosa. J. Bacteriol. 188, 6026–6033. doi: 10.1128/JB.00409-406
Khasnis, A. A., and Nettleman, M. D. (2005). Global warming and infectious disease. Arch. Med. Res. 36, 689–696. doi: 10.1016/j.arcmed.2005.03.041
Kiekens, S., Sass, A., Van Nieuwerburgh, F., Deforce, D., and Coenye, T. (2018). The Small RNA ncS35 regulates growth in Burkholderia cenocepacia J2315. mSphere 3:e0579-17. doi: 10.1128/msphere.00579-517
Kim, S., Reyes, D., Beaume, M., Francois, P., and Cheung, A. (2014). Contribution of teg49 Small RNA in the 5′ upstream transcriptional region of sarA to virulence in Staphylococcus aureus. Infect. Immun. 82, 4369–4379. doi: 10.1128/IAI.02002-2014
Kim, V. N., Han, J., and Siomi, M. C. (2009). Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. 10, 127–139.
Kirchman, D. L., Meon, B., Cottrell, M. T., Hutchins, D. A., Weeks, D., and Bruland, K. W. (2000). Carbon versus iron limitation of bacterial growth in the California upwelling regime. Limnol. Oceanogr. 45, 1681–1688. doi: 10.4319/lo.2000.45.8.1681
Kluger, M. J., Kozak, W., Conn, C. A., Leon, L. R., and Soszynski, D. (1998). Role of fever in disease. Ann. N. Y. Acad. Sci. 856, 224–233. doi: 10.1111/j.1749-6632.1998.tb08329.x
Krämer, A., Akmatov, M., and Kretzschmar, M. (2009). “Principles of infectious disease epidemiology,” in Modern Infectious Disease Epidemiology. Statistics for Biology and Health, eds A. Krämer, M. Kretzschmar, and K. Krickeberg, (New York, NY: Springer), 85–99. doi: 10.1007/978-0-387-93835-6_5
Kröger, C., Rothhardt, J. E., Brokatzky, D., Felsl, A., Kary, S. C., Heermann, R., et al. (2018). The small RNA RssR regulates myo-inositol degradation by Salmonella enterica. Sci. Rep. 8:17739. doi: 10.1038/s41598-018-35784-35788
Kuijl, C., and Neefjes, J. (2009). New insight into the everlasting host-pathogen arms race. Nat. Immunol. 10, 808–809. doi: 10.1038/ni0809-808
Lalaouna, D., Baude, J., Wu, Z., Tomasini, A., Chicher, J., Marzi, S., et al. (2019). RsaC sRNA modulates the oxidative stress response of Staphylococcus aureus during manganese starvation. Nucleic Acids Res. 47, 9871–9887. doi: 10.1093/nar/gkz728
Lalaouna, D., Eyraud, A., Chabelskaya, S., Felden, B., and Massé, E. (2014). Regulatory RNAs involved in bacterial antibiotic resistance. PLoS Pathog. 10:e04299. doi: 10.1371/journal.ppat.1004299
Lalaouna, D., Simoneau-Roy, M., Lafontaine, D., and Massé, E. (2013). Regulatory RNAs and target mRNA decay in prokaryotes. Biochim. Biophys. Acta Gene Regul. Mech. 1829, 742–747. doi: 10.1016/j.bbagrm.2013.02.013
Langdon, J. H (2016). “Case Study 1. The darwinian paradigm: an evolving world view,” in The Science of Human Evolution, (Cham: Springer), 1–8. doi: 10.1007/978-3-319-41585-7
Law, C. O. K., Huang, C., Pan, Q., Lee, J., Hao, Q., Chan, T. F., et al. (2019). A small RNA transforms the multidrug resistance of Pseudomonas aeruginosa to drug susceptibility. Mol. Ther. Nucleic Acids 16, 218–228. doi: 10.1016/j.omtn.2019.02.011
Lee, J., and Zhang, L. (2015). The hierarchy quorum sensing network in Pseudomonas aeruginosa. Protein Cell 6, 26–41. doi: 10.1007/s13238-014-0100-x
Lee, R. C., Feinbaum, R. L., and Ambros, V. (1993). The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 75, 843–854. doi: 10.1016/0092-8674(93)90529-y
Legüe, M., and Calixto, A. (2019). RNA language in Caenorhabditis elegans and bacteria interspecies communication and memory. Curr. Opin. Syst. Biol. 13, 16–22. doi: 10.1016/j.coisb.2018.08.005
Leid, J. G., Willson, C. J., Shirtliff, M. E., Hassett, D. J., Parsek, M. R., and Jeffers, A. K. (2005). The exopolysaccharide alginate protects Pseudomonas aeruginosa biofilm bacteria from IFN-γ-mediated macrophage killing. J. Immunol. 175, 7512–7518. doi: 10.4049/jimmunol.175.11.7512
Lejars, M., Kobayashi, A., and Hajnsdorf, E. (2019). Physiological roles of antisense RNAs in prokaryotes. Biochimie 164, 3–16. doi: 10.1016/j.biochi.2019.04.015
Lenz, D. H., Mok, K. C., Lilley, B. N., Kulkarni, R. V., Wingreen, N. S., and Bassler, B. L. (2004). The small RNA chaperone Hfq and multiple small RNAs control quorum sensing in Vibrio harveyi and Vibrio cholerae. Cell 118, 69–82. doi: 10.1016/j.cell.2004.06.009
Li, S., Xu, X., Zheng, Z., Zheng, J., Shakeel, M., and Jin, F. (2019). MicroRNA expression profiling of Plutella xylostella after challenge with B. thuringiensis. Dev. Comp. Immunol. 93, 115–124. doi: 10.1016/j.dci.2018.12.008
Liao, L., Liu, C., Zeng, Y., Zhao, B., Zhang, J., and Chen, B. (2019). Multipartite genomes and the sRNome in response to temperature stress of an Arctic Pseudoalteromonas fuliginea BSW20308. Environ. Microbiol. 21, 272–285. doi: 10.1111/1462-2920.14455
Lindsley, C. W. (2016). Lost in translation: the death of basic science. ACS Chem. Neurosci. 7, 1024–1024. doi: 10.1021/acschemneuro.6b00206
Liu, H., Wang, X., Wang, H. D., Wu, J., Ren, J., Meng, L., et al. (2012). Escherichia coli noncoding RNAs can affect gene expression and physiology of Caenorhabditis elegans. Nat. Commun. 3, 1–11. doi: 10.1038/ncomms2071
Liu, R., and Ochman, H. (2007). Stepwise formation of the bacterial flagellar system. Proc. Natl. Acad. Sci. U.S.A. 104, 7116–7121. doi: 10.1073/pnas.0700266104
Liu, X., and Matsumura, P. (1994). The FlhD/FlhC complex, a transcriptional activator of the Escherichia coli flagellar class II operons. J. Bacteriol. 176, 7345–7351. doi: 10.1128/jb.176.23.7345-7351.1994
Loh, E., Righetti, F., Eichner, H., Twittenhoff, C., and Narberhaus, F. (2018). RNA thermometers in bacterial pathogens. Microbiol. Spectr. 6:RWR-0012-2017. doi: 10.1128/microbiolspec.RWR-0012-2017
Lowy, F. D. (1998). Medical progress: Staphylococcus aureus infections. N. Engl. J. Med. 339, 520–532. doi: 10.1056/NEJM199808203390806
Lybecker, M. C., Abel, C. A., Feig, A. L., and Samuels, D. S. (2010). Identification and function of the RNA chaperone Hfq in the Lyme disease spirochete Borrelia burgdorferi. Mol. Microbiol. 78, 622–635. doi: 10.1111/j.1365-2958.2010.07374.x
Lybecker, M. C., and Samuels, D. S. (2007). Temperature-induced regulation of RpoS by a small RNA in Borrelia burgdorferi. Mol. Microbiol. 64, 1075–1089. doi: 10.1111/j.1365-2958.2007.05716.x
Lyu, L., Zhang, X., Li, C., Yang, T., Wang, J., Pan, L., et al. (2019). Small RNA profiles of serum exosomes derived from individuals with latent and active tuberculosis. Front. Microbiol. 10:1174. doi: 10.3389/fmicb.2019.01174
Lyu, Y., Wu, J., and Shi, Y. (2019). Metabolic and physiological perturbations of Escherichia coli W3100 by bacterial small RNA RyhB. Biochimie 162, 144–155. doi: 10.1016/j.biochi.2019.04.016
Mackowiak, P. A. (1981). Direct effects of hyperthermia on pathogenic microorganisms: teleologic implications with regard to fever. Rev. Infect. Dis. 3, 508–520. doi: 10.1093/CLINIDS/3.3.508
Mahenthiralingam, E., Urban, T. A., and Goldberg, J. B. (2005). The multifarious, multireplicon Burkholderia cepacia complex. Nat. Rev. Microbiol. 3, 144–156. doi: 10.1038/nrmicro1085
Mai, J., Rao, C., Watt, J., Sun, X., Lin, C., Zhang, L., et al. (2019). Mycobacterium tuberculosis 6C sRNA binds multiple mRNA targets via C-rich loops independent of RNA chaperones. Nucleic Acids Res. 47, 4292–4307. doi: 10.1093/nar/gkz149
Malgaonkar, A., and Nair, M. (2019). Quorum sensing in Pseudomonas aeruginosa mediated by RhlR is regulated by a small RNA PhrD. Sci. Rep. 9:89. doi: 10.1038/s41598-018-36488-36489
Mallia, P., and Johnston, S. L. (2007). Influenza infection and COPD. Int. J. COPD 2, 55–64. doi: 10.2147/copd.2007.2.1.55
Manna, A. C., Kim, S., Cengher, L., Corvaglia, A., Leo, S., Francois, P., et al. (2018). Small RNA teg49 is derived from a sarA transcript and regulates virulence genes independent of SarA in Staphylococcus aureus. Infect. Immun. 86:17. doi: 10.1128/IAI.00635-617
Marden, J. N., Diaz, M. R., Walton, W. G., Gode, C. J., Betts, L., Urbanowski, M. L., et al. (2013). An unusual CsrA family member operates in series with RsmA to amplify posttranscriptional responses in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U.S.A. 110, 15055–15060. doi: 10.1073/pnas.1307217110
Marinho, C. M., Dos Santos, P. T., Kallipolitis, B. H., Johansson, J., Ignatov, D., Guerreiro, D. N., et al. (2019). The σB-dependent regulatory sRNA Rli47 represses isoleucine biosynthesis in Listeria monocytogenes through a direct interaction with the ilvA transcript. RNA Biol. 16, 1424–1437. doi: 10.1080/15476286.2019.1632776
Massé, E., and Gottesman, S. (2002). A small RNA regulates the expression of genes involved in iron metabolism in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 99, 4620–4625. doi: 10.1073/pnas.032066599
Massé, E., Majdalani, N., and Gottesman, S. (2003). Regulatory roles for small RNAs in bacteria. Curr. Opin. Microbiol. 6, 120–124. doi: 10.1016/S1369-5274(03)00027-24
McPhearson, R. M., DePaola, A., Zywno, S. R., Motes, M. L., and Guarino, A. M. (1991). Antibiotic resistance in Gram-negative bacteria from cultured catfish and aquaculture ponds. Aquaculture 99, 203–211. doi: 10.1016/0044-8486(91)90241-X
Melson, E. M., and Kendall, M. M. (2019). The sRNA DicF integrates oxygen sensing to enhance enterohemorrhagic Escherichia coli virulence via distinctive RNA control mechanisms. Proc. Natl. Acad. Sci. U.S.A. 116, 14210–14215. doi: 10.1073/pnas.1902725116
Mey, A. R., Butz, H. A., and Payne, S. M. (2015). Vibrio cholerae CsrA regulates ToxR levels in response to amino acids and is essential for virulence. mBio 6:15. doi: 10.1128/mBio.01064-1015
Milillo, M. A., Trotta, A., Serafino, A., Marin Franco, J. L., Marinho, F. V., Alcain, J., et al. (2019). Bacterial RNA contributes to the down-modulation of MHC-II expression on monocytes/macrophages diminishing CD4+ T cell responses. Front. Immunol. 10:2181. doi: 10.3389/fimmu.2019.02181
Miller, C. L., Romero, M., Karna, S. L. R., Chen, T., Heeb, S., and Leung, K. P. (2016). RsmW, Pseudomonas aeruginosa small non-coding RsmA-binding RNA upregulated in biofilm versus planktonic growth conditions. BMC Microbiol. 16:155. doi: 10.1186/s12866-016-0771-y
Moresco, E. M. Y., LaVine, D., and Beutler, B. (2011). Toll-like receptors. Curr. Biol. 21:39. doi: 10.1016/j.cub.2011.05.039
Mustachio, L. M., Aksit, S., Mistry, R. H., Scheffler, R., Yamada, A., and Liu, J. M. (2012). The Vibrio cholerae mannitol transporter is regulated posttranscriptionally by the MtlS small regulatory RNA. J. Bacteriol. 194, 598–606. doi: 10.1128/JB.06153-6111
Nadal Jimenez, P., Koch, G., Thompson, J. A., Xavier, K. B., Cool, R. H., and Quax, W. J. (2012). The multiple signaling systems regulating virulence in Pseudomonas aeruginosa. Microbiol. Mol. Biol. Rev. 76, 46–65. doi: 10.1128/mmbr.05007-5011
Nakatsu, Y., Matsui, H., Yamamoto, M., Noutoshi, Y., Toyoda, K., and Ichinose, Y. (2019). Quorum-dependent expression of rsmX and rsmY, small non-coding RNAs, in Pseudomonas syringae. Microbiol. Res. 22, 72–78. doi: 10.1016/j.micres.2019.04.004
Napoli, C., Lemieux, C., and Jorgensen, R. (1990). Introduction of a chimeric chalcone synthase gene into petunia results in reversible co-suppression of homologous genes in trans. Plant Cell 2, 279–289. doi: 10.1105/tpc.2.4.279
Narberhaus, F. (2010). Translational control of bacterial heat shock and virulence genes by temperature-sensing mRNAs. RNA Biol. 7:501. doi: 10.4161/rna.7.1.10501
Narberhaus, F., Waldminghaus, T., and Chowdhury, S. (2006). RNA thermometers. FEMS Microbiol. Rev. 30, 3–16. doi: 10.1111/j.1574-6976.2005.004.x
Ng, D., Harn, T., Altindal, T., Kolappan, S., Marles, J. M., Lala, R., et al. (2016). The Vibrio cholerae minor pilin TcpB initiates assembly and retraction of the toxin-Coregulated Pilus. PLoS Pathog. 12:e1006109. doi: 10.1371/journal.ppat.1006109
Oh, C. S., and Beer, S. V. (2005). Molecular genetics of Erwinia amylovora involved in the development of fire blight. FEMS Microbiol. Lett. 253, 185–192. doi: 10.1016/j.femsle.2005.09.051
Okamura, K., and Lai, E. C. (2008). Endogenous small interfering RNAs in animals. Nat. Rev. Mol. Cell Biol. 9, 673–678. doi: 10.1038/nrm2479
Olejniczak, M., and Storz, G. (2017). ProQ/FinO-domain proteins: another ubiquitous family of RNA matchmakers? Mol. Microbiol. 104, 905–915. doi: 10.1111/mmi.13679
O’Neill, J. (2014). The review on antimicrobial resistance tackling drug-resistant infections globally: final report and recommendations. U. K. Rev. Antimicrob. Resist. 80. Available online at: https://wellcomecollection.org/works/thvwsuba
Ong, S. T., Shan, Ho, J. Z., Ho, B., and Ding, J. L. (2006). Iron-withholding strategy in innate immunity. Immunobiology 211, 295–314. doi: 10.1016/j.imbio.2006.02.004
Ostberg, J. R., Taylor, S. L., Baumann, H., and Repasky, E. A. (2000). Regulatory effects of fever-range whole-body hyperthermia on the LPS-induced acute inflammatory response. J. Leukoc. Biol. 68, 815–820. doi: 10.1189/jlb.68.6.815
Pallen, M. J., Beatson, S. A., and Bailey, C. M. (2005). Bioinformatics analysis of the locus for enterocyte effacement provides novel insights into type-III secretion. BMC Microbiol. 5:9. doi: 10.1186/1471-2180-5-9
Papenfort, K., and Vogel, J. (2010). Regulatory RNA in bacterial pathogens. Cell Host Microb. 8, 116–127. doi: 10.1016/j.chom.2010.06.008
Pena, R. T., Blasco, L., Ambroa, A., González-Pedrajo, B., Fernández-García, L., López, M., et al. (2019). Relationship between quorum sensing and secretion systems. Front. Microbiol. 10:1100. doi: 10.3389/fmicb.2019.01100
Pita, T., Feliciano, J. R., and Leitão, J. H. (2018). Small noncoding regulatory RNAs from Pseudomonas aeruginosa and Burkholderia cepacia complex. Int. J. Mol. Sci. 19:3759. doi: 10.3390/ijms19123759
Poole, K. (2012). Bacterial stress responses as determinants of antimicrobial resistance. J. Antimicrob. Chemother. 67, 2069–2089. doi: 10.1093/jac/dks196
Popitsch, N., Bilusic, I., Rescheneder, P., Schroeder, R., and Lybecker, M. (2017). Temperature-dependent sRNA transcriptome of the Lyme disease spirochete. BMC Genomics 18:28. doi: 10.1186/s12864-016-3398-3393
Pounds, J. A., Bustamante, M. R., Coloma, L. A., Consuegra, J. A., Fogden, M. P. L., Foster, P. N., et al. (2006). Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 439, 161–167. doi: 10.1038/nature04246
Puzari, M., Sharma, M., and Chetia, P. (2018). Emergence of antibiotic resistant Shigella species: a matter of concern. J. Infect. Public Health 11, 451–454. doi: 10.1016/j.jiph.2017.09.025
Radolf, J. D., Caimano, M. J., Stevenson, B., and Hu, L. T. (2012). Of ticks, mice and men: Understanding the dual-host lifestyle of Lyme disease spirochaetes. Nat. Rev. Microbiol. 10, 87–99. doi: 10.1038/nrmicro2714
Ramamoorthi, N., Narasimhan, S., Pal, U., Bao, F., Yang, X. F., Fish, D., et al. (2005). The Lyme disease agent exploits a tick protein to infect the mammalian host. Nature 436, 573–577. doi: 10.1038/nature03812
Ratner, H. K., Escalera-Maurer, A., Le Rhun, A., Jaggavarapu, S., Wozniak, J. E., Crispell, E. K., et al. (2019). Catalytically Active Cas9 mediates transcriptional interference to facilitate bacterial virulence. Mol. Cell 75, 498–510. doi: 10.1016/j.molcel.2019.05.029
Records, A. R., and Gross, D. C. (2010). Sensor kinases RetS and LadS regulate Pseudomonas syringae Type VI secretion and virulence factors. J. Bacteriol. 192, 3584–3596. doi: 10.1128/JB.00114-110
Rochat, T., Bohn, C., Morvan, C., Le Lam, T. N., Razvi, F., Pain, A., et al. (2018). The conserved regulatory RNA RsaE down-regulates the arginine degradation pathway in Staphylococcus aureus. Nucleic Acids Res. 46, 8803–8816. doi: 10.1093/nar/gky584
Rodó, X., Pascual, M., Fuchs, G., and Faruque, A. S. G. (2002). ENSO and cholera: a nonstationary link related to climate change? Proc. Natl. Acad. Sci. U.S.A. 99, 12901–12906. doi: 10.1073/pnas.182203999
Rohmer, L., Hocquet, D., and Miller, S. I. (2011). Are pathogenic bacteria just looking for food? metabolism and microbial pathogenesis. Trends Microbiol. 19, 341–348. doi: 10.1016/j.tim.2011.04.003
Romeo, T., and Babitzke, P. (2018). Global regulation by CsrA and Its RNA antagonists. Microbiol. Spectr. 6, 1321–1330. doi: 10.1128/microbiolspec.rwr-0009-2017
Romero, D., Traxler, M. F., López, D., and Kolter, R. (2011). Antibiotics as signal molecules. Chem. Rev. 111, 5492–5505. doi: 10.1021/cr2000509
Romero, M., Silistre, H., Lovelock, L., Wright, V. J., Chan, K.-G., Hong, K.-W., et al. (2018). Genome-wide mapping of the RNA targets of the Pseudomonas aeruginosa riboregulatory protein RsmN. Nucleic Acids Res. 46, 6823–6840. doi: 10.1093/nar/gky324
Ross, J. A., Thorsing, M., Lillebæk, E. M. S., Teixeira Dos Santos, P., and Kallipolitis, B. H. (2019). The LhrC sRNAs control expression of T cell-stimulating antigen TcsA in Listeria monocytogenes by decreasing tcsA mRNA stability. RNA Biol. 16, 270–281. doi: 10.1080/15476286.2019.1572423
Roy, B., Ahamed, S. T., Bandyopadhay, B., and Giri, N. (2020). Development of quinolone resistance and prevalence of different virulence genes among Shigella flexneri and Shigella dysenteriae in environmental water samples. Lett. Appl. Microbiol. 13262. doi: 10.1111/lam.13262
Ruano-Gallego, D., Álvarez, B., and Fernández, L. Á (2015). Engineering the controlled assembly of filamentous injectisomes in E. coli K-12 for protein translocation into mammalian cells. ACS Synth. Biol. 4, 1030–1041. doi: 10.1021/acssynbio.5b00080
Rübsam, H., Kirsch, F., Reimann, V., Erban, A., Kopka, J., Hagemann, M., et al. (2018). The iron-stress activated RNA 1 (IsaR1) coordinates osmotic acclimation and iron starvation responses in the cyanobacterium Synechocystis sp. PCC 6803. Environ. Microbiol. 20, 2757–2768. doi: 10.1111/1462-2920.14079
Sahni, A., Hajjari, M., Raheb, J., Foroughmand, A. M., and Asgari, M. (2019). The non-coding RNA rprA can increase the resistance to ampicillin in Escherichia coli. Microb. Pathog. 129, 266–270. doi: 10.1016/j.micpath.2019.02.021
Saliba, A. E., Santos, C., and Vogel, J. (2017). New RNA-seq approaches for the study of bacterial pathogens. Curr. Opin. Microbiol. 35, 78–87. doi: 10.1016/j.mib.2017.01.001
Sanderson, S., Campbell, D. J., and Shastri, N. (1995). Identification of a CD4+ T cell-stimulating antigen of pathogenic bacteria by expression cloning. J. Exp. Med. 182, 1751–1757. doi: 10.1084/jem.182.6.1751
Santiago-Frangos, A., Fröhlich, K. S., Jeliazkov, J. R., Małecka, E. M., Marino, G., Gray, J. J., et al. (2019). Caulobacter crescentus Hfq structure reveals a conserved mechanism of RNA annealing regulation. Proc. Natl. Acad. Sci. U.S.A. 166, 10978–10987. doi: 10.1073/pnas.1814428116
Sassi, M., Augagneur, Y., Mauro, T., Ivain, L., Chabelskaya, S., Hallier, M., et al. (2015). SRD: A Staphylococcus regulatory RNA database. RNA 21, 1005–1017. doi: 10.1261/rna.049346.114
Sauder, A. B., and Kendall, M. M. (2018). After the fact(or): posttranscriptional gene regulation in enterohemorrhagic Escherichia coli O157:H7. J. Bacteriol. 200:e00228-18. doi: 10.1128/JB.00228-218
Schachterle, J. K., Zeng, Q., and Sundin, G. W. (2019). Three Hfq-dependent small RNAs regulate flagellar motility in the fire blight pathogen Erwinia amylovora. Mol. Microbiol. 111, 1476–1492. doi: 10.1111/mmi.14232
Schuster, M., Hawkins, A. C., Harwood, C. S., and Greenberg, E. P. (2004). The Pseudomonas aeruginosa RpoS regulon and its relationship to quorum sensing. Mol. Microbiol. 51, 973–985. doi: 10.1046/j.1365-2958.2003.03886.x
Segovia, C., Arias-Carrasco, R., Yañez, A. J., Maracaja-Coutinho, V., and Santander, J. (2018). Core non-coding RNAs of Piscirickettsia salmonis. PLoS One 13:e0197206. doi: 10.1371/journal.pone.0197206
Sellge, G., and Kufer, T. A. (2015). PRR-signaling pathways: learning from microbial tactics. Semin. Immunol. 27, 75–84. doi: 10.1016/j.smim.2015.03.009
Sheehan, L. M., and Caswell, C. C. (2018). An account of evolutionary specialization: the AbcR small RNAs in the rhizobiales. Mol. Microbiol. 107, 24–33. doi: 10.1111/mmi.13869
Shimizu, K. (2013). Regulation systems of bacteria such as Escherichia coli in response to nutrient limitation and environmental stresses. Metabolites 4, 1–35. doi: 10.3390/metabo4010001
Shin, G. Y., Schachterle, J. K., Shyntum, D. Y., Moleleki, L. N., Coutinho, T. A., and Sundin, G. W. (2019). Functional characterization of a global virulence regulator Hfq and identification of Hfq-dependent sRNAs in the plant pathogen Pantoea ananatis. Front. Microbiol. 10:2075. doi: 10.3389/fmicb.2019.02075
Shyp, V., Tankov, S., Ermakov, A., Kudrin, P., English, B. P., Ehrenberg, M., et al. (2012). Positive allosteric feedback regulation of the stringent response enzyme RelA by its product. EMBO Rep. 13, 835–839. doi: 10.1038/embor.2012.106
Silva, I. J., Barahona, S., Eyraud, A., Lalaouna, D., Figueroa-Bossi, N., Massé, E., et al. (2019). SraL sRNA interaction regulates the terminator by preventing premature transcription termination of rho mRNA. Proc. Natl. Acad. Sci. U.S.A. 116, 3042–3051. doi: 10.1073/pnas.1811589116
Sinha, D., Matz, L. M., Cameron, T. A., and de Lay, N. R. (2018). Poly(A) polymerase is required for RyhB sRNA stability and function in Escherichia coli. RNA 24, 1496–1511. doi: 10.1261/rna.067181.118
Slingenbergh, J., Gilbert, M., De Balogh, K., and Wint, W. (2004). Ecological sources of zoonotic diseases. OIE Rev. Sci. Tech. 23, 467–484. doi: 10.20506/rst.23.2.1492
Smirnov, A., Förstner, K. U., Holmqvist, E., Otto, A., Günster, R., Becher, D., et al. (2016). Grad-seq guides the discovery of ProQ as a major small RNA-binding protein. Proc. Natl. Acad. Sci. U.S.A. 113, 11591–11596. doi: 10.1073/pnas.1609981113
Sonnleitner, E., Gonzalez, N., Sorger-Domenigg, T., Heeb, S., Richter, A. S., Backofen, R., et al. (2011). The small RNA PhrS stimulates synthesis of the Pseudomonas aeruginosa quinolone signal. Mol. Microbiol. 80, 868–885. doi: 10.1111/j.1365-2958.2011.07620.x
Sparling, P. F. (1983). Bacterial virulence and pathogenesis: an overview. Rev. Infect. Dis. 5, 637–646.
Steere, A. C., Strle, F., Wormser, G. P., Hu, L. T., Branda, J. A., Hovius, J. W. R., et al. (2016). Lyme borreliosis. Nat. Rev. Dis. Prim. 2, 1–19. doi: 10.1038/nrdp.2016.90
Streinzer, M., Paulus, H. F., and Spaethe, J. (2009). Floral colour signal increases short-range detectability of a sexually deceptive orchid to its bee pollinator. J. Exp. Biol. 212, 1365–1370. doi: 10.1242/jeb.027482
Sudo, N., Soma, A., Iyoda, S., Oshima, T., Ohto, Y., Saito, K., et al. (2018). Small RNA Esr41 inversely regulates expression of LEE and flagellar genes in enterohaemorrhagic Escherichia coli. Microbiology 164, 821–834. doi: 10.1099/mic.0.000652
Sudo, N., Soma, A., Muto, A., Iyoda, S., Suh, M., Kurihara, N., et al. (2014). A novel small regulatory RNA enhances cell motility in enterohemorrhagic Escherichia coli. J. Gen. Appl. Microbiol. 60, 44–50. doi: 10.2323/jgam.60.44
Suh, S. J., Silo-Suh, L., Woods, D. E., Hassett, D. J., West, S. E. H., and Ohman, D. E. (1999). Effect of rpoS mutation on the stress response and expression of virulence factors in Pseudomonas aeruginosa. J. Bacteriol. 181, 3890–3897. doi: 10.1128/jb.181.13.3890-3897.1999
Sundberg, L. R., Ketola, T., Laanto, E., Kinnula, H., Bamford, J. K. H., Penttinen, R., et al. (2016). Intensive aquaculture selects for increased virulence and interference competition in bacteria. Proc. R. Soc. B Biol. Sci. 283:3069. doi: 10.1098/rspb.2015.3069
Swaminathan, B., and Gerner-Smidt, P. (2007). The epidemiology of human listeriosis. Microb. Infect. 9, 1236–1243. doi: 10.1016/j.micinf.2007.05.011
Tanji, T., and Ip, Y. T. (2005). Regulators of the toll and Imd pathways in the drosophila innate immune response. Trends Immunol. 26, 193–198. doi: 10.1016/j.it.2005.02.006
Taylor, A. F. (2016). Small molecular replicators go organic. Nature 537, 627–628. doi: 10.1038/537627a
Thairu, M. W., Cheng, S., and Hansen, A. K. (2018). A sRNA in a reduced Mutualistic symbiont genome regulates its own gene expression. Mol. Ecol. 27, 1766–1776. doi: 10.1111/mec.14424
Thairu, M. W., and Hansen, A. K. (2019). Changes in aphid host plant diet influence the small-RNA expression profiles of its obligate nutritional symbiont, Buchnera. mBio 10:e01733-19. doi: 10.1128/mBio.01733-1719
Thi Bach Nguyen, H., Romero, A. D., Amman, F., Sorger-Domenigg, T., and Tata, M. (2018). Negative control of RpoS synthesis by the sRNA ReaL in Pseudomonas aeruginosa. Front. Microbiol. 9:2488. doi: 10.3389/fmicb.2018.02488
Thomason, M. K., Voichek, M., Dar, D., Addis, V., Fitzgerald, D., Gottesman, S., et al. (2019). A rhlI 5′ UTR-Derived sRNA regulates RhlR-dependent quorum sensing in Pseudomonas aeruginosa. mBio 10:e02253-19. doi: 10.1128/mBio.02253-2219
Tomasini, A., François, P., Howden, B. P., Fechter, P., Romby, P., and Caldelari, I. (2014). The importance of regulatory RNAs in Staphylococcus aureus. Infect. Genet. Evol. 21, 616–626. doi: 10.1016/j.meegid.2013.11.016
Toyofuku, M., Nomura, N., and Eberl, L. (2019). Types and origins of bacterial membrane vesicles. Nat. Rev. Microbiol. 17, 13–24. doi: 10.1038/s41579-018-0112-112
Trewavas, A. J. (2001). The population/biodiversity paradox. Agricultural efficiency to save wilderness. Plant Physiol. 125, 174–179. doi: 10.1104/pp.125.1.174
Trullàs, R. (2019). El Fracaso De La Ciencia Con Adjetivos, Por Ramon Trullàs. El Periódico. Available online at: https://www.elperiodico.com/es/opinion/20180201/el-fracaso-de-la-ciencia-con-adjetivos-articulo-ramon-trullas-6594038 (accessed December 23, 2019).
Valentini, M., Gonzalez, D., Mavridou, D. A., and Filloux, A. (2018). Lifestyle transitions and adaptive pathogenesis of Pseudomonas aeruginosa. Curr. Opin. Microbiol. 41, 15–20. doi: 10.1016/j.mib.2017.11.006
van der Krol, A. R., Mur, L. A., Beld, M., Mol, J. N., and Stuitje, A. R. (1990). Flavonoid genes in petunia: addition of a limited number of gene copies may lead to a suppression of gene expression. Plant Cell 2, 291–299. doi: 10.1105/tpc.2.4.291
Van Elsas, J. D., Semenov, A. V., Costa, R., and Trevors, J. T. (2011). Survival of Escherichia coli in the environment: fundamental and public health aspects. ISME J. 5, 173–183. doi: 10.1038/ismej.2010.80
Wassenaar, T. M., and Gaastra, W. (2001). Bacterial virulence: can we draw the line? FEMS Microbiol. Lett. 201, 1–7. doi: 10.1111/j.1574-6968.2001.tb10724.x
Waters, L. S., and Storz, G. (2009). Regulatory RNAs in bacteria. Cell 136, 615–628. doi: 10.1016/j.cell.2009.01.043
Wei, B. L., Brun-Zinkernagel, A. M., Simecka, J. W., Prüß, B. M., Babitzke, P., and Romeo, T. (2001). Positive regulation of motility and flhDC expression by the RNA-binding protein CsrA of Escherichia coli. Mol. Microbiol. 40, 245–256. doi: 10.1046/j.1365-2958.2001.02380.x
Weiberg, A., Wang, M., Lin, F. M., Zhao, H., Zhang, Z., Kaloshian, I., et al. (2013). Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science 342, 118–123. doi: 10.1126/science.1239705
Wernegreen, J. J. (2002). Genome evolution in bacterial endosymbionts of insects. Nat. Rev. Genet. 3, 850–861. doi: 10.1038/nrg931
Werren, J. H., Baldo, L., and Clark, M. E. (2008). Wolbachia: master manipulators of invertebrate biology. Nat. Rev. Microbiol. 6, 741–751. doi: 10.1038/nrmicro1969
Westermann, A. J., Barquist, L., and Vogel, J. (2017). Resolving host-pathogen interactions by dual RNA-seq. PLoS Pathog. 13:e06033. doi: 10.1371/journal.ppat.1006033
Westermann, A. J., Förstner, K. U., Amman, F., Barquist, L., Chao, Y., Schulte, L. N., et al. (2016). Dual RNA-seq unveils noncoding RNA functions in host-pathogen interactions. Nature 529, 496–501. doi: 10.1038/nature16547
Westermann, A. J., Gorski, S. A., and Vogel, J. (2012). Dual RNA-seq of pathogen and host. Nat. Rev. Microbiol. 10, 618–630. doi: 10.1038/nrmicro2852
Westermann, A. J., Venturini, E., Sellin, M. E., Förstner, K. U., Hardt, W. D., and Vogel, J. (2019). The major RNA-binding protein ProQ impacts virulence gene expression in Salmonella enterica serovar Typhimurium. mBio 10:e002504-18. doi: 10.1128/mBio.02504-2518
Wightman, B., Ha, I., and Ruvkun, G. (1993). Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 75, 855–862. doi: 10.1016/0092-8674(93)90530-4
Wilderman, P. J., Sowa, N. A., FitzGerald, D. J., FitzGerald, P. C., Gottesman, S., Ochsner, U. A., et al. (2004). Identification of tandem duplicate regulatory small RNAs in Pseudomonas aeruginosa involved in iron homeostasis. Proc. Natl. Acad. Sci. U.S.A. 101, 9792–9797. doi: 10.1073/pnas.0403423101
Wilms, I., Voss, B., Hess, W. R., Leichert, L. I., and Narberhaus, F. (2011). Small RNA-mediated control of the Agrobacterium tumefaciens GABA binding protein. Mol. Microbiol. 80, 492–506. doi: 10.1111/j.1365-2958.2011.07589.x
Witzany, G. (2010). Biocommunication and natural genome editing. World J. Biol. Chem. 1:348. doi: 10.4331/wjbc.v1.i11.348
Woolfit, M., Algama, M., Keith, J. M., McGraw, E. A., and Popovici, J. (2015). Discovery of putative small non-coding RNAs from the obligate intracellular bacterium Wolbachia pipientis. PLoS One 10:e0118595. doi: 10.1371/journal.pone.0118595
Xi, D., Li, Y., Yan, J., Li, Y., Wang, X., and Cao, B. (2020). Small RNA coaR contributes to intestinal colonization in Vibrio cholerae via the two-component system EnvZ/OmpR. Environ. Microbiol. 1462. doi: 10.1111/1462-2920.14906
Yakhnin, A. V., Baker, C. S., Vakulskas, C. A., Yakhnin, H., Berezin, I., Romeo, T., et al. (2013). CsrA activates flhDC expression by protecting flhDC mRNA from RNase E-mediated cleavage. Mol. Microbiol. 87, 851–866. doi: 10.1111/mmi.12136
Yuan, X., Zeng, Q., Khokhani, D., Tian, F., Severin, G. B., Waters, C. M., et al. (2019). A feed-forward signalling circuit controls bacterial virulence through linking cyclic di-GMP and two mechanistically distinct sRNAs, ArcZ and RsmB. Environ. Microbiol. 21, 2755–2771. doi: 10.1111/1462-2920.14603
Zapf, R. L., Wiemels, R. E., Keogh, R. A., Holzschu, D. L., Howell, K. M., Trzeciak, E., et al. (2019). The small RNA Teg41 regulates expression of the alpha phenol-soluble modulins and is required for virulence in Staphylococcus aureus. mBio 10:e02484-18. doi: 10.1128/mBio.02484-2418
Zeng, G., Yang, J., and Zhu, G. (2019). Cross-kingdom small RNAs among animals, plants and microbes. Cells 8:371. doi: 10.3390/cells8040371
Zeng, Q., and Sundin, G. W. (2014). Genome-wide identification of Hfq-regulated small RNAs in the fire blight pathogen Erwinia amylovora discovered small RNAs with virulence regulatory function. BMC Genom. 15:414. doi: 10.1186/1471-2164-15-414
Zhang, M. G., and Liu, J. M. (2019). Transcription of cis antisense small RNA MtlS in Vibrio cholerae is regulated by transcription of its target gene, mtlA. J. Bacteriol. 201:e00178-19. doi: 10.1128/JB.00178-119
Keywords: small RNA, bacteria, infectious disease, information transfer, host-pathogen interaction
Citation: González Plaza JJ (2020) Small RNAs as Fundamental Players in the Transference of Information During Bacterial Infectious Diseases. Front. Mol. Biosci. 7:101. doi: 10.3389/fmolb.2020.00101
Received: 01 January 2020; Accepted: 04 May 2020;
Published: 16 June 2020.
Edited by:
Olga N. Ozoline, Institute of Cell Biophysics (RAS), RussiaReviewed by:
Claudio Valverde, National University of Quilmes, ArgentinaAlfredo Berzal-Herranz, Instituto de Parasitología y Biomedicina López-Neyra, Spain
Copyright © 2020 González Plaza. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juan José González Plaza, gonzalez_plaza@fld.czu.cz; gonzalezplazajuanjose@gmail.com