Profiling of Volatile Organic Compounds in Wild Indigenous Medicinal Ginger (Zingiber barbatum Wall.) from Myanmar

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. The VOC Profile in the Headspace of Six Z. barbatum Rhizomes
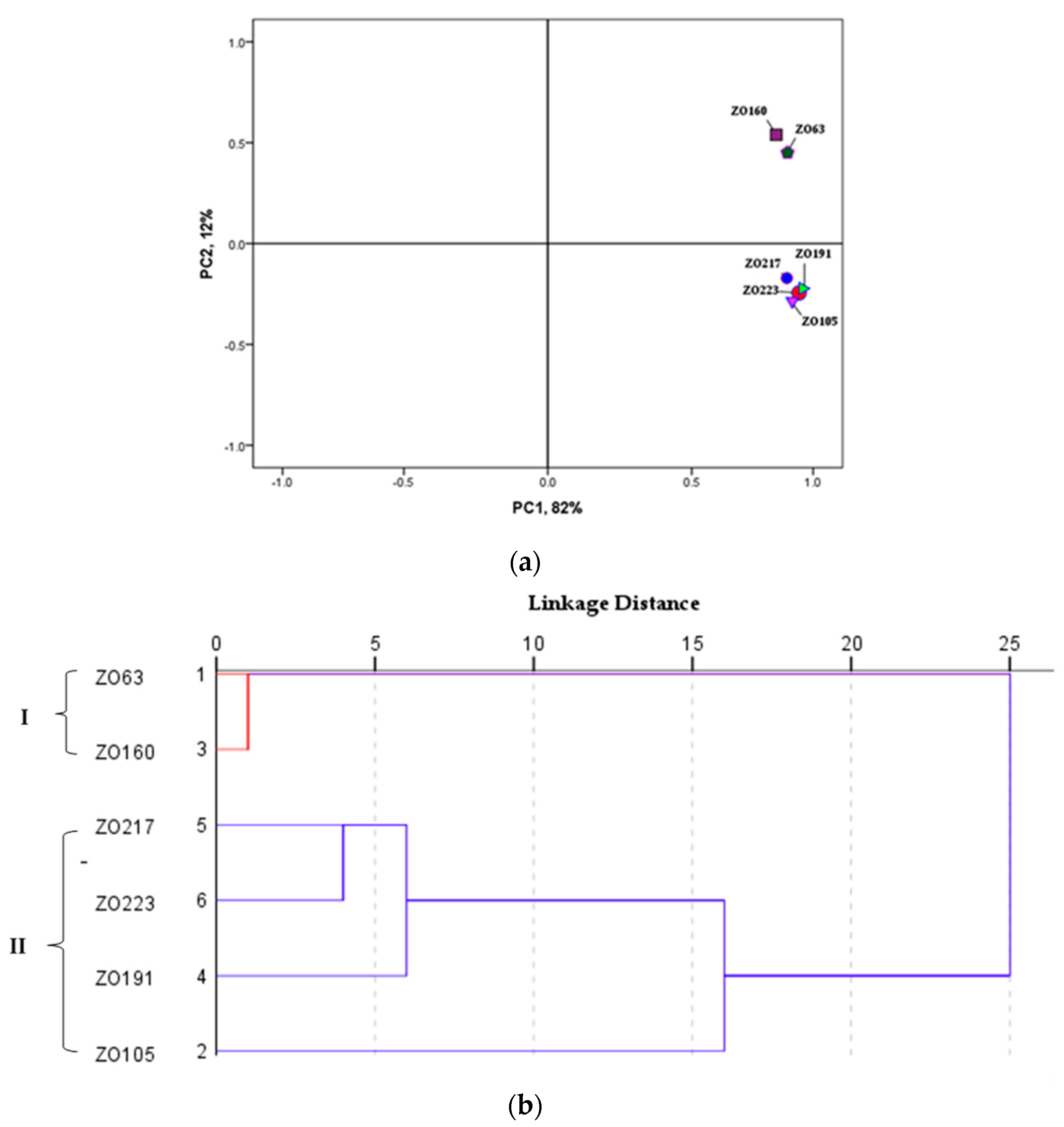
2.2. Multivariate Data Analysis (MVDA) of Omics Data
2.2.1. Principal Component Analysis (PCA) and Orthogonal Partial Least Square Projection to Latent Structures Differential Analysis (OPLS-DA)
2.2.2. Principal Components Analysis (PCA) and Hierarchical Cluster Analysis (HCA)
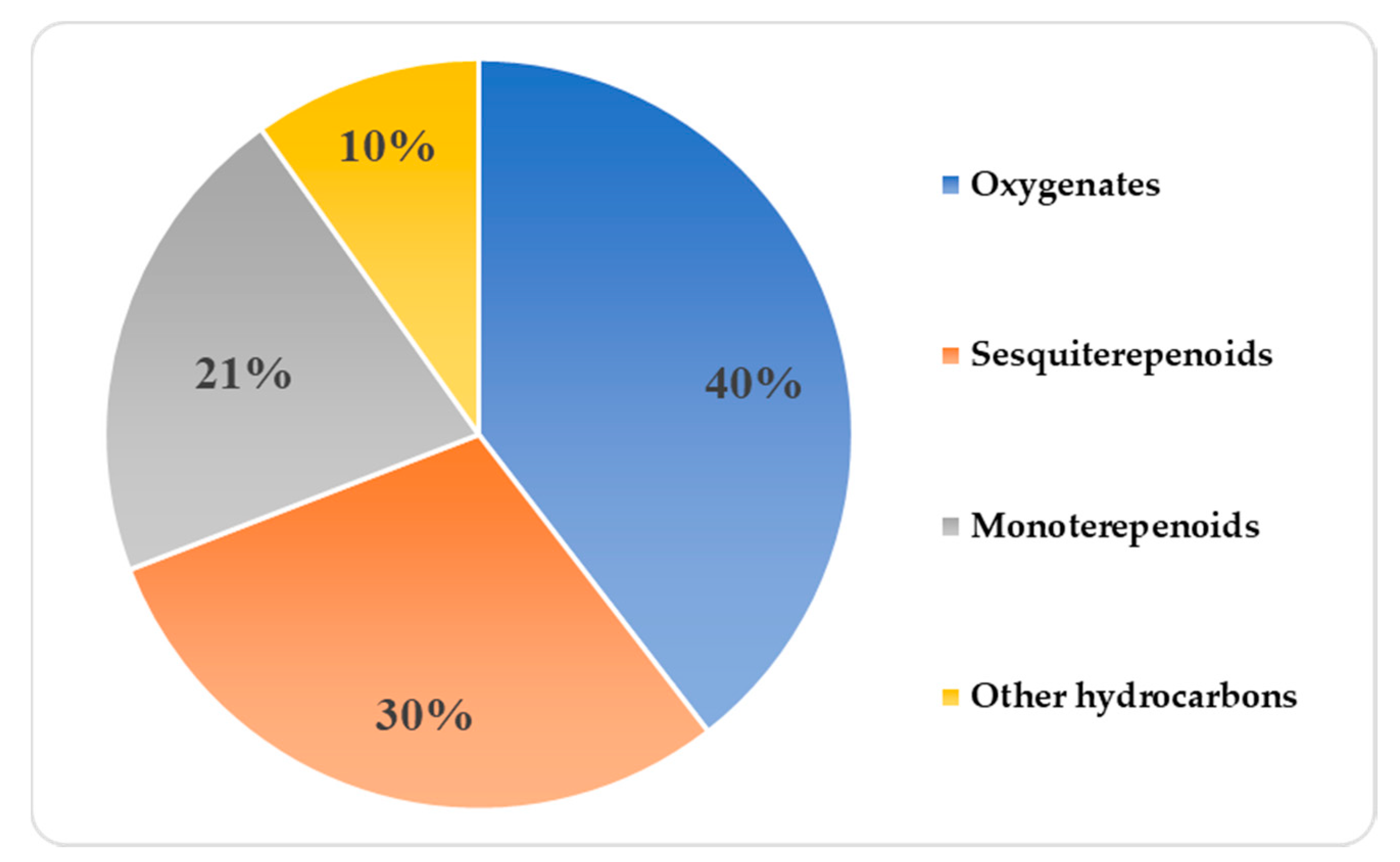
2.3. Composition of VOCs in the VOC Profile of Z. barbatum Samples
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.1.1. Sample Selection
4.1.2. Experimental Design, Sample Collection, and Sample Preparation
4.2. Sample Processing
4.2.1. Homogenization and Aliquot Preparation
4.2.2. Chemicals
4.2.3. VOC Extraction and Headspace Collection by a SPME Fiber
4.3. GC-TOF-MS Analysis
4.4. Data Processing and Provisional Identification of VOCs in HS Samples of Z. barbatum Species
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Species | Accession (GRC UT) | Status | Collection Site | Altitude (m a.s.l.) | Year of Acquisition by SMTA | |
|---|---|---|---|---|---|---|
| City | Region/State | |||||
| Z. barbatum | ZO 63 | Landrace | Nattalin | Bago | 4 | 2004 |
| Z. barbatum | ZO 105 | Landrace | Pyon oo lwin | Mandalay | 1070 | 2004 |
| Z. barbatum | ZO 160 | Landrace | Thayarwaddy | Bago | 15 | 2007 |
| Z. barbatum | ZO 191 | Landrace | Pin Da Ya | Shan | 1164 | 2008 |
| Z. barbatum | ZO 217 | Landrace | Aung Ban | Shan | 1286 | 2009 |
| Z. barbatum | ZO 223 | Landrace | Kyauk Pa Daung | Mandalay | 595 | 2009 |
References
- Conrath, U.; Pieterse, C.M.J.; Mauch-Mani, B. Priming in plant-pathogen interactions. Trends Plant Sci. 2002, 7, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, I.T.; Halitschke, R.; Paschold, A.; Von Dahl, C.C.; Preston, C.A. Volatile signaling in plant-plant interactions: “Talking trees” in the genomics era. Science 2006, 311, 812–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennaoum, Z.; Benhassaini, H. Chemical Ecology of Ruta sp.: VOC, Chemotaxonomy and Allelochemistry. In Biodiversity and Chemotaxonomy; Ramawat, K.G., Ed.; Springer Nature Switzerland AG: Cham, Switzerland, 2019; Volume 24, pp. 295–318. [Google Scholar]
- Materić, D.; Bruhn, D.; Turner, C.; Morgan, G.; Mason, N.; Gauci, V. Methods in plant foliar volatile organic vompounds research. Appl. Plant Sci. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Llusia, J.; Penuelas, J.; Gimeno, B. Seasonal and species-specific response of VOC emissions by mediterranean woody plant to elevated ozone concentrations. Atmos. Environ. 2002, 36, 3931–3938. [Google Scholar] [CrossRef]
- Vivaldo, G.; Masi, E.; Taiti, C.; Caldarelli, G.; Mancuso, S. The network of plants volatile organic compounds. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, A.A. Current trends in the evolutionary ecology of plant defence. Funct. Ecol. 2011, 25, 420–432. [Google Scholar] [CrossRef]
- D’Alessandro, M.; Turlings, T.C.J. Advances and challenges in the identification of volatiles that mediate interactions among plants and arthropods. Analyst 2006, 131, 24–32. [Google Scholar] [CrossRef] [Green Version]
- The Plant List. Available online: http://www.theplantlist.org/1.1/ (accessed on 22 December 2019).
- Zhang, M.; Viennois, E.; Prasad, M.; Zhang, Y.; Wang, L.; Zhang, Z.; Han, M.K.; Xiao, B.; Xu, C.; Srinivasan, S.; et al. Edible ginger-derived nanoparticles: A novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer. Biomaterials 2016, 101, 321–340. [Google Scholar] [CrossRef] [Green Version]
- Singh, G.; Maurya, S.; Catalan, C.; de Lampasona, M.P. Studies on essential oils, Part 42: Chemical, antifungal, antioxidant and sprout suppressant studies on ginger essential oil and its oleoresin. Flavour Fragr. J. 2005, 20. [Google Scholar] [CrossRef]
- Ozaki, Y.; Kawahara, N.; Harada, M. Anti-inflammatory effect of Zingiber cassumunar Roxb. and its active principles. Chem. Pharm. Bull. 1991, 39, 2353–2356. [Google Scholar] [CrossRef] [Green Version]
- Nile, S.H.; Park, S.W. Chromatographic analysis, antioxidant, anti-inflammatory, and xanthine oxidase inhibitory activities of ginger extracts and its reference compounds. Ind. Crops Prod. 2015, 70, 238–244. [Google Scholar] [CrossRef]
- An, K.; Zhao, D.; Wang, Z.; Wu, J.; Xu, Y.; Xiao, G. Comparison of different drying methods on Chinese ginger (Zingiber officinale Roscoe): Changes in volatiles, chemical profile, antioxidant properties, and microstructure. Food Chem. 2016, 197, 1292–1300. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.V.; Murthy, P.S.; Manjunatha, J.R.; Bettadaiah, B.K. Synthesis and quorum sensing inhibitory activity of key phenolic compounds of ginger and their derivatives. Food Chem. 2014, 159, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Pithayanukul, P.; Tubprasert, J.; Wuthi-Udomlert, M. In vitro antimicrobial activity of Zingiber cassumunar (Plai) oil and a 5% Plai oil gel. Phytother. Res. 2007, 21, 164–169. [Google Scholar] [CrossRef]
- Mesomo, M.C.; Corazza, M.L.; Ndiaye, P.M.; Dalla Santa, O.R.; Cardozo, L.; Scheer, A.D.P. Supercritical CO2 extracts and essential oil of ginger (Zingiber officinale R.): Chemical composition and antibacterial activity. J. Supercrit. Fluids. 2013, 80, 44–49. [Google Scholar] [CrossRef]
- Sasidharan, I.; Menon, A.N. Comparative chemical composition and antimicrobial activity fresh and dried ginger oils (Zingiber officinale Roscoe). Int. J. Curr. Pharm. Res. 2010, 2, 40–43. [Google Scholar]
- Citronberg, J.; Bostick, R.; Ahearn, T.; Turgeon, D.K.; Ruffin, M.T.; Djuric, Z.; Sen, A.; Brenner, D.E.; Zick, S.M. Effects of ginger supplementation on cell-cycle biomarkers in the normal-appearing colonic mucosa of patients at increased risk for colorectal cancer: Results from a pilot, randomized, and controlled trial. Cancer Prev. Res. 2013, 6, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Murakami, A.; Takahashi, D.; Kinoshita, T.; Koshimizu, K.; Kim, H.W.; Yoshihiro, A.; Nakamura, Y.; Jiwajinda, S.; Terao, J.; Ohigashi, H. Zerumbone, a Southeast Asian ginger sesquiterpene, markedly suppresses free radical generation, proinflammatory protein production, and cancer cell proliferation accompanied by apoptosis: The α,β-unsaturated carbonyl group is a prerequisite. Carcinogenesis 2002, 23, 795–802. [Google Scholar] [CrossRef] [Green Version]
- Takada, Y.; Murakami, A.; Aggarwal, B.B. Zerumbone abolishes NF-κB and IκBα kinase activation leading to suppression of antiapoptotic and metastatic gene expression, upregulation of apoptosis, and downregulation of invasion. Oncogene 2005, 24, 6957–6969. [Google Scholar] [CrossRef] [Green Version]
- Kirana, C.; Record, I.; Mclntosh, G.H.; Jones, P.P. Screening for Antitumor Activity of 11 Species of Indonesian Zingiberaceae Using Human MCF-7 and HT-29 Cancer Cells. Pharm. Biol. 2003, 41, 271–276. [Google Scholar] [CrossRef]
- Vernin, G.; Parkanyi, C. Chemistry of ginger. In Ginger: The Genus Zingiber; Ravindran, P.N., Nirmal Babu, K., Eds.; CRC Press: NY, USA, 2005; pp. 87–180. [Google Scholar]
- Mao, Q.Q.; Xu, X.Y.; Cao, S.Y.; Gan, R.Y.; Corke, H.; Beta, T.; Li, H.B. Bioactive compounds and bioactivities of ginger (Zingiber officinale Roscoe). Foods 2019, 8, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wohlmuth, H.; Smith, M.K.; Brooks, L.O.; Myers, S.P.; Leach, D.N. Essential oil composition of diploid and tetraploid clones of ginger (Zingiber officinale Roscoe) grown in Australia. J. Agric. Food Chem. 2006, 54, 1414–1419. [Google Scholar] [CrossRef] [PubMed]
- Kurobayashi, Y.; Sakakibara, H.; Yanai, T.; Yajima, I.; Hayashi, K. Volatile flavor compounds of myoga (Zingiber Mioga). Agric. Biol. Chem. 1991, 55, 1655–1657. [Google Scholar]
- Sukatta, U.; Rugthaworn, P.; Punjee, P.; Chidchenchey, S.; Keeratinijakal, V. Chemical composition and physical properties of oil from plai (Zingiber cassumunar Roxb.) obtained by hydro distillation and hexane extraction. Kasetsart J. Nat. Sci. 2009, 43, 212–217. [Google Scholar]
- Tan, J.W.; Israf, D.A.; Tham, C.L. Major bioactive compounds in essential oils extracted from the rhizomes of Zingiber zerumbet (L) Smith: A mini-review on the anti-allergic and immunomodulatory properties. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.K.; Singh, V.; Ali, M. Chemical composition and antimicrobial activity of fresh rhizome essential oil of Zingiber officinale Roscoe. Pharmacog. J. 2016, 8, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Sharifi-Rad, M.; Varoni, E.M.; Salehi, B.; Sharifi-Rad, J.; Matthews, K.R.; Ayatollahi, S.A.; Kobarfard, F.; Ibrahim, S.A.; Mnayer, D.; Zakaria, Z.A.; et al. Plants of the genus Zingiber as a source of bioactive phytochemicals: From tradition to pharmacy. Molecules 2017, 22, 2145. [Google Scholar] [CrossRef] [Green Version]
- Valente, J.; Zuzarte, M.; Gonçalves, M.J.; Lopes, M.C.; Cavaleiro, C.; Salgueiro, L.; Cruz, M.T. Antifungal, antioxidant and anti-inflammatory activities of Oenanthe crocata L. essential oil. Food Chem. Toxicol. 2013, 62, 349–354. [Google Scholar] [CrossRef]
- Franz, C.; Novak, J. Sources of Essential Oils. In Handbook of Essential Oils Science, Technology, and Applications; CRC Press: Boca Raton, FL, USA, 2015; pp. 43–86. [Google Scholar]
- Aung, M.M. Taxonomic Study of the Genus Zingiber Mill. (Zingiberaceae) in Myanmar. Ph.D. Thesis, Kochi University, Kochi, Japan, 2016. [Google Scholar]
- Awale, S.; Linn, T.Z.; Than, M.M.; Swe, T.; Saiki, I.; Kadota, S. The healing art of traditional medicines in Myanmar. J. Tradit. Med. 2006, 23, 47–68. [Google Scholar]
- Wicaksana, N. Characterization and Diversity Assessment of Zingiber Species with Special Reference to Underutilized Medicinal Z. barbatum. Ph.D. Thesis, University of Tsukuba, Tsukuba, Japan, 2012. [Google Scholar]
- Wicaksana, N.; Gilani, S.A.; Ahmad, D.; Kikuchi, A.; Watanabe, K.N. Morphological and molecular characterization of underutilized medicinal wild ginger (Zingiber barbatum Wall.) from Myanmar. Plant Genet. Resour. 2011, 9, 531–542. [Google Scholar] [CrossRef]
- Tholl, D.; Boland, W.; Hansel, A.; Loreto, F.; Röse, U.S.R.; Schnitzler, J.-P. Practical approaches to plant volatile analysis. Plant J. 2006, 45, 540–560. [Google Scholar] [CrossRef] [PubMed]
- Ormeño, E.; Goldstein, A.; Niinemets, Ü. Extracting and trapping biogenic volatile organic compounds stored in plant species. Trends Analyt. Chem. 2011, 30, 978–989. [Google Scholar] [CrossRef]
- George, M.J.; Njobeh, P.B.; Gbashi, S.; Adegoke, G.O.; Dubery, I.A.; Madala, N.E. Rapid Screening of volatile organic compounds from Aframomum danielli seeds using headspace solid phase microextraction coupled to gas chromatography mass spectrometry. Int. J. Anal. Chem. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Kusano, M.; Kobayashi, M.; Iizuka, Y.; Fukushima, A.; Saito, K. Unbiased profiling of volatile organic compounds in the headspace of Allium plants using an in-tube extraction device. BMC Res. Notes 2016, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R.P. Identification of Essential Oil Compounds by Gas Chromatography/Quadrupole Mass Spectroscopy, 4th ed.; Allured: Carol Stream, IL, USA, 2012. [Google Scholar]
- König, W.A.; Joulain, D.; Hochmuth, D.H. Terpenoids Library—MassFinder. Available online: https://massfinder.com/wiki/Terpenoids_Library (accessed on 20 March 2020).
- Skogerson, K.; Wohlgemuth, G.; Barupal, D.K.; Fiehn, O. The volatile compound BinBase mass spectral database. BMC Bioinform. 2011, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, S.E.; Ausloos, P.; Clifton, C.L.; Klassen, J.K.; Lias, S.G.; Mikaya, A.I.; Sparkman, O.D.; Tchekhovskoi, D.V.; Zaikin, V.; Zhu, D. Evaluation of the NIST/EPA/NIH Mass Spectral Library. Abstr. Pap. Am. Chem. S. 1999, 218, 368. [Google Scholar]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef] [Green Version]
- ChemSpider Search and Share Chemistry. Available online: https://www.chemspider.com/ (accessed on 20 December 2019).
- Trygg, J. O2-PLS for qualitative and quantitative analysis in multivariate calibration. J. Chemom. 2002, 16, 283–293. [Google Scholar] [CrossRef]
- Barker, M.; Rayens, W. Partial least squares for discrimination. J. Chemom. 2003, 17, 166–173. [Google Scholar] [CrossRef]
- Worley, B.; Powers, R. Multivariate Analysis in Metabolomics. Curr. Metabolomics 2013, 1, 92–107. [Google Scholar]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Kettaneh, N.; Berglund, A.; Wold, S. PCA and PLS with very large data sets. Comput. Stat. Data Anal. 2005, 48, 69–85. [Google Scholar] [CrossRef]
- Eriksson, L.; Byrne, T.; Johansson, E.; Trygg, J.; Vikström, C. Multi- and Megavariate Data Analysis Basic Principles and Applications, 3rd ed.; Umetrics Academy: Umeå, Sweden, 2013. [Google Scholar]
- Perveen, S. Introductory Chapter: Terpenes and Terpenoids. In Terpenes and Terpenoids; Perveen, S., Al-Taweel, A., Eds.; IntechOpen: London, UK, 2018; pp. 1–12. [Google Scholar]
- Sobral, M.V.; Xavier, A.L.; Lima, T.C.; de Sousa, D.P. Antitumor activity of monoterpenes found in essential oils. Sci. World J. 2014, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Koziol, A.; Stryjewska, A.; Librowski, T.; Salat, K.; Gawel, M.; Moniczewski, A.; Lochynski, S. An overview of the pharmacological properties and potential applications of natural monoterpenes. Mini-Rev. Med. Chem. 2014, 14, 1156–1168. [Google Scholar] [CrossRef] [PubMed]
- Repetto, M.G.; Boveris, A. Bioactivity of sesquiterpenes: Compounds that protect from alcohol-induced gastric mucosal lesions and oxidative damage. Mini-Rev. Med. Chem. 2010, 10, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Iijima, Y.; Koeduka, T.; Suzuki, H.; Kubota, K. Biosynthesis of geranial, a potent aroma compound in ginger rhizome (Zingiber officinale): Molecular cloning and characterization of geraniol dehydrogenase. Plant Biotechnol. 2014, 534, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, O. Identification of the characteristic odorants in fresh rhizomes of ginger (Zingiber officinale Roscoe) using aroma extract dilution analysis and modified multidimensional gas chromatography-mass spectroscopy. J. Agric. Food Chem. 1995, 43, 2941–2945. [Google Scholar] [CrossRef]
- Dong, L.; Miettinen, K.; Goedbloed, M.; Verstappen, F.W.A.; Voster, A.; Jongsma, M.A.; Memelink, J.; van der Krol, S.; Bouwmeester, H.J. Characterization of two geraniol synthases from Valeriana officinalis and Lippia dulcis: Similar activity but difference in subcellular localization. Metab. Eng. 2013, 20, 198–211. [Google Scholar] [CrossRef]
- Brilli, F.; Loreto, F.; Baccelli, I. Exploiting plant volatile organic compounds (VOCS) in agriculture to improve sustainable defense strategies and productivity of crops. Front Plant Sci. 2019, 10, 1–8. [Google Scholar] [CrossRef]
- Şanli, A.; Karadoğan, T. Geographical Impact on Essential Oil Composition of endemic Kundmannia Anatolica Hub.-Mor. (Apiaceae). Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 131–137. [Google Scholar] [CrossRef]
- Demasi, S.; Caser, M.; Lonati, M.; Cioni, P.L.; Pistelli, L.; Najar, B.; Scariot, V. Latitude and altitude influence secondary metabolite production in peripheral alpine populations of the mediterranean species Lavandula angustifolia Mill. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.P.; Wang, S.; Zhang, J.; Yang, G.; Zhao, M.X.; Ma, W.F.; Zhang, X.B.; Li, X.; Han, B.X.; Chen, N.F.; et al. Effects of ecological factors on secondary metabolites and inorganic elements of Scutellaria baicalensis and analysis of geoherblism. Sci. China Life Sci. 2013, 56, 1047–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, A.; Baldwin, I.T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef] [PubMed]
- Schnee, C.; Köllner, T.G.; Held, M.; Turlings, T.C.J.; Gershenzon, J.; Degenhardt, J. The products of a single maize sesquiterpene synthase form a volatile defense signal that attracts natural enemies of maize herbivores. Proc. Natl. Acad. Sci. USA 2006, 103, 1129–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dicke, M.; Loreto, F. Induced plant volatiles: From genes to climate change. Trends Plant Sci. 2010, 15, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Loreto, F.; Schnitzler, J.P. Abiotic stresses and induced BVOCs. Trends Plant Sci. 2010, 15, 154–166. [Google Scholar] [CrossRef]
- Smyth, G.K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, 1–25. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]



| Compounds | ZO63 | ZO105 | ZO160 | ZO191 | ZO217 | ZO223 |
|---|---|---|---|---|---|---|
| 1,3,5-Cycloheptatriene | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Hexanal | 0.00 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| 2-methylbutan-2-yl acetate | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| (Z)-3-Octene | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Heptanal | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| γ-Butyrolactone | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| α-Thujene | 0.30 ± 0.09 a | 0.56 ± 0.34 a | 0.22 ± 0.07 a | 0.46 ± 0.28 a | 0.15 ± 0.14 a | 0.16 ± 0.09 a |
| 2-Octanone | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Sabinene | 0.25 ± 0.13 a | 0.32 ± 0.15 a | 0.25 ± 0.28 a | 0.53 ± 0.32 a | 0.10 ± 0.04 a | 0.16 ± 0.16 a |
| Butyl pivalate | 0.08 ± 0.04 a,b | 0.06 ± 0.03 a,b,c | 0.06 ± 0.03 a,b,c | 0.02 ± 0.01 b,c | 0.09 ± 0.03 a | 0.01 ± 0.00 c |
| Decane | 0.08 ± 0.01 a | 0.08 ± 0.01 a | 0.08 ± 0.01 a | 0.09 ± 0.01 a | 0.08 ± 0.01 a | 0.08 ± 0.01 a |
| Octanal | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a | 0.01 ± 0.00 a |
| α-Phellandrene | 0.12 ± 0.02 a,b | 0.21 ± 0.13 a | 0.09 ± 0.01 a,b | 0.12 ± 0.06 a,b | 0.04 ± 0.01 b | 0.07 ± 0.02 b |
| 1,4-Dichlorobenzene | 0.04 ± 0.00 a | 0.04 ± 0.01 a | 0.04 ± 0.01 a | 0.05 ± 0.00 a | 0.04 ± 0.01 a | 0.04 ± 0.00 a |
| α-Terpinene | 0.33 ± 0.08 a,b | 0.64 ± 0.43 a | 0.21 ± 0.06 a,b | 0.36 ± 0.19 a,b | 0.09 ± 0.05 b | 0.17 ± 0.08 a,b |
| o-Cymene | 0.76 ± 0.29 a | 1.51 ± 0.90 a | 0.65 ± 0.14 a | 0.79 ± 0.49 a | 0.52 ± 0.04 a | 0.59 ± 0.15 a |
| Limonene | 0.02 ± 0.01 a | 0.03 ± 0.02 a | 0.01 ± 0.00 a | 0.02 ± 0.01 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a |
| β-Phellandrene | 0.12 ± 0.03 a,b | 0.23 ± 0.13 a | 0.09 ± 0.02 b | 0.14 ± 0.07 a,b | 0.03 ± 0.03 b | 0.07 ± 0.03 b |
| 3-Methyldecane | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a,b | 0.00 ± 0.00 b | 0.00 ± 0.00 a | 0.00 ± 0.00 b |
| 2-Methyldecane | 0.02 ± 0.01 a | 0.02 ± 0.00 a | 0.03 ± 0.01 a | 0.03 ± 0.01 a | 0.02 ± 0.00 a | 0.03 ± 0.01 a |
| γ-Terpinene | 0.48 ± 0.12 a,b | 0.94 ± 0.68 a | 0.30 ± 0.10 a,b | 0.55 ± 0.30 a,b | 0.13 ± 0.07 b | 0.23 ± 0.12 a,b |
| 1-Octanol | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a | 0.01 ± 0.00 a |
| (Z)-Sabinenhydrate | 0.02 ± 0.02 a | 0.01 ± 0.01 a | 0.01 ± 0.01 a | 0.02 ± 0.01 a | 0.00 ± 0.01 a | 0.01 ± 0.01 a |
| (E)-Sabinenhydrate | 0.07 ± 0.07 a | 0.04 ± 0.05 a | 0.03 ± 0.04 a | 0.08 ± 0.05 a | 0.02 ± 0.02 a | 0.03 ± 0.03 a |
| (4E)-7-Methyl-4-decene | 0.02 ± 0.00 a,b | 0.01 ± 0.00 a,b | 0.01 ± 0.00 a,b | 0.01 ± 0.00 a | 0.01 ± 0.00 b | 0.01 ± 0.00 b |
| 4-Tolualdehyde | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Terpinolene | 0.11 ± 0.03 a,b | 0.21 ± 0.15 a | 0.07 ± 0.02 a,b | 0.12 ± 0.07 a,b | 0.03 ± 0.02 b | 0.05 ± 0.03 a,b |
| p-Cymenene | 0.00 ± 0.00 a | 0.01 ± 0.01 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Decane | 0.05 ± 0.01 a | 0.06 ± 0.01 a | 0.05 ± 0.01 a | 0.07 ± 0.01 a | 0.05 ± 0.00 a | 0.06 ± 0.02 a |
| Nonanal | 0.03 ± 0.03 a | 0.06 ± 0.05 a | 0.04 ± 0.02 a | 0.04 ± 0.05 a | 0.05 ± 0.02 a | 0.03 ± 0.02 a |
| 2,4,6-Trimethyldecane | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a |
| (E)-3-caren-2-ol | 0.00 ± 0.00 a,b | 0.001±0.00 a | 0.00 ± 0.00 ab | 0.00 ± 0.00 a,b | 0.00 ± 0.00 b | 0.00 ± 0.00 a,b |
| Octyl acetate | 0.01 ± 0.00 a | 0.01 ± 0.00 a,b | 0.01 ± 0.00 a,b | 0.01 ± 0.00 a,b | 0.01 ± 0.00 a,b | 0.01 ± 0.00 b |
| L-camphor | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a |
| 1-Nonanol | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Tetrahydrolinalyl acetate | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Terpinen-4-ol | 0.72 ± 0.04 b | 1.25 ± 0.21 a | 0.63 ± 0.10 a,c | 0.79 ± 0.09 b | 0.26 ± 0.06 c | 0.87 ± 0.32 b |
| Naphthalene | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| 5,6-Dimethylundecane | 0.02 ± 0.00 a | 0.01 ± 0.00 a | 0.02 ± 0.00 a | 0.02 ± 0.00 a | 0.02 ± 0.00 a | 0.02 ± 0.00 a |
| (Z)-Sabinene hydrate acetate | 0.01 ± 0.00 a,b | 0.03 ± 0.01 a | 0.01 ± 0.00 b | 0.02 ± 0.01 a,b | 0.01 ± 0.00 b | 0.01 ± 0.01 b |
| 1,3-Di-tert-butylbenzene | 0.03 ± 0.01 a | 0.03 ± 0.01 a | 0.03 ± 0.01 a | 0.03 ± 0.01 a | 0.03 ± 0.02 a | 0.02 ± 0.01 a |
| 4,6-Dimethyldodecane | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Bornyl acetate | 0.15 ± 0.04 a | 0.05 ± 0.02 b | 0.12 ± 0.03 a | 0.03 ± 0.01 b | 0.03 ± 0.01 b | 0.02 ± 0.01 b |
| Terpinen-4-ol acetate | 0.02 ± 0.01 a | 0.03 ± 0.01 a | 0.01 ± 0.01 a | 0.03 ± 0.01 a | 0.01 ± 0.00 a | 0.02 ± 0.01 a |
| Tridecane | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Valeric acid | 0.01 ± 0.00 a | 0.00 ± 0.00 b | 0.00 ± 0.00 a,b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b |
| 2,7,10-Trimethyldodecane | 0.00 ±0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.01 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| 2,5-Bornanediol | 0.00 ± 0.00 b | 0.00 ± 0.00 a | 0.00 ± 0.00 b | 0.00 ± 0.00 a | 0.00 ± 0.00 b | 0.00 ± 0.00 a |
| β-Terpinyl acetate | 0.09 ± 0.02 a | 0.10 ± 0.04 a | 0.07 ± 0.04 a,b | 0.07 ± 0.02 a,b | 0.03 ± 0.01 b | 0.04 ± 0.02 a,b |
| 3-Hydroxy-2,4,4-trimethylpentyl 2-methylpropanoate | 0.03 ± 0.00 a | 0.03 ± 0.00 a | 0.03 ± 0.00 a | 0.03 ± 0.00 a | 0.03 ± 0.00 a | 0.03 ± 0.01 a |
| α-Ylangene | 0.01 ± 0.01 a | 0.00 ± 0.00 b | 0.01 ± 0.00 a,b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b |
| β-Elemene | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.01 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Tetradecane | 0.04 ± 0.01 a | 0.03 ± 0.01 a | 0.03 ± 0.01 a | 0.04 ± 0.01 a | 0.04 ± 0.00 a | 0.03 ± 0.01 a |
| γ-Elemene | 0.01 ± 0.02 a | 0.00 ± 0.00 a | 0.01 ± 0.01 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| α-Bergamotene | 0.00 ± 0.00 a | 0.00 ±0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| 12-Chloro-5-dodecyne | 0.01 ± 0.00 a | 0.00 ± 0.00 b | 0.00 ± 0.00 a,b | 0.00 ± 0.00 a,b | 0.00 ± 0.00 b | 0.00 ± 0.00 b |
| Dihydrocurcumene | 0.01 ± 0.01 a | 0.00 ± 0.00 a | 0.01 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| β-Farnesene | 0.03 ± 0.01 a | 0.00 ± 0.00 b | 0.02 ± 0.01 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b |
| Selina-5,11-diene | 0.02 ± 0.02 a | 0.00 ± 0.00 a | 0.01 ± 0.01 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| 3,11-Acoradiene | 0.02 ± 0.02 a | 0.01 ± 0.00 a | 0.02 ± 0.02 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| α-Curcumene | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| γ-Curcumene | 0.04 ± 0.04 a | 0.00 ± 0.00 a | 0.03 ± 0.02 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Cuparene | 0.22 ± 0.20 a | 0.02 ± 0.01 a | 0.19 ± 0.11 a | 0.03 ± 0.02 a | 0.013 ± 0.01 a | 0.10 ± 0.01 a |
| γ-Amorphene | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Germacrene A | 0.44 ± 0.43 a | 0.00 ± 0.00 a | 0.39 ± 0.22 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| α-Zingiberene | 0.38 ± 0.34 a | 0.02 ± 0.02 b | 0.34 ± 0.17 a,b | 0.02 ± 0.02 b | 0.01 ± 0.01 b | 0.01 ± 0.00 b |
| Valencene | 0.05 ± 0.05 a | 0.00 ± 0.00 a | 0.05 ± 0.03 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| 2,4-Di-tert-butylphenol | 0.78 ± 0.08 a | 0.70 ± 0.14 a | 0.80 ± 0.10 a | 0.80 ± 0.11 a | 0.81 ± 0.13 a | 0.66 ± 0.19 a |
| (E)-β-Guaiene | 0.14 ± 0.11 a | 0.00 ± 0.00 b | 0.11 ± 0.06 a,b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b |
| β-Bisabolene | 0.05 ± 0.04 a | 0.00 ± 0.00 b,c | 0.05 ± 0.02 a,b | 0.01 ± 0.00 b,c | 0.00 ± 0.00 c | 0.00 ± 0.00 b,c |
| σ-Cadinene | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| β-Sesquiphellandrene | 1.19 ± 0.80 a | 0.11 ± 0.07 b | 1.32 ± 0.68 a | 0.17 ± 0.11 b | 0.09 ± 0.03 b | 0.09 ± 0.05 b |
| 7-epi-α-Selinene | 0.08 ± 0.07 a | 0.00 ± 0.00 b | 0.07 ± 0.03 a,b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b |
| 1-Iodoundecane | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| (E)-γ-Macrocarpene | 0.01 ± 0.01 a | 0.00 ± 0.00 a | 0.01 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Germacrene B | 0.01 ± 0.01 a | 0.00 ± 0.00 a | 0.01 ± 0.01 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| 2,2,4-Trimethyl-1,3-pentadienol diisobutyrate | 0.02 ± 0.00 a | 0.02 ± 0.00 a | 0.02 ± 0.01 a | 0.02 ± 0.01 a | 0.02 ± 0.01 a | 0.01 ± 0.00 a |
| 2-Allyl-1,4-dimethoxy-3-methylbenzene | 0.02 ± 0.01 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a | 0.03 ± 0.02 a | 0.03 ± 0.02 a | 0.02 ± 0.01 a |
| Hexadecane | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a | 0.02 ± 0.01 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a |
| Elemol acetate | 0.05 ± 0.01 a | 0.00 ± 0.00 c | 0.02 ± 0.02 b | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c |
| 2-Tetradecyloxirane | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shukurova, M.K.; Asikin, Y.; Chen, Y.; Kusano, M.; Watanabe, K.N. Profiling of Volatile Organic Compounds in Wild Indigenous Medicinal Ginger (Zingiber barbatum Wall.) from Myanmar. Metabolites 2020, 10, 248. https://doi.org/10.3390/metabo10060248
Shukurova MK, Asikin Y, Chen Y, Kusano M, Watanabe KN. Profiling of Volatile Organic Compounds in Wild Indigenous Medicinal Ginger (Zingiber barbatum Wall.) from Myanmar. Metabolites. 2020; 10(6):248. https://doi.org/10.3390/metabo10060248
Chicago/Turabian StyleShukurova, Musavvara Kh., Yonathan Asikin, Yanhang Chen, Miyako Kusano, and Kazuo N. Watanabe. 2020. "Profiling of Volatile Organic Compounds in Wild Indigenous Medicinal Ginger (Zingiber barbatum Wall.) from Myanmar" Metabolites 10, no. 6: 248. https://doi.org/10.3390/metabo10060248