Coastal Pine-Oak Glacial Refugia in the Mediterranean Basin: A Biogeographic Approach Based on Charcoal Analysis and Spatial Modelling

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
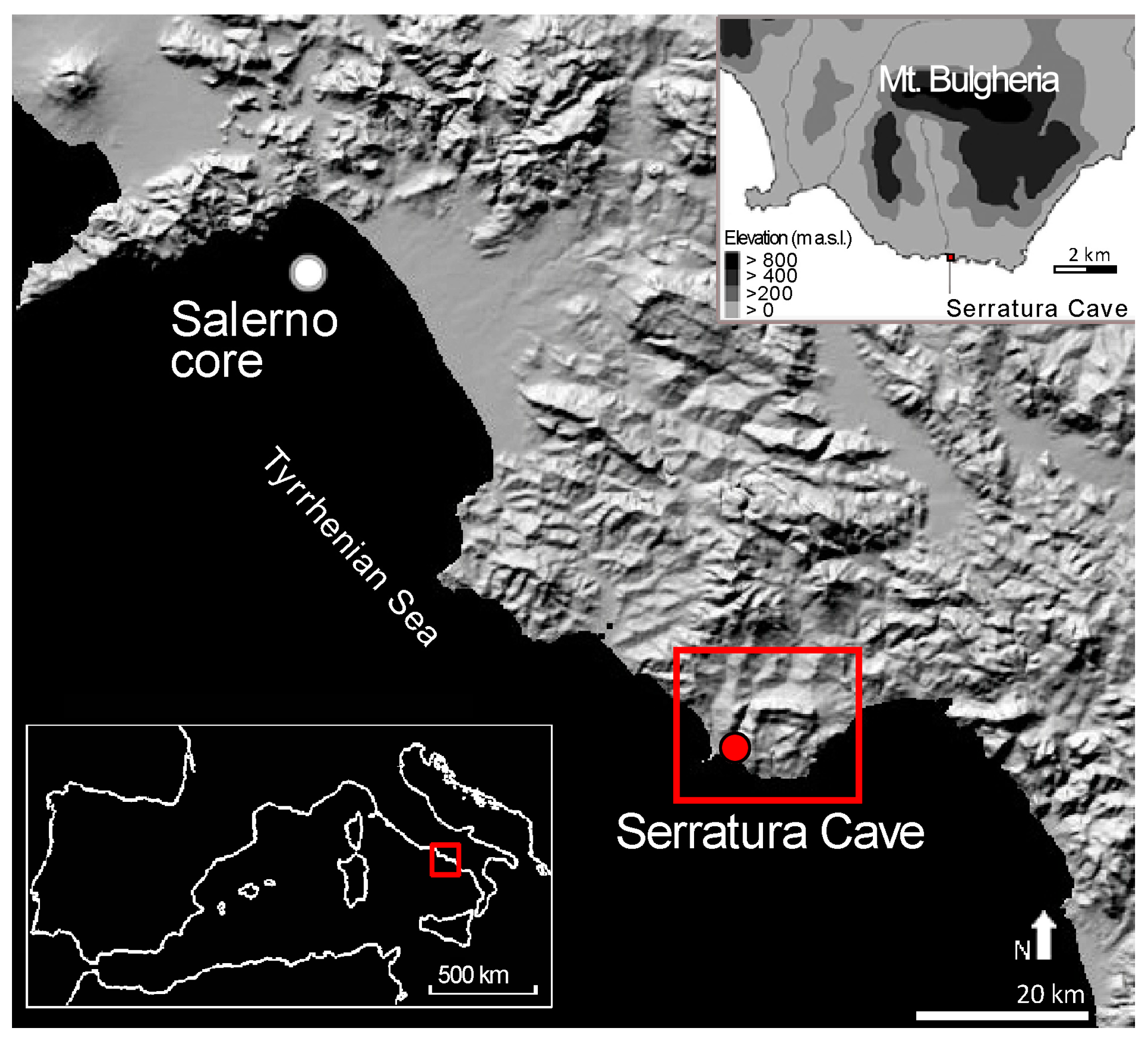
2.1. Study Area
2.2. Charcoal Analysis
2.3. Ecological Niche Model
2.3.1. Training and Projection Area
2.3.2. Data Collection
2.3.3. Environmental Variables
2.3.4. Maxent Models
2.3.5. Model Validation
2.3.6. Resolving Taxonomic Ambiguity by Means of ENM
3. Results
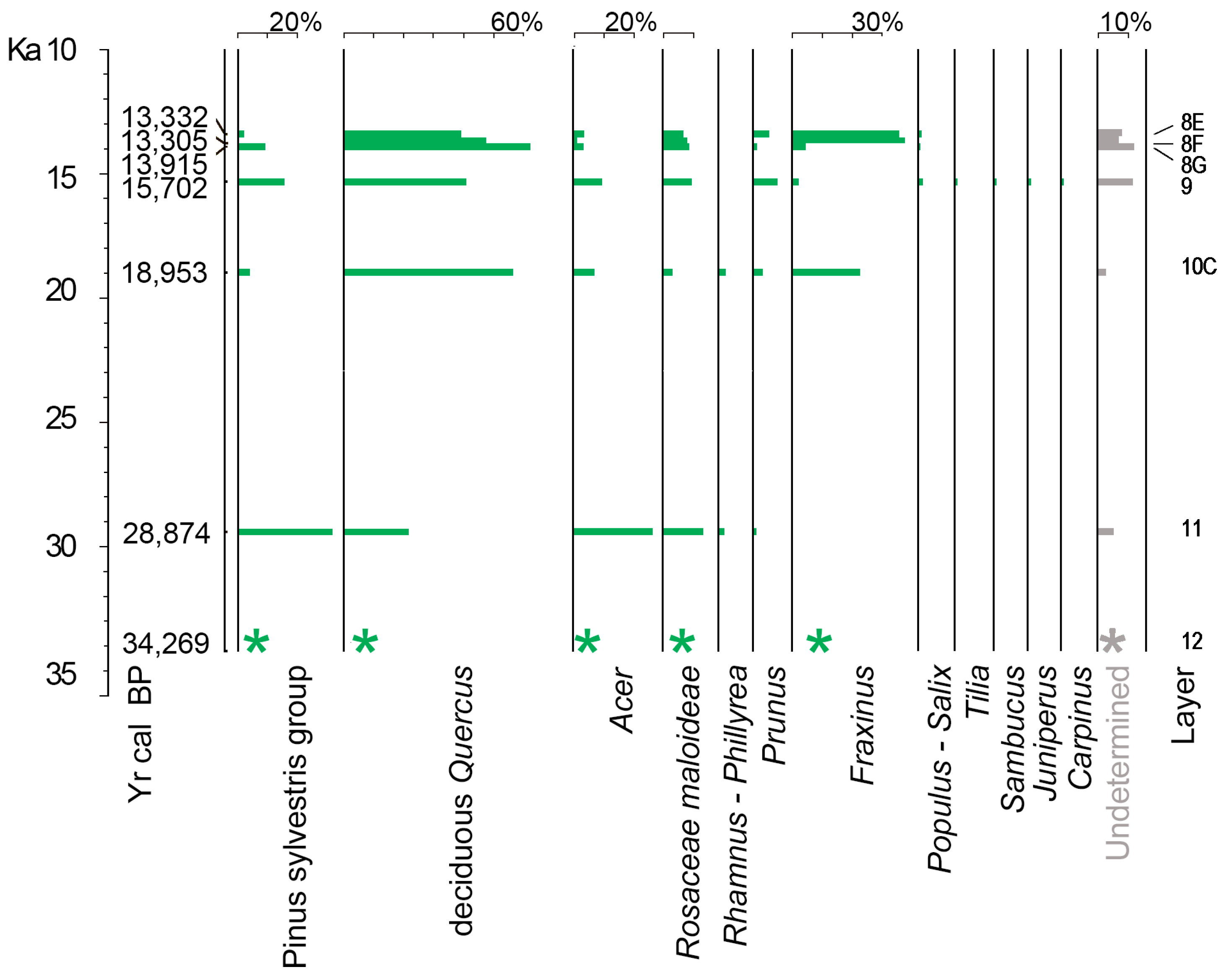
3.1. Charcoal Analysis
3.2. Ecological Niche Models of Pinus spp. and Q. pubescens
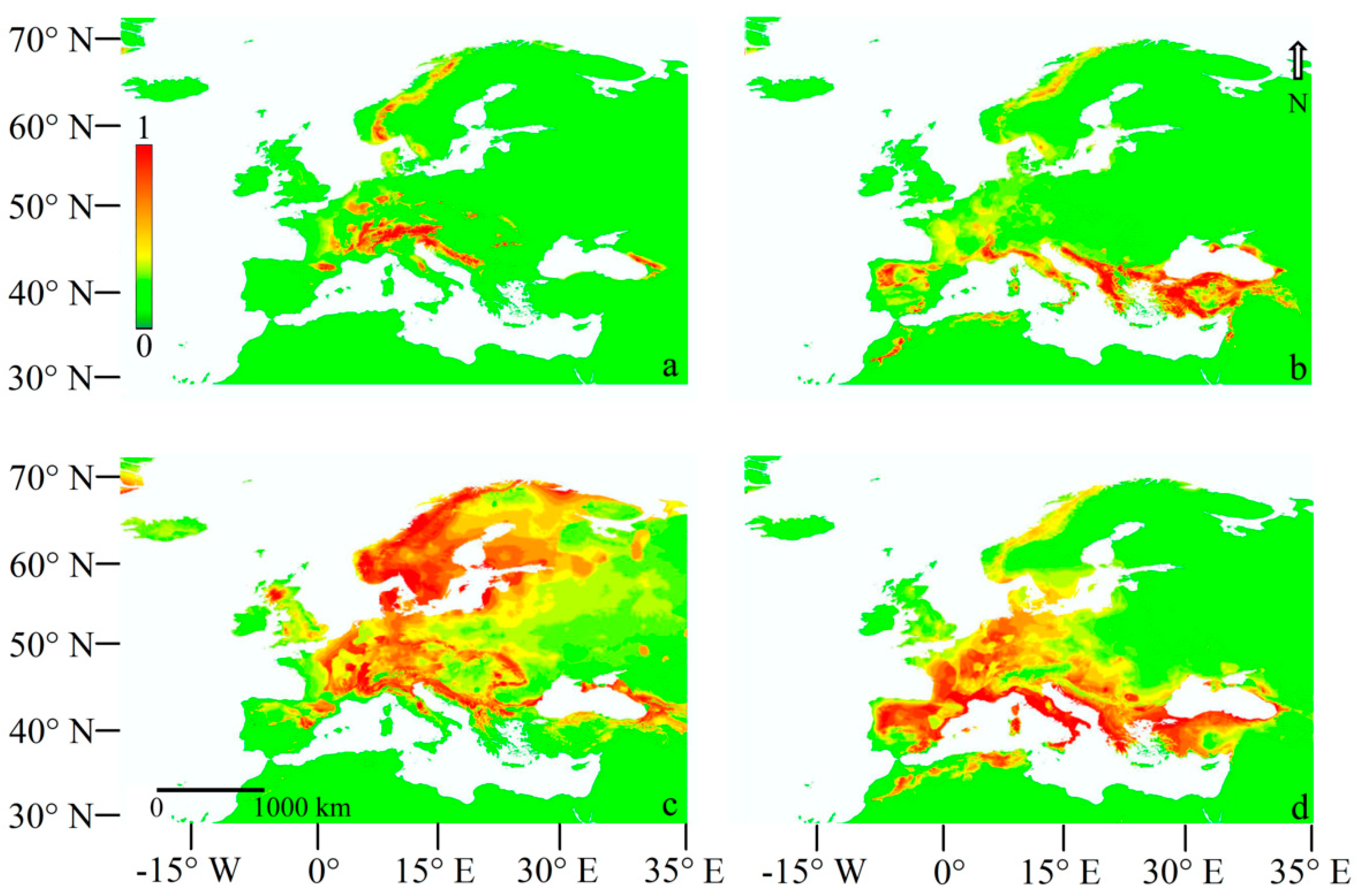
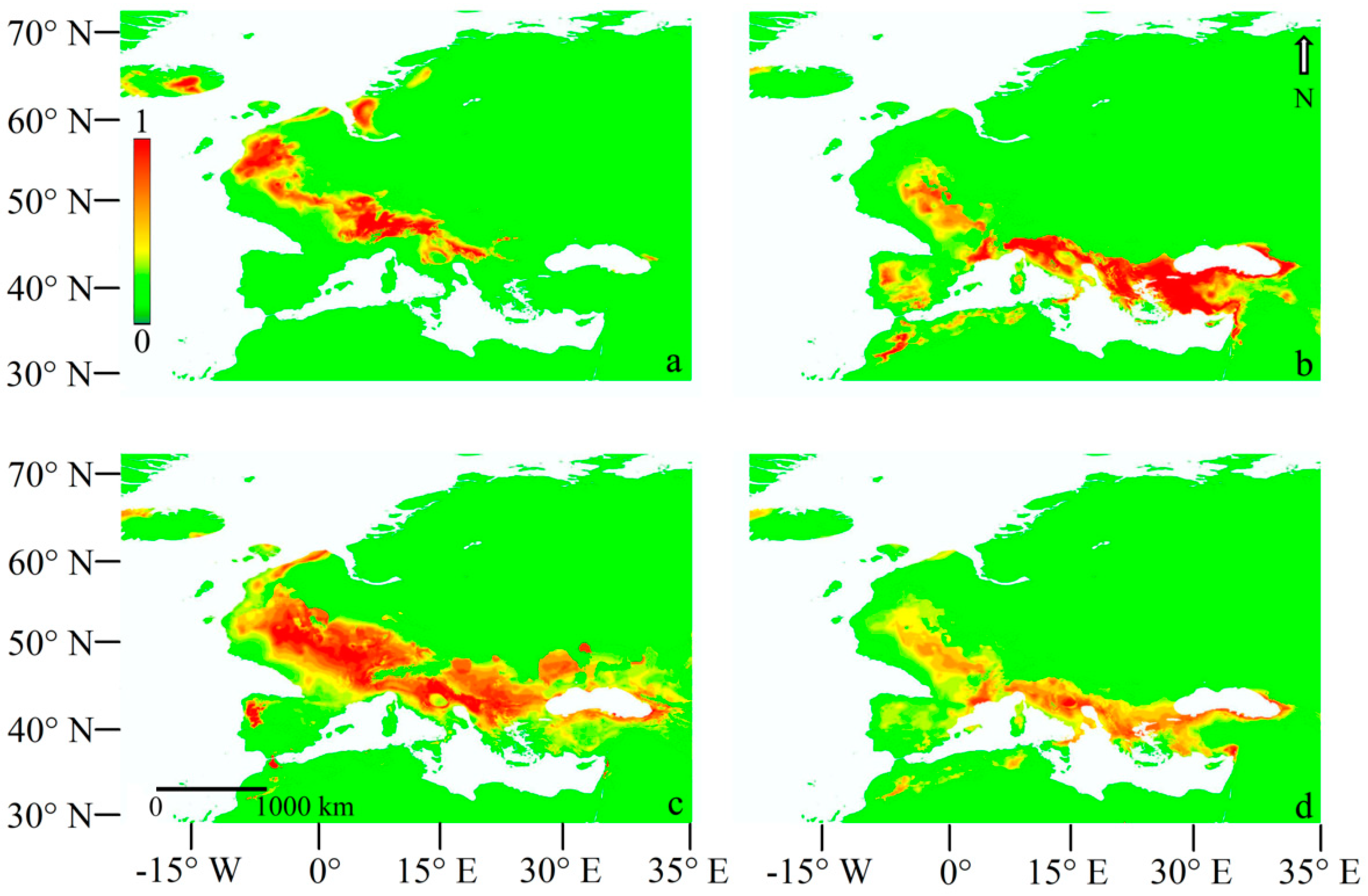
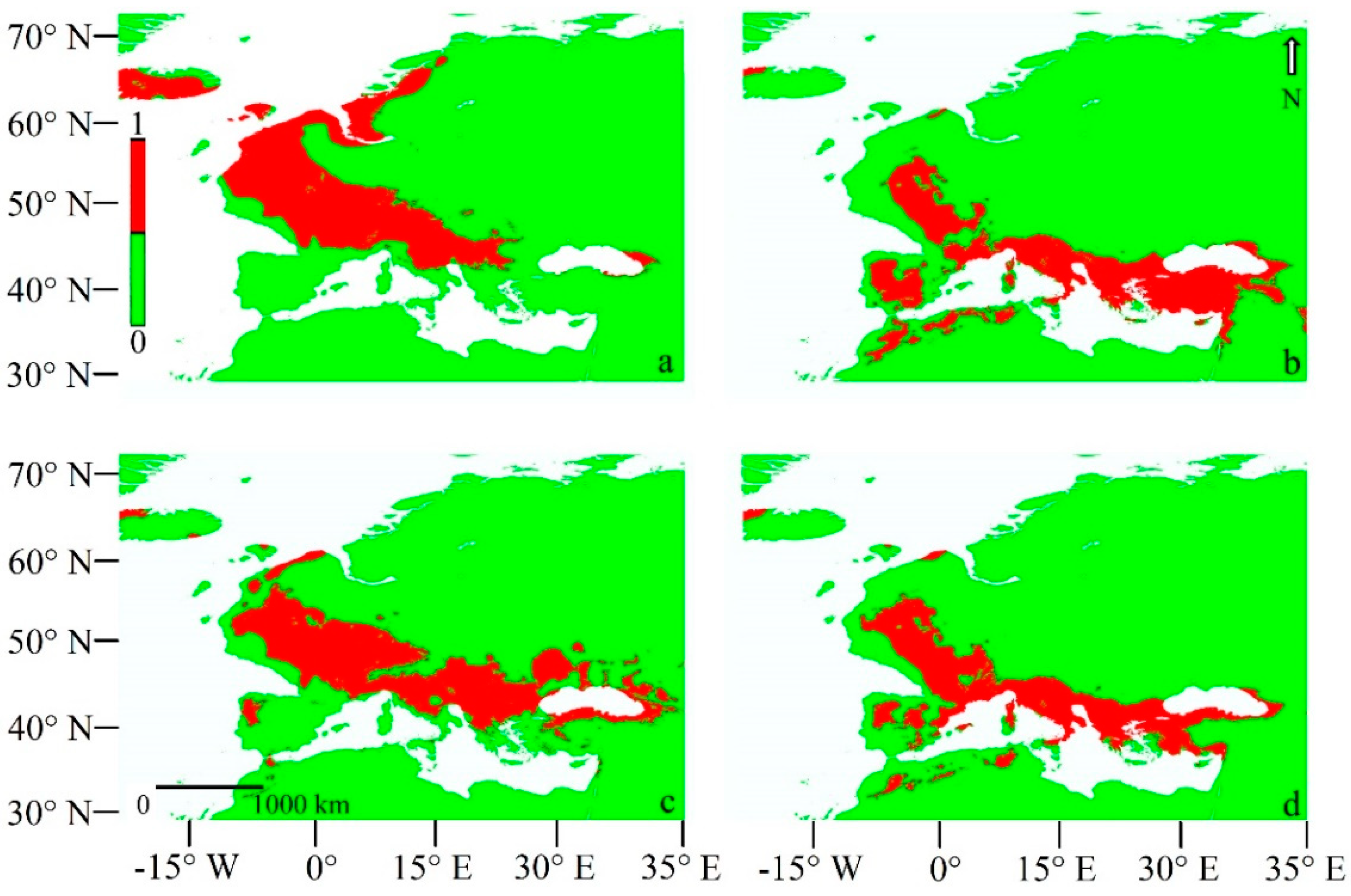
3.2.1. Current Models
3.2.2. Last Glacial Maximum Projection Models
3.2.3. LGM Projection Models Adding the Q. pubescens Distribution
4. Discussion
4.1. Pinus Type Sylvestris: Ecological Niche Models and Ecological Considerations
4.2. Vegetation Cover of the Pleniglacial
4.3. Vegetation Cover of the Late Glacial
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bennett, K.D.; Tzedakis, P.C.; Willis, K.J. Quaternary refugia of north European trees. J. Biogeogr. 1991, 18, 103–115. [Google Scholar] [CrossRef]
- Brewer, S.; Cheddadi, R.; De Beaulieu, J.L.; Reille, M. The spread of deciduous Quercus throughout Europe since the last glacial period. Ecol. Manag. 2002, 156, 27–48. [Google Scholar] [CrossRef]
- Cheddadi, R.; Vendramin, G.G.; Litt, T.; François, L.; Kageyama, M.; Lorentz, S.; Laurent, J.-M.; De Beaulieu, J.-L.; Sadori, L.; Jost, A.; et al. Imprints of glacial refugia in the modern genetic diversity of Pinus sylvestris. Glob. Ecol. Biogeogr. 2006, 15, 271–282. [Google Scholar] [CrossRef]
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef]
- Gavin, D.G.; Fitzpatrick, M.C.; Gugger, P.F.; Heath, K.D.; Rodríguez-Sánchez, F.; Dobrowski, S.Z.; Hampe, A.; Sheng Hu, F.; Ashcroft, M.B.; Bartlein, P.J.; et al. Climate refugia: Joint inference from fossil records, species distribution models and phylogeography. New Phytol. 2014, 204, 37–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbard, P.; Ehlers, J. Extent and chronology of Quaternary glaciation. Episodes 2008, 31, 211–218. [Google Scholar]
- Quézel, P.; Médail, F. Ecologie et Biogéographie des Forêts du Bassin Méditerranéen; Elsevier: Paris, France, 2003. [Google Scholar]
- Gómez, A.; Lunt, D.H. Refugia within refugia: Patterns of phylogeographic concordance in the Iberian Peninsula. In Phylogeography of Southern European Refugia; Weiss, S., Ferrand, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 155–188. [Google Scholar]
- Willis, K.J.; Whittaker, R.J. The refugial debate. Science 2000, 287, 1406–1407. [Google Scholar] [CrossRef]
- Figueiral, I.; Mosbrugger, V. A review of charcoal analysis as a tool for assessing Quaternary and Tertiary environments: Achievements and limits. Paleogeogr. Paleoclimatol. Paleoecol. 2000, 164, 397–407. [Google Scholar] [CrossRef]
- Taberlet, P.; Cheddadi, R. Quaternary refugia and persistence of biodiversity. Science 2002, 297, 2009–2010. [Google Scholar] [CrossRef]
- Waltari, E.; Hijmans, R.J.; Peterson, A.T.; Nyári, Á.S.; Perkins, S.L.; Guralnick, R.P. Locating Pleistocene refugia: Comparing phylogeographic and ecological niche model predictions. PLoS ONE 2007, 2, e563. [Google Scholar] [CrossRef] [Green Version]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Renfrew, C.; Bahn, P.G. Archaeology: Theories, Methods and Practice; Thames and Hudson: London, UK, 2016. [Google Scholar]
- Scheel-Ybert, R. Anthracology: Charcoal Analysis. In Encyclopedia of Global Archaeology; Smith, C., Ed.; Springer: New York, NY, USA, 2018; pp. 1–11. [Google Scholar]
- Svenning, J.-C.; Fløjgaard, C.; Marske, K.A.; Nógues-Bravo, D.; Normand, S. Applications of species distribution modeling to paleobiology. Quat. Sci. Rev. 2011, 30, 2930–2947. [Google Scholar] [CrossRef]
- Eduardo, A.A.; Martinez, P.A.; Gouveia, S.F.; da Silva Santos, F.; de Aragao, W.S.; Morales-Barbero, J.; Kerber, L.; Liparini, A. Extending the paleontology–biogeography reciprocity with ENMs: Exploring models and data in reducing fossil taxonomic uncertainty. PLoS ONE 2018, 13, e0194725. [Google Scholar] [CrossRef]
- Jurestovsky, D.; Joyner, T.A. Applications of species distribution modeling for paleontological fossil detection: Late Pleistocene models of Saiga (Artiodactyla: Bovidae, Saiga tatarica). Paleobiodivers. Paleoenviron. 2018, 98, 277–285. [Google Scholar] [CrossRef]
- Ribeiro, M.M.; Roque, N.; Ribeiro, S.; Gavinhos, C.; Castanheira, I.; Quinta-Nova, L.; Albuquerque, T.; Gerassis, S. Bioclimatic modeling in the Last Glacial Maximum, Mid-Holocene and facing future climatic changes in the strawberry tree (Arbutus unedo L.). PLoS ONE 2019, 14, e0210062. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. J. R. Meteorol. Soc. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Tang, C.Q.; Dong, Y.-F.; Herrando-Moraira, S.; Matsui, T.; Ohashi, H.; He, L.-Y.; Nakao, K.; Tanaka, N.; Tomita, M.; Li, X.-S. Potential effects of climate change on geographic distribution of the Tertiary relict tree species Davidia involucrata in China. Sci. Rep. 2017, 7, 43822. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Ji, X.; Obata, S.; Pais, A.; Dong, Y.; Peet, R.; Xiang, Q.-Y.J. Resolving relationships and phylogeographic history of the Nyssa sylvatica complex using data from RAD-seq and species distribution modeling. Mol. Phylogenetics Evol. 2018, 126, 1–16. [Google Scholar] [CrossRef]
- Bailey, G.; Carrion, J.S.; Fa, D.A.; Finlayson, C.; Finlayson, G.; Rodriguez-Vidal, J. The coastal shelf of the Mediterranean and beyond: Corridor and refugium for human populations in the Pleistocene. Quat. Sci. Rev. 2008, 27, 2095–2099. [Google Scholar] [CrossRef]
- Ermolli, E.R.; di Pasquale, G. Vegetation dynamics of south-western Italy in the last 28 kyr inferred from pollen analysis of a Tyrrhenian Sea core. Veg. Hist. Archaeobot. 2002, 11, 211–220. [Google Scholar] [CrossRef]
- Svenning, J.-C.; Normand, S.; Kageyama, M. Glacial refugia of temperate trees in Europe: Insights from species distribution modelling. J. Ecol. 2008, 96, 1117–1127. [Google Scholar] [CrossRef]
- Spada, F.; Cutini, M.; Paura, B. Floristic changes along the topographical gradient in montane grasslands in Monti Picentini (Campania, SW Italy). Ann. Bot. 2010, 0, 86–98. [Google Scholar]
- Bucci, G.; Borghetti, M. Understory vegetation as a useful predictor of natural regeneration and canopy dynamics in Pinus sylvestris forests in Italy. Acta Oecol. 1997, 18, 485–501. [Google Scholar] [CrossRef]
- Santangelo, N.; Santo, A.; Guida, D.; Lanzara, R.; Siervo, V. The geosites of the Cilento-Vallo di Diano National Park (Campania region, southern Italy). Quat. Vol. Spec. 2005, 18, 104–114. [Google Scholar]
- Martini, F. Grotta della Serratura a Marina di Camerota.Culture e Ambienti dei Complessi Olocenici; Garlatti & Razzai: Firenze, Italy, 1993. [Google Scholar]
- Martini, I.; Ronchitelli, A.; Arrighi, S.; Capecchi, G.; Ricci, S.; Scaramucci, S.; Spagnolo, V.; Gambassini, P.; Moroni, A. Cave clastic sediments as a tool for refining the study of human occupation of prehistoric sites: Insights from the cave site of La Cala (Cilento, southern Italy). J. Quat. Sci. 2018, 33, 586–596. [Google Scholar] [CrossRef]
- Bonanomi, G.; Rita, A.; Allevato, E.; Cesarano, G.; Saulino, L.; Di Pasquale, G.; Allegrezza, M.; Pesaresi, S.; Borghetti, M.; Rossi, S. Anthropogenic and environmental factors affect the tree line position of Fagus sylvatica along the Apennines (Italy). J. Biogeogr. 2018, 45, 2595–2608. [Google Scholar] [CrossRef]
- Greguss, P. Identification of Living Gymnosperms on the Basis of Xylotomy; Akadémiai Kiadó: Budapest, Hungary, 1955. [Google Scholar]
- Greguss, P. Holzanatomie der Europäischen Laubhölzer und Sträucher; Akadémiai Kiadó: Budapest, Hungary, 1959. [Google Scholar]
- Schweingruber, F.H. Anatomie Europäischer Hölzer: Ein Atlas zur Bestimmung Europäischer Baum-, Strauch- und Zwergstrauchhölzer; P. Haupt: Bern, Switzerland; Stuttgart, Germany, 1990. [Google Scholar]
- Vernet, J.L.; Ogereau, P.; Figueiral, I.; Machado Yanes, C.; Uzquiano, P. Guide D’identification des Charbons de Bois Préhistoriques du Sud-Ouest de l’Europe; CNRS: Paris, France, 2001. [Google Scholar]
- Cambini, A. Micrografia Comparata dei Legni del Genere Quercus; Riconoscimento Microscopico del Legno delle Querce Italiane; CNR: Rome, Italy, 1967. [Google Scholar]
- Chabal, L.; Fabre, J.F.; Terral, I.; Théry-Parisot, I. L’anthracologie. In La Botanique; Bourquin-Mignot, C., Brochier, J.E., Chabal, L., Crozat, S., Fabre, L., Guibal, F., Marinval, P., Terral, J.F., Théry-Parisot, I., Eds.; Errance: Paris, France, 1999; pp. 43–104. [Google Scholar]
- Chabal, L. La représentativité paléo-écologique des charbons de bois archéologiques issus du bois de feu. Bull. Soc. Bot. Fr. Actual. Bot. 1992, 139, 213–236. [Google Scholar] [CrossRef]
- Heinz, C.; Thiébault, S. Characterization and paleoecological significance of archaeological charcoal assemblages during late and post-glacial phases in southern France. Quat. Res. 1998, 50, 56–68. [Google Scholar] [CrossRef]
- Asouti, E.; Austin, P. Reconstructing woodland vegetation and its exploitation by past societies, based on the analysis and interpretation of archaeological wood charcoal macro-remains. Environ. Archaeol. 2005, 10, 1–18. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Allevato, E.; Cocchiararo, A.; Moser, D.; Pacciarelli, M.; Saracino, A. Late Holocene persistence of Abies alba in low-mid altitude deciduous forests of central and southern Italy: New perspectives from charcoal data. J. Veg. Sci. 2014, 25, 1299–1310. [Google Scholar] [CrossRef]
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological maps for the main European woody species. Data Brief 2017, 12, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Ancillotto, L.; Mori, E.; Bosso, L.; Agnelli, P.; Russo, D. The Balkan long-eared bat (Plecotus kolombatovici) occurs in Italy-first confirmed record and potential distribution. Mamm. Biol. 2019, 96, 61–67. [Google Scholar] [CrossRef]
- Shiferaw, H.; Schaffner, U.; Bewket, W.; Alamirew, T.; Zeleke, G.; Teketay, D.; Eckert, S. Modelling the current fractional cover of an invasive alien plant and drivers of its invasion in a dryland ecosystem. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.L.; Bennett, J.R.; French, C.M. ENMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosso, L.; Ancillotto, L.; Smeraldo, S.; D’Arco, S.; Migliozzi, A.; Conti, P.; Russo, D. Loss of potential bat habitat following a severe wildfire: A model-based rapid assessment. Int. J. Wildland Fire 2018, 27, 756–769. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Smeraldo, S.; Bosso, L.; Fraissinet, M.; Bordignon, L.; Brunelli, M.; Ancillotto, L.; Russo, D. Modelling risks posed by wind turbines and power lines to soaring birds: The black stork (Ciconia nigra) in Italy as a case study. Biodivers. Conserv. 2020, 29, 1–18. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Raffini, F.; Bertorelle, G.; Biello, R.; D’Urso, G.; Russo, D.; Bosso, L. From Nucleotides to Satellite Imagery: Approaches to Identify and Manage the Invasive Pathogen Xylella fastidiosa and Its Insect Vectors in Europe. Sustainability 2020, 12, 4508. [Google Scholar] [CrossRef]
- Bertolino, S.; Sciandra, C.; Bosso, L.; Russo, D.; Lurz, P.W.; Di Febbraro, M. Spatially explicit models as tools for implementing effective management strategies for invasive alien mammals. Mammal Rev. 2020, 50, 187–199. [Google Scholar] [CrossRef]
- Iverson, L.R.; Rebbeck, J.; Peters, M.P.; Hutchinson, T.; Fox, T. Predicting Ailanthus altissima presence across a managed forest landscape in southeast Ohio. For. Ecosyst. 2019, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Sillero, N.; Poboljšaj, K.; Lešnik, A.; Šalamun, A. Influence of landscape factors on amphibian roadkills at the national level. Diversity 2019, 11, 13. [Google Scholar] [CrossRef] [Green Version]
- Zarzo-Arias, A.; Penteriani, V.; del Mar Delgado, M.; Torre, P.P.; Garcia-Gonzalez, R.; Mateo-Sánchez, M.C.; Garcia, P.V.; Dalerum, F. Identifying potential areas of expansion for the endangered brown bear (Ursus arctos) population in the Cantabrian Mountains (NW Spain). PLoS ONE 2019, 14, e0209972. [Google Scholar] [CrossRef] [PubMed]
- Fois, M.; Bacchetta, G.; Cuena-Lombrana, A.; Cogoni, D.; Pinna, M.S.; Sulis, E.; Fenu, G. Using extinctions in species distribution models to evaluate and predict threats: A contribution to plant conservation planning on the island of Sardinia. Environ. Conserv. 2018, 45, 11–19. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Wang, R.; Li, Q.; He, S.; Liu, Y.; Wang, M.; Jiang, G. Modeling and mapping the current and future distribution of Pseudomonas syringae pv. actinidiae under climate change in China. PLoS ONE 2018, 13, e0192153. [Google Scholar]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Leipold, M.; Tausch, S.; Poschlod, P.; Reisch, C. Species distribution modeling and molecular markers suggest longitudinal range shifts and cryptic northern refugia of the typical calcareous grassland species Hippocrepis comosa (horseshoe vetch). Ecol. Evol. 2017, 7, 1919–1935. [Google Scholar] [CrossRef] [Green Version]
- Roces-Díaz, J.V.; Jiménez-Alfaro, B.; Chytrý, M.; Díaz-Varela, E.R.; Álvarez-Álvarez, P. Glacial refugia and mid-Holocene expansion delineate the current distribution of Castanea sativa in Europe. Paleogeogr. Paleoclimatol. Paleoecol. 2018, 491, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Dakhil, M.A.; Xiong, Q.; Farahat, E.A.; Zhang, L.; Pan, K.; Pandey, B.; Olatunji, O.A.; Tariq, A.; Wu, X.; Zhang, A. Past and future climatic indicators for distribution patterns and conservation planning of temperate coniferous forests in southwestern China. Ecol. Indic. 2019, 107, 105559. [Google Scholar] [CrossRef]
- Paz, A.; González, A.; Crawford, A.J. Testing effects of Pleistocene climate change on the altitudinal and horizontal distributions of frogs from the Colombian Andes: A species distribution modeling approach. Front. Biogeogr. 2019, 11, e37055. [Google Scholar] [CrossRef] [Green Version]
- Archis, J.N.; Akcali, C.; Stuart, B.L.; Kikuchi, D.; Chunco, A.J. Is the future already here? The impact of climate change on the distribution of the eastern coral snake (Micrurus fulvius). PeerJ 2018, 6, e4647. [Google Scholar] [CrossRef]
- Jarnevich, C.S.; Hayes, M.A.; Fitzgerald, L.A.; Adams, A.A.Y.; Falk, B.G.; Collier, M.A.; Lea’R, B.; Klug, P.E.; Naretto, S.; Reed, R.N. Modeling the distributions of tegu lizards in native and potential invasive ranges. Sci. Rep. 2018, 8, 10193. [Google Scholar] [CrossRef] [PubMed]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elith, J.; Kearney, M.S.; Phillips, S.J. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Stoklosa, J.; Daly, C.; Foster, S.D.; Ashcroft, M.B.; Warton, D.I. A climate of uncertainty: Accounting for error in climate variables for species distribution models. Methods Ecol. Evol. 2015, 6, 412–423. [Google Scholar] [CrossRef]
- Costa, M.; Morla, C.; Sainz Ollero, H. Los Bosques Ibéricos: Una Interpretación Geobotánica; Iberian Forests: A Geobotanical Interpretation; Planeta: Madrid, Spain, 1997. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea Vol. 1; Cambridge University Press: Cambridge, UK, 1964. [Google Scholar]
- Bernetti, G. Selvicoltura Speciale; Unione Tipografico-Editrice Torinese: Torino, Italy, 1995. [Google Scholar]
- Finlayson, G.; Finlayson, C.; Pacheco, F.G.; Vidal, J.R.; Carrión, J.S.; Espejo, J.R. Caves as archives of ecological and climatic changes in the Pleistocene—The case of Gorham’s Cave, Gibraltar. Quat. Int. 2008, 181, 55–63. [Google Scholar] [CrossRef]
- Lambeck, K.; Purcell, A. Sea-level change in the Mediterranean Sea since the LGM: Model predictions for tectonically stable areas. Quat. Sci. Rev. 2005, 24, 1969–1988. [Google Scholar] [CrossRef]
- Di Donato, V.; Esposito, P.; Russo-Ermolli, E.; Scarano, A.; Cheddadi, R. Coupled atmospheric and marine paleoclimatic reconstruction for the last 35 ka in the Sele Plain–Gulf of Salerno area (southern Italy). Quat. Int. 2008, 190, 146–157. [Google Scholar] [CrossRef]
- Mix, A.C.; Bard, E.; Schneider, R. Environmental processes of the ice age: Land, oceans, glaciers (EPILOG). Quat. Sci. Rev. 2001, 20, 627–657. [Google Scholar] [CrossRef]
- Tzedakis, P.C. Seven ambiguities in the Mediterranean paleoenvironmental narrative. Quat. Sci. Rev. 2007, 26, 2042–2066. [Google Scholar] [CrossRef]
- Allen, J.R.; Huntley, B. Weichselian palynological records from southern Europe: Correlation and chronology. Quat. Int. 2000, 73, 111–125. [Google Scholar] [CrossRef]
- Ellenberg, H. Zeigerwerte der Gefäßpflanzen Mitteleuropas; Scripta Geobotanica IX; Erich Goltze KG: Göttingen, Germany, 1979. [Google Scholar]
- Bertolini, M.; Fedozzi, S.; Martini, F.; Sala, B. Late Glacial and Holocene climatic oscillations inferred from the variations in the micromammal associations at Grotta della Serratura (Marina di Camerota, Salerno, S. Italy). Quaternario 1996, 9, 561–566. [Google Scholar]
- Sala, B. Climatic changes in the Quaternary inferred from variations in the Mammal associations. Allionia 1996, 9, 89–94. [Google Scholar]
- Martini, F.; Colonese, A.C.; Giuseppe, Z.D.; Ghinassi, M.; Vetro, D.L.; Ricciardi, S. Human-environment relationships during the Late Glacial-Early Holocene transition: Some examples from Campania, Calabria and Sicily. Méditerranée 2009, 112, 89–94. [Google Scholar] [CrossRef]
- Follieri, M.; Giardini, M.; Magri, D.; Sadori, L. Palynostratigraphy of the last glacial period in the volcanic region of central Italy. Quat. Int. 1998, 47, 3–20. [Google Scholar] [CrossRef]
- Magri, D. Advances in Italian palynological studies: Late Pleistocene and Holocene records. GFF 2007, 129, 337–344. [Google Scholar] [CrossRef]
- Ricci Lucchi, M.R. Vegetation dynamics during the last Interglacial–Glacial cycle in the Arno coastal plain (Tuscany, western Italy): Location of a new tree refuge. Quat. Sci. Rev. 2008, 27, 2456–2466. [Google Scholar] [CrossRef]
- Fiorentino, G. L’analisi antracologica della sepoltura Ostuni 1 di S. Maria di Agnano: Considerazioni paleoambientali e paletnologiche. In Il Riparo di Agnano nel Paleolitico Superiore: La Sepoltura Ostuni 1 e Suoi Simboli; Coppola, D., Ed.; Università di Roma Tor Vergata: Rome, Italy, 2012; pp. 197–201. [Google Scholar]
- García-Amorena, I.; Manzaneque, F.G.; Rubiales, J.M.; Granja, H.M.; de Carvalho, G.S.; Morla, C. The Late Quaternary coastal forests of western Iberia: A study of their macroremains. Paleogeogr. Paleoclimatol. Paleoecol. 2007, 254, 448–461. [Google Scholar] [CrossRef]
- Hallé, F.; Oldemann, R.A.A.; Tomlinson, P.B. Tropical Trees and Forests: An Architectural Analysis; Springer: Berlin, Germany, 1978. [Google Scholar]
- Farjon, A. Pines: Drawings and Description of the Genus Pinus, 2nd ed.; Brill Academic Pub: Leiden, The Netherlands, 2018. [Google Scholar]
- Todaro, L.; Andreu, L.; D’Alessandro, C.M.; Gutiérrez, E.; Cherubini, P.; Saracino, A. Response of Pinus leucodermis to climate and anthropogenic activity in the National Park of Pollino (Basilicata, Southern Italy). Biol. Conserv. 2007, 137, 507–519. [Google Scholar] [CrossRef]
- Quézel, P.; Barbero, M. Signification phytoécologique et phytosociologique des peuplements naturels de Pin de Salzmann en France. Ecol. Mediterr. 1988, 14, 41–63. [Google Scholar] [CrossRef]
- Akman, Y. The vegetation of Beynam forest. Commun. Fac. Sci. Univ. Ank. 1972, 16, 31–54. [Google Scholar] [CrossRef]
- Emberger, L. La végétation de la region méditérranéenne. Essai d’une classification des groupements végétaux. Rev. Gen. Bot. 1930, 503, 642–662. [Google Scholar]
- Orombelli, G.; Ravazzi, C.; Cita, M.B. Osservazioni sul significato dei termini LGM (UMG), Tardoglaciale e postglaciale in ambito globale, italiano ed alpino. Quaternario 2005, 18, 147–156. [Google Scholar]
- Watts, W.A.; Allen, J.R.M.; Huntley, B.; Fritz, S.C. Vegetation history and climate of the last 15,000 years at Laghi di Monticchio, southern Italy. Quat. Sci. Rev. 1996, 15, 113–132. [Google Scholar] [CrossRef]
- Follieri, M.; Magri, D.; Sadori, L. 250,000-year pollen record from Valle di Castiglione (Roma). Pollen Spores 1988, 30, 329–356. [Google Scholar]
- Kelly, M.G.; Huntley, B. An 11 000-year record of vegetation and environment from Lago di Martignano, Latium, Italy. J. Quat. Sci. 1991, 6, 209–224. [Google Scholar] [CrossRef]
- Leroy, S.A.G.; Giralt, S.; Francus, P.; Seret, G. The high sensitivity of the palynological record in the Vico maar lacustrine sequence (Latium, Italy) highlights the climatic gradient through Europe for the last 90 ka. Quat. Sci. Rev. 1996, 15, 189–201. [Google Scholar] [CrossRef]
- Magri, D.; Sadori, L. Late Pleistocene and Holocene pollen stratigraphy at Lago di Vico, central Italy. Veg. Hist. Archaeobot. 1999, 8, 247–260. [Google Scholar] [CrossRef]
- Magri, D. Late Quaternary vegetation history at Lagaccione near Lago di Bolsena (central Italy). Rev. Paleobot. Palynol. 1999, 106, 171–208. [Google Scholar] [CrossRef]
- Nicol-Pichard, S.; Dubar, M. Reconstruction of late-glacial and holocene environments in southeast France based on the study of a 66-m long core from Biot, Alpes Maritimes. Veg. Hist. Archaeobot. 1998, 7, 11–15. [Google Scholar] [CrossRef]
- Colonese, A.C.; Zanchetta, G.; Fallick, A.E.; Martini, F.; Manganelli, G.; Drysdale, R.N. Stable isotope composition of Helix ligata (Müller, 1774) from Late Pleistocene–Holocene archaeological record from Grotta della Serratura (Southern Italy): Paleoclimatic implications. Glob. Planet. Chang. 2010, 71, 249–257. [Google Scholar] [CrossRef]
- Rubiales, J.M.; García-Amorena, I.; Hernández, L.; Génova, M.; Martínez, F.; Manzaneque, F.G.; Morla, C. Late Quaternary dynamics of pinewoods in the Iberian mountains. Rev. Paleobot. Palynol. 2010, 162, 476–491. [Google Scholar] [CrossRef]
- Carrión, J.S.; Finlayson, C.; Fernández, S.; Finlayson, G.; Allué, E.; López-Sáez, J.A.; López-García, P.; Gil-Romera, G.; Bailey, G.; González-Sampériz, P. A coastal reservoir of biodiversity for Upper Pleistocene human populations: Paleoecological investigations in Gorham’s Cave (Gibraltar) in the context of the Iberian Peninsula. Quat. Sci. Rev. 2008, 27, 2118–2135. [Google Scholar] [CrossRef]
- Carrión, J.S.; Yll, E.I.; Walker, M.J.; Legaz, A.J.; Chaín, C.; López, A. Glacial refugia of temperate, Mediterranean and Ibero-North African flora in south-eastern Spain: New evidence from cave pollen at two Neanderthal man sites. Glob. Ecol. Biogeogr. 2003, 12, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Huntley, B.; Birks, H.J.B. An Atlas of Past and Present Pollen Maps for Europe: 0–13000 Years Ago; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar]
- Tzedakis, P.C. Vegetation change through glacial—Interglacial cycles: A long pollen sequence perspective. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1994, 345, 403–432. [Google Scholar]
- Willis, K.; Mc Elwain, J. The Evolution of Plants; Oxford University Press: Oxford, UK, 2013. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Layer | Cultural Facies | 14C Dating (yr BP) | Calibrated Ages ± 2σ (cal yr BP) | Median cal yr BP | Chronostratigraphy |
|---|---|---|---|---|---|
| 8E | Final Epigravettian | 11,490 ± 160 | 13,708–13,057 | 13,332 | Late Glacial |
| 8F | 11,460 ± 80 | 13,455–13,135 | 13,305 | Late Glacial | |
| 8G | 12,060 ± 90 | 14,150–13,730 | 13,915 | Late Glacial | |
| 9 | Evolved Epigravettian | 13,100 ± 120 | 16,063–15,302 | 15,702 | Late Glacial |
| 10C | 15,700 ± 110 | 19,234–18,724 | 18,953 | Pleniglacial | |
| 11 | Gravettian | 24,380 ± 1530 | 32,574–25,925 | 28,874 | Pleniglacial |
| 12 | 29,020 ± 2650 | 43,290–28,645 | 34,269 | Pleniglacial |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Pasquale, G.; Saracino, A.; Bosso, L.; Russo, D.; Moroni, A.; Bonanomi, G.; Allevato, E. Coastal Pine-Oak Glacial Refugia in the Mediterranean Basin: A Biogeographic Approach Based on Charcoal Analysis and Spatial Modelling. Forests 2020, 11, 673. https://doi.org/10.3390/f11060673
Di Pasquale G, Saracino A, Bosso L, Russo D, Moroni A, Bonanomi G, Allevato E. Coastal Pine-Oak Glacial Refugia in the Mediterranean Basin: A Biogeographic Approach Based on Charcoal Analysis and Spatial Modelling. Forests. 2020; 11(6):673. https://doi.org/10.3390/f11060673
Chicago/Turabian StyleDi Pasquale, Gaetano, Antonio Saracino, Luciano Bosso, Danilo Russo, Adriana Moroni, Giuliano Bonanomi, and Emilia Allevato. 2020. "Coastal Pine-Oak Glacial Refugia in the Mediterranean Basin: A Biogeographic Approach Based on Charcoal Analysis and Spatial Modelling" Forests 11, no. 6: 673. https://doi.org/10.3390/f11060673