Decline and Passive Restoration of Forest Vegetation Around the Yeocheon Industrial Complex of Southern Korea


Abstract
:1. Introduction
2. Materials and Methods
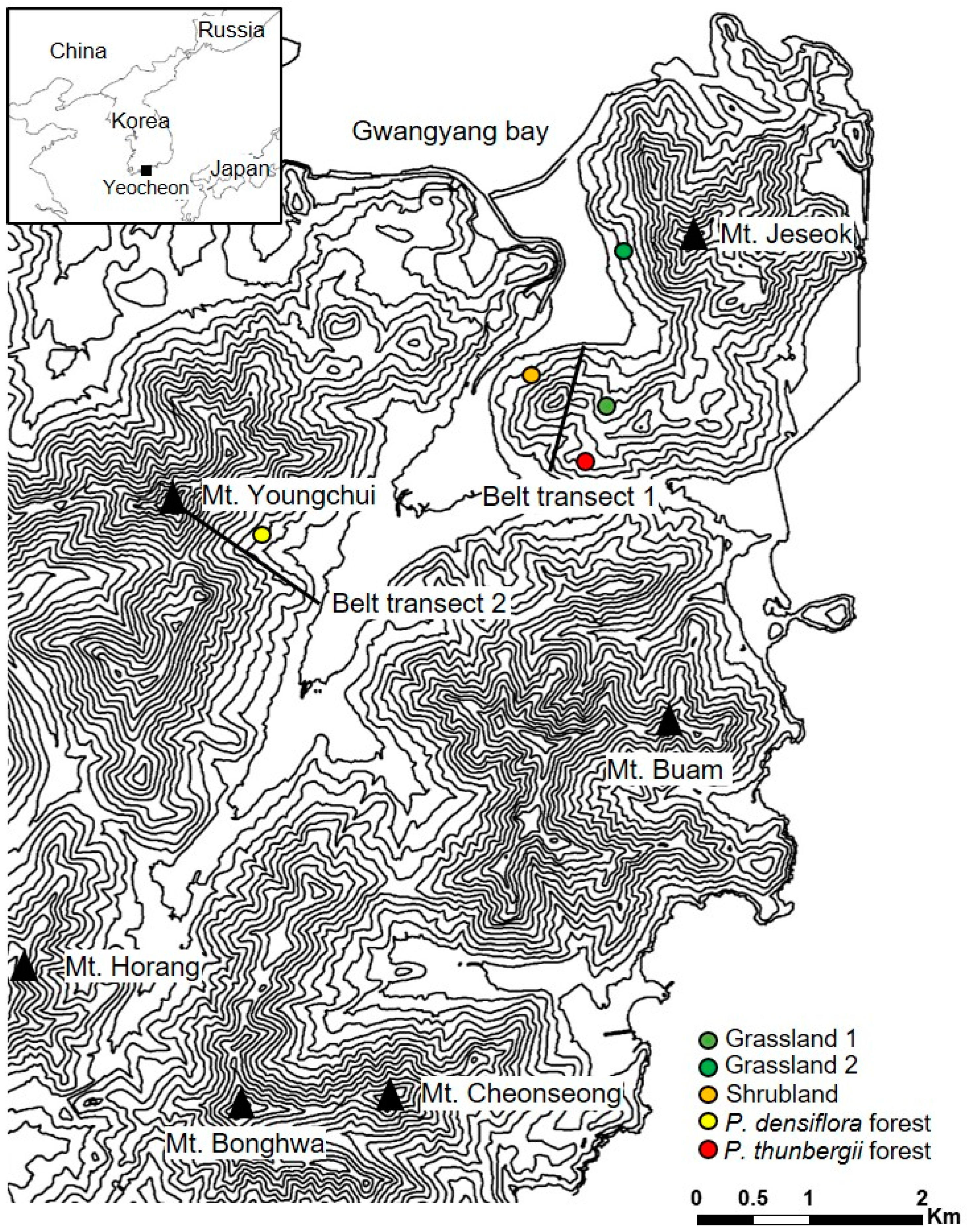
2.1. Study Area
2.2. Experimental Design
2.3. Vegetation Survey and Data Processing
2.4. Statistical Analyses
3. Results
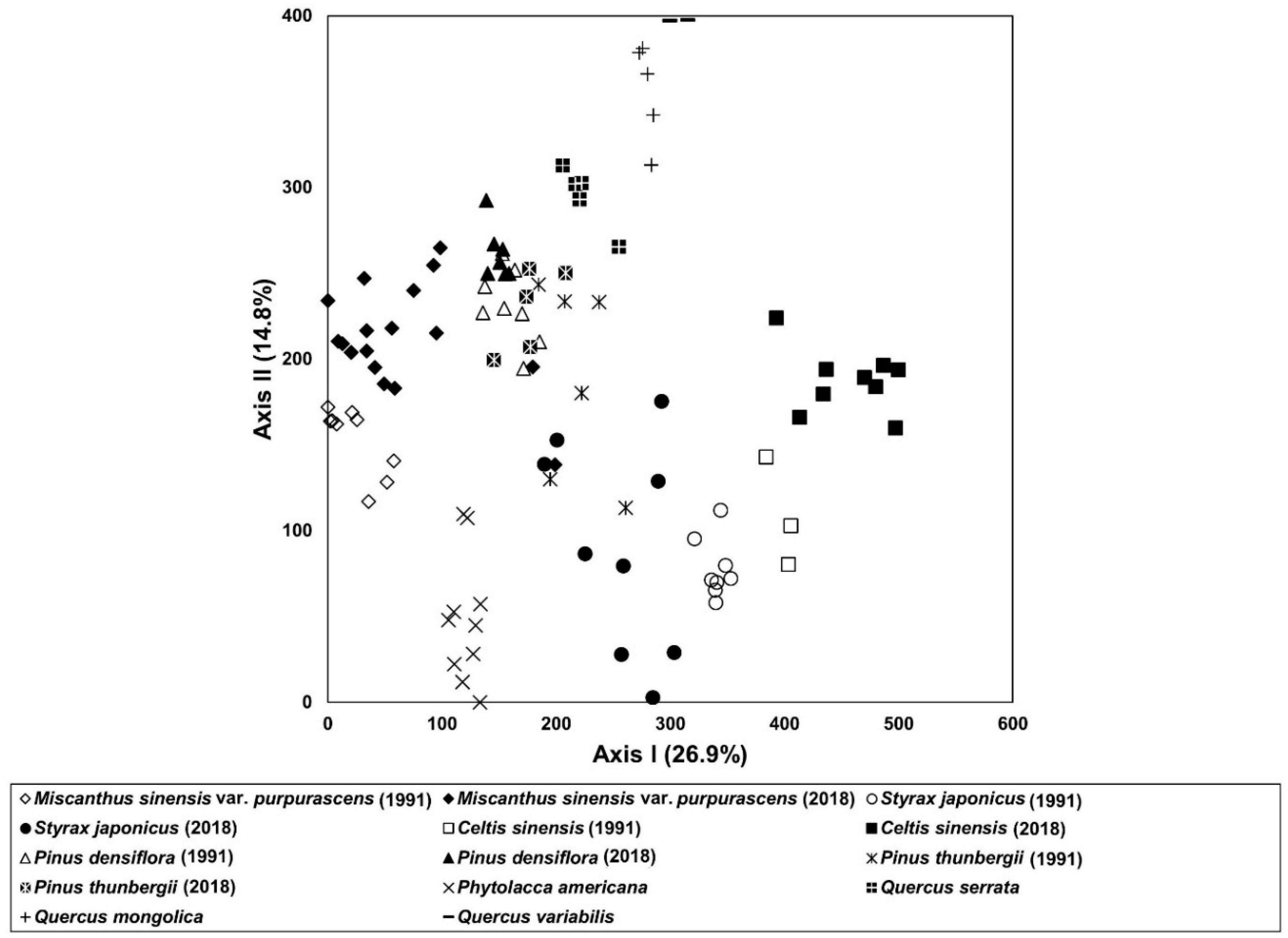
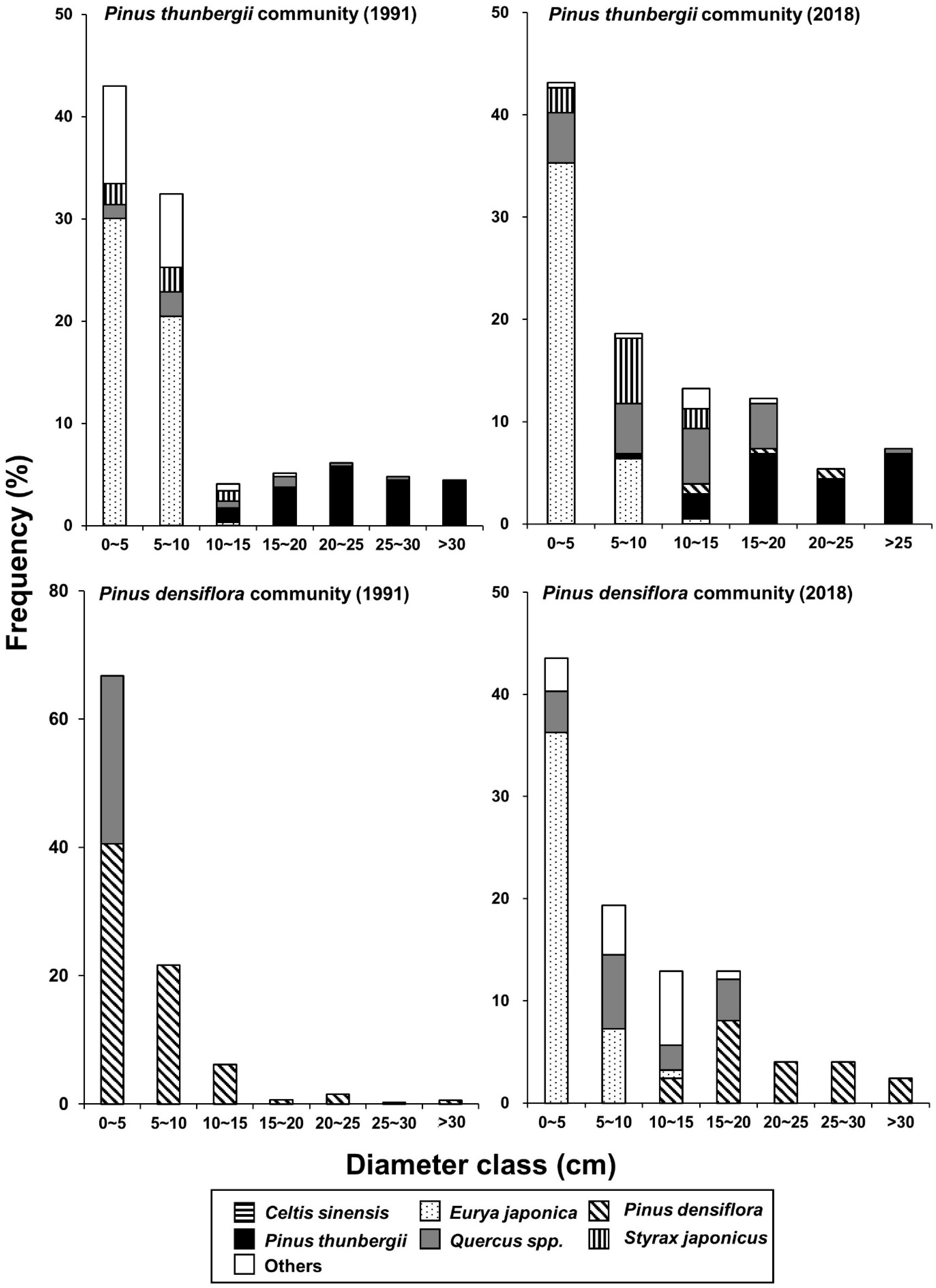
3.1. Vegetation Sequence
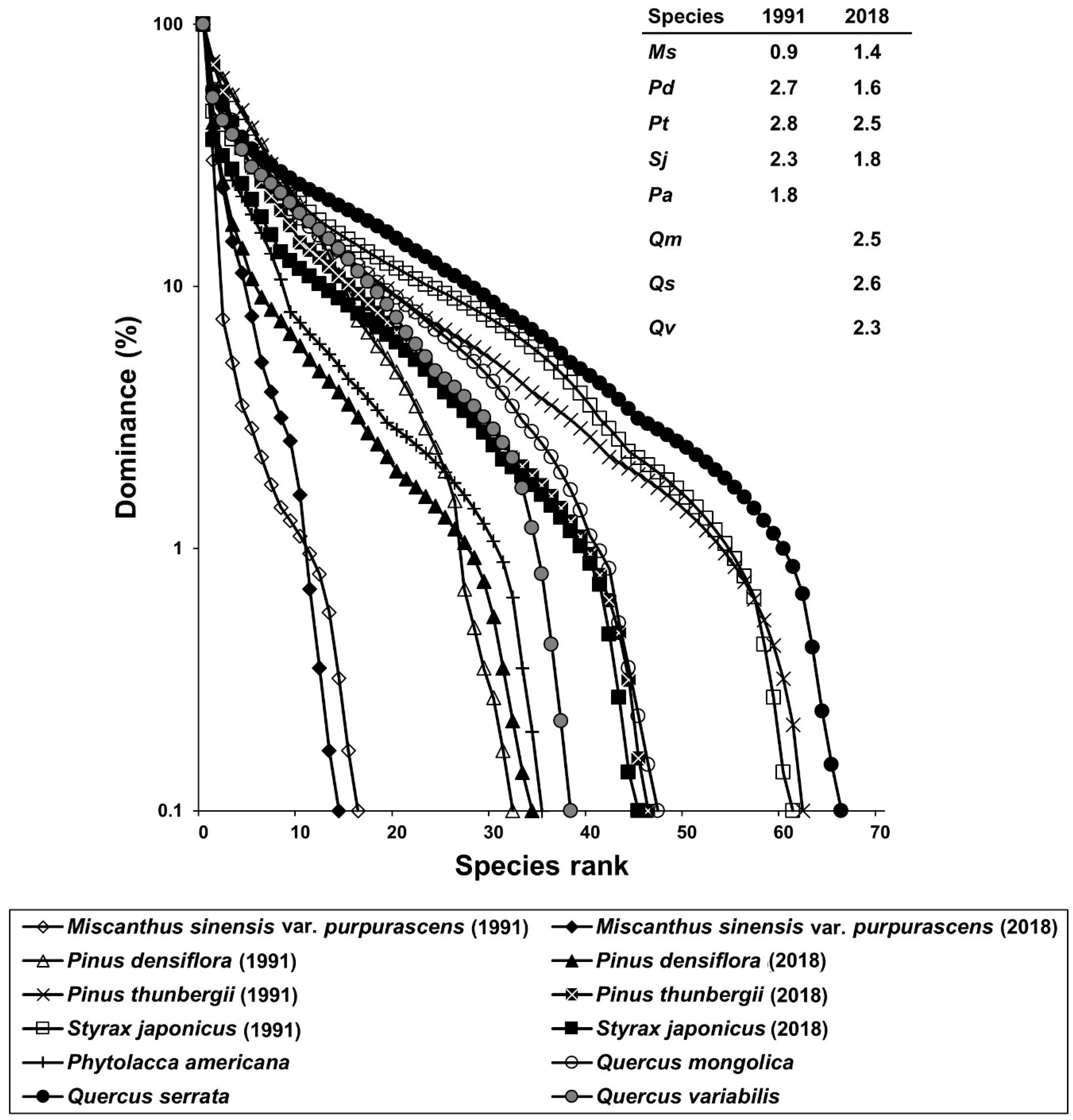
3.2. Species Diversity
3.3. Vegetation Dynamics
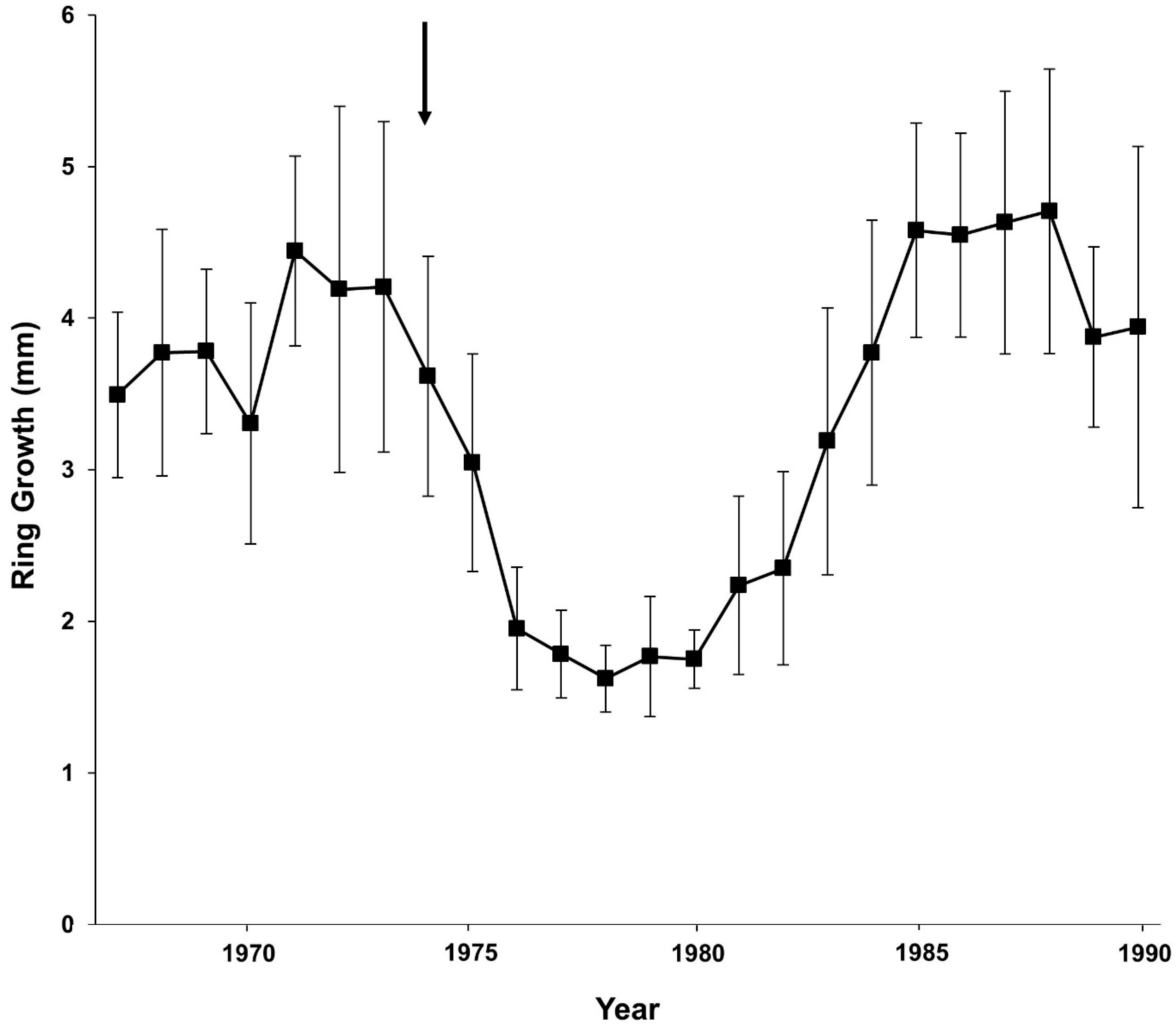
3.4. Yearly Changes in Annual Rings
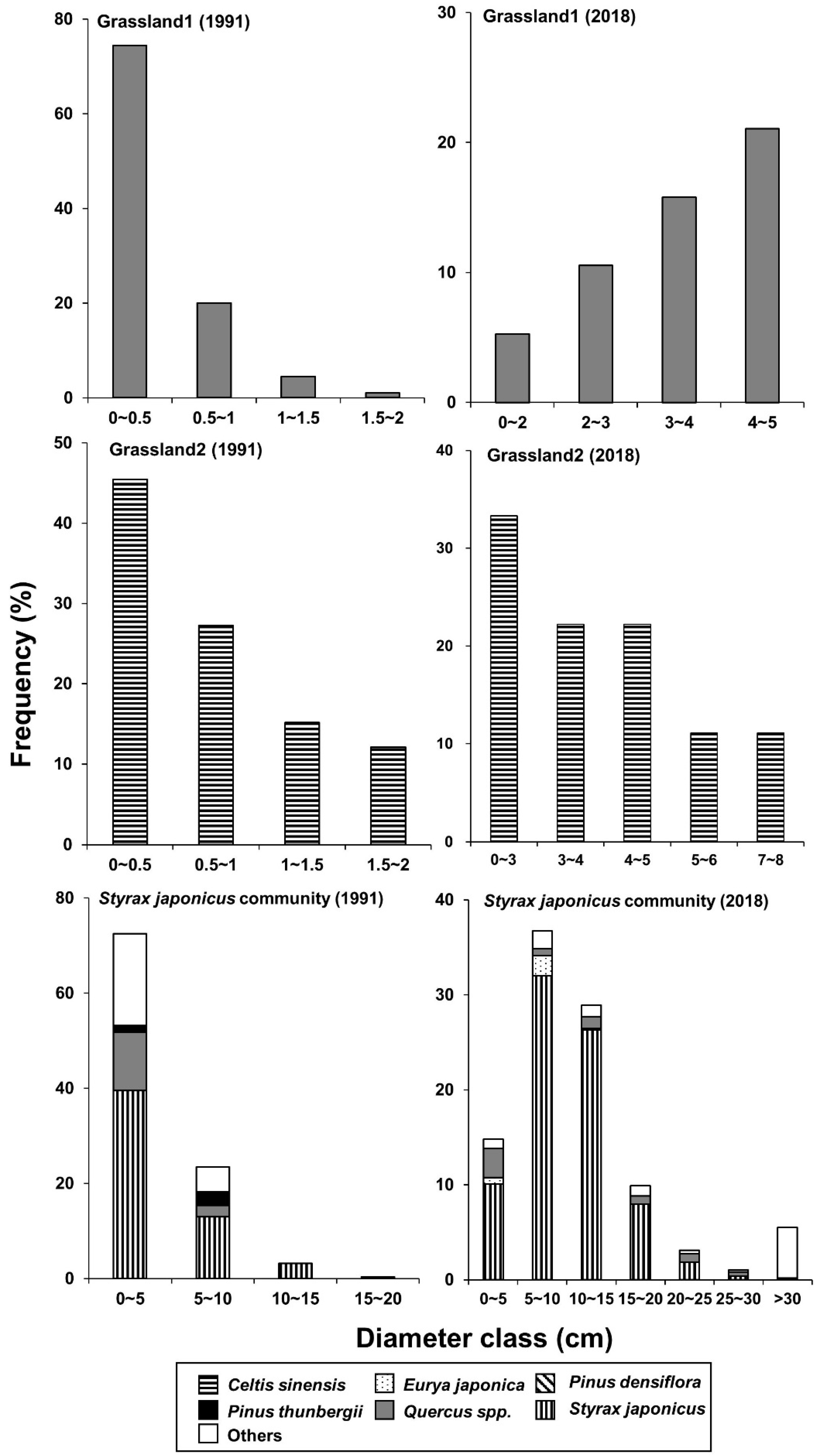
3.5. Age Distribution of Two Dominant Species
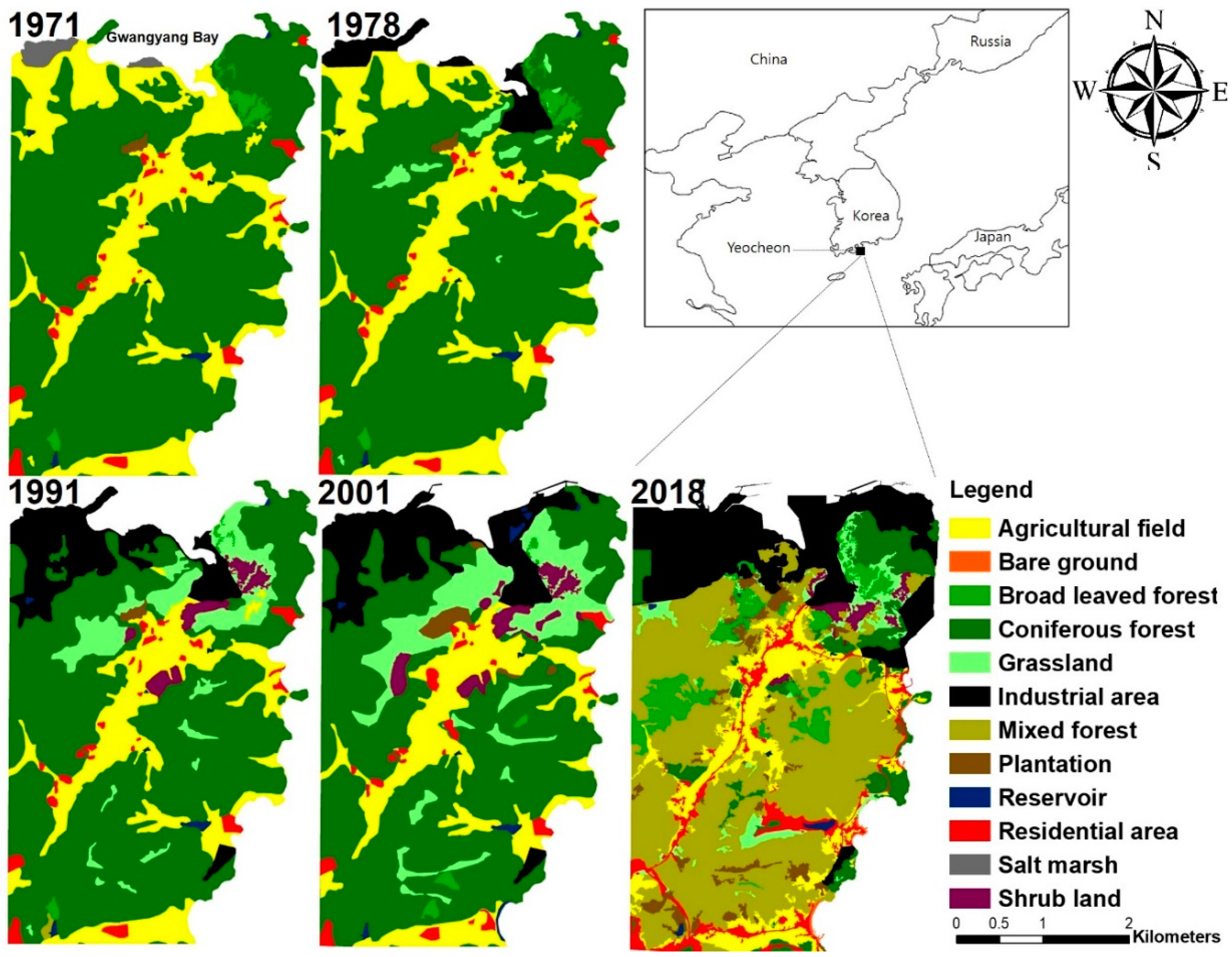
3.6. Landscape Structure and Change
4. Discussion
4.1. Characteristics and Establishment Background of Vegetation Around the Yeocheon Industrial Complex
4.2. Relationship Between Environmental Factors and Spatial Distribution of Vegetation
4.3. Future Prospects Based on Vegetation Dynamics
4.4. Implications for Restoration
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
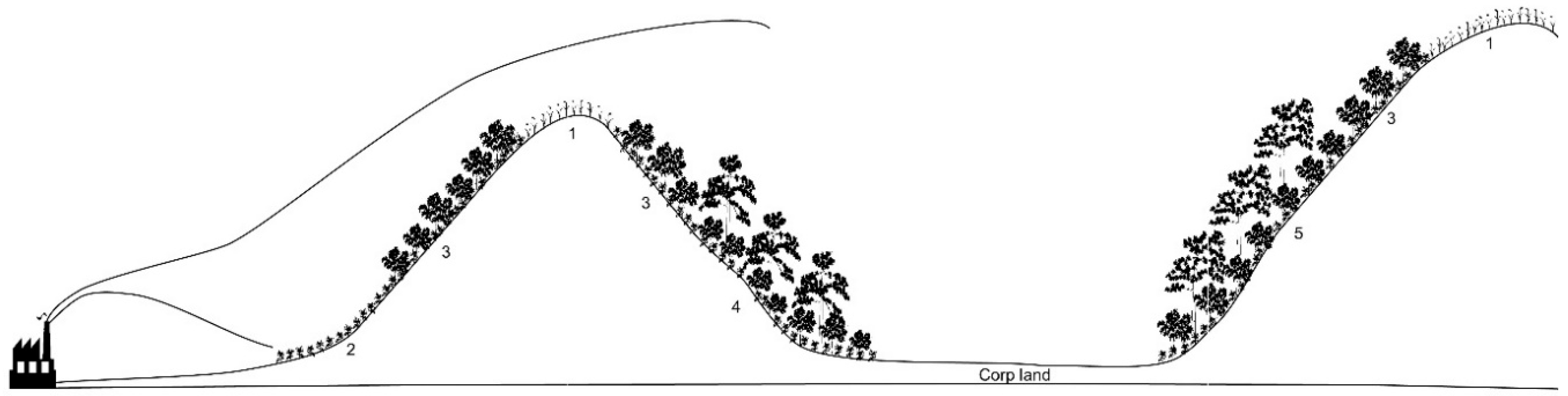
Appendix A. Landscape Change Progressed for the Last 50 Years around the Yeocheon Industrial Complex

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
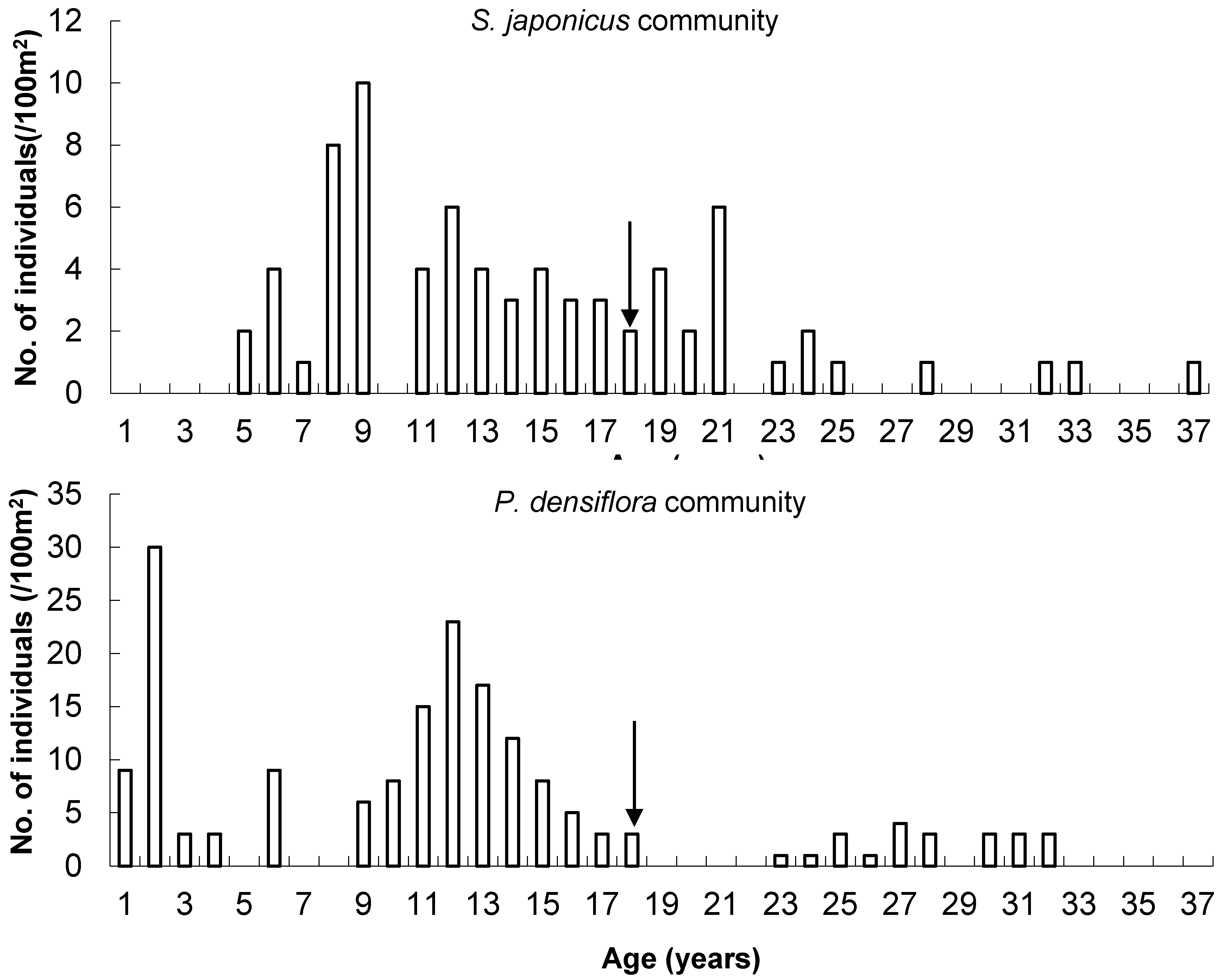{kind=link}
{kind=link}
{kind=link}
| Landscape Element | 1971 | 1978 | 1991 | 2001 | 2018 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| km2 | % | km2 | % | km2 | % | km2 | % | km2 | % | |
| Forest | ||||||||||
| Broad leaved forest | 0.6 | 1.1 | 0.6 | 1.1 | 0.2 | 0.3 | 0.3 | 0.6 | 3.6 | 6.3 |
| Coniferous forest | 38.5 | 71.4 | 37.8 | 69.9 | 33.2 | 61.0 | 30.4 | 53.7 | 4.6 | 8.1 |
| Mixed forest | - | - | - | - | 0.5 | 0.9 | - | - | 24.0 | 42.2 |
| Plantation | 0.1 | 0.3 | - | - | 0.1 | 0.3 | 0.6 | 1.0 | 1.9 | 3.3 |
| Shrub land | - | - | - | - | 1.1 | 2.0 | 1.5 | 2.7 | 0.7 | 1.3 |
| Grassland | - | - | 0.8 | 1.5 | 4.3 | 7.9 | 6.4 | 11.3 | 2.2 | 3.8 |
| Subtotal | 39.2 | 72.7 | 39.2 | 72.5 | 39.3 | 72.3 | 39.2 | 69.3 | 36.9 | 65.0 |
| Others | ||||||||||
| Agricultural field | 12.7 | 23.6 | 11.6 | 21.4 | 8.5 | 15.5 | 8.5 | 15.0 | 6.1 | 10.8 |
| Bare ground | - | - | - | - | 0.1 | 0.2 | ||||
| Industrial area | - | 1.9 | 3.6 | 5.3 | 9.8 | 6.8 | 12.1 | 10.4 | 18.4 | |
| Reservoir | 0.2 | 0.4 | 0.2 | 0.4 | 0.2 | 0.4 | 0.4 | 0.8 | 0.2 | 0.3 |
| Residential area | 1.2 | 2.2 | 1.2 | 2.2 | 1.1 | 2.1 | 1.6 | 2.9 | 3.0 | 5.3 |
| Salt marsh | 0.6 | 1.2 | - | - | - | - | ||||
| Subtotal | 14.7 | 27.3 | 14.9 | 27.5 | 15.1 | 27.7 | 17.3 | 30.7 | 19.9 | 35.0 |
| Total | 54.0 | 100.0 | 54.1 | 100.0 | 54.5 | 100.0 | 56.5 | 100.0 | 56.8 | 100.0 |
References
- Freedman, B. Environmental Ecology: The Ecological Effects of Pollution, Disturbance, and Other Stresses; Elsevier: Amsterdam, The Netherlands, 1995. [Google Scholar]
- Gunn, J.M. Restoration and Recovery of an Industrial Region: Progress in Restoring the Smelter-Damaged Landscape near Sudbury, Canada; Springer Science & Business Media: New York, NY, USA, 1995. [Google Scholar]
- Seinfeld, J.H.; Pandis, S.N. Atmospheric Chemistry and Physics: From Air Pollution to Climate Change, 3rd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Smith, W.H. Air Pollution and Forests: Interactions between Air Contaminants and Forest Ecosystems; Springer Science & Business Media: Berlin, Germany, 1990. [Google Scholar]
- Morecroft, M.D.; Cape, J.N.; Parr, T.W.; Brown, J.C.; Caporn, S.J.M.; Carroll, J.A.; Emmett, B.A.; Harmens, H.; Hill, M.O.; Lane, A.M.J.; et al. Monitoring the impacts of air pollution (acidification, eutrophication and ground-level ozone) on terrestrial habitats in the UK: A Scoping Study; Environment and Heritage Service: Belfast, Northern Ireland, 2005; 293p. [Google Scholar]
- Rockström, J.; Steffen, W.; Noone, K.; Persson, Å.; Chapin, F.S., III; Lambin, E.; Lenton, T.M.; Scheffer, M.; Folke, C.; Schellnhuber, H.J. Planetary boundaries: Exploring the safe operating space for humanity. Ecol. Soc. 2009, 14, 32. [Google Scholar] [CrossRef]
- Gheorghe, I.F.; Ion, B. The effects of air pollutants on vegetation and the role of vegetation in reducing atmospheric pollution. In The Impact of Air Pollution on Health, Economy, Environment and Agricultural Sources; Khallaf, M., Ed.; InTech: London, UK, 2011; pp. 241–280. ISBN 978-953-307-528-0. [Google Scholar]
- Kelly, F.J.; Fussell, J.C. Air pollution and public health: Emerging hazards and improved understanding of risk. Environ. Geochem. Health 2015, 37, 631–649. [Google Scholar] [CrossRef] [Green Version]
- Spiegel, J.; Maystre, L.Y. Environmental pollution control and prevention. In Encyclopaedia of Occupational Health and Safety; International Labour Office: Geneva, Switzerland, 1998; Volume 4, Chapter 55- Environmental Pollution Control. [Google Scholar]
- Pirrone, N.; Cinnirella, S.; Feng, X.; Finkelman, R.B.; Friedli, H.R.; Leaner, J.; Mason, R.; Mukherjee, A.B.; Stracher, G.B.; Streets, D. Global mercury emissions to the atmosphere from anthropogenic and natural sources. Atmos. Chem. Phys. 2010, 10. [Google Scholar]
- Labordena, M.; Neubauer, D.; Folini, D.; Patt, A.; Lilliestam, J. Blue skies over China: The effect of pollution-control on solar power generation and revenues. PLoS ONE 2018, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Perera, F. Pollution from fossil-fuel combustion is the leading environmental threat to global pediatric health and equity: Solutions exist. Int. J. Environ. Res. Public Health 2018, 15, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ministry of Environment. Annual Report of Ambient Air Quality in Korea. Available online: http://library.me.go.kr (accessed on 17 February 2020).
- Seinfeld, J.H.; Pandis, S.N. Atmospheric Chemistry and Physics: From Air Pollution to Climate Change, 1st ed.; John Wiley & Sons: Hoboken, NJ, USA, 1998. [Google Scholar]
- Kato, N.; Akimoto, H. Anthropogenic emissions of SO2 and NOx in Asia: Emission inventories. Atmos. Environ. Part A Gen. Top. 1992, 26, 2997–3017. [Google Scholar] [CrossRef]
- Lee, C.S.; Cho, Y.C.; Shin, H.C. Effects of restoration practiced by introducing tolerant plants to the industrially degraded forest ecosystem around the Ulsan, Korea. In Proceedings of the 16th International Conference, Society for Ecological Restoration International, Victoria, BC, Canada, 25–27 August 2004; pp. 1–11. [Google Scholar]
- Lee, C.S.; Lee, K.S.; Hwangbo, J.K.; You, Y.H.; Kim, J.H. Selection of tolerant plants and their arrangement to restore a forest ecosystem damaged by air pollution. J Water Air Soil Pollut. 2004, 156, 251–273. [Google Scholar] [CrossRef]
- Lee, C.S.; Moon, J.S.; Cho, Y.C. Effects of soil amelioration and tree planting on restoration of an air-pollution damaged forest in south Korea. Water Air Soil Pollut. 2007, 179, 239–254. [Google Scholar] [CrossRef]
- Legge, A.H.; Krupa, S.V. Air Pollutants and Their Effects on the Terrestrial Ecosystem; John Wiley & Sons Inc.: Hoboken, NJ, USA, 1986. [Google Scholar]
- Darrall, N. The effect of air pollutants on physiological processes in plants. Plant Cell Environ. 1989, 12, 1–30. [Google Scholar] [CrossRef]
- Council, N.R. Improving Risk Communication; National Academies: Washington, DC, USA, 1989. [Google Scholar]
- Paoletti, E.; Bytnerowicz, A.; Andersen, C.; Augustaitis, A.; Ferretti, M.; Grulke, N.; Günthardt-Goerg, M.S.; Innes, J.; Johnson, D.; Karnosky, D. Impacts of air pollution and climate change on forest ecosystems—Emerging research needs. Sci. World J. 2007, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Moore, B.; Allard, G. Abiotic disturbances and their influence on forest health: A review. In Forest Health and Biosecurity Working Paper; Forestry Department, Food and Agriculture Organization: Rome, Italy, 2011; p. 44. [Google Scholar]
- Kharuk, V. Air pollution impacts on subarctic forests at Noril’sk, Siberia. In Forest Dynamics in Heavily Polluted Regions; Report No. 1 of the IUFRO Task Force on Environmental Change; Innes, J.L., Oleksyn, J., Eds.; CABI: Wallingford, UK, 2000; pp. 77–86. [Google Scholar]
- Rigina, O.; Kozlov, M. The impacts of air pollution on the northern taiga forests of the Kola Peninsula, Russian Federation. In Forest Dynamics in Heavily Polluted Region; Report No. 1 of the IUFRO Task Force on Environmental Change; Innes, J.L., Oleksyn, J., Eds.; CABI: Wallingford, UK, 2000; pp. 37–65. [Google Scholar]
- Winterhalder, K. Landscape degradation by smelter emissions near Sudbury, Canada, and subsequent amelioration and restoration. In Forest Dynamics in Heavily Polluted Regions; Report No. 1 of the IUFRO Task Force on Environmental Change; Innes, J.L., Oleksyn, J., Eds.; CABI: Wallingford, UK, 2000; pp. 87–119. [Google Scholar]
- Luttermann, A.; Freedman, B. Risks to forests in heavily polluted regions. In Forest Dynamics in Heavily Polluted Regions; Report No. 1 of the IUFRO Task Force on Environmental Change; Innes, J.L., Oleksyn, J., Eds.; CABI: Wallingford, UK,, 2000; pp. 9–26. [Google Scholar]
- Cooke, S.J.; Sack, L.; Franklin, C.E.; Farrell, A.P.; Beardall, J.; Wikelski, M.; Chown, S.L. What is conservation physiology? Perspectives on an increasingly integrated and essential science. Conserv. Physiol. 2013, 1, cot001. [Google Scholar] [CrossRef] [PubMed]
- Freedman, B. Ecological effects of environmental stressors. In Oxford Research Encyclopedia of Environmental Science; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Kozlowski, T. Impacts of air pollution on forest ecosystems. BioScience 1980, 30, 88–93. [Google Scholar] [CrossRef]
- Tilman, D.; Lehman, C. Human-caused environmental change: Impacts on plant diversity and evolution. Proc. Natl. Acad. Sci. USA 2001, 98, 5433–5440. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.S.; Cho, Y.C. Selection of pollution-tolerant trees for restoration of degraded forests and evaluation of the experimental restoration practices at the Ulsan Industrial Complex, Korea. In Ecology, Planning, and Management of Urban Forests; Carreiro, M.M., Song, Y.C., Wu, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 369–392. [Google Scholar]
- Kim, G.S.; Pee, J.H.; An, J.H.; Lim, C.H.; Lee, C.S. Selection of air pollution tolerant plants through the 20-years-long transplanting experiment in the Yeocheon industrial area, southern Korea. Anim. Cells Syst. 2015, 19, 208–215. [Google Scholar] [CrossRef] [Green Version]
- Acid Deposition Monitoring Network in East Asia (EANET); Task Force on Research Coordination (TFRC); Scientific Advisory Committee (SAC). Review on the State of Air Pollution in East Asia; United Nations Environment Programme Asia and the Pacific: Bangkok, Thailand, 2015; p. 430. [Google Scholar]
- Lee, C.S.; Jung, S.; Lim, B.S.; Kim, A.R.; Lim, C.H.; Lee, H. Forest decline under progress in the urban forest of Seoul, Central Korea. In Deforestation around the World; Mohd, N.S., Zulkiflee, A.L., Gabriel De, O., Nathaniel, B., Yosio, S., Carlos, A.C., Eds.; IntechOpen: London, UK, 2019. [Google Scholar]
- Hallgren, J.E. Physiological and biochemical effects of sulfur dioxide on plants. In Sulfur in the Environment Part 2; Willey: New York, NY, USA, 1978; pp. 163–209. [Google Scholar]
- You, Y.H.; Lee, C.S.; Kim, J.H. Selection of tolerant species among Korean major woody plants to restore Yeocheon Industrial Complex Area. Korean J. Ecol. 1998, 21, 337–344. [Google Scholar]
- Woo, S.Y.; Lee, D.K.; Lee, Y.K. Net photosynthetic rate, ascorbate peroxidase and glutathione reductase activities of Erythrina orientalis in polluted and non-polluted areas. Photosynthetica 2007, 45, 293–295. [Google Scholar] [CrossRef]
- Baek, S.G.; Woo, S.Y. Physiological and biochemical responses of two tree species in urban areas to different air pollution levels. Photosynthetica 2010, 48, 23–29. [Google Scholar] [CrossRef]
- Kramer, P.; Kozlowski, T. Physiology of Woody Plants; Academic Press: New York, NY, USA, 1979. [Google Scholar]
- Cox, R. Sensitivity of forest plant reproduction to long range transported air pollutants: In vitro and in vivo sensitivity of Oenothera parviflora L. pollen to simulated acid rain. J. New Phytol. 1984, 97, 63–70. [Google Scholar] [CrossRef]
- Lovett, G.M.; Tear, T.H.; Evers, D.C.; Findlay, S.E.; Cosby, B.J.; Dunscomb, J.K.; Driscoll, C.T.; Weathers, K.C. Effects of air pollution on ecosystems and biological diversity in the eastern United States. Ann. N. Y. Acad. Sci. 2009, 1162, 99–135. [Google Scholar] [CrossRef] [PubMed]
- Lal, N. Effects of acid rain on plant growth and development. J. Sci. Technol. 2016, 11, 85–108. [Google Scholar]
- Treshow, M. Interaction of air pollutants and plant diseases. In Responses Plants Air Pollut. Brian, M.J., Kozlowski, T.T., Eds.; Academic Press: New York, NY, USA, 1975; pp. 307–334. [Google Scholar]
- Williams, W.T. Air pollution diseases in the Californian forests. A base line for smog disease on pondeeosa and Jeffrey pines in the Sequoia and Los Padres National Forests, California. Environ. Sci. Technol. 1980, 14, 179–182. [Google Scholar] [CrossRef]
- Chakraborty, S.; Tiedemann, A.; Teng, P.S. Climate change: Potential impact on plant diseases. Environ. Pollut. 2000, 108, 317–326. [Google Scholar] [CrossRef]
- Percy, K.E.; Awmack, C.S.; Lindroth, R.L.; Kubiske, M.E.; Kopper, B.J.; Isebrands, J.; Pregitzer, K.S.; Hendrey, G.R.; Dickson, R.E.; Zak, D.R. Altered performance of forest pests under atmospheres enriched by CO2 and O3. Nature 2002, 420, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.H. Communities and Ecosystems; Macmillan: New York, NY, USA, 1975; Volume 2. [Google Scholar]
- Taoda, H. Effect of urbanization on the evergreen broadleaf forest in Tokyo, Japan. Bull. Yokohama Phytosociol. Soc. Jpn. 1979, 16, 161–165. [Google Scholar]
- Lee, C.S.; Cho, Y.C.; Lee, A.N. Restoration planning for the Seoul metropolitan area, Korea. In Ecology, Planning, and Management of Urban Forests; Springer: Berlin/Heidelberg, Germany, 2008; pp. 393–419. [Google Scholar]
- Lee, C.S.; Chun, Y.M.; Lee, H.; Pi, J.H.; Lim, C.H. Establishment, Regeneration, and Succession of Korean Red Pine (Pinus densiflora S. et Z.) Forest in Korea. In Conifers; IntechOpen: London, UK, 2018. [Google Scholar]
- Kercher, J.; Axelrod, M.; Bingham, G. Forecasting Effects of S02 Pollution on Growth and Succession in a Western Conifer Forest. In Proceedings of the Symposium on Effects of Air Pollutants on Mediterranean and Temperate Forest Ecosystems, Riverside, CA, USA, 22–27 June 1980; p. 200. [Google Scholar]
- Jean-Pierre, G. What is the impact of air pollutants on vegetation? Encycl. Environ. 2019. Life-Nature under Tension, ISSN 2555-0950. Available online: https://www.encyclopedie-environnement.org/en/life/impact-air-pollutants-on-vegetation (accessed on 17 February 2020).
- Korea Forest Service. Forest Geographic Information System. Available online: http://www.forest.go.kr/newkfsweb/html/HtmlPage.do?pg=/fgis/UI_KFS_5002_020200.html&mn=KFS_02_04_03_04_02&orgld=fgis (accessed on 21 March 2019).
- Korea Meteorological Administration. Automatic Weather System (AWS). Available online: http://data.kma.go.kr/data/grnd/selectAwsRItmList.do?pgmNo=56 (accessed on 21 March 2019).
- Oh, K.C. A Survey Report on Vegetation before Construction of the Petrochemical Industrial Complex in the Yeosu Region; Korea Institute of Science and Technology: Seoul, Korea, 1979. [Google Scholar]
- National Institute of Ecology, National Ecosystem Survey. Available online: https://www.nie-ecobank.kr/spceinfo/main.do (accessed on 14 February 2020).
- Ministry of Environment. Selection and Breeding of Tolerant Species and Bio-Indicator to Air Pollution and Acid Rain; Ministry of Environment: Seoul, Korea, 1996; p. 353.
- Ministry of Environment. Korea National Long-Term Ecological Research: Final Reports. Ministry of Environment: Seoul, Korea, 2013. [Google Scholar]
- Kim, G.S.; Lim, C.H.; Lim, Y.K.; Jung, S.H.; Lee, C.S. Diagnostic assessment on vegetation damage due to hydrofluoric gas leak accident and restoration planning to mitigate the damage in a forest ecosystem around Hube Globe in Gumi. J. Wetl. Res. 2015, 17, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Ellenberg, D.; Mueller-Dombois, D. Aims and Methods of Vegetation Ecology; Wiley: New York, NY, USA, 1974. [Google Scholar]
- Korea National Arboretum (KNA). Korean Plant Names Index. Available online: http://www.nature.go.kr/main/Main.do (accessed on 13 February 2020).
- Braun-Blanquet, J. Pflanzensoziologie, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 1964. [Google Scholar]
- Hill, M. Decorana—A Fortran program for detrended correspondence analysis and reciprocal averaging. Vegetatio 1979, 42, 47–58. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Kent, M.; Coker, P. Vegetation Description and Analysis: A Practical Approach; John Wiley & Sons: Hoboken, NJ, USA, 1992. [Google Scholar]
- Lee, C.S.; You, Y.H.; Robinson, G.R. Secondary succession and natural habitat restoration in abandoned rice fields of central Korea. Restor. Ecol. 2002, 10, 306–314. [Google Scholar] [CrossRef]
- Wright, S.J.; Muller-Landau, H.C.; Condit, R.; Hubbell, S.P. Gap-dependent recruitment, realized vital rates, and size distributions of tropical trees. Ecology 2003, 84, 3174–3185. [Google Scholar] [CrossRef]
- White, E.P.; Ernest, S.M.; Kerkhoff, A.J.; Enquist, B.J. Relationships between body size and abundance in ecology. Trends Ecol. Evol. 2007, 22, 323–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deb, P.; Sundriyal, R. Tree regeneration and seedling survival patterns in old-growth lowland tropical rainforest in Namdapha National Park, north-east India. For. Ecol. Manag. 2008, 255, 3995–4006. [Google Scholar] [CrossRef]
- Westphal, C.; Tremer, N.; von Oheimb, G.; Hansen, J.; von Gadow, K.; Härdtle, W. Is the reverse J-shaped diameter distribution universally applicable in European virgin beech forests? For. Ecol. Manag. 2006, 223, 75–83. [Google Scholar] [CrossRef]
- Peet, R.K. Forest vegetation of the Colorado front range. Vegetatio 1981, 45, 3–75. [Google Scholar] [CrossRef]
- Dyakov, N.R. Successional Pattern, Stand Structure and Regeneration of Forest Vegetation According to Local Environmental Gradients. Ecol. Balk. 2013, 5, 1. [Google Scholar]
- Lykke, A.M. Assessment of species composition change in savanna vegetation by means of woody plants’ size class distributions and local information. Biodivers. Conserv. 1998, 7, 1261–1275. [Google Scholar] [CrossRef]
- Bin, Y.; Ye, W.; Muller-Landau, H.C.; Wu, L.; Lian, J.; Cao, H. Unimodal tree size distributions possibly result from relatively strong conservatism in intermediate size classes. PLoS ONE 2012, 7, e52596. [Google Scholar] [CrossRef] [Green Version]
- Chew, R.M.; Chew, A.E. The primary productivity of a desert-shrub (Larrea tridentata) community. Ecol. Monogr. 1965, 35, 355–375. [Google Scholar] [CrossRef]
- Barbour, M.G.; Burk, J.H.; Pitts, W.D. Terrestrial Plant Ecology; Benjamin/Cummings: San Francisco, CA, USA, 1999; Volume 3. [Google Scholar]
- Lee, C. Regeneration of Pinus densiflora community around the Yeocheon industrial complex disturbed by air pollution. Korean J. Ecol. 1993, 16, 305–316. [Google Scholar]
- Statistical Package for the Social Sciences, IBM SPSS Statistics 24; International Business Machines Co.: New York, NY, USA, 2020.
- Ter Braak, C.J. CANOCO—A FORTRAN Program for Canonical Community Ordination by [partial][etrended][canonical] Correspondence Analysis, Principal Components Analysis and Redundancy Analysis (version 2.1); MLV: Wageningen, The Netherlands, 1988. [Google Scholar]
- Nriagu, J.O. Environmental Impacts of Smelters; Wiley: New York, NY, USA, 1984. [Google Scholar]
- Pryde, P.R. Environmental Management in the Soviet Union; Cambridge University Press: Cambridge, UK, 1991; Volume 4. [Google Scholar]
- Peterson, D. Troubled Lands: The Legacy of Soviet Environmental Destruction; Westview Press: Boulder, CO, USA, 1993. [Google Scholar]
- Hansen, J.R.; Hansson, R.; Norris, S. The State of the European Arctic Environment; Office for Official Publications of the European Communities: Copenhagen, Denmark, 1996. [Google Scholar]
- Bormann, F. The effects of air pollution on the New England landscape. Ambio 1982, 11, 338–346. [Google Scholar]
- National Institute of Environmental Research (NIER). Studies on the Accumulation of Environmental Pollutants and its Correlation with Tree Growth in the Industrial Complexes; National Institute of Environmental Research: Incheon, Korea, 1984. [Google Scholar]
- National Institute of Environmental Research (NIER). A Study on the Evaluation and the Prevention of Damage to Plant Communities by Air Pollution (Ⅱ): A Comparison of the Structure and Productivity of Plant Communities around Industrial Complexes and an Unpolluted Area; National Institute of Environmental Research: Incheon, Korea, 1990; Volume 12, pp. 181–235. [Google Scholar]
- Larcher, W. Physiological Plant Ecology; Springer: Berlin, Germany, 1975. [Google Scholar]
- Rydval, M.; Wilson, R. The impact of industrial SO2 pollution on north Bohemia conifers. Water Air Soil Pollut. 2012, 223, 5727–5744. [Google Scholar] [CrossRef]
- Bai, L.; Wang, J.; Ma, X.; Lu, H. Air pollution forecasts: An overview. Int. J. Environ. Res. Public Health 2018, 15, 780. [Google Scholar] [CrossRef] [Green Version]
- Gordon, A.G.; Gorham, E. Ecological aspects of air pollution from an iron-sintering plant at Wawa, Ontario. Can. J. Bot. 1963, 41, 1063–1078. [Google Scholar] [CrossRef]
- Sousa, W.P. The role of disturbance in natural communities. Ann. Rev. Ecol. Syst. 1984, 15, 353–391. [Google Scholar] [CrossRef]
- Lee, C.; Hong, S. Landscape ecological perspectives in the structure and dynamics of fire-disturbed vegetation in a rural landscape, eastern Korea. J Landsc. Ecol. Appl. Land Eval. Dev. Conserv. Itc Publ. 2001, 81–94. [Google Scholar]
- An, J.H.; Lim, C.H.; Cho, Y.C.; Lee, C.S. Early recovery process and restoration planning of burned pine forests in central eastern Korea. J. For. Res. 2019, 30, 243–255. [Google Scholar] [CrossRef]
- Woodwell, G.M. Effects of pollution on the structure and physiology of ecosystems. Science 1970, 168, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.W.; Hauer, R.J.; Werner, L.P. Urban Forestry: Planning and Managing Urban Greenspaces; Waveland press: Long Grove, IL, USA, 1998. [Google Scholar]
- Oh, W.S.; Lee, C.S. Recovery of Ecosystem Service Functions through Ecological Restoration Practice: A Case Study of Coal Mine Spoils, Samcheok, Central Eastern Korea. J. Korean J. Environ. Biol. Anim. Cells 2014, 32, 102–111. [Google Scholar] [CrossRef]
- SERI (Society Ecological Restoration International Science & Policy Working Group). The SER International Primer on Ecological Restoration; Society for Ecological Restoration International: Tucson, Arizona, USA, 2004. [Google Scholar]
- McDonald, T.; Gann, G.; Jonson, J.; Dixon, K. International Standards for the Practice of Ecological Restoration–Including Principles and Key Concepts; Society for Ecological Restoration: Washington, DC, USA, 2016. [Google Scholar]
- Bradshaw, A. Ecological principles and land reclamation practice. J. Landsc. Plan. 1984, 11, 35–48. [Google Scholar] [CrossRef]
- Kamiyama, A.; Masashi, A.; Kondo, M.; Yoshimizu, K. Advanced Greening Technology; Soft Science Co: Tokyo, Japan, 1989; p. 291. [Google Scholar]
- Zahawi, R.A.; Reid, J.L.; Holl, K.D. Hidden costs of passive restoration. Restor. Ecol. 2014, 22, 284–287. [Google Scholar] [CrossRef]
- Prach, K.; del Moral, R. Passive restoration is often quite effective: Response to Zahawi et al. (2014). Restor. Ecol. 2015, 23, 344–346. [Google Scholar] [CrossRef]









| Period | Mean (mm) | Standard Deviation (mm) | t-Value | p |
|---|---|---|---|---|
| Suppressed (1974 to 1985) | 2.64 | 1.49 | 6.99 | 0.00 |
| Non-suppressed (other period) | 4.12 | 1.71 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Lim, B.S.; Kim, D.U.; Kim, A.R.; Seol, J.W.; Lim, C.H.; Kil, J.H.; Moon, J.S.; Lee, C.S. Decline and Passive Restoration of Forest Vegetation Around the Yeocheon Industrial Complex of Southern Korea. Forests 2020, 11, 674. https://doi.org/10.3390/f11060674
Lee H, Lim BS, Kim DU, Kim AR, Seol JW, Lim CH, Kil JH, Moon JS, Lee CS. Decline and Passive Restoration of Forest Vegetation Around the Yeocheon Industrial Complex of Southern Korea. Forests. 2020; 11(6):674. https://doi.org/10.3390/f11060674
Chicago/Turabian StyleLee, Hansol, Bong Soon Lim, Dong Uk Kim, A Reum Kim, Jae Won Seol, Chi Hong Lim, Ji Hyun Kil, Jeong Sook Moon, and Chang Seok Lee. 2020. "Decline and Passive Restoration of Forest Vegetation Around the Yeocheon Industrial Complex of Southern Korea" Forests 11, no. 6: 674. https://doi.org/10.3390/f11060674