Cationic Dendrimer G2-S16 Inhibits Herpes Simplex Type 2 Infection and Protects Mice Vaginal Microbiome

 , and
, and 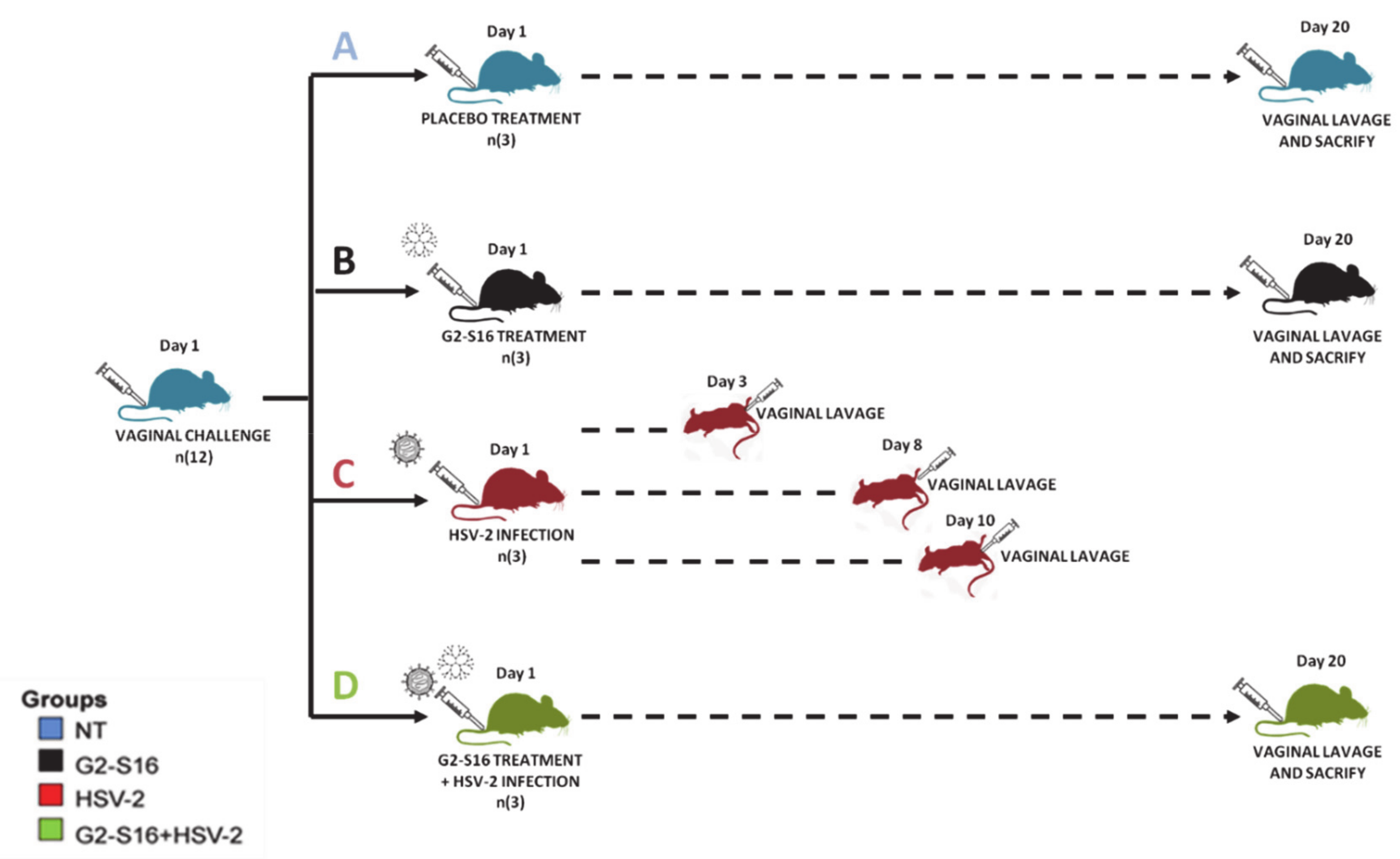{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Dendrimer, Cell Culture Semen Samples, and Virus Strain
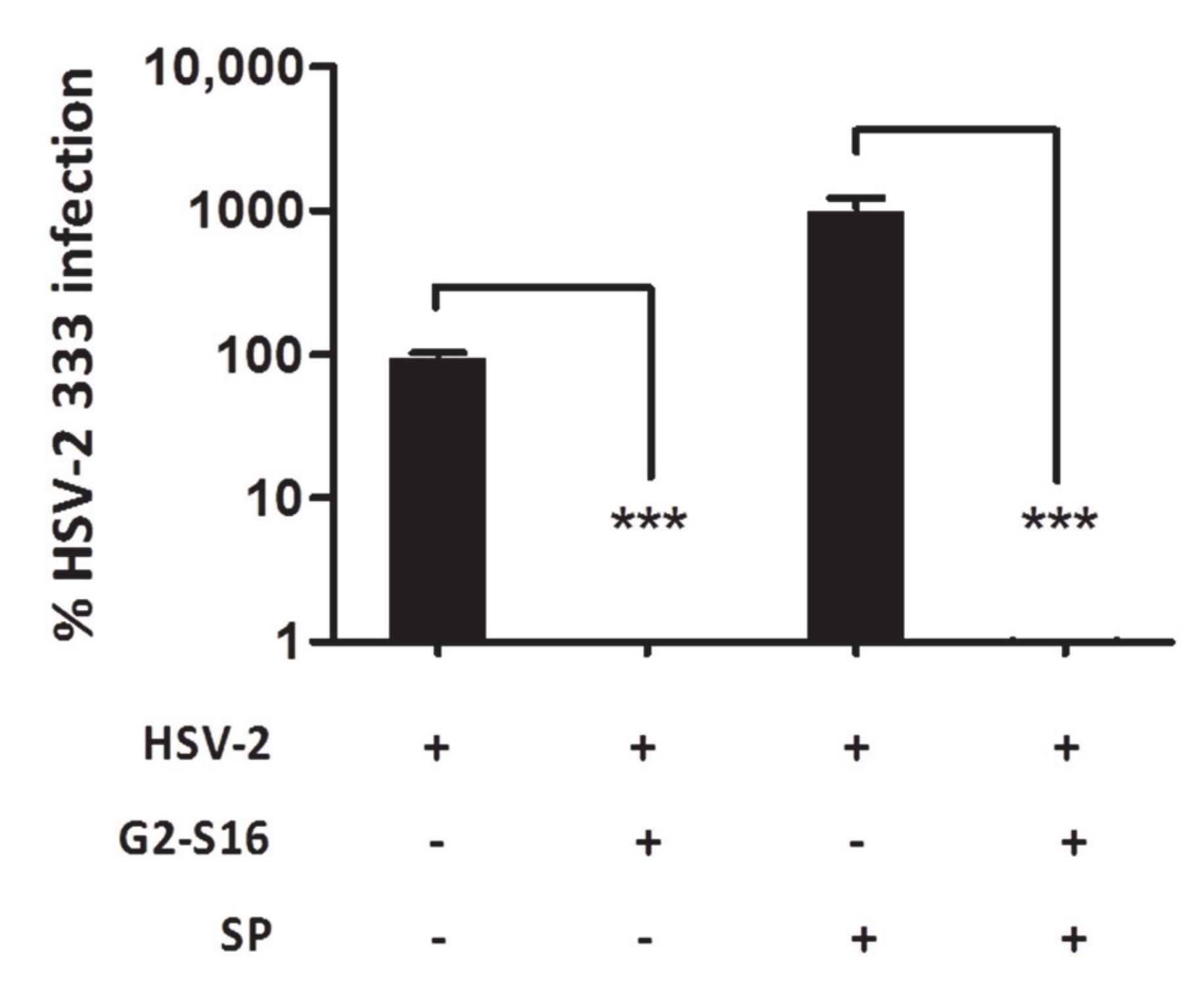
2.2. Efficacy of G2-S16 Polyanionic Carbosilane Dendrimer against HSV-2 Infection in Presence of Semen
2.3. Experimental Mice In Vivo Design
2.4. DNA Extraction, Microbiome Sequencing, and Sequence Analysis
2.5. Statistical Analysis
3. Results
3.1. Inhibition of G2-S16 Polyanionic Carbosilane Dendrimer in Presence of Semen
3.2. Experimental Design and Vaginal Microbiome Analyses
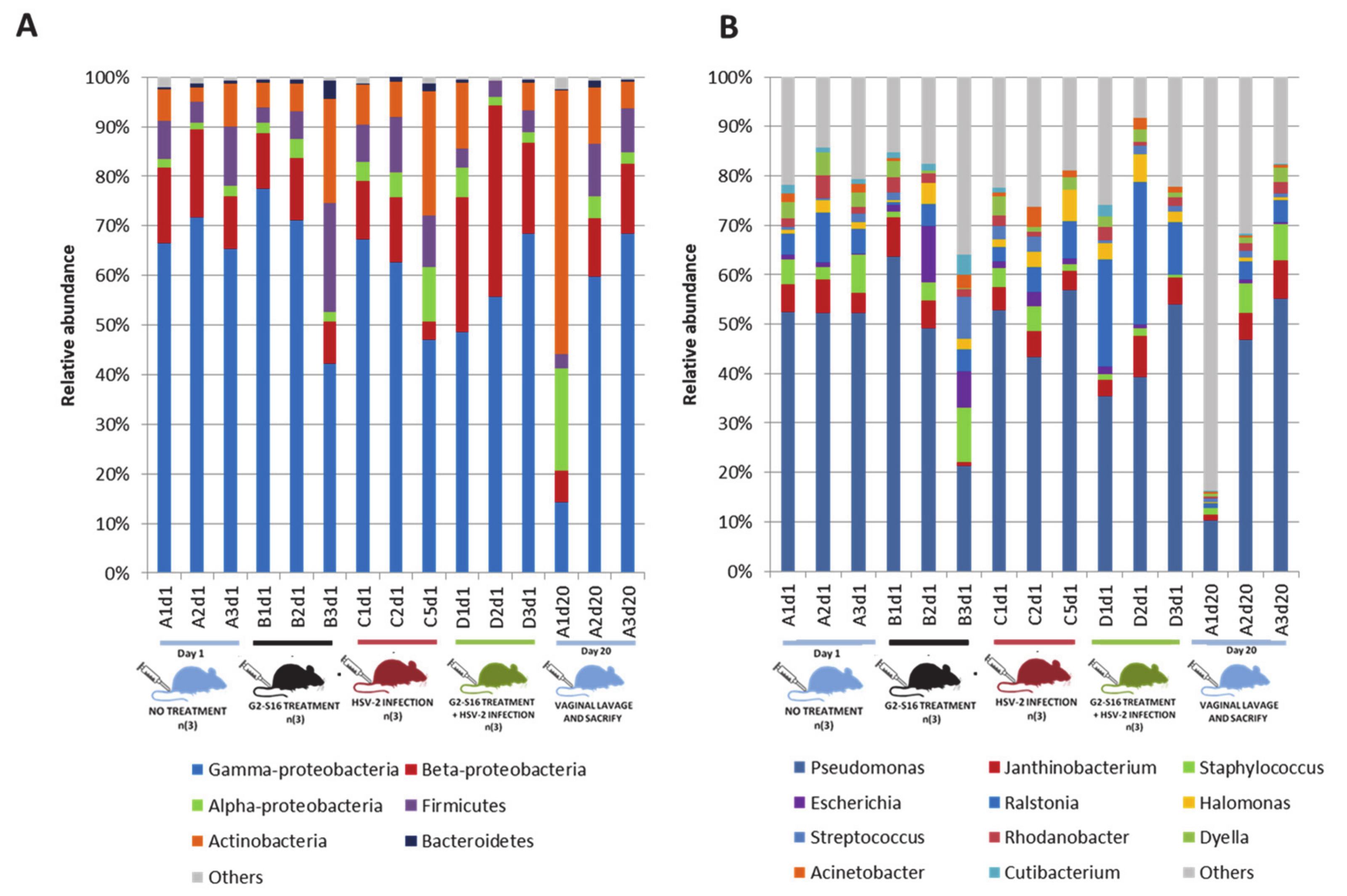
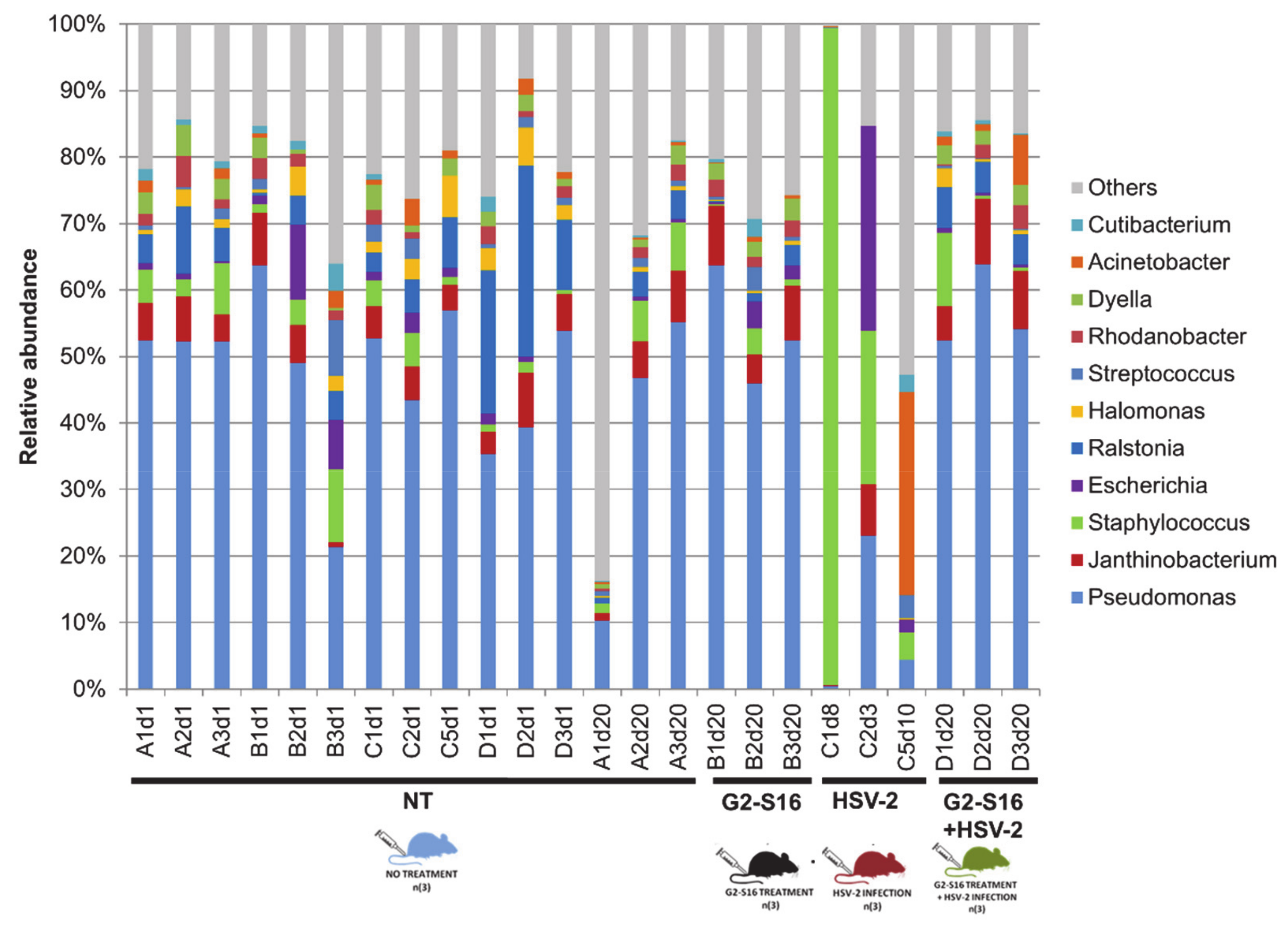
3.3. Microbial Community of Healthy Female Mice Vagina
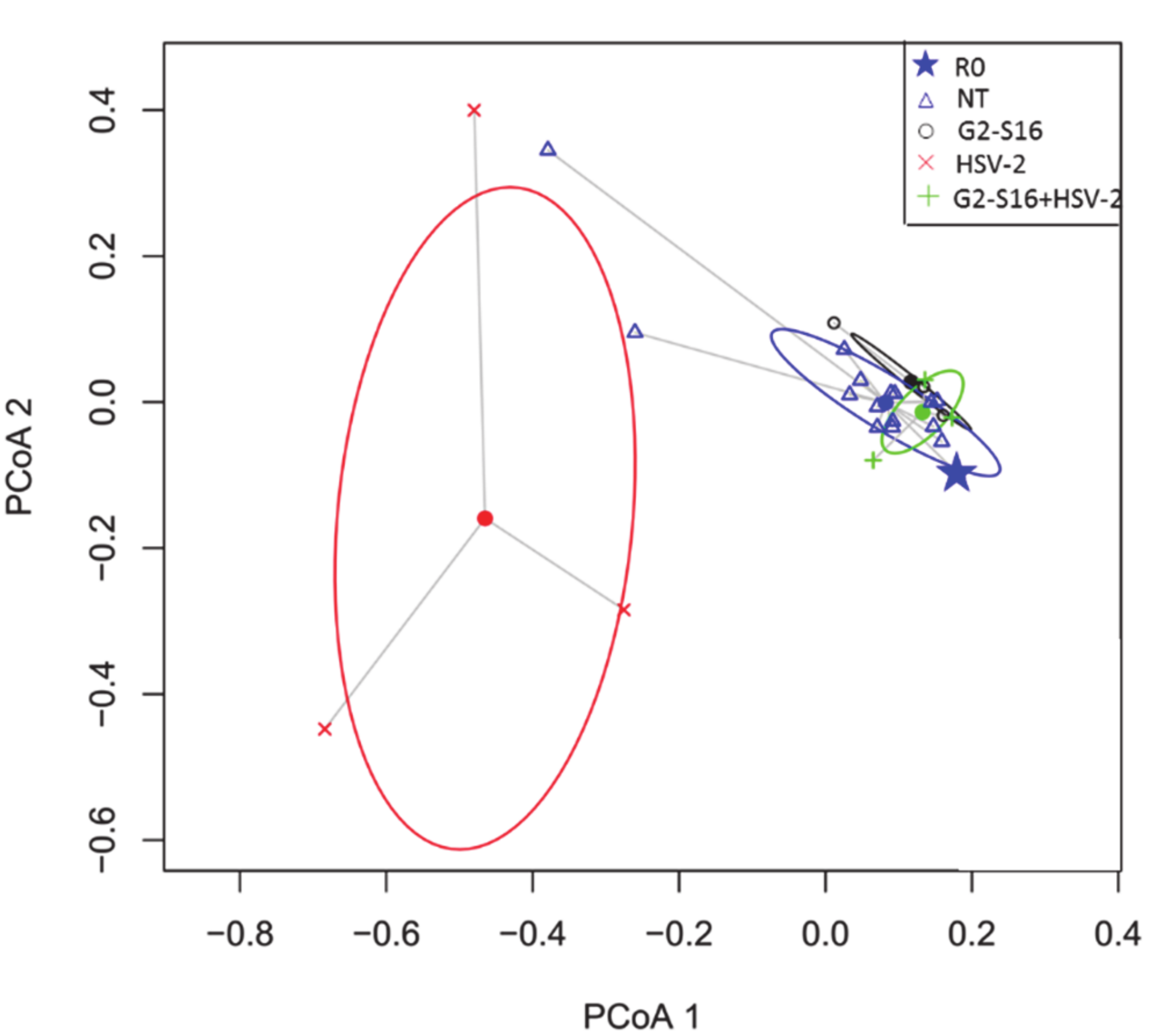
3.4. Variability of Microbial Composition in HSV-2 Infection
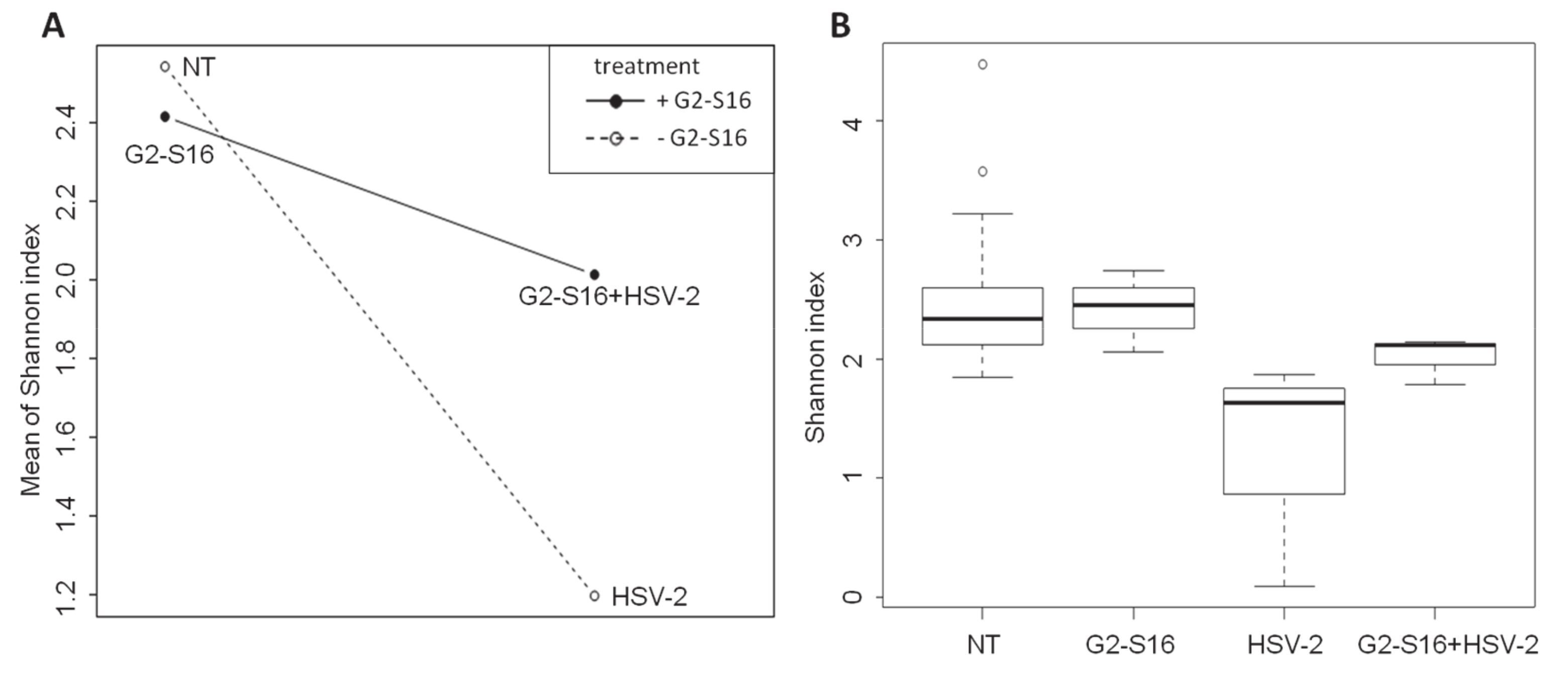
3.5. Effect of the G2-S16 Polyanionic Carbosilane Dendrimer on Microbial Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| STIs | Sexual transmission infections |
| BV | Bacterial vaginosis |
| TFV | Tenofovir |
| HSV-2 | Herpes simplex virus type 2 |
| HIV-1 | Human immunodeficiency virus type 1 |
| BLT | Bone-liver-thymus |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| FBS | Fetal bovine serum |
| IgG | Human immunoglobulin G |
| PFU | Plaque-forming unit |
| MOI | Multiplicity of infection |
| PBS | Phosphate buffer saline |
| HEC | Hydroxyethyl-cellulose |
| NT | Nontreated |
| SP | Seminal plasma |
| PCoA | Principal coordinate analysis |
| HCV | Hepatitis C virus |
References
- Grant, R.; Hamer, D.; Hope, T.; Johnston, R.; Lange, J.; Lederman, M.M.; Lieberman, J.; Miller, C.J.; Moore, J.P.; Mosier, N.E.; et al. Whither or Wither Microbicides? Science 2008, 321, 532–534. [Google Scholar] [CrossRef] [Green Version]
- Karim, Q.A.; Karim, S.S.A.; Frohlich, J.A.; Grobler, A.C.; Baxter, C.; Mansoor, L.E.; Kharsany, A.B.M.; Sibeko, S.; Mlisana, K.; Omar, Z.; et al. Effectiveness and Safety of Tenofovir Gel, an Antiretroviral Microbicide, for the Prevention of HIV Infection in Women. Science 2010, 329, 1168–1174. [Google Scholar] [CrossRef] [Green Version]
- Zirafi, O.; Kim, K.-A.; Roan, N.R.; Kluge, S.F.; Müller, J.A.; Jiang, S.; Mayer, B.; Greene, W.C.; Kirchhoff, F.; Münch, J. Semen enhances HIV infectivity and impairs the antiviral efficacy of microbicides. Sci. Transl. Med. 2014, 6, 262ra157. [Google Scholar] [CrossRef] [Green Version]
- Klatt, N.R.; Cheu, R.; Birse, K.; Zevin, A.S.; Perner, M.; Noël-Romas, L.; Grobler, A.C.; Westmacott, G.; Xie, I.Y.; Butler, J.; et al. Vaginal bacteria modify HIV tenofovir microbicide efficacy in African women. Science 2017, 356, 938–945. [Google Scholar] [CrossRef] [Green Version]
- Velloza, J.; Heffron, R. The Vaginal Microbiome and its Potential to Impact Efficacy of HIV Pre-exposure Prophylaxis for Women. Curr. HIV/AIDS Rep. 2017, 14, 153–160. [Google Scholar] [CrossRef]
- Krasnopolsky, V.N.; Prilepskaya, V.N.; Polatti, F.; Zarochentseva, N.V.; Bayramova, G.R.; Caserini, M.; Palmieri, R. Efficacy of Vitamin C Vaginal Tablets as Prophylaxis for Recurrent Bacterial Vaginosis: A Randomised, Double-Blind, Placebo-Controlled Clinical Trial. J. Clin. Med. Res. 2013, 5, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Brotman, R.M. Vaginal microbiome and sexually transmitted infections: An epidemiologic perspective. J. Clin. Investig. 2011, 121, 4610–4617. [Google Scholar] [CrossRef]
- Eastment, M.C.; McClelland, R.S. Vaginal microbiota and susceptibility to HIV. AIDS 2018, 32, 687–698. [Google Scholar] [CrossRef]
- Petrova, M.I.; Broek, M.V.D.; Balzarini, J.; Vanderleyden, J.; Lebeer, S. Vaginal microbiota and its role in HIV transmission and infection. FEMS Microbiol. Rev. 2013, 37, 762–792. [Google Scholar] [CrossRef] [Green Version]
- Atashili, J.; Poole, C.; Ndumbe, P.M.; Adimora, A.A.; Smith, J.S. Bacterial vaginosis and HIV acquisition: A meta-analysis of published studies. AIDS 2008, 22, 1493–1501. [Google Scholar] [CrossRef] [Green Version]
- Achkar, J.M.; Fries, B.C. Candida Infections of the Genitourinary Tract. Clin. Microbiol. Rev. 2010, 23, 253–273. [Google Scholar] [CrossRef] [Green Version]
- Cherpes, T.L.; Melan, M.; Kant, J.A.; Cosentino, L.A.; Meyn, L.A.; Hillier, S.L. Genital Tract Shedding of Herpes Simplex Virus Type 2 in Women: Effects of Hormonal Contraception, Bacterial Vaginosis, and Vaginal Group B Streptococcus Colonization. Clin. Infect. Dis. 2005, 40, 1422–1428. [Google Scholar] [CrossRef]
- Nardis, C.; Mosca, L.; Mastromarino, P. Vaginal microbiota and viral sexually transmitted diseases. Ann. Ig. 2013, 25, 443–456. [Google Scholar]
- Jaramillo-Ruiz, D.; De La Mata, F.J.; Gomez, R.; Correa-Rocha, R.; Muñoz-Fernández, M. Ángeles Nanotechnology as a New Therapeutic Approach to Prevent the HIV-Infection of Treg Cells. PLoS ONE 2016, 11, e0145760. [Google Scholar] [CrossRef]
- Ceña-Diez, R.; García-Broncano, P.; De La Mata, F.J.; Gomez-Ramirez, R.; Resino, S.; Muñoz-Fernández, M. Ángeles G2-S16 dendrimer as a candidate for a microbicide to prevent HIV-1 infection in women. Nanoscale 2017, 9, 9732–9742. [Google Scholar] [CrossRef]
- Rasines, B.; Sánchez-Nieves, J.; Maiolo, M.; Malý, M.; Chonco, L.; Jimenez, J.L.; Muñoz-Fernández, M. Ángeles; De La Mata, F.J.; Gómez, R. Synthesis, structure and molecular modelling of anionic carbosilane dendrimers. Dalton Trans. 2012, 41, 12733. [Google Scholar] [CrossRef]
- Muñoz-Fernández, M.A.; Ceña-Diez, R.; Vacas-Córdoba, E.; García-Broncano, P.; De La Mata, F.J.; Gomez, R.; Malý, M. Prevention of vaginal and rectal herpes simplex virus type 2 transmission in mice: Mechanism of antiviral action. Int. J. Nanomed. 2016, 11, 2147–2162. [Google Scholar] [CrossRef] [Green Version]
- Sepúlveda-Crespo, D.; Ceña-Diez, R.; Jiménez, J.L.; Muñoz-Fernández, M. Ángeles Mechanistic Studies of Viral Entry: An Overview of Dendrimer-Based Microbicides As Entry Inhibitors Against Both HIV and HSV-2 Overlapped Infections. Med. Res. Rev. 2016, 37, 149–179. [Google Scholar] [CrossRef]
- Sepúlveda-Crespo, D.; Mj, S.; Tager, A.M.; Vrbanac, V.; Gomez-Ramirez, R.; De La Mata, F.J.; Jiménez, J.L.; Muñoz-Fernández, M. Ángeles Prevention vaginally of HIV-1 transmission in humanized BLT mice and mode of antiviral action of polyanionic carbosilane dendrimer G2-S16. Nanomed. Nanotechnol. Boil. Med. 2015, 11, 1299–1308. [Google Scholar] [CrossRef]
- Chonco, L.; Pion, M.; Vacas, E.; Rasines, B.; Maly, M.; Serramía, M.; Fernández, L.A.L.; De La Mata, F.J.; Álvarez, S.; Gomez, R.; et al. Carbosilane dendrimer nanotechnology outlines of the broad HIV blocker profile. J. Control. Release 2012, 161, 949–958. [Google Scholar] [CrossRef]
- Torres, L.; Ortiz, T.; Tang, Q. Enhancement of Herpes Simplex Virus (HSV) Infection by Seminal Plasma and Semen Amyloids Implicates a New Target for the Prevention of HSV Infection. Viruses 2015, 7, 2057–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Merino, I.; Cuevas, N.D.L.; Jiménez, J.L.; Gallego, J.; Gómez, C.; Prieto, C.; Serramía, M.J.; Lorente, R.; Muñoz-Fernández, M. Ángeles The Spanish HIV BioBank: A model of cooperative HIV research. Retrovirology 2009, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Broncano, P.; Ceña-Diez, R.; De La Mata, F.J.; Gomez-Ramirez, R.; Resino, S.; Muñoz-Fernández, M. Ángeles Efficacy of carbosilane dendrimers with an antiretroviral combination against HIV-1 in the presence of semen-derived enhancer of viral infection. Eur. J. Pharmacol. 2017, 811, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Córdoba, E.V.; Arnaiz, E.; Relloso, M.; Sánchez-Torres, C.; García, F.; Pérez-Álvarez, L.; Gomez-Ramirez, R.; De La Mata, F.J.; Pion, M.; Muñoz-Fernández, M. Ángeles Development of sulphated and naphthylsulphonated carbosilane dendrimers as topical microbicides to prevent HIV-1 sexual transmission. AIDS 2013, 27, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Toka, F.N.; Gierynska, M.; Rouse, B.T. Codelivery of CCR7 Ligands as Molecular Adjuvants Enhances the Protective Immune Response against Herpes Simplex Virus Type 1. J. Virol. 2003, 77, 12742–12752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallichan, W.S.; Woolstencroft, R.N.; Guarasci, T.; McCluskie, M.J.; Davis, H.L.; Rosenthal, K.L. Intranasal immunization with CpG oligodeoxynucleotides as an adjuvant dramatically increases IgA and protection against herpes simplex virus-2 in the genital tract. J. Immunol. 2001, 166, 3451–3457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parr, M.B.; Kepple, L.; McDermott, M.R.; Drew, M.D.; Bozzola, J.J.; Parr, E.L. A mouse model for studies of mucosal immunity to vaginal infection by herpes simplex virus type 2. Lab. Investig. 1994, 70, 369–380. [Google Scholar]
- Knight, R.; Vrbanac, A.; Taylor, B.C.; Aksenov, A.; Callewaert, C.; Debelius, J.; González, A.; Kosciolek, T.; McCall, L.-I.; McDonald, D.; et al. Best practices for analysing microbiomes. Nat. Rev. Genet. 2018, 16, 410–422. [Google Scholar] [CrossRef] [Green Version]
- Rotmistrovsky, K.; Agarwala, R. BMTagger: Best Match Tagger for removing human reads from metagenomics datasets. Bioinformatics 2011. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Ulyantsev, V.; Kazakov, S.V.; Dubinkina, V.B.; Tyakht, A.V.; Alexeev, D.G. MetaFast: Fast reference-free graph-based comparison of shotgun metagenomic data. Bioinformatics 2016, 32, 2760–2767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnabas, R.V.; Wasserheit, J.N.; Huang, Y.; Janes, H.E.; Morrow, R.; Fuchs, J.; E Mark, K.; Casapia, M.; Mehrotra, D.V.; Buchbinder, S.P.; et al. Impact of Herpes Simplex Virus Type 2 on HIV-1 acquisition and progression in an HIV vaccine trial (the Step Study). JAIDS J. Acquir. Immune Defic. Syndr. 2011, 57, 1. [Google Scholar] [CrossRef]
- Tang, Q.; Roan, N.R.; Yamamura, Y. Seminal Plasma and Semen Amyloids Enhance Cytomegalovirus Infection in Cell Culture. J. Virol. 2013, 87, 12583–12591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roan, N.R.; Münch, J.; Arhel, N.J.; Mothes, W.; Neidleman, J.; Kobayashi, A.; Smith-McCune, K.; Kirchhoff, F.; Greene, W.C. The Cationic Properties of SEVI Underlie Its Ability To Enhance Human Immunodeficiency Virus Infection. J. Virol. 2008, 83, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Barfod, K.K.; Roggenbuck, M.; Hansen, L.H.; Schjørring, S.; Larsen, S.T.; Sørensen, S.J.; Krogfelt, K. The murine lung microbiome in relation to the intestinal and vaginal bacterial communities. BMC Microbiol. 2013, 13, 303. [Google Scholar] [CrossRef] [Green Version]
- Esber, A.; Miguel, R.D.V.; Cherpes, T.L.; Klebanoff, M.A.; Gallo, M.F.; Turner, A.N. Risk of Bacterial Vaginosis Among Women With Herpes Simplex Virus Type 2 Infection: A Systematic Review and Meta-analysis. J. Infect. Dis. 2015, 212, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef]
- Van De Wijgert, J.H.H.M. The vaginal microbiome and sexually transmitted infections are interlinked: Consequences for treatment and prevention. PLoS Med. 2017, 14, e1002478. [Google Scholar] [CrossRef] [Green Version]
- Lewis, F.M.T.; Bernstein, K.T.; Aral, S.O. Vaginal Microbiome and Its Relationship to Behavior, Sexual Health, and Sexually Transmitted Diseases. Obstet. Gynecol. 2017, 129, 643–654. [Google Scholar] [CrossRef]
- Ravel, J.; Gajer, P.; Fu, L.; Mauck, C.K.; Koenig, S.S.K.; Sakamoto, J.; Motsinger-Reif, A.A.; Doncel, G.F.; Zeichner, S.L. Twice-Daily Application of HIV Microbicides Alters the Vaginal Microbiota. mBio 2012, 3, e00370-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, S.; Liu, C.; Mitchell, C.; Fiedler, T.L.; Thomas, K.K.; Agnew, K.J.; Marrazzo, J.; Fredricks, D.N. Temporal Variability of Human Vaginal Bacteria and Relationship with Bacterial Vaginosis. PLoS ONE 2010, 5, e10197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koiter, T.R.; Hazenberg, M.P.; Van Der Schoot, P. Regulation of the bacterial microflora of the vagina in cyclic female rats. J. Exp. Zool. 1977, 202, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Swartz, J.D.; Lachman, M.; Westveer, K.; O’Neill, T.; Geary, T.; Kott, R.W.; Berardinelli, J.G.; Hatfield, P.G.; Thomson, J.; Roberts, A.; et al. Characterization of the Vaginal Microbiota of Ewes and Cows Reveals a Unique Microbiota with Low Levels of Lactobacilli and Near-Neutral pH. Front. Veter. Sci. 2014, 1, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, E.A.; Beasley, D.; Dunn, R.R.; Archie, E.A. Lactobacilli Dominance and Vaginal pH: Why Is the Human Vaginal Microbiome Unique? Front. Microbiol. 2016, 7, 4. [Google Scholar] [CrossRef]
- Neuendorf, E.; Gajer, P.; Bowlin, A.K.; Marques, P.X.; Ma, B.; Yang, H.; Fu, L.; Humphrys, M.S.; Forney, L.J.; Myers, G.S.; et al. Chlamydia caviae infection alters abundance but not composition of the guinea pig vaginal microbiota. Pathog. Dis. 2015, 73. [Google Scholar] [CrossRef]
- Schwebke, J.R. Role of vaginal flora as a barrier to HIV acquisition. Curr. Infect. Dis. Rep. 2001, 3, 152–155. [Google Scholar] [CrossRef]
- Ledru, S.; Méd, N.; Ledru, E.; Bazie, A.; Chiron, J. HIV-1 infection associated with abnormal vaginal flora morphology and bacterial vaginosis. Lancet 1997, 350, 1251–1252. [Google Scholar] [CrossRef]
- Chen, S.; Wang, Y.; Chen, F.; Yang, H.; Gan, M.; Zheng, S.J. A Highly Pathogenic Strain of Staphylococcus sciuri Caused Fatal Exudative Epidermitis in Piglets. PLoS ONE 2007, 2, e147. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.D.; Pradhan, A.K.; Green, S.J.; Naqib, A.; Odoyo-June, E.; Gaydos, C.A.; Barry, S.; Landay, A.; Bailey, R.C. Microbial Diversity of Genital Ulcers of HSV-2 Seropositive Women. Sci. Rep. 2017, 7, 15475. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerrero-Beltrán, C.; Garcia-Heredia, I.; Ceña-Diez, R.; Rodriguez-Izquierdo, I.; Serramía, M.J.; Martinez-Hernandez, F.; Lluesma-Gomez, M.; Martinez-Garcia, M.; Muñoz-Fernández, M.Á. Cationic Dendrimer G2-S16 Inhibits Herpes Simplex Type 2 Infection and Protects Mice Vaginal Microbiome. Pharmaceutics 2020, 12, 515. https://doi.org/10.3390/pharmaceutics12060515
Guerrero-Beltrán C, Garcia-Heredia I, Ceña-Diez R, Rodriguez-Izquierdo I, Serramía MJ, Martinez-Hernandez F, Lluesma-Gomez M, Martinez-Garcia M, Muñoz-Fernández MÁ. Cationic Dendrimer G2-S16 Inhibits Herpes Simplex Type 2 Infection and Protects Mice Vaginal Microbiome. Pharmaceutics. 2020; 12(6):515. https://doi.org/10.3390/pharmaceutics12060515
Chicago/Turabian StyleGuerrero-Beltrán, Carlos, Inmaculada Garcia-Heredia, Rafael Ceña-Diez, Ignacio Rodriguez-Izquierdo, María Jesús Serramía, Francisco Martinez-Hernandez, Mónica Lluesma-Gomez, Manuel Martinez-Garcia, and María Ángeles Muñoz-Fernández. 2020. "Cationic Dendrimer G2-S16 Inhibits Herpes Simplex Type 2 Infection and Protects Mice Vaginal Microbiome" Pharmaceutics 12, no. 6: 515. https://doi.org/10.3390/pharmaceutics12060515