Assessment of the Chemical Diversity and Potential Toxicity of Benthic Cyanobacterial Blooms in the Lagoon of Moorea Island (French Polynesia)


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Species Identification
2.2. Metabolites Fingerprinting
2.3. Evaluation of Cytotoxic Activity Using the Neuroblastoma Cell-Based Assay
2.4. Evaluation of Toxicity Using Artemia salina Lethality Bioassay
3. Results and Discussion
3.1. Identification of Benthic Cyanobacterial Blooms
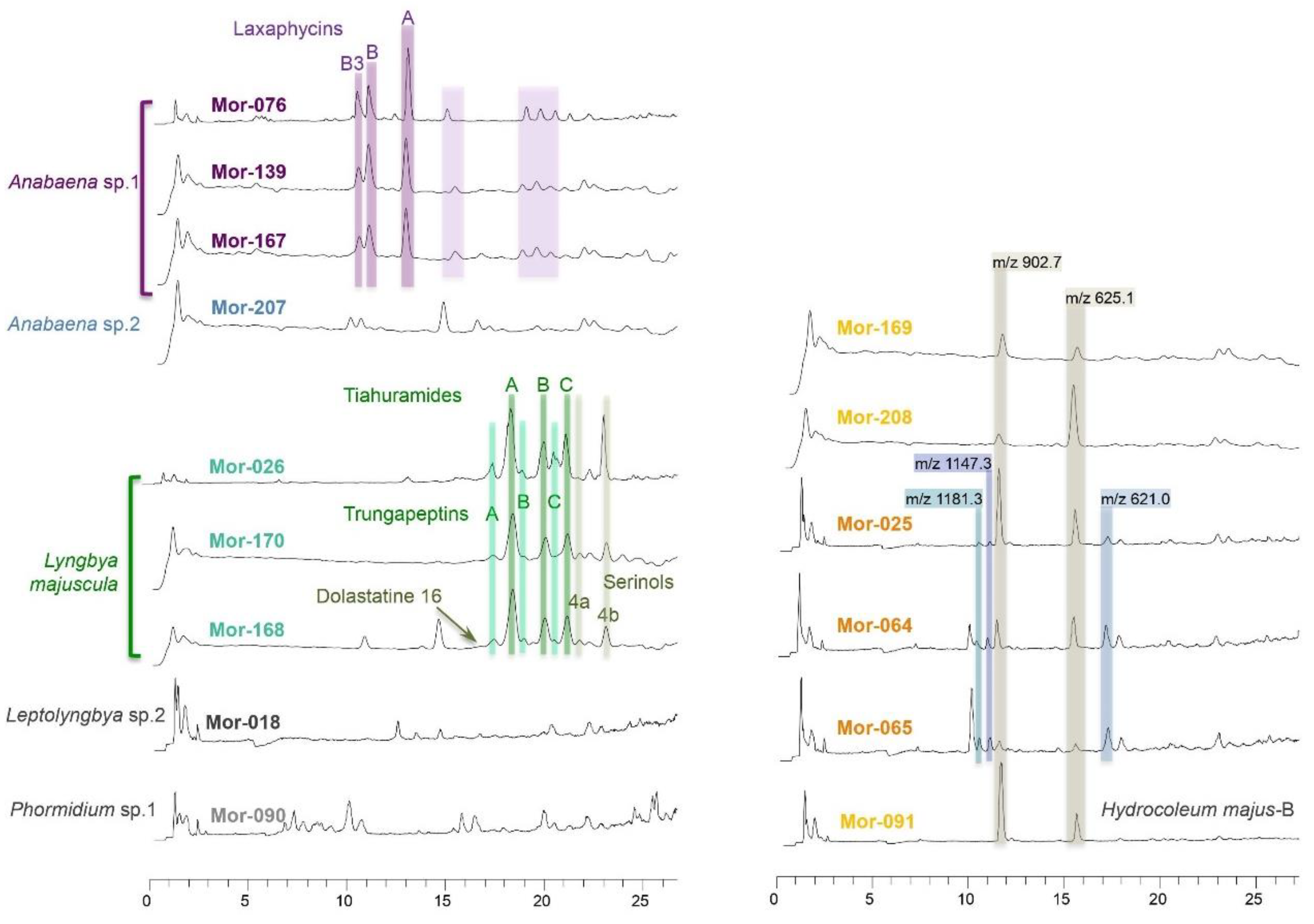
3.2. Chemical Profiles of Benthic Cyanobacterial Blooms
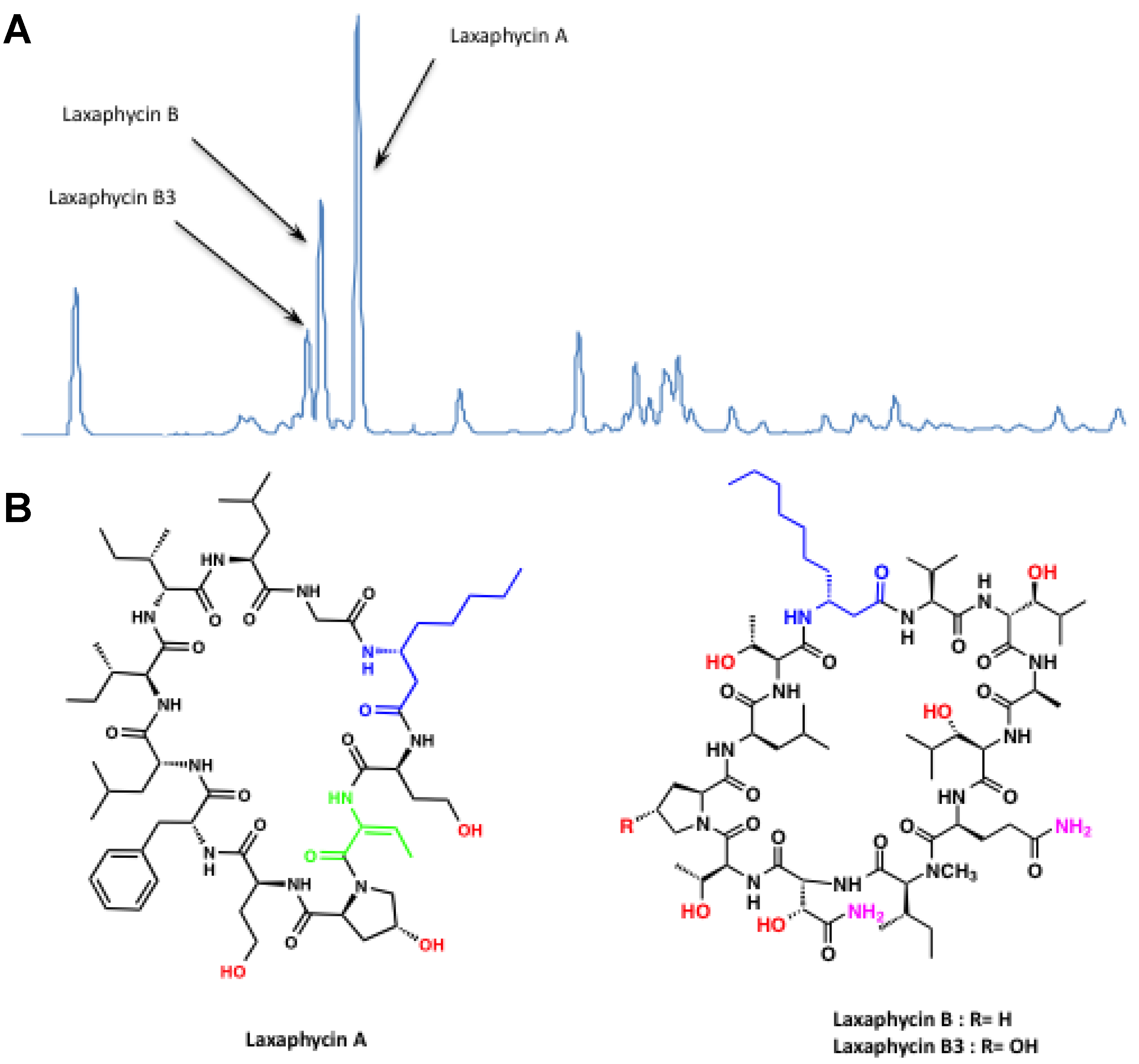
3.2.1. Anabaena sp.1 Blooms
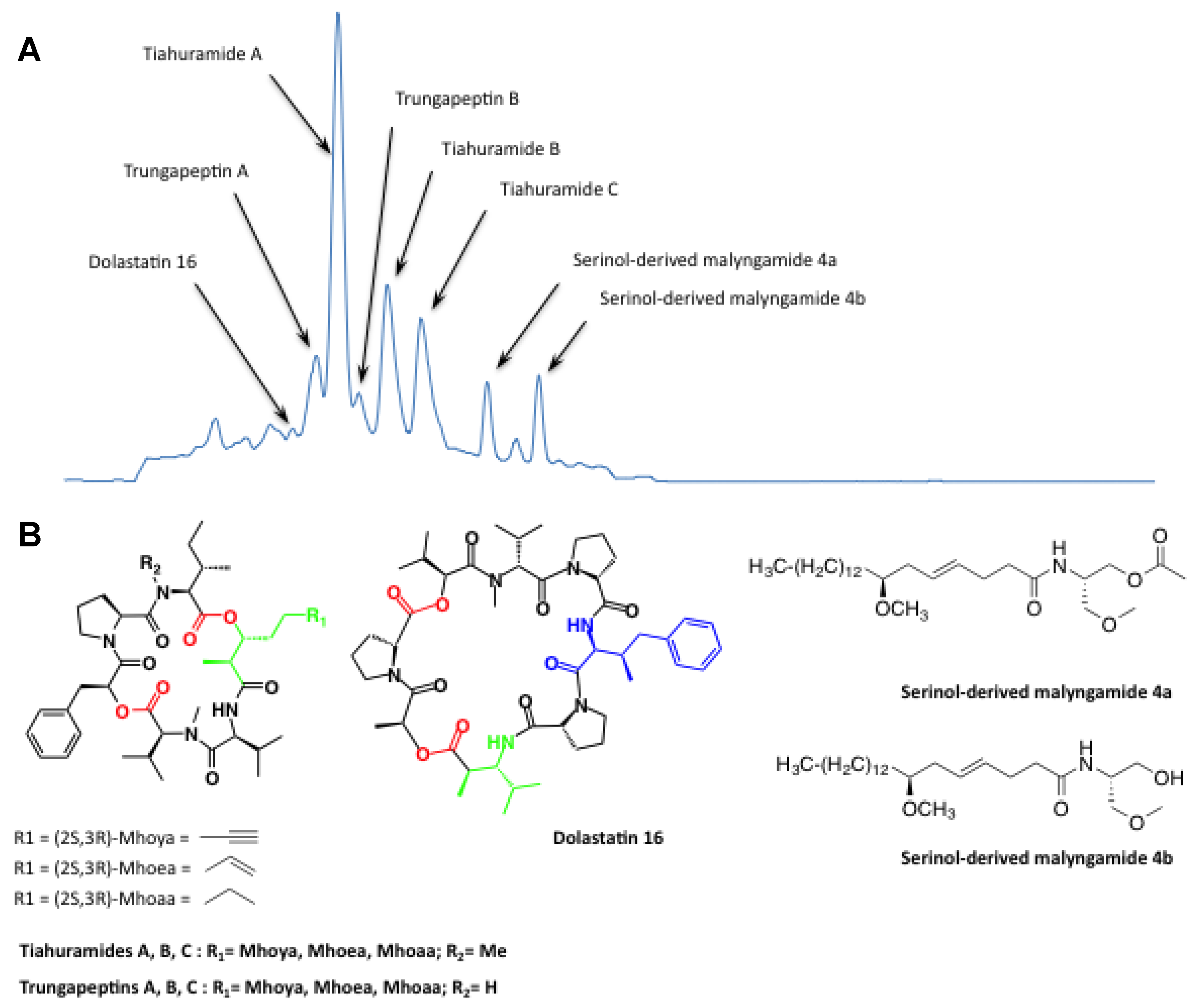
3.2.2. Lyngbya Majuscula Blooms
3.2.3. Other Blooms
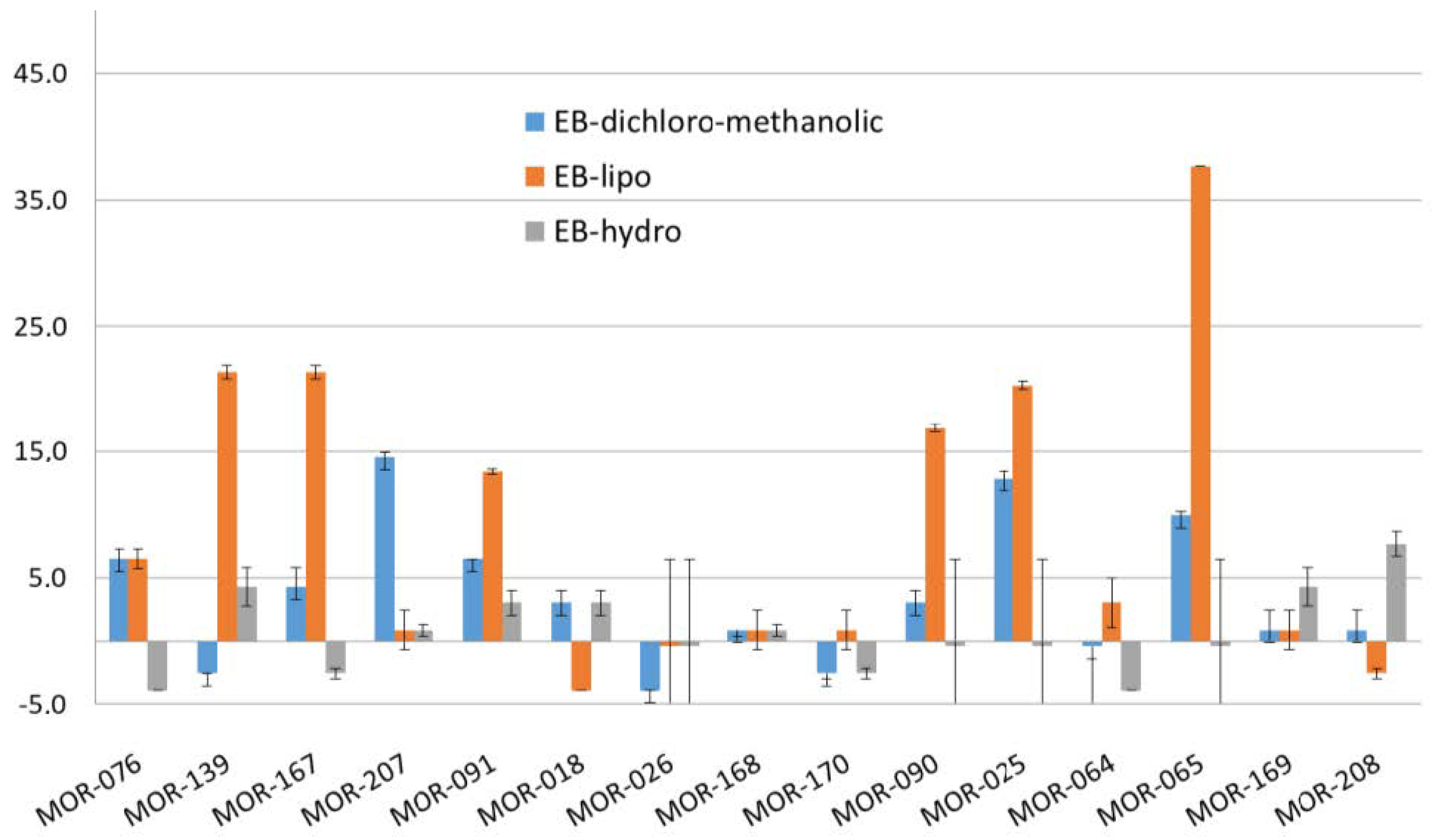
3.3. Toxicity of Benthic Cyanobacterial Blooms
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brocks, J.J.; Logan, G.A.; Buick, R.; Summons, R.E. Archean molecular fossils and the early rise of eukaryotes. Science 1999, 285, 1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, A.K.; Tiwari, D.N.; Rai, A.N. Cyanobacteria: From Basic Science to Applications; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Bernard, C.; Ballot, A.; Thomazeau, S.; Maloufi, S.; Furey, A.; Mankiewicz-Boczek, J.; Pawlik-Skowrońska, B.; Capelli, C.; Salmaso, N. Appendix 2: Cyanobacteria associated with the production of cyanotoxins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 501–525. [Google Scholar]
- Du, X.; Liu, H.; Yuan, L.; Wang, Y.; Ma, Y.; Wang, R.; Chen, X.; Losiewicz, D.M.; Guo, H.; Zhang, H. The diversity of cyanobacterial toxins on structural characterization, distribution and identification: A systematic review. Toxins 2019, 11, 530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burja, A.M.; Banaigs, B.; Abou-Mansour, E.; Grant Burgess, J.; Wright, P.C. Marine cyanobacteria - a prolific source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar] [CrossRef]
- Tan, L.T. Bioactive natural products from marine cyanobacteria for drug discovery. Phytochemistry 2007, 68, 954–979. [Google Scholar] [CrossRef] [PubMed]
- Salvador-Reyes, L.A.; Luesch, H. Biological targets and mechanisms of action of natural products from marine cyanobacteria. Nat. Prod. Rep. 2015, 32, 478–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, Y.; Zhang, J.; He, S.; Yan, X. New peptides isolated from marine cyanobacteria, an overview over the past decade. Mar. Drugs 2017, 15, 132. [Google Scholar] [CrossRef] [Green Version]
- Roué, M.; Gugger, M.; Golubic, S.; Amzil, Z.; Aráoz, R.; Turquet, J.; Chinain, M.; Laurent, D. Marine cyanotoxins potentially harmful to human health. In Outstanding Marine Molecules; La Barre, S., Kornprobst, J.-M., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2014; pp. 3–22. [Google Scholar]
- Osborne, N.J.T.; Webb, P.M.; Shaw, G.R. The toxins of Lyngbya majuscula and their human and ecological health effects. Environ. Int. 2001, 27, 381–392. [Google Scholar] [CrossRef]
- Osborne, N.; Seawright, A.; Shaw, G. Dermal toxicology of Lyngbya majuscula from Moreton Bay, Queensland, Australia. Harmful Algae 2008, 7, 584–589. [Google Scholar] [CrossRef]
- Taylor, M.S.; Stahl-Timmins, W.; Redshaw, C.H.; Osborne, N.J. Toxic alkaloids in Lyngbya majuscula and related tropical marine cyanobacteria. Harmful Algae 2014, 31, 1–8. [Google Scholar] [CrossRef]
- Laurent, D.; Kerbrat, A.-S.; Darius, H.T.; Girard, E.; Golubic, S.; Benoit, E.; Sauviat, M.-P.; Chinain, M.; Molgo, J.; Pauillac, S. Are cyanobacteria involved in Ciguatera Fish Poisoning-like outbreaks in New Caledonia? Harmful Algae 2008, 7, 827–838. [Google Scholar] [CrossRef]
- Golubic, S.; Abed, R.M.M.; Palińska, K.; Pauillac, S.; Chinain, M.; Laurent, D. Marine toxic cyanobacteria: Diversity, environmental responses and hazards. Toxicon 2010, 56, 836–841. [Google Scholar] [CrossRef] [PubMed]
- Méjean, A.; Peyraud-Thomas, C.; Kerbrat, A.S.; Golubic, S.; Pauillac, S.; Chinain, M.; Laurent, D. First identification of the neurotoxin homoanatoxin-a from mats of Hydrocoleum lyngbyaceum (marine cyanobacterium) possibly linked to giant clam poisoning in New Caledonia. Toxicon 2010, 56, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Laurent, D.; Kerbrat, A.S.; Darius, H.T.; Rossi, F.; Yeeting, B.; Haddad, M.; Golubic, S.; Pauillac, S.; Chinain, M. Ciguatera Shellfish Poisoning (CSP): A new ecotoxicological phenomenon from cyanobacteria to humans via giant clams. In Food Chains: New Research; Jensen, M.A., Muller, D.W., Eds.; Nova Science Publishers: Hauppauge, NY, USA, 2012; pp. 1–44. [Google Scholar]
- Kuffner, I.B.; Walters, L.J.; Becerro, M.A.; Paul, V.J.; Ritson-Williams, R.l.; Beach, K.S. Inhibition of coral recruitment by macroalgae and cyanobacteria. Mar. Ecol. Prog. Ser. 2006, 323, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Kramarsky-Winter, E.; Arotsker, L.; Rasoulouniriana, D.; Siboni, N.; Loya, Y.; Kushmaro, A. The possible role of cyanobacterial filaments in coral black band disease pathology. Microb. Ecol. 2014, 67, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Morrow, K.M.; Ritson-Williams, R.; Ross, C.; Liles, M.R.; Paul, V.J. Macroalgal extracts induce bacterial assemblage shifts and sublethal tissue stress in Caribbean corals. PLoS ONE 2012, 7, e44859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef]
- Zubia, M.; Vieira, C.; Palinska, K.; Roué, M.; Gaertner, J.-C.; Zloch, I.; Grellier, M.; Golubic, S. Benthic cyanobacteria on coral reefs of Moorea Island (French Polynesia): Diversity response to habitat quality. Hydrobiologia 2019, 843, 61–78. [Google Scholar] [CrossRef]
- Bonnard, I.; Rolland, M.; Francisco, C.; Banaigs, B. Total structure and biological properties of laxaphycins A and B, cyclic lipopeptides from the marine cyanobacterium Lyngbya majuscula. Lett. Pept. Sci. 1997, 4, 289–292. [Google Scholar] [CrossRef]
- Bonnard, I.; Rolland, M.; Salmon, J.-M.; Debiton, E.; Barthomeuf, C.; Banaigs, B. Total structure and inhibition of tumor cell proliferation of laxaphycins. J. Med. Chem. 2007, 50, 1266–1279. [Google Scholar] [CrossRef]
- Bornancin, L.; Boyaud, F.; Mahiout, Z.; Bonnard, I.; Mills, S.C.; Banaigs, B.; Inguimbert, N. Isolation and synthesis of laxaphycin B-type peptides: A case study and clues to their biosynthesis. Mar. Drugs 2015, 13, 7285–7300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levert, A.; Alvariño, R.; Bornancin, L.; Abou Mansour, E.; Burja, A.M.; Genevière, A.-M.; Bonnard, I.; Alonso, E.; Botana, L.; Banaigs, B. Structures and activities of tiahuramides A–C, cyclic depsipeptides from a Tahitian collection of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2018, 81, 1301–1310. [Google Scholar] [CrossRef] [PubMed]
- Bornancin, L.; Alonso, E.; Alvariño, R.; Inguimbert, N.; Bonnard, I.; Botana, L.M.; Banaigs, B. Structure and biological evaluation of new cyclic and acyclic laxaphycin-A type peptides. Bioorg. Med. Chem. 2019, 27, 1966–1980. [Google Scholar] [CrossRef] [PubMed]
- Villeneuve, A.; Laurent, D.; Chinain, M.; Gugger, M.; Humbert, J.-F. Molecular characterization of the diversity and potential toxicity of cyanobacterial mats in two tropical lagoons in the South Pacific Ocean. J. Phycol. 2012, 48, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Hokama, Y.; Dickey, R.W.; Granade, H.R.; Lewis, R.; Yasumoto, T.; Wekell, M.M. Detection of sodium channel toxins: Directed cytotoxicity assays of purified ciguatoxins, brevetoxins, saxitoxins, and seafood extracts. J. AOAC Int. 1995, 78, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Chinain, M.; Gatti, C.M.; Roué, M.; Darius, H.T. Ciguatera poisoning in French Polynesia: Insights into the novel trends of an ancient disease. New Microbes New Infect. 2019, 31, 100565. [Google Scholar] [CrossRef]
- Skinner, M.P.; Brewer, T.D.; Johnstone, R.; Fleming, L.E.; Lewis, R.J. Ciguatera fish poisoning in the Pacific Islands (1998 to 2008). PLoS Negl. Trop. Dis. 2011, 5, e1416. [Google Scholar] [CrossRef]
- Chinain, M.; Darius, H.T.; Gatti, C.M.; Roué, M. Biotoxines marines et sécurité sanitaire des ressources lagonaires en Polynésie française. In La Sécurité Alimentaire (Les Cahiers de Droit de la Santé Juridiques, Historiques et Prospectifs n°27); Mu Si Yan, L., Qi, J., De Raulin, A., Eds.; LEH Editions: Bordeaux, France, 2018; pp. 119–143. [Google Scholar]
- Nogle, L.M.; Gerwick, W.H. Isolation of four new cyclic depsipeptides, antanapeptins A−D, and dolastatin 16 from a Madagascan collection of Lyngbya majuscula. J. Nat. Prod. 2002, 65, 21–24. [Google Scholar] [CrossRef]
- Pettit, G.R.; Xu, J.-P.; Hogan, F.; Williams, M.D.; Doubek, D.L.; Schmidt, J.M.; Cerny, R.L.; Boyd, M.R. Isolation and structure of the human cancer cell growth inhibitory cyclodepsipeptide dolastatin 16. J. Nat. Prod. 1997, 60, 752–754. [Google Scholar] [CrossRef]
- Wan, F.; Erickson, K.L. Serinol-derived malyngamides from an Australian cyanobacterium. J. Nat. Prod. 1999, 62, 1696–1699. [Google Scholar] [CrossRef]
- Bunyajetpong, S.; Yoshida, W.Y.; Sitachitta, N.; Kaya, K. Trungapeptins A−C, cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2006, 69, 1539–1542. [Google Scholar] [CrossRef] [PubMed]
- Viallon, J.; Chinain, M.; Darius, T.H. Revisiting the neuroblastoma cell-based assay (CBA-N2a) for the improved detection of marine toxins active on voltage gated sodium channels (VGSCs). Toxins 2020, 12, 281. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.N.; Ferrigni, N.R.; Putnam, J.E.; Jacobsen, L.B.; Nichols, D.E.; McLaughlin, J.L. Brine shrimp: A convenient general bioassay for active plant constituents. Planta Med. 1982, 45, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Rajabi, S.; Ramazani, A.; Hamidi, M.; Naji, T. Artemia salina as a model organism in toxicity assessment of nanoparticles. DARU J. Pharm. Sci. 2015, 23, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lernout, T.; Thiria, J.; Maltaverne, E.; Salim, M.; Turquet, J.; Lajoinie, G.; Solet, J.L.; Filleul, L. Alerte aux cyanobactéries sur la plage de N’Gouja, Mayotte, avril 2010. Bulletin de Veille Sanitaire de la Cellule de l’Institut de Veille Sanitaire en Régoin Océan Indien 2012, 9, 12–14. [Google Scholar]
- Charpy, L.; Casareto, B.E.; Langlade, M.J.; Suzuki, Y. Cyanobacteria in coral reef ecosystems: A review. J. Mar. Biol. 2012, 2012, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Frankmölle, W.P.; Knübel, G.; Moore, R.E.; Patterson, G.M. Antifungal cyclic peptides from the terrestrial blue-green alga Anabaena laxa. II. Structures of laxaphycins A, B, D and E. J. Antibiot. 1992, 45, 1458–1466. [Google Scholar] [CrossRef] [Green Version]
- Gerwick, W.H.; Jiang, Z.D.; Agarwal, S.K.; Farmer, B.T. Total structure of hormothamnin A, a toxic cyclic undecapeptide from the tropical marine cyanobacterium Hormothamnion enteromorphoides. Tetrahedron 1992, 48, 2313–2324. [Google Scholar] [CrossRef]
- MacMillan, J.B.; Ernst-Russell, M.A.; de Ropp, J.S.; Molinski, T.F. Lobocyclamides A−C, lipopeptides from a cryptic cyanobacterial mat containing Lyngbya confervoides. J. Org. Chem. 2002, 67, 8210–8215. [Google Scholar] [CrossRef]
- Luo, S.; Krunic, A.; Kang, H.-S.; Chen, W.-L.; Woodard, J.L.; Fuchs, J.R.; Swanson, S.M.; Orjala, J. Trichormamides A and B with antiproliferative activity from the cultured freshwater cyanobacterium Trichormus sp. UIC 10339. J. Nat. Prod. 2014, 77, 1871–1880. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Kang, H.-S.; Krunic, A.; Chen, W.-L.; Yang, J.; Woodard, J.L.; Fuchs, J.R.; Hyun Cho, S.; Franzblau, S.G.; Swanson, S.M.; et al. Trichormamides C and D, antiproliferative cyclic lipopeptides from the cultured freshwater cyanobacterium cf. Oscillatoria sp. UIC 10045. Bioorg. Med. Chem. 2015, 23, 3153–3162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maru, N.; Ohno, O.; Uemura, D. Lyngbyacyclamides A and B, novel cytotoxic peptides from marine cyanobacteria Lyngbya sp. Tetrahedron Lett. 2010, 51, 6384–6387. [Google Scholar] [CrossRef]
- Zhaxybayeva, O.; Gogarten, J.P.; Charlebois, R.L.; Doolittle, W.F.; Papke, R.T. Phylogenetic analyses of cyanobacterial genomes: Quantification of horizontal gene transfer events. Genome Res. 2006, 16, 1099–1108. [Google Scholar] [CrossRef] [Green Version]
- Sitachitta, N.; Williamson, R.T.; Gerwick, W.H. Yanucamides A and B, two new depsipeptides from an assemblage of the marine cyanobacteria Lyngbya majuscula and Schizothrix species. J. Nat. Prod. 2000, 63, 197–200. [Google Scholar] [CrossRef]
- Luesch, H.; Pangilinan, R.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Pitipeptolides A and B, new cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2001, 64, 304–307. [Google Scholar] [CrossRef]
- Montaser, R.; Paul, V.J.; Luesch, H. Pitipeptolides C–F, antimycobacterial cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula from Guam. Phytochemistry 2011, 72, 2068–2074. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Lee, P.P.F.; Tan, L.T. Hantupeptin A, a cytotoxic cyclic depsipeptide from a Singapore collection of Lyngbya majuscula. J. Nat. Prod. 2009, 72, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Lee, P.P.F.; Tan, L.T. Hantupeptins B and C, cytotoxic cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. Phytochemistry 2010, 71, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Tan, L.T. Lagunamides A and B: Cytotoxic and antimalarial cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010, 73, 1810–1814. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Chan, K.P.; Chen, D.Y.-K.; Tan, L.T. Lagunamide C, a cytotoxic cyclodepsipeptide from the marine cyanobacterium Lyngbya majuscula. Phytochemistry 2011, 72, 2369–2375. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, P.D.; Byrum, T.; Liu, W.-T.; Dorrestein, P.C.; Gerwick, W.H. Viequeamide A, a cytotoxic member of the kulolide superfamily of cyclic depsipeptides from a marine button cyanobacterium. J. Nat. Prod. 2012, 75, 1560–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakao, Y.; Yoshida, W.Y.; Szabo, C.M.; Baker, B.J.; Scheuer, P.J. More peptides and other diverse constituents of the marine mollusk Philinopsis speciosa. J. Org. Chem. 1998, 63, 3272–3280. [Google Scholar] [CrossRef]
- Montaser, R.; Abboud, K.A.; Paul, V.J.; Luesch, H. Pitiprolamide, a proline-rich dolastatin 16 analogue from the marine cyanobacterium Lyngbya majuscula from Guam. J. Nat. Prod. 2011, 74, 109–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engene, N.; Gunasekera, S.P.; Gerwick, W.H.; Paul, V.J. Phylogenetic inferences reveal a large extent of novel biodiversity in chemically rich tropical marine cyanobacteria. Appl. Environ. Microb. 2013, 79, 1882–1888. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.C.; Monroe, E.A.; Podell, S.; Hess, W.R.; Klages, S.; Esquenazi, E.; Niessen, S.; Hoover, H.; Rothmann, M.; Lasken, R.S.; et al. Genomic insights into the physiology and ecology of the marine filamentous cyanobacterium Lyngbya majuscula. Proc. Natl. Acad. Sci. USA 2011, 108, 8815–8820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engene, N.; Choi, H.; Esquenazi, E.; Rottacker, E.C.; Ellisman, M.H.; Dorrestein, P.C.; Gerwick, W.H. Underestimated biodiversity as a major explanation for the perceived rich secondary metabolite capacity of the cyanobacterial genus Lyngbya. Environ. Microbiol. 2011, 13, 1601–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engene, N.; Rottacker, E.C.; Kaštovský, J.; Byrum, T.; Choi, H.; Ellisman, M.H.; Komárek, J.; Gerwick, W.H. Moorea producens gen. nov., sp. nov. and Moorea bouillonii comb. nov., tropical marine cyanobacteria rich in bioactive secondary metabolites. Int. J. Syst. Evol. Microbiol. 2012, 62, 1171–1178. [Google Scholar] [CrossRef]
- MarinLit. Available online: http://pubs.rsc.org/marinlit/ (accessed on 25 August 2019).
- Clark, B.R.; Engene, N.; Teasdale, M.E.; Rowley, D.C.; Matainaho, T.; Valeriote, F.A.; Gerwick, W.H. Natural products chemistry and taxonomy of the marine cyanobacterium Blennothrix cantharidosmum. J. Nat. Prod. 2008, 71, 1530–1537. [Google Scholar] [CrossRef] [Green Version]
- Gbankoto, A.; Vigo, J.; Dramane, K.; Banaigs, B.; Aina, E.; Salmon, J.-M. Cytotoxic effect of laxaphycins A and B on human lymphoblastic cells (CCRF-CEM) using digitised videomicrofluorometry. In Vivo 2005, 19, 577–582. [Google Scholar]
- Yasumoto, T. Fish poisoning due to toxins of microalgal origins in the Pacific. Toxicon 1998, 36, 1515–1518. [Google Scholar] [CrossRef]
- Ito, E.; Satake, M.; Yasumoto, T. Pathological effects of lyngbyatoxin A upon mice. Toxicon 2002, 40, 551–556. [Google Scholar] [CrossRef]
- Schwarz, D.; Orf, I.; Kopka, J.; Hagemann, M. Recent applications of metabolomics toward cyanobacteria. Metabolites 2013, 3, 72–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Réveillon, D.; Tunin-Ley, A.; Grondin, I.; Othmani, A.; Zubia, M.; Bunet, R.; Turquet, J.; Culioli, G.; Briand, J.-F. Exploring the chemodiversity of tropical microalgae for the discovery of natural antifouling compounds. J. Appl. Phycol. 2019, 31, 319–333. [Google Scholar] [CrossRef] [Green Version]
- Aysun, A.; Yağmur, K.; Yusuf, B. Cell proliferation and cytotoxicity assays. Curr. Pharm. Biotechnol. 2016, 17, 1213–1221. [Google Scholar] [CrossRef]






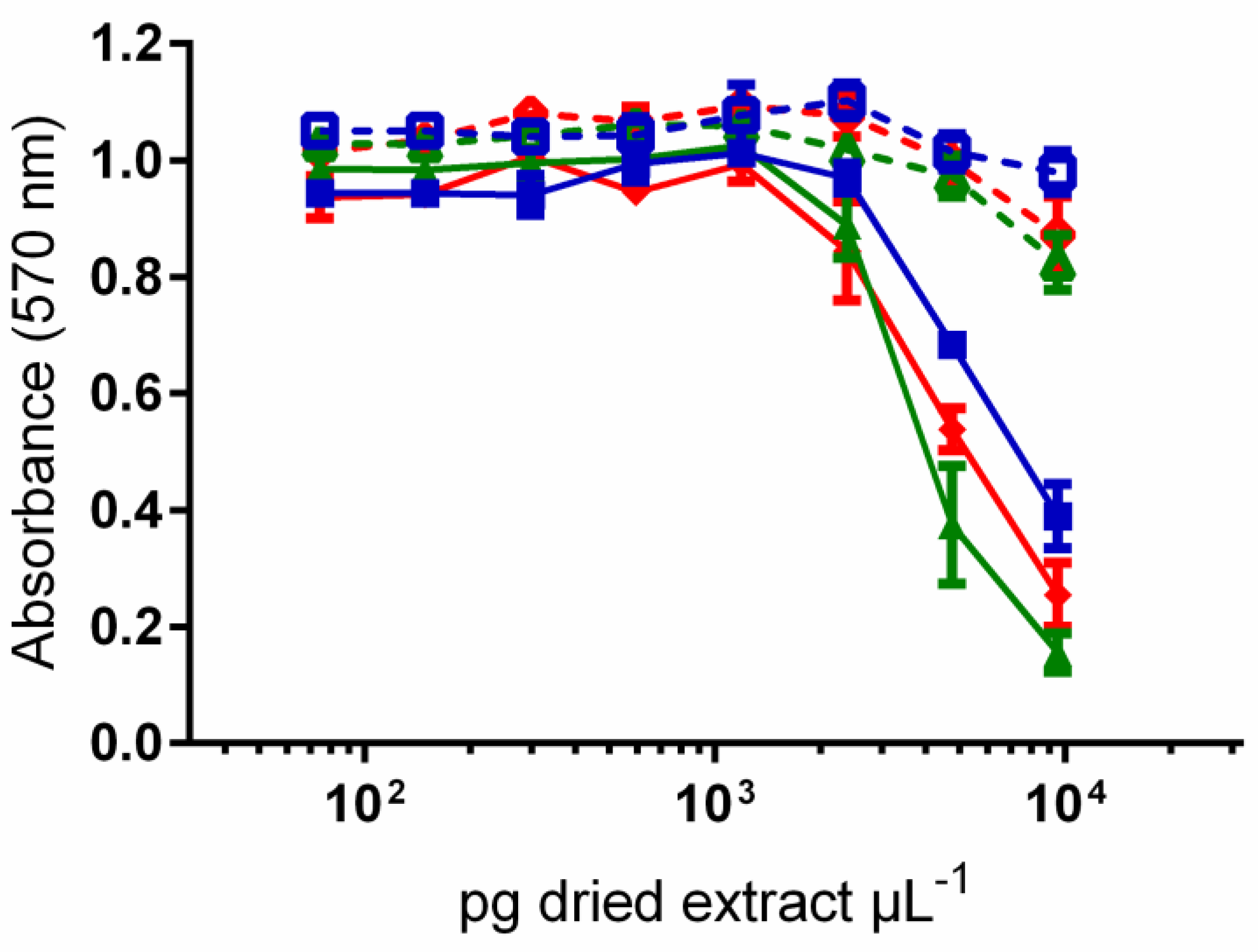
 /
/  ), MOR-139 (
), MOR-139 (  /
/  ) and MOR-167 (
) and MOR-167 (  /
/  ) collected in the lagoon of Moorea Island. Data represent the means ± SE of values obtained from three independent neuroblastoma cell-based assays (CBA-N2a), each point run in triplicate.
/ ), MOR-139 ( / ) and MOR-167 ( / ) collected in the lagoon of Moorea Island. Data represent the means ± SE of values obtained from three independent neuroblastoma cell-based assays (CBA-N2a), each point run in triplicate.
) collected in the lagoon of Moorea Island. Data represent the means ± SE of values obtained from three independent neuroblastoma cell-based assays (CBA-N2a), each point run in triplicate.
/ ), MOR-139 ( / ) and MOR-167 ( / ) collected in the lagoon of Moorea Island. Data represent the means ± SE of values obtained from three independent neuroblastoma cell-based assays (CBA-N2a), each point run in triplicate.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
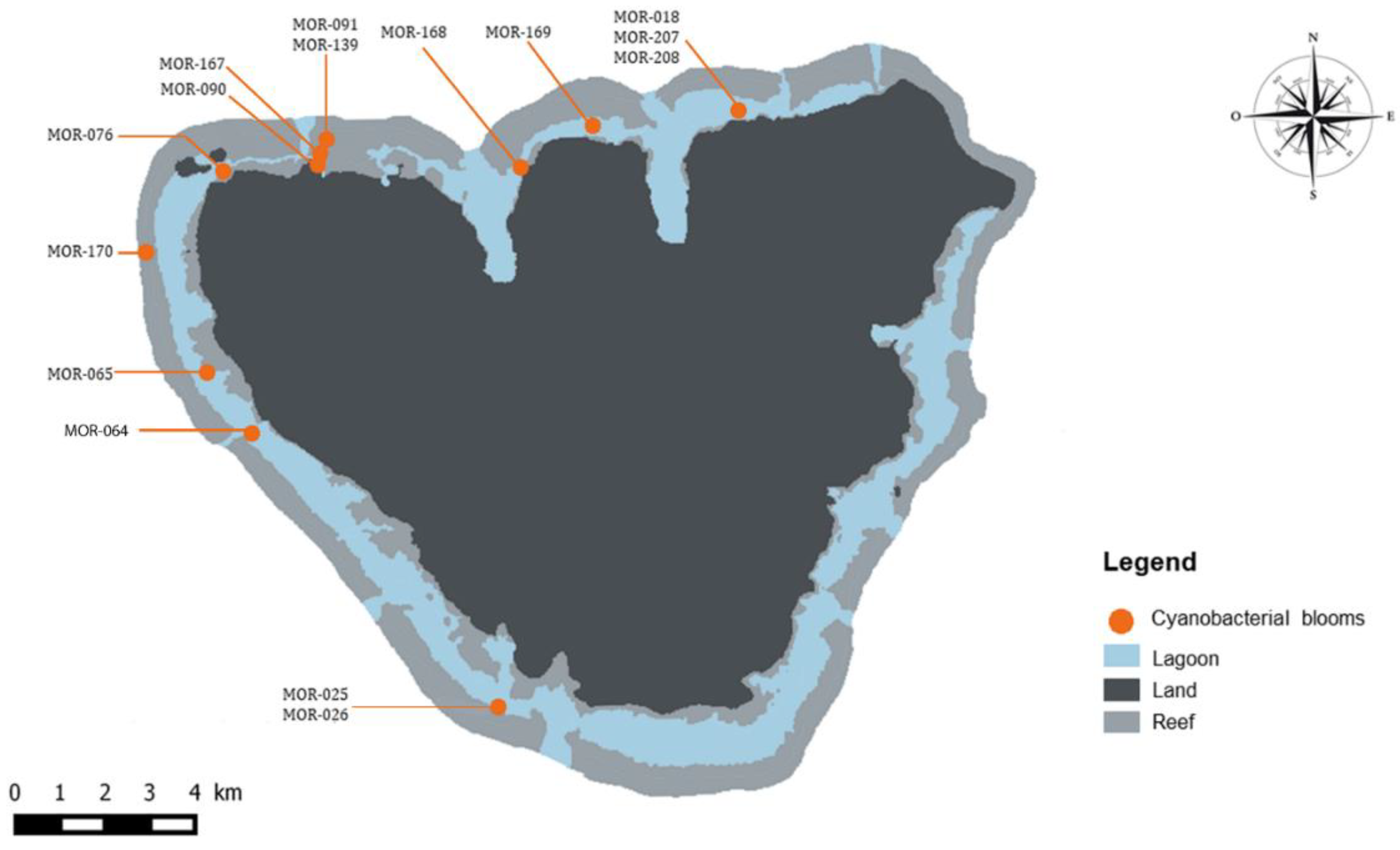
| Species Dominance | N° Voucher | Latitude S | Longitude W |
|---|---|---|---|
| MOR-076 | 17°29’26.60’’ | 149°54’40.50’’ | |
| Anabaena sp.1 | MOR-139 | 17°29’13.20’’ | 149°53’24.20’’ |
| MOR-167 | 17°29’19.46’’ | 149°53’28.32’’ | |
| Anabaena sp.2 | MOR-207 | 17°28’47.63’’ | 149’48’27.78’’ |
| Hydrocoleum majus-B | MOR-091 | 17°29’13.20’’ | 149°53’24.20’’ |
| Leptolyngbya sp.2 | MOR-018 | 17°28’47.63’’ | 149’48’27.78’’ |
| MOR-026 | 17°35’30.50’’ | 149°51’18.00’’ | |
| Lyngbya majuscula | MOR-168 | 17°29’21.91’’ | 149°51’05.26’’ |
| MOR-170 | 17°28’58.87’’ | 149°50’06.47’’ | |
| Phormidium sp.1 | MOR-090 | 17°29’23.00’’ | 149°53’36.40’’ |
| MOR-025 | 17°35’30.50’’ | 149°51’18.00’’ | |
| MOR-064 | 17°32’24.24’’ | 149°54’16.05’’ | |
| Plurispecific mats 1 | MOR-065 | 17°31’43.11’’ | 149°54’48.52’’ |
| MOR-169 | 17°28’58.91’’ | 149°50’10.32’’ | |
| MOR-208 | 17°28’47.63’’ | 149’48’27.78’’ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonnard, I.; Bornancin, L.; Dalle, K.; Chinain, M.; Zubia, M.; Banaigs, B.; Roué, M. Assessment of the Chemical Diversity and Potential Toxicity of Benthic Cyanobacterial Blooms in the Lagoon of Moorea Island (French Polynesia). J. Mar. Sci. Eng. 2020, 8, 406. https://doi.org/10.3390/jmse8060406
Bonnard I, Bornancin L, Dalle K, Chinain M, Zubia M, Banaigs B, Roué M. Assessment of the Chemical Diversity and Potential Toxicity of Benthic Cyanobacterial Blooms in the Lagoon of Moorea Island (French Polynesia). Journal of Marine Science and Engineering. 2020; 8(6):406. https://doi.org/10.3390/jmse8060406
Chicago/Turabian StyleBonnard, Isabelle, Louis Bornancin, Klervi Dalle, Mireille Chinain, Mayalen Zubia, Bernard Banaigs, and Mélanie Roué. 2020. "Assessment of the Chemical Diversity and Potential Toxicity of Benthic Cyanobacterial Blooms in the Lagoon of Moorea Island (French Polynesia)" Journal of Marine Science and Engineering 8, no. 6: 406. https://doi.org/10.3390/jmse8060406