Thymus zygis subsp. zygis an Endemic Portuguese Plant: Phytochemical Profiling, Antioxidant, Anti-Proliferative and Anti-Inflammatory Activities

 , , , , and
, , , , and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Standards and Reagents
2.2. Plant Material
2.3. Preparation of Extracts
2.4. Total Phenolic Compound Content
2.5. Total Flavonoid Content
2.6. Total Ortho-Diphenol Content
2.7. In Vitro Antioxidant Activity Assessment
2.7.1. ABTS•+ Scavenging Assay
2.7.2. Hydroxyl Radicals Scavenging Assay
2.7.3. Nitric Oxide Radical Scavenging Assay
2.8. Determination of the Phenolic Profile by High Performance Liquid Chromatography with Diode Array Detector and High Performance Liquid Chromatography with Electrospray Ionization and Tandem Mass Spectrometry Detection
2.9. Quantification of Oleanolic Acid (OA) and Ursolic Acid UA) in Hydroethanolic Extracts
2.10. In Vitro Cell-Based Assays
2.10.1. Cell Maintenance and Handling
2.10.2. Cell Viability/Cytotoxicity or Anti-Proliferative Activity Assay
2.10.3. Anti-Inflammatory Activity
2.11. Data and Statistical Analysis
3. Results and Discussion
3.1. Extract Yield and Chemical Composition of T. zygis Extracts
3.2. T.zygis subps. zygis Aqueous Decoction and Hydroethanolic Extracts Phenolic Profiles
3.3. Oleanolic Acid and Ursolic Acid Contents
3.4. In Vitro Antioxidant Activity
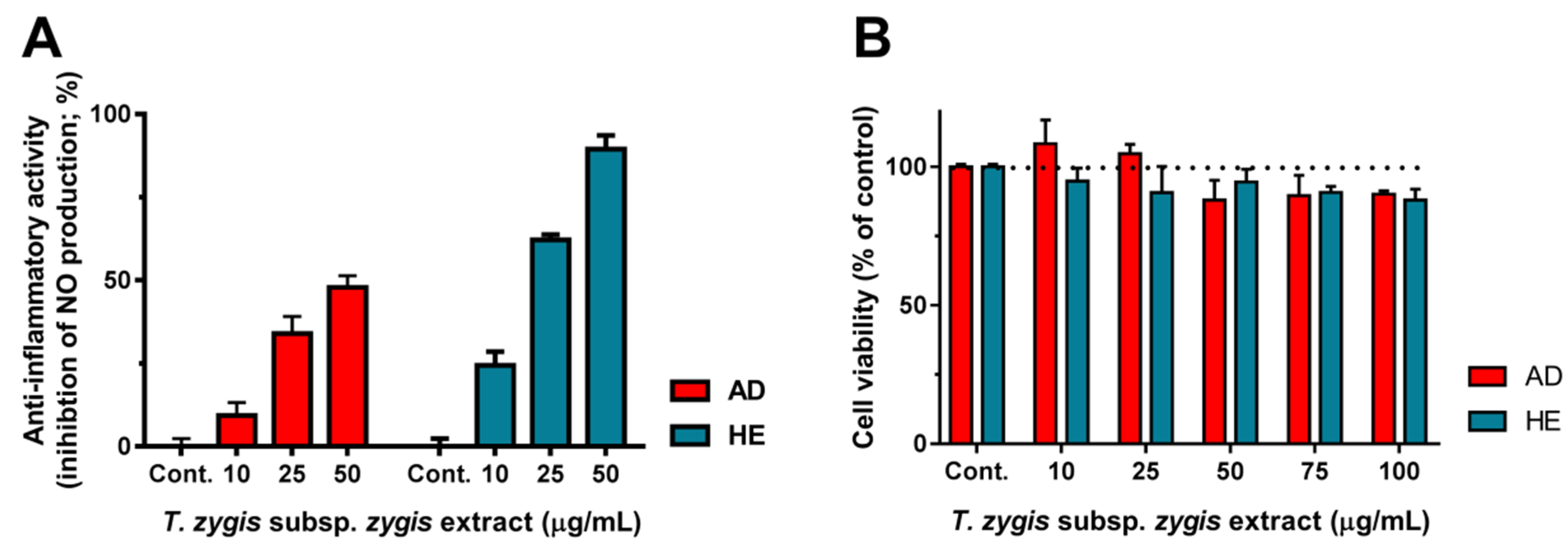
3.5. Anti-Proliferative Effect of T. zygis subsp. zygis Extracts
3.6. Anti-Inflammatory Effect of T. zygis subsp. zygis Extracts
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stahl-Biskup, E.; Sáez, F. Thyme, The genus Thymus, 1st ed.; CRC Press—Taylor and Francis: London, UK, 2003; p. 346. [Google Scholar]
- Soorni, A.; Borna, T.; Alemardan, A.; Chakrabarti, M.; Hunt, A.G.; Bombarely, A. Transcriptome Landscape Variation in the Genus Thymus. Genes 2019, 10, 620. [Google Scholar] [CrossRef] [Green Version]
- Leal, F.; Taghouti, M.; Nunes, F.M.; Silva, A.M.; Coelho, A.C.; Matos, M. Thymus Plants: A Review—Micropropagation, Molecular and Antifungal Activity. In Active Ingredients from Aromatic and Medicinal Plants. El-Shemy, H.A., Ed.; InTech: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Ghasemi Pirbalouti, A.; Emami Bistghani, Z.; Malekpoor, F. An overview on genus Thymus. J. Herb. Drugs (Int. J. Med. Herbs) 2015, 6, 93–100. [Google Scholar]
- Morales Valverde, R. Taxonomía de los géneros Thymus (excluida de la sección serpyllum) y Thymbra en la Península Ibérica; CSIC-Real Jardín Botánico (RJB): Madrid, Spain, 1986. [Google Scholar]
- Moldão-Martins, M.; Bernardo-Gil, M.G.; Beirão Da Costa, M.L.; Rouzet, M. Seasonal variation in yield and composition of Thymus zygis L. subsp. sylvestris essential oil. Flavour Fragr. J. 1999, 14, 177–182. [Google Scholar] [CrossRef]
- Dandlen, S.A.; Miguel, M.G.; Duarte, J.; Faleiro, M.L.; Sousa, M.J.; Lima, A.S.; Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G. Acetylcholinesterase Inhibition Activity of Portuguese Thymus Species Essential Oils. J. Essent. Oil Bear. Plants 2011, 14, 140–150. [Google Scholar] [CrossRef]
- Euro+Med. Thymus zygis L.—Euro+Med PlantBase—The Information Resource for Euro-Mediterranean Plant Diversity. Available online: http://ww2.bgbm.org/EuroPlusMed/PTaxonDetail.asp?NameId=92470&PTRefFk=8000000 (accessed on 20 April 2020).
- Franco, J.D.A. Nova Flora de Portugal (Continente e Açores), Volume II; Sociedade Astória Lda: Lisboa, Portugal, 1984. [Google Scholar]
- Sáez, F. Essential oil variability of Thymus zygis growing wild in southeastern spain. Phytochemistry 1995, 40, 819–825. [Google Scholar] [CrossRef]
- Rodrigues, V.; Cabral, C.; Évora, L.; Ferreira, I.; Cavaleiro, C.; Cruz, M.T.; Salgueiro, L. Chemical composition, anti-inflammatory activity and cytotoxicity of Thymus zygis L. subsp. sylvestris (Hoffmanns. & Link) Cout. essential oil and its main compounds. Arab. J. Chem. 2019, 12, 3236–3243. [Google Scholar] [CrossRef] [Green Version]
- Sotomayor, J.A.; Martínez, R.M.; García, A.J.; Jordán, M.J. Thymus zygis Subsp. Gracilis: Watering Level Effect on Phytomass Production and Essential Oil Quality. J. Agric. Food Chem. 2004, 52, 5418–5424. [Google Scholar] [CrossRef]
- Gonçalves, M.J.; Cruz, M.T.; Cavaleiro, C.; Lopes, M.C.; Salgueiro, L. Chemical, antifungal and cytotoxic evaluation of the essential oil of Thymus zygis subsp. sylvestris. Ind. Crops Prod. 2010, 32, 70–75. [Google Scholar] [CrossRef]
- Blanco Salas, J.; Ruiz Téllez, T.; Vázquez Pardo, F.M.; Cases Capdevila, M.A.; Pérez-Alonso, M.J.; Gervasini Rodríguez, C. Short communication. Influence of phenological stage on the antioxidant activity of Thymus zygis s. l. essential oil. Span. J. Agric. Res. 2012, 2, 461–465. [Google Scholar] [CrossRef] [Green Version]
- Rota, M.C.; Herrera, A.; Martínez, R.M.; Sotomayor, J.A.; Jordán, M.J. Antimicrobial activity and chemical composition of Thymus vulgaris, Thymus zygis and Thymus hyemalis essential oils. Food Control 2008, 19, 681–687. [Google Scholar] [CrossRef]
- Soares, J.R.; Dinis, T.C.; Cunha, A.P.; Almeida, L. Antioxidant activities of some extracts of Thymus zygis. Free Radic. Res. 1997, 26, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Stankovic, M.S.; Radic, Z.S.; Blanco-Salas, J.; Vazquez-Pardo, F.M.; Ruiz-Tellez, T. Screening of selected species from Spanish flora as a source of bioactive substances. Ind. Crops Prod. 2017, 95, 493–501. [Google Scholar] [CrossRef]
- Jordán, M.J.; Martínez, R.M.; Martínez, C.; Moñino, I.; Sotomayor, J.A. Polyphenolic extract and essential oil quality of Thymus zygis ssp. gracilis shrubs cultivated under different watering levels. Ind. Crops Prod. 2009, 29, 145–153. [Google Scholar] [CrossRef]
- Afonso, A.F.; Pereira, O.R.; Válega, M.; Silva, A.M.S.; Cardoso, S.M. Metabolites and Biological Activities of Thymus zygis, Thymus pulegioides, and Thymus fragrantissimus Grown under Organic Cultivation. Molecules 2018, 23, 1514. [Google Scholar] [CrossRef] [Green Version]
- Khouya, T.; Ramchoun, M.; Hmidani, A.; Amrani, S.; Harnafi, H.; Benlyas, M.; Filali Zegzouti, Y.; Alem, C. Anti-inflammatory, anticoagulant and antioxidant effects of aqueous extracts from Moroccan thyme varieties. Asian Pac. J. Trop. Biomed. 2015, 5, 636–644. [Google Scholar] [CrossRef] [Green Version]
- Hmidani, A.; Bouhlali, E.D.T.; Khouya, T.; Ramchoun, M.; Filali-Zegzouti, Y.; Alem, C.; Benlyas, M. Antioxidant, anti-inflammatory and anticoagulant activities of three Thymus species grown in southeastern Morocco. Future J. Pharm. Sci. 2019, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- European Medicines Agency. Committee on Herbal Medicinal Products. Community herbal monograph on Thymus vulgaris L. and Thymus zygis L., herba. Available online: https://www.ema.europa.eu/en/documents/herbal-monograph/final-community-herbal-monograph-thymus-vulgaris-l-thymus-zygis-l-herba_en.pdf (accessed on 15 April 2020).
- European Medicines Agency. Thyme: Thymus vulgaris L. and Thymus zygis L., herba. Available online: https://www.ema.europa.eu/en/documents/herbal-summary/thyme-summary-public_en.pdf (accessed on 15 April 2020).
- Nieto, G.; Bañón, S.; Garrido, M.D. Effect of supplementing ewes’ diet with thyme (Thymus zygis ssp. gracilis) leaves on the lipid oxidation of cooked lamb meat. Food Chem. 2011, 125, 1147–1152. [Google Scholar] [CrossRef]
- Boutoial, K.; Garcia, V.; Rovira, S.; Ferrandini, E.; Abdelkhalek, O.; Lopez, M.B. Effect of feeding goats with distilled and non-distilled thyme leaves (Thymus zygis subp. gracilis) on milk and cheese properties. J. Dairy Res. 2013, 80, 448–456. [Google Scholar] [CrossRef]
- Direção-Geral de Agricultura e Desenvolvimento Rural (DGADR). Queijo Rabaçal PDO. Available online: https://tradicional.dgadr.gov.pt/en/categories/cheese-and-other-dairy-products/374-queijo-rabacal-dop-en (accessed on 15 April 2020).
- Martins-Gomes, C.; Taghouti, M.; Schäfer, J.; Bunzel, M.; Silva, A.M.; Nunes, F.M. Chemical characterization and bioactive properties of decoctions and hydroethanolic extracts of Thymus carnosus Boiss. J. Funct. Foods 2018, 43, 154–164. [Google Scholar] [CrossRef]
- Machado, M.; Felizardo, C.; Fernandes-Silva, A.A.; Nunes, F.M.; Barros, A. Polyphenolic compounds, antioxidant activity and L-phenylalanine ammonia-lyase activity during ripening of olive cv. “Cobrancosa” under different irrigation regimes. Food Res. Int. 2013, 51, 412–421. [Google Scholar] [CrossRef]
- Ferreira, S.S.; Silva, A.M.; Nunes, F.M. Citrus reticulata Blanco peels as a source of antioxidant and anti-proliferative phenolic compounds. Ind. Crops Prod. 2018, 111, 141–148. [Google Scholar] [CrossRef]
- Ferreira, S.S.; Silva, P.; Silva, A.M.; Nunes, F.M. Effect of harvesting year and elderberry cultivar on the chemical composition and potential bioactivity: A three-year study. Food Chem. 2020, 302, 125366. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Tang, M.C.; Wu, J.M. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Taghouti, M.; Martins-Gomes, C.; Schafer, J.; Felix, L.M.; Santos, J.A.; Bunzel, M.; Nunes, F.M.; Silva, A.M. Thymus pulegioides L. as a rich source of antioxidant, anti-proliferative and neuroprotective phenolic compounds. Food Funct. 2018, 9, 3617–3629. [Google Scholar] [CrossRef] [PubMed]
- Sreejayan Rao, M.N.A. Nitric oxide scavenging by curcuminoids. J. Pharm. Pharmcol. 1997, 49, 105–107. [Google Scholar] [CrossRef]
- Carbone, C.; Martins-Gomes, C.; Caddeo, C.; Silva, A.M.; Musumeci, T.; Pignatello, R.; Puglisi, G.; Souto, E.B. Mediterranean essential oils as precious matrix components and active ingredients of lipid nanoparticles. Int. J. Pharm. 2018, 548, 217–226. [Google Scholar] [CrossRef]
- Andreani, T.; Kiill, C.P.; de Souza, A.L.R.; Fangueiro, J.F.; Fernandes, L.; Doktorovova, S.; Santos, D.L.; Garcia, M.L.; Gremiao, M.P.D.; Souto, E.B.; et al. Surface engineering of silica nanoparticles for oral insulin delivery: Characterization and cell toxicity studies. Colloid Surf. B 2014, 123, 916–923. [Google Scholar] [CrossRef] [Green Version]
- Severino, P.; Andreani, T.; Jager, A.; Chaud, M.V.; Santana, M.H.A.; Silva, A.M.; Souto, E.B. Solid lipid nanoparticles for hydrophilic biotech drugs: Optimization and cell viability studies (Caco-2 & HEPG-2 cell lines). Eur. J. Med. Chem. 2014, 81, 28–34. [Google Scholar] [CrossRef]
- Silva, A.M.; Martins-Gomes, C.; Coutinho, T.E.; Fangueiro, J.F.; Sanchez-Lopez, E.; Pashirova, T.N.; Andreani, T.; Souto, E.B. Soft Cationic Nanoparticles for Drug Delivery: Production and Cytotoxicity of Solid Lipid Nanoparticles (SLNs). Appl. Sci. 2019, 9, 4438. [Google Scholar] [CrossRef] [Green Version]
- Taghouti, M.; Martins-Gomes, C.; Schäfer, J.; Santos, J.A.; Bunzel, M.; Nunes, F.M.; Silva, A.M. Chemical Characterization and Bioactivity of Extracts from Thymus mastichina: A Thymus with a Distinct Salvianolic Acid Composition. Antioxidants 2020, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Taghouti, M.; Martins-Gomes, C.; Félix, L.M.; Schäfer, J.; Santos, J.A.; Bunzel, M.; Nunes, F.M.; Silva, A.M. Polyphenol composition and biological activity of Thymus citriodorus and Thymus vulgaris: Comparison with endemic Iberian Thymus species. Food Chem. 2020. (under review). [Google Scholar]
- Martins-Gomes, C.; Nunes, F.M.; Sampaio, A.; Souto, E.B.; Silva, A.M. Rosmarinic Acid: Sources, Bioactivities and Health Benefits. In Phytochemicals: Plant Sources and Potential Health Benefits; Ryan, I., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2019; pp. 109–146. [Google Scholar]
- Ziani, B.E.C.; Heleno, S.A.; Bachari, K.; Dias, M.I.; Alves, M.J.; Barros, L.; Ferreira, I.C.F.R. Phenolic compounds characterization by LC-DAD- ESI/MSn and bioactive properties of Thymus algeriensis Boiss. & Reut. and Ephedra alata Decne. Food Res. Int. 2019, 116, 312–319. [Google Scholar] [CrossRef] [Green Version]
- Janicsák, G.; Veres, K.; Zoltán Kakasy, A.; Máthé, I. Study of the oleanolic and ursolic acid contents of some species of the Lamiaceae. Biochem. Syst. Ecol. 2006, 34, 392–396. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C.; Aruoma, O.I. The deoxyribose method: A simple “test-tube” assay for determination of rate constants for reactions of hydroxyl radicals. Anal. Biochem. 1987, 165, 215–219. [Google Scholar] [CrossRef]
- Aruoma, O.I.; Chaudhary, S.S.; Grootveld, M.; Halliwell, B. Binding of iron(II) ions to the pentose sugar 2-deoxyribose. J. Inorg. Biochem. 1989, 35, 149–155. [Google Scholar] [CrossRef]
- Dorman, H.J.D.; Peltoketo, A.; Hiltunen, R.; Tikkanen, M.J. Characterisation of the antioxidant properties of de-odourised aqueous extracts from selected Lamiaceae herbs. Food Chem. 2003, 83, 255–262. [Google Scholar] [CrossRef]
- Chung, S.; Osawa, T.; Kawakishi, S. Hydroxyl radical scavenging effects of spices and scavengers from brown mustard. Biosci. Biotechnol. Biochem. 1997, 61, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Petrović, S.; Ušjak, L.; Milenković, M.; Arsenijević, J.; Drobac, M.; Drndarević, A.; Niketić, M. Thymus dacicus as a new source of antioxidant and antimicrobial metabolites. J. Funct. Foods 2017, 28, 114–121. [Google Scholar] [CrossRef]
- Silva, A.M.; Alvarado, H.L.; Abrego, G.; Martins-Gomes, C.; Garduño-Ramirez, M.L.; García, M.L.; Calpena, A.C.; Souto, E.B. In Vitro Cytotoxicity of Oleanolic/Ursolic Acids-Loaded in PLGA Nanoparticles in Different Cell Lines. Pharmaceutics 2019, 11, 362. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Guo, W.-J.; Yang, Q.-Y. Effects of ursolic acid and oleanolic acid on human colon carcinoma cell line HCT15. World J. Gastroenterol. 2002, 8, 493–495. [Google Scholar] [CrossRef]
- Wiemann, J.; Heller, L.; Csuk, R. Targeting cancer cells with oleanolic and ursolic acid derived hydroxamates. Bioorganic Med. Chem. Lett. 2016, 26, 907–909. [Google Scholar] [CrossRef] [PubMed]
- Sharmila, R.; Manoharan, S. Anti-tumor activity of rosmarinic acid in 7, 12-dimethylbenz (a) anthracene (DMBA) induced skin carcinogenesis in Swiss albino mice. Indian J. Exp. Biol. 2012, 50, 187–194. [Google Scholar]
- Wu, C.-F.; Hong, C.; Klauck, S.M.; Lin, Y.-L.; Efferth, T. Molecular mechanisms of rosmarinic acid from Salvia miltiorrhiza in acute lymphoblastic leukemia cells. J. Ethnopharmacol. 2015, 176, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Tang, L.; Yi, Q. Salvianolic Acids: Potential Source of Natural Drugs for the Treatment of Fibrosis Disease and Cancer. Front Pharmacol. 2019, 10, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, C.-Y.; Ho, Y.-C.; Lin, C.-W.; Yang, W.-E.; Yu, Y.-L.; Tsai, M.-C.; Yang, S.-F.; Su, S.-C. Salvianolic acid A suppresses MMP-2 expression and restrains cancer cell invasion through ERK signaling in human nasopharyngeal carcinoma. J. Ethnopharmacol. 2020, 252, 112601. [Google Scholar] [CrossRef]
- Ismaili, H.; Sosa, S.; Brkic, D.; Fkih-Tetouani, S.; Ilidrissi, A.; Touati, D.; Aquino, R.P.; Tubaro, A. Topical anti-inflammatory activity of extracts and compounds from Thymus broussonettii. J. Pharm. Pharmcol. 2002, 54, 1137–1140. [Google Scholar] [CrossRef]
- Laszczyk, M.N. Pentacyclic triterpenes of the lupane, oleanane and ursane group as tools in cancer therapy. Planta Med. 2009, 75, 1549–1560. [Google Scholar] [CrossRef] [Green Version]
- Checker, R.; Sandur, S.K.; Sharma, D.; Patwardhan, R.S.; Jayakumar, S.; Kohli, V.; Sethi, G.; Aggarwal, B.B.; Sainis, K.B. Potent anti-inflammatory activity of ursolic acid, a triterpenoid antioxidant, is mediated through suppression of NF-kappaB, AP-1 and NF-AT. PLoS ONE 2012, 7, e31318. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, Y.; Murakami, A.; Ohigashi, H. Ursolic acid: An anti- and pro-inflammatory triterpenoid. Mol. Nutr. Food Res. 2008, 52, 26–42. [Google Scholar] [CrossRef]
- Mlala, S.; Oyedeji, O.A.; Gondwe, M.; Oyedeji, O.O. Ursolic Acid and Its Derivatives as Bioactive Agents. Molecules 2019, 24, 2751. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; He, Y.; Yin, T.; Chen, H.; Gao, S.; Hu, M. Metabolism of Phenolic Compounds in LPS-stimulated Raw264.7 Cells Can Impact Their Anti-inflammatory efficacy: Indication of Hesperetin. J. Agric. Food Chem. 2018, 66, 6042–6052. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Pan, M.-H.; Wu, Q.-L.; Park, C.-H.; Juliani, H.R.; Ho, C.-T.; Simon, J.E. LC-MS method for the simultaneous quantitation of the anti-inflammatory constituents in oregano (Origanum species). J. Agric. Food Chem. 2010, 58, 7119–7125. [Google Scholar] [CrossRef] [PubMed]
- Sobeh, M.; Rezq, S.; Cheurfa, M.; Abdelfattah, A.O.M.; Rashied, M.H.R.; El-Shazly, M.A.; Yasri, A.; Wink, M.; Mahmoud, F.M. Thymus algeriensis and Thymus fontanesii: Chemical Composition, In Vivo Antiinflammatory, Pain Killing and Antipyretic Activities: A Comprehensive Comparison. Biomolecules 2020, 10, 599. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hydroethanolic Extract | Aqueous Decoction | ||
|---|---|---|---|
| Extraction yield (%, w/w) | 22.83 ± 0.96 | 29.70 ± 0.99 * | |
| Total phenols (mg Caffeic acid eq./g) | Ext. | 195.81 ± 7.07 | 97.31 ± 7.67 * |
| D.P. | 44.70 ± 1.61 | 28.90 ± 2.28 * | |
| Total flavonoids (mg Catechin eq./g) | Ext. | 269.49 ± 10.39 | 124.80 ± 11.62 * |
| D.P. | 61.52 ± 2.37 | 37.07 ± 3.45 * | |
| Ortho-diphenols (mg Caffeic acid eq./g) | Ext. | 139.79 ± 1.28 | 78.55 ± 0.80 * |
| D.P. | 31.91 ± 0.29 | 23.33 ± 0.24 * | |
| ABTS•+ (mmol Trolox eq./g) | Ext. | 1.08 ± 0.15 | 0.76 ± 0.14 |
| D.P. | 0.25 ± 0.03 | 0.23 ± 0.04 | |
| •OH radicals + EDTA (% inhibition) | 66.28 ±1.20 | ||
| •OH radicals − EDTA (% inhibition) | 43.15 ±2.88 | ||
| NO• radicals (% inhibition, after 120 min) | 29.32 ± 1.67 |
| Compound | R.T. (min) | ESI-MS2 | Quantification | |||||
|---|---|---|---|---|---|---|---|---|
| HE | AD | E. M. E. | ||||||
| mg/g D.P. | mg/g Extract | mg/g D.P. | mg/g Extract | |||||
| 1 | Salvianic acid A | 18.48 ± 0.22 | [197] | n.q. | n.q. | 0.17 ± 0.01 | 0.58 ± 0.4 | |
| 2 | Eriodictyol-di-O-hexoside | 21.81 ± 0.19 | [611]:449;287 | n.q. | n.q. | n.d. | n.d. | |
| 3 | Chlorogenic acid | 23.72 ± 0.21 | [353]:191;179;173;135 | n.q. | n.q. | n.d. | n.d. | |
| 4 | Unknown | 24.72 ± 0.22 | [563]:545;517;455 | n.q. | n.q. | n.q. | n.q. | |
| 5 | Hydroxyjasmonic acid–hexoside | 24.89 ± 0.10 | [387]:369;225;207;163 | 0.03 ± 0.01 | 0.14 ± 0.03 | 0.31 ± 0.01 | 1.07 ± 0.02 | * |
| 6 | Apigenin-(6,8)-C-diglucoside | 25.02 ± 0.18 | [593]:575;503;473;383 353 | 0.17 ± 0.04 | 0.76 ± 0.15 | 0.56 ± 0.03 | 1.85 ± 0.1 | * |
| 7 | Caffeic acid | 25.52 ± 0.22 | [179]:135 | 0.08 ± 0.02 | 0.36 ± 0.09 | 0.56 ± 0.01 | 1.89 ± 0.02 | * |
| 8 | Unknown | 26.05 ± 0.19 | [495]:486;451;375;368 | n.q. | n.q. | n.q. | n.q. | |
| 9 | Eriodictyol-(?)-O-hexoside | 26.16 ± 0.23 | [449]:287 | 2.00 ± 0.26 | 8.77 ± 1.13 | 1.43 ± 0.05 | 4.80 ± 0.17 | * |
| 10 | Unknown | 27.95 ± 0.95 | [367]:193;173;155;137;111 | n.q. | n.q. | n.q. | n.q. | |
| 11 | Prolithospermic acid | 28.73 ± 0.25 | [357]:313;269;245;203 | n.d. | n.d. | n.q. | n.q. | |
| 12 | Naringenin-O-hexoside | 29.4 ± 0.26 | [433]:313;271;267;137 | n.d. | n.d. | n.q. | n.q. | |
| 13 | Quercetin-(?)-O-hexoside | 30.48 ± 0.04 | [463]:301 | 0.92 ± 0.17 | 4.05 ± 0.76 | 0.39 ± 0.06 | 1.31 ± 0.22 | |
| 14 | Naringenin-O-hexoside | 30.88 ± 0.34 | [433]:313;271 | n.q. | n.q. | n.q. | n.q. | |
| 15 | Luteolin-O-hexoside | 32.74 ± 0.41 | [447]:285 | 0.83 ± 0.12 | 3.64 ± 0.51 | 0.70 ± 0.05 | 2.38 ± 0.16 | * |
| 16 | Luteolin-(?)-O-rutinoside | 33.87 ± 0.30 | [593]:285 | n.q. | n.q. | n.q. | n.q. | |
| 17 | Quercetin-(?)-O-hexuronide | 34.42 ± 0.28 | [477]:301 | n.q. | n.q. | n.q. | n.q. | |
| 18 | Luteolin-(?)-O-hexoside | 34.84 ± 0.30 | [447]:285 | 4.44 ± 0.57 | 19.46 ± 2.49 | 4.23 ± 0.30 | 14.23 ± 1.00 | |
| 19 | Salvianolic acid B/E isomer 2 | 37.30 ± 0.28 | [717]:555;519;475;357;295 | n.q. | n.q. | n.q. | n.q. | |
| 20 | Quercetin-(?)-acetyl-hexoside | 38.06 ± 0.38 | [549]:531;505;486;416;345;301 | 1.03 ± 0.22 | 4.52 ± 0.87 | 0.98 ± 0.15 | 3.30 ± 0.5 | |
| 21 | Salvianolic acid A isomer | 38.46 ± 0.34 | [493]:383;313;295 | n.q. | n.q. | n.d. | n.d. | |
| 22 | Luteolin-(?)-O-hexorunide | 38.77 ± 0.34 | [461]:285;175 | 2.92 ± 0.32 | 12.78 ± 1.40 | 3.01 ± 0.32 | 10.14 ± 1.06 | |
| 23 | Chrysoeriol-(?)-O-hexoside | 39.90 ± 0.33 | [461]:299;160 | n.q. | n.q. | n.q. | n.q. | |
| 24 | Rosmarinic acid | 39.44 ± 0.38 | [359]:223;179;161 | 11.11 ± 1.39 | 48.65 ± 5.34 | 4.18 ± 0.78 | 14.07 ± 2.62 | * |
| 25 | Salvianolic acid I | 41.37 ± 0.88 | [537]:493;448;359;339;313;295 | 3.31 ± 0.48 | 14.52 ± 2.10 | 0.95 ± 0.20 | 3.21 ± 0.65 | * |
| 26 | Salvianolic acid K | 42.33 ± 0.30 | [555]:537;493;359 | 2.36 ± 0.37 | 10.33 ± 1.62 | 2.11 ± 0.30 | 7.10 ± 1.03 | |
| 27 | Quercetin-(?)-O-hexoside-hexuronide | 43.36 ± 0.38 | [639]:301 | n.q. | n.q. | n.q. | n.q. | |
| 28 | Apigenin-(?)-O-hexuronide | 44.73 ± 0.44 | [445]:269;175 | n.q. | n.q. | n.q. | n.q. | |
| 29 | Chrysoeriol-(?)-O-hexuronide | 45.86 ± 0.30 | [475]:299 | n.q. | n.q. | n.q. | n.q. | |
| 30 | Unknown | 49.50 ± 0.44 | [551]:519;359;339;313;221;179 | n.q. | n.q. | n.d. | n.d. | |
| 31 | Oleanolic acid a | 35.85 ± 0.05 | - | 0.22 ± 0.03 | 0.99 ± 0.15 | n.d. | n.d. | |
| 32 | Ursolic acid a | 36.91 ± 0.05 | - | 0.48 ± 0.08 | 2.17 ± 0.35 | n.d. | n.d. | |
| Total phenolic compounds | 29.22 ± 3.47 | 127.98 ± 15.20 | 19.58 ± 2.25 | 65.93 ± 7.56 | * | |||
| Total phenolic acid | 16.90 ± 2.03 | 74.00 ± 8.92 | 8.30 ± 1.29 | 27.93 ± 4.35 | * | |||
| Total flavonoids | 12.32 ± 1.44 | 53.98 ± 6.31 | 11.28 ± 0.96 | 37.99 ± 3.21 | ||||
| IC50 (µg/mL) | Exposure Time Effect | Extraction Method Effect | ||||
|---|---|---|---|---|---|---|
| AD | HE | AD | HE | |||
| Caco-2 | 24 h | 746.10 ± 6.35 | 202.20 ± 5.59 | * | * | * |
| 48 h | 604.70 ± 6.70 | 85.01 ± 15.10 | * | |||
| HepG2 | 24 h | 719.20 ± 8.65 | 264.90 ± 10.03 | * | * | * |
| 48 h | 638.02 ± 5.24 | 82.19 ± 2.46 | * | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.M.; Martins-Gomes, C.; Souto, E.B.; Schäfer, J.; Santos, J.A.; Bunzel, M.; Nunes, F.M. Thymus zygis subsp. zygis an Endemic Portuguese Plant: Phytochemical Profiling, Antioxidant, Anti-Proliferative and Anti-Inflammatory Activities. Antioxidants 2020, 9, 482. https://doi.org/10.3390/antiox9060482
Silva AM, Martins-Gomes C, Souto EB, Schäfer J, Santos JA, Bunzel M, Nunes FM. Thymus zygis subsp. zygis an Endemic Portuguese Plant: Phytochemical Profiling, Antioxidant, Anti-Proliferative and Anti-Inflammatory Activities. Antioxidants. 2020; 9(6):482. https://doi.org/10.3390/antiox9060482
Chicago/Turabian StyleSilva, Amélia M., Carlos Martins-Gomes, Eliana B. Souto, Judith Schäfer, João A. Santos, Mirko Bunzel, and Fernando M. Nunes. 2020. "Thymus zygis subsp. zygis an Endemic Portuguese Plant: Phytochemical Profiling, Antioxidant, Anti-Proliferative and Anti-Inflammatory Activities" Antioxidants 9, no. 6: 482. https://doi.org/10.3390/antiox9060482