A Mixture of Algae and Extra Virgin Olive Oils Attenuates the Cardiometabolic Alterations Associated with Aging in Male Wistar Rats

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Treatment
2.3. Serum Measurements
2.4. Serum Lipid Extraction and Fatty Acid Analysis
2.5. Experiments of Vascular Reactivity
2.6. Incubation of Aorta Segments in Presence/Absence of Insulin (10−7 M):
2.7. Nitrite and Nitrate Concentrations in the Culture Medium:
2.8. Protein Analysis in Arterial Tissue by Western Blot
2.9. RNA Extraction and Purification
2.10. Quantitative Real-Time PCR
2.11. Statistical Analysis
3. Results
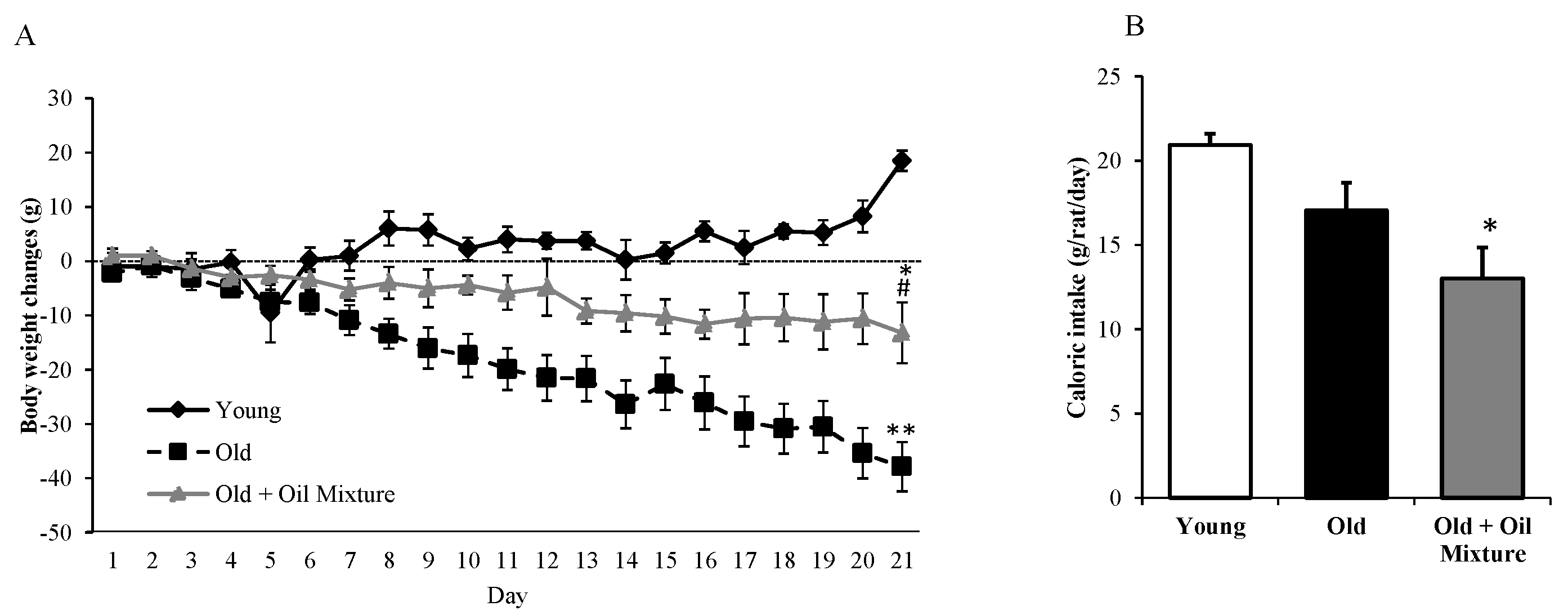
3.1. Body Weight and Food Intake
3.2. Organ Weights
3.3. Lipid Profile, Serum Levels of Metabolic Hormones and HOMA-IR Index
3.4. Serum Inflammatory Parameters
3.5. Percentage of Fatty Acids in the Serum
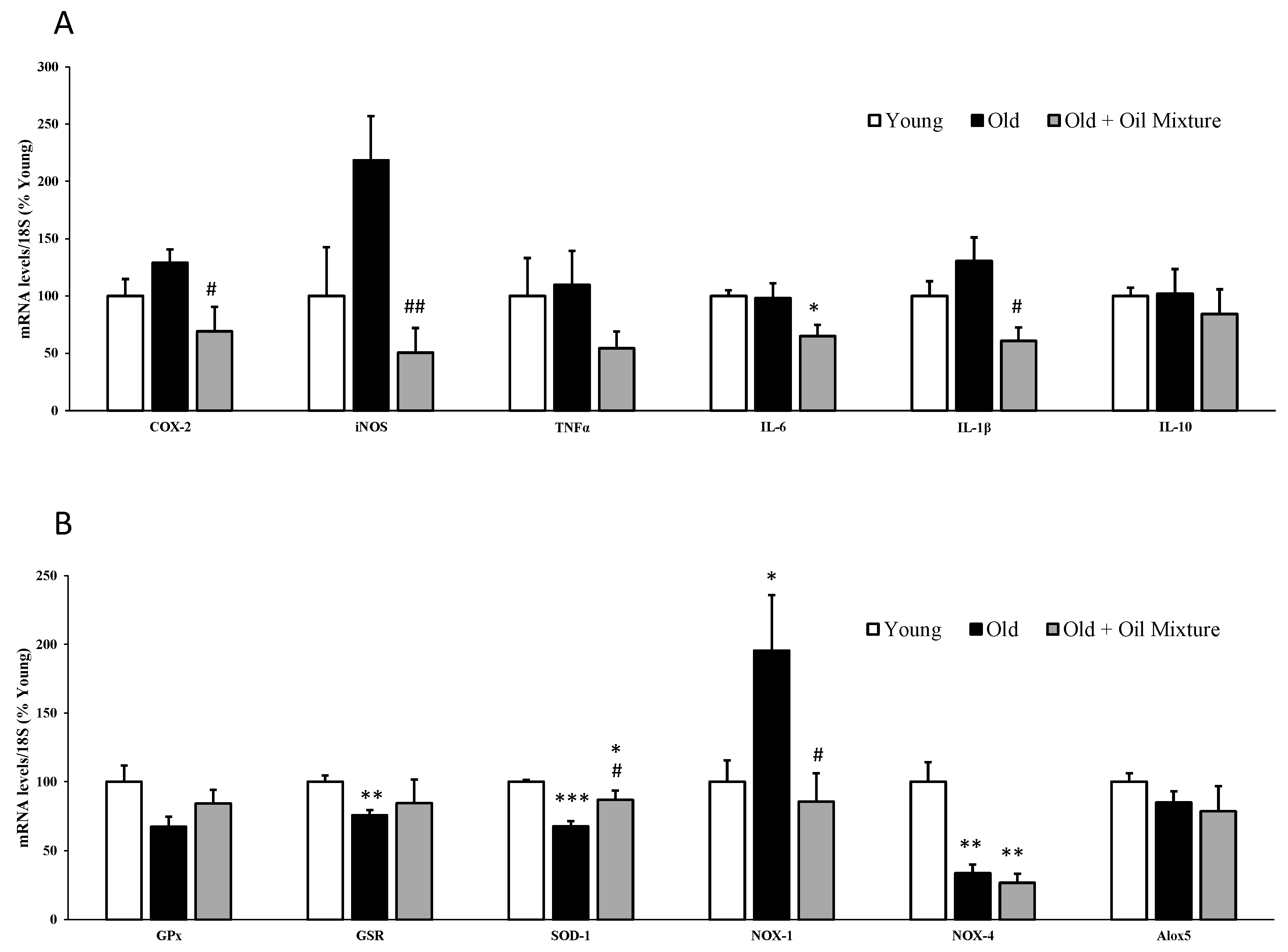
3.6. Gene Expression of Metabolic, Inflammatory and Oxidative Stress Markers in the Liver
3.7. Gene Expression of Inflammatory and Oxidative Stress Markers in the Heart
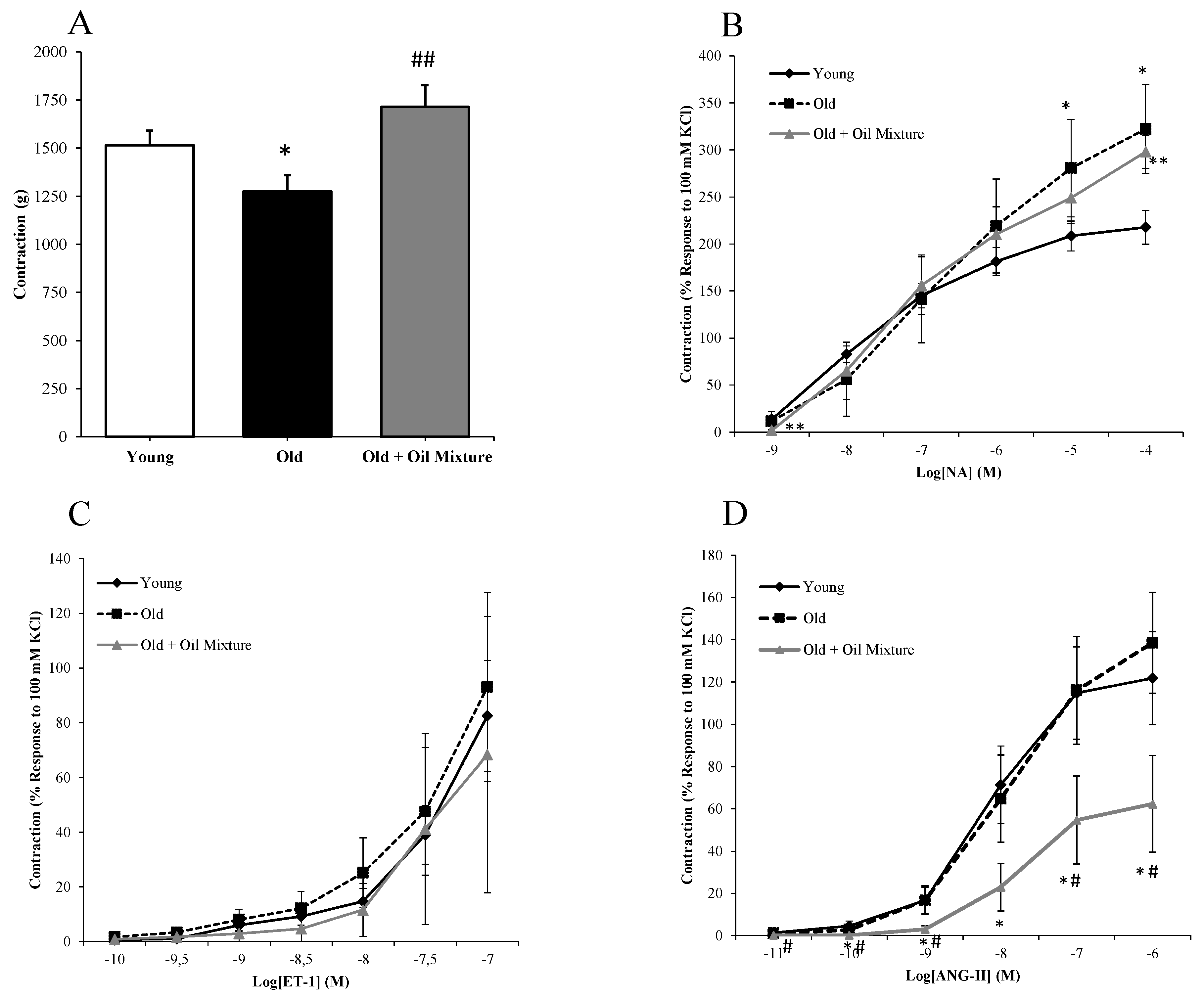
3.8. Aortic Vasoconstriction
3.9. Endothelium-Dependent and Independent Aortic Relaxation
3.10. Aortic Relaxation in Response to Insulin
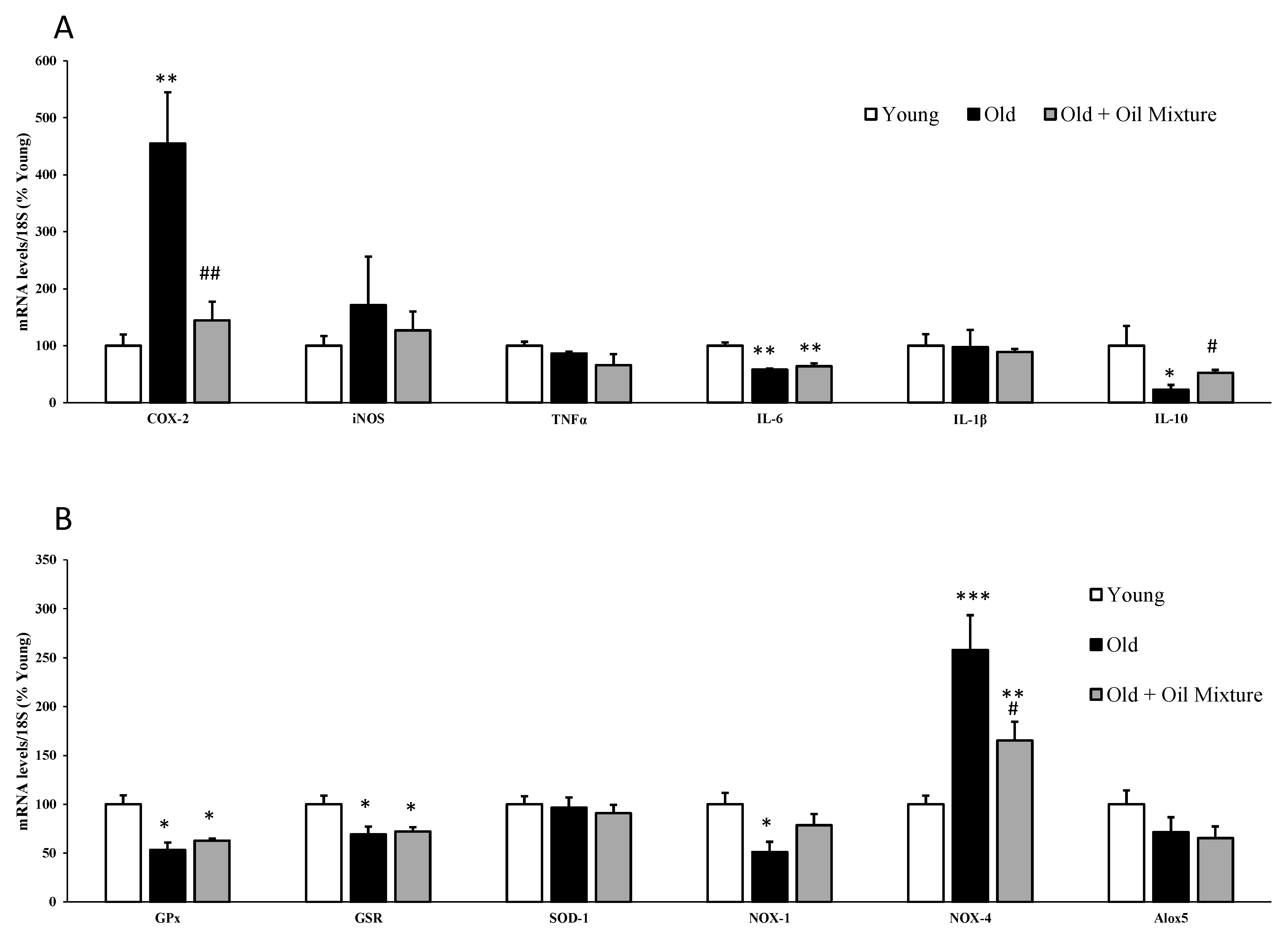
3.11. Gene Expression of Inflammatory and Oxidative Stress Markers in the Aorta
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Veronica, G.; Esther, R.-R.M. Aging, Metabolic Syndrome and the Heart. Aging Dis. 2012, 3, 269–279. [Google Scholar]
- Anderson, R.M.; Weindruch, R. Metabolic reprogramming in dietary restriction. Interdiscip. Top. Gerontol. 2007, 35, 18–38. [Google Scholar] [CrossRef] [PubMed]
- López-Lluch, G.; Navas, P. Calorie restriction as an intervention in ageing. J. Physiol. 2016, 594, 2043–2060. [Google Scholar] [CrossRef]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 Fatty Acids EPA and DHA: Health Benefits Throughout Life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Elherik, K.; Bolton-Smith, C.; Barr, R.; Hill, A.; Murrie, I.; Belch, J. The effects of dietary fatty acid supplementation on endothelial function and vascular tone in healthy subjects. Cardiovasc. Res. 2003, 59, 955–962. [Google Scholar] [CrossRef] [Green Version]
- Morgan, D.R.; Dixon, L.J.; Hanratty, C.G.; El-Sherbeeny, N.; Hamilton, P.B.; McGrath, L.T.; Leahey, W.J.; Johnston, G.D.; McVeigh, G.E. Effects of Dietary Omega-3 Fatty Acid Supplementation on Endothelium-Dependent Vasodilation in Patients with Chronic Heart Failure. Am. J. Cardiol. 2006, 97, 547–551. [Google Scholar] [CrossRef]
- Goodfellow, J.; Bellamy, M.F.; Ramsey, M.W.; Jones, C.J.H.; Lewis, M.J. Dietary supplementation with marine omega-3 fatty acids improve systemic large artery endothelial function in subjects with hypercholesterolemia. J. Am. Coll. Cardiol. 2000, 35, 265–270. [Google Scholar] [CrossRef] [Green Version]
- McVeigh, G.E.; Brennan, G.M.; Johnston, G.D.; McDermott, B.J.; McGrath, L.T.; Henry, W.R.; Andrews, J.W.; Hayes, J.R. Dietary fish oil augments nitric oxide production or release in patients with Type 2 (non-insulin-dependent) diabetes mellitus. Diabetologia 1993, 36, 33–38. [Google Scholar] [CrossRef]
- Gao, H.; Geng, T.; Huang, T.; Zhao, Q. Fish oil supplementation and insulin sensitivity: A systematic review and meta-analysis. Lipids Heal. Dis. 2017, 16, 131. [Google Scholar] [CrossRef] [Green Version]
- Park, M.H.; Kim, D.H.; Lee, E.K.; Kim, N.D.; Im, D.S.; Lee, J.; Yu, B.P.; Chung, H.Y. Age-related inflammation and insulin resistance: A review of their intricate interdependency. Arch. Pharmacol. Res. 2014, 37, 1507–1514. [Google Scholar] [CrossRef] [Green Version]
- Muniyappa, R.; Quon, M. Insulin action and insulin resistance in vascular endothelium. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Amor, S.; Carro, B.M.; Rubio, C.; Carrascosa, J.; Hu, W.; Huang, Y.; García-Villalón, A.L.; Granado, M. Study of insulin vascular sensitivity in aortic rings and endothelial cells from aged rats subjected to caloric restriction: Role of perivascular adipose tissue. Exp. Gerontol. 2018, 109, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Granado, M.; Amor, S.; Carro, B.M.; Guerra-Menéndez, L.; Tejera-Muñoz, A.; González-Hedström, D.; Rubio, C.; Carrascosa, J.M.; García-Villalón, Á.L. Caloric restriction attenuates aging-induced cardiac insulin resistance in male Wistar rats through activation of PI3K/Akt pathway. Nutr. Metab. Cardiovasc. Dis. 2019, 29, 97–105. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2018; ISBN 978-92-5-130562-1. [Google Scholar]
- Naylor, R.L.; Goldburg, R.J.; Primavera, J.H.; Kautsky, N.; Beveridge, M.C.M.; Clay, J.; Folke, C.; Lubchenco, J.; Mooney, H.; Troell, M. Effect of aquaculture on world fish supplies. Nature 2000, 405, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craddock, J.C.; Neale, E.P.; Probst, Y.C.; Peoples, G.E. Algal supplementation of vegetarian eating patterns improves plasma and serum docosahexaenoic acid concentrations and omega-3 indices: A systematic literature review. J. Hum. Nutr. Diet. 2017, 30, 693–699. [Google Scholar] [CrossRef] [Green Version]
- Adarme-Vega, T.C.; Thomas-Hall, S.R.; Schenk, P.M. Towards sustainable sources for omega-3 fatty acids production. Curr. Opin. Biotechnol. 2014, 26, 14–18. [Google Scholar] [CrossRef]
- Nobili, V.; Alisi, A.; Musso, G.; Scorletti, E.; Calder, P.C.; Byrne, C.D. Omega-3 fatty acids: Mechanisms of benefit and therapeutic effects in pediatric and adult NAFLD. Crit. Rev. Clin. Lab. Sci. 2015, 53, 106–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenihan-Geels, G.; Bishop, K.S.; Ferguson, L.R. Alternative Sources of Omega-3 Fats: Can we Find a Sustainable Substitute for Fish? Nutrients 2013, 5, 1301–1315. [Google Scholar] [CrossRef]
- Kazuo, M.; Miyashita, K. Prevention of Fish Oil Oxidation. J. Oleo Sci. 2019, 68, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Parkinson, L.; Cicerale, S. The Health Benefiting Mechanisms of Virgin Olive Oil Phenolic Compounds. Molecules 2016, 21, 1734. [Google Scholar] [CrossRef] [Green Version]
- Soto-Alarcon, S.A.; Valenzuela, R.; Valenzuela, A.; Videla, L. Liver Protective Effects of Extra Virgin Olive Oil: Interaction between Its Chemical Composition and the Cell-signaling Pathways Involved in Protection. Endocrine Metab. Immune Disord. Drug Targets 2017, 18, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Buckland, G.; Mayén, A.-L.; Agudo, A.; Travier, N.; Navarro-Mateu, F.; Sánchez, M.J.; Chirlaque, M.D.; Barricarte, A.; Ardanaz, E.; Moreno-Iribas, C.; et al. Olive oil intake and mortality within the Spanish population (EPIC-Spain). Am. J. Clin. Nutr. 2012, 96, 142–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapsell, L.C. Foods and food components in the Mediterranean diet: Supporting overall effects. BMC Med. 2014, 12, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Gómez, Y.; Lopez-Miranda, J.; Blanco-Colio, L.M.; Marín, C.; Pérez-Martínez, P.; Ruano, J.; Paniagua, J.A.; Rodríguez, F.; Egido, J.; Jiménez, F.P. Olive oil and walnut breakfasts reduce the postprandial inflammatory response in mononuclear cells compared with a butter breakfast in healthy men. Atherosclerosis 2009, 204, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Thandapilly, S.J.; Raj, P.; Louis, X.L.; Perera, D.; Yamanagedara, P.; Zahradka, P.; Taylor, C.G.; Netticadan, T. Canola oil rich in oleic acid improves diastolic heart function in diet-induced obese rats. J. Physiol. Sci. 2016, 67, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.-H.; Chu, P.-M.; Kao, C.-L.; Cheng, Y.-H.; Hung, C.-H.; Tsai, K.-L. Oleic acid activates MMPs up-regulation through SIRT1/PPAR-γ inhibition: A probable linkage between obesity and coronary arterial disease. J. Biochem. 2016, 160, 217–225. [Google Scholar] [CrossRef]
- Perdomo, L.; Beneit, N.; Otero, Y.F.; Escribano, O.; Diaz-Castroverde, S.; Gomez-Hernandez, A.; Benito, M. Protective role of oleic acid against cardiovascular insulin resistance and in the early and late cellular atherosclerotic process. Cardiovasc. Diabetol. 2015, 14, 75. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, F.; Sandra, D.; Vincenzo, C.; Angela, Z.; Taglieri, I.; Francesca, V.; Lara, T. The Nutraceutical Value of Olive Oil and Its Bioactive Constituents on the Cardiovascular System. Focusing on Main Strategies to Slow Down Its Quality Decay during Production and Storage. Nutrients 2019, 11, 1962. [Google Scholar] [CrossRef] [Green Version]
- Ghanbari, R.; Anwar, F.; Alkharfy, K.M.; Gilani, A.-H.; Saari, N. Valuable Nutrients and Functional Bioactives in Different Parts of Olive (Olea europaea L.)—A Review. Int. J. Mol. Sci. 2012, 13, 3291–3340. [Google Scholar] [CrossRef]
- Gonzalez-Hedström, E.A.D.; Granado, M.; García-Inarejos, A.M. Protective Effects of Extra Virgin Olive Oil Components Against Storage-induced Omega 3 Oxidation in Algae Oil. Unpublished.
- Drews, B.; Milojevic, V.; Giller, K.; Ulbrich, S. Fatty acid profile of blood plasma and oviduct and uterine fluid during early and late luteal phase in the horse. Theriogenology 2018, 114, 258–265. [Google Scholar] [CrossRef]
- Olmstead, I.L.; Hill, D.R.; Dias, D.A.; Jayasinghe, N.S.; Callahan, D.; Kentish, S.; Scales, P.J.; Martin, G.J. A quantitative analysis of microalgal lipids for optimization of biodiesel and omega-3 production. Biotechnol. Bioeng. 2013, 110, 2096–2104. [Google Scholar] [CrossRef] [PubMed]
- Miranda, K.M.; Espey, M.G.; Wink, D.A. A Rapid, Simple Spectrophotometric Method for Simultaneous Detection of Nitrate and Nitrite. Nitric Oxide 2001, 5, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P. A reagent for the sencillo-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. Biotechniques 1993, 15, 532. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Maeda, M.; Scaglia, N.; Igal, R.A. Regulation of fatty acid synthesis and Δ9-desaturation in senescence of human fibroblasts. Life Sci. 2009, 84, 119–124. [Google Scholar] [CrossRef]
- Vessby, B.; Tengblad, S.; Lithell, H. Insulin sensitivity is related to the fatty acid composition of serum lipids and skeletal muscle phospholipids in 70-year-old men. Diabetologia 1994, 37, 1044–1050. [Google Scholar] [CrossRef]
- Kim, O.; Lim, H.; Lee, M.; Kim, J.; Lee, J. Association of fatty acid composition in serum phospholipids with metabolic syndrome and arterial stiffness. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 366–374. [Google Scholar] [CrossRef]
- Arnold, L.E.; Young, A.S.; Belury, M.A.; Cole, R.M.; Gracious, B.; Seidenfeld, A.M.; Wolfson, H.; Fristad, M.A. Omega-3 Fatty Acid Plasma Levels Before and after Supplementation: Correlations with Mood and Clinical Outcomes in the Omega-3 and Therapy Studies. J. Child Adolesc. Psychopharmacol. 2017, 27, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Kondo, K.; Morino, K.; Nishio, Y.; Kondo, M.; Nakao, K.; Nakagawa, F.; Ishikado, A.; Sekine, O.; Yoshizaki, T.; Kashiwagi, A.; et al. A fish-based diet intervention improves endothelial function in postmenopausal women with type 2 diabetes mellitus: A randomized cruce trial. Metabolism 2014, 63, 930–940. [Google Scholar] [CrossRef]
- Harris, W.S. The Omega-3 Index: Clinical Utility for Therapeutic Intervention. Curr. Cardiol. Rep. 2010, 12, 503–508. [Google Scholar] [CrossRef]
- Hellhammer, J.; Hero, T.; Franz, N.; Contreras, C.; Schubert, M. Omega-3 fatty acids administered in phosphatidylserine improved certain aspects of high chronic stress in men. Nutr. Res. 2012, 32, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.S.; Lo, M.J.; Kau, M.M. Glucocorticoids and aging. J. Formos. Med. Assoc. 1997, 96, 792–801. [Google Scholar]
- Goldberg, E.L.; Dixit, V.D. Drivers of age-related inflammation and strategies for healthspan extension. Immunol. Rev. 2015, 265, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Khansari, N.; Shakiba, Y.; Mahmoudi, M. Chronic inflammation and oxidative stress as a major cause of age-related diseases and cancer. Recent Patents Inflamm. Allergy Drug Discov. 2009, 3, 73–80. [Google Scholar] [CrossRef]
- de Moraes, I.M.; Gonçalves-De-Albuquerque, C.F.; Kurz, A.R.M.; Oliveira, F.M.D.J.; De Abreu, V.H.P.; Torres, R.; Carvalho, V.F.; Estato, V.; Bozza, P.T.; Sperandio, M.; et al. Omega-9 Oleic Acid, the Main Compound of Olive Oil, Mitigates Inflammation during Experimental Sepsis. Oxidative Med. Cell. Longev. 2018, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acharya, P.; Talahalli, R.R. Aging and Hyperglycemia Intensify Dyslipidemia-Induced Oxidative Stress and Inflammation in Rats: Assessment of Restorative Potentials of ALA and EPA + DHA. Inflammation 2018, 42, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Halade, G.V.; Rahman, M.; Bhattacharya, A.; Barnes, J.L.; Chandrasekar, B.; Fernandes, G.; Barnes, J. Docosahexaenoic acid-enriched fish oil attenuates kidney disease and prolongs median and maximal life span of autoimmune lupus-prone mice. J. Immunol. 2010, 184, 5280–5286. [Google Scholar] [CrossRef] [Green Version]
- Kalupahana, N.S.; Claycombe, K.J.; Moustaid-Moussa, N. (n-3) Fatty Acids Alleviate Adipose Tissue Inflammation and Insulin Resistance: Mechanistic Insights. Adv. Nutr. 2011, 2, 304–316. [Google Scholar] [CrossRef] [Green Version]
- Albracht-Schulte, K.; Kalupahana, N.S.; Ramalingam, L.; Wang, S.; Rahman, S.M.; Robert-McComb, J.; Moustaid-Moussa, N. Omega-3 fatty acids in obesity and metabolic syndrome: A mechanistic update. J. Nutr. Biochem. 2018, 58, 1–16. [Google Scholar] [CrossRef]
- Vessby, B.; Uusitupa, M.; Hermansen, K.; Riccardi, G.; Rivellese, A.A.; Tapsell, L.C.; Nälsén, C.; Berglund, L.; Louheranta, A.; Rasmussen, B.M.; et al. Substituting dietary saturated for monounsaturated fat impairs insulin sensitivity in healthy men and women: The KANWU Study. Diabetologia 2001, 44, 312–319. [Google Scholar] [CrossRef] [Green Version]
- di Nicolantonio, J.J.; O’Keefe, J.H. Good Fats versus Bad Fats: A Comparison of Fatty Acids in the Promotion of Insulin Resistance, Inflammation, and Obesity. Mo. Med. 2017, 114, 303–307. [Google Scholar]
- Heshmati, J.; Morvaridzadeh, M.; Maroufizadeh, S.; Akbari, A.; Yavari, M.; Amirinejad, A.; Maleki-Hajiagha, A.; Sepidarkish, M.; Amirinejhad, A. Omega-3 fatty acids supplementation and oxidative stress parameters: A systematic review and meta-analysis of clinical trials. Pharmacol. Res. 2019, 149, 104462. [Google Scholar] [CrossRef]
- Mollace, V.; Gliozzi, M.; Carresi, C.; Musolino, V.; Oppedisano, F. Re-assessing the mechanism of action of n? 3 PUFAs. Int. J. Cardiol. 2013, 170, 8. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Liu, R.; Xia, C.; Chen, Y.; Dong, Z.; Huang, B.; Li, R.; Li, M.; Xu, C. Effects of different fatty acids on BRL3A rat liver cell damage. J. Cell. Physiol. 2020. [Google Scholar] [CrossRef]
- Bhattacharjee, B.; Pal, P.K.; Chattopadhyay, A.; Bandyopadhyay, D. Oleic acid protects against cadmium induced cardiac and hepatic tissue injury in male Wistar rats: A mechanistic study. Life Sci. 2020, 244, 117324. [Google Scholar] [CrossRef] [PubMed]
- Al-Shudiefat, A.A.R.; Sharma, A.K.; Bagchi, R.; Dhingra, S.; Singal, P.K. Oleic acid mitigates TNF-α-induced oxidative stress in rat cardiomyocytes. Mol. Cell. Biochem. 2012, 372, 75–82. [Google Scholar] [CrossRef]
- Singh, P.K.; Gari, M.; Choudhury, S.; Shukla, A.; Gangwar, N.; Garg, S.K. Oleic Acid Prevents Isoprenaline-Induced Cardiac Injury: Effects on Cellular Oxidative Stress, Inflammation and Histopathological Alterations. Cardiovasc. Toxicol. 2019, 20, 28–48. [Google Scholar] [CrossRef]
- Liu, T.; Wen, H.; Li, H.; Xu, H.; Xiao, N.; Liu, R.; Chen, L.; Sun, Y.; Song, L.; Bai, C.; et al. Oleic Acid Attenuates Ang II (Angiotensin II)-Induced Cardiac Remodeling by Inhibiting FGF23 (Fibroblast Growth Factor 23) Expression in Mice. Hypertension 2020, 75, 680–692. [Google Scholar] [CrossRef]
- Innes, J.; Calder, P.C. Marine Omega-3 (N-3) Fatty Acids for Cardiovascular Health: An Update for 2020. Int. J. Mol. Sci. 2020, 21, 1362. [Google Scholar] [CrossRef] [Green Version]
- Sekikawa, A.; Cui, C.; Sugiyama, D.; Fabio, A.; Harris, W.S.; Zhang, X. Effect of High-Dose Marine Omega-3 Fatty Acids on Atherosclerosis: A Systematic Review and Meta-Analysis of Randomized Clinical Trials. Nutrients 2019, 11, 2599. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Núñez, B.; Dijck-Brouwer, D.J.; Muskiet, F.A. The relation of saturated fatty acids with low-grade inflammation and cardiovascular disease. J. Nutr. Biochem. 2016, 36, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Lennon-Edwards, S.; Schellhardt, T.A.; Kuczmarski, J.M. Antioxidant Defense Is Increased in Aged Hearts Following Omega-3 Supplementation in the Absence of Changes in Inflammation. Physiol. Res. 2015, 64, 433–438. [Google Scholar] [CrossRef]
- Kenny, D.; Warltier, D.C.; Pleuss, J.A.; Hoffmann, R.G.; Goodfriend, T.L.; Egan, B.M. Effect of omega-3 fatty acids on the vascular response to angiotensin in normotensive men. Am. J. Cardiol. 1992, 70, 1347–1352. [Google Scholar] [CrossRef]
- Drenjančević, I.; Jelaković, B.; Lombard, J.; Kunert, M.; Kibel, A.; Gros, M. High-salt diet and hypertension: Focus on the renin-angiotensin system. Kidney Blood Press. Res. 2010, 34, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mazzolai, L.; Hayoz, D. The renin-angiotensin system and atherosclerosis. Curr. Hypertens. Rep. 2006, 8, 47–53. [Google Scholar] [CrossRef]
- Davda, R.K.; Stepniakowski, K.T.; Lu, G.; Ullian, M.E.; Goodfriend, T.L.; Egan, B.M. Oleic Acid Inhibits Endothelial Nitric Oxide Synthase by a Protein Kinase C–Independent Mechanism. Hypertension 1995, 26, 764–770. [Google Scholar] [CrossRef]
- Gremmels, H.; Bevers, L.M.; Fledderus, J.; Braam, B.; Van Zonneveld, A.J.; Verhaar, M.C.; Joles, J. Oleic acid increases mitochondrial reactive oxygen species production and decreases endothelial nitric oxide synthase activity in cultured endothelial cells. Eur. J. Pharmacol. 2015, 751, 67–72. [Google Scholar] [CrossRef]
- Zgheel, F.; Perrier, S.; Remila, L.; Houngue, U.; Mazzucotelli, J.-P.; Morel, O.; Auger, C.; Schini-Kerth, V. EPA: DHA 6:1 is a superior omega-3 PUFAs formulation attenuating platelets-induced contractile responses in porcine coronary and human internal mammary artery by targeting the serotonin pathway via an increased endothelial formation of nitric oxide. Eur. J. Pharmacol. 2019, 853, 41–48. [Google Scholar] [CrossRef]
- Limbu, R.; Cottrell, G.S.; McNeish, A.J. Characterisation of the vasodilation effects of DHA and EPA, n-3 PUFAs (fish oils), in rat aorta and mesenteric resistance arteries. PLoS ONE 2018, 13, e0192484. [Google Scholar] [CrossRef]
- Sato, K.; Chino, D.; Sugimoto, T.; Kanai, K.; Obara, K.; Miyauchi, S.; Tanaka, Y. Pharmacological Characteristics of the Inhibitory Effects of Docosahexaenoic Acid on Vascular Contractions Studied in Rat Mesenteric Artery. Pharmacology 2014, 93, 229–243. [Google Scholar] [CrossRef]
- Sametz, W.; Jeschek, M.; Juan, H.; Wintersteiger, R. Influence of Polyunsaturated Fatty Acids on Vasoconstrictions Induced by 8-iso-PGF2α and 8-iso-PGE2. Pharmacology 2000, 60, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liang, X.; Wang, L.; Lu, X.; Huang, J.; Cao, J.; Li, H.; Gu, D. Effect of omega-3 fatty acids supplementation on endothelial function: A meta-analysis of randomized controlled trials. Atherosclerosis 2012, 221, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Zanetti, M.; Cappellari, G.G.; Barbetta, D.; Semolic, A.; Barazzoni, R. Omega 3 Polyunsaturated Fatty Acids Improve Endothelial Dysfunction in Chronic Renal Failure: Role of eNOS Activation and of Oxidative Stress. Nutrients 2017, 9, 895. [Google Scholar] [CrossRef] [Green Version]
- Villalpando, D.M.; Navarro, R.; del Campo, L.; Largo, C.; Muñoz, D.; Tabernero, M.; Baeza, R.; Otero, C.; García, H.S.; Ferrer, M. Effect of Dietary Docosahexaenoic Acid Supplementation on the Participation of Vasodilator Factors in Aorta from Orchidectomized Rats. PLoS ONE 2015, 10, e0142039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimokawa, H.; Vanhoutte, P.M. Dietary omega 3 fatty acids and endothelium-dependent relaxations in porcine coronary arteries. Am. J. Physiol. Circ. Physiol. 1989, 256, 968–973. [Google Scholar] [CrossRef]
- Gaertner, S.; Auger, C.; Farooq, M.A.; Pollet, B.; Khemais-Benkhiat, S.; Niazi, Z.R.; Schrevens, S.; Park, S.-H.; Toti, F.; Stephan, D.; et al. Oral Intake of EPA: DHA 6:1 by Middle-Aged Rats for One Week Improves Age-Related Endothelial Dysfunction in Both the Femoral Artery and Vein: Role of Cyclooxygenases. Int. J. Mol. Sci. 2020, 21, 920. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.A.; Gaertner, S.; Amoura, L.; Niazi, Z.R.; Park, S.-H.; Qureshi, A.W.; Oak, M.-H.; Toti, F.; Schini-Kerth, V.B.; Auger, C. Intake of omega-3 formulation EPA: DHA 6:1 by old rats for 2 weeks improved endothelium-dependent relaxations and normalized the expression level of ACE/AT1R/NADPH oxidase and the formation of ROS in the mesenteric artery. Biochem. Pharmacol. 2020, 173, 113749. [Google Scholar] [CrossRef]
- Sloboda, N.; Feve, B.; Thornton, S.N.; Nzietchueng, R.; Regnault, V.; Simon, G.; Labat, C.; Louis, H.; Max, J.-P.; Muscat, A.; et al. Fatty Acids Impair Endothelium-Dependent Vasorelaxation: A Enlace Between Obesity and Arterial Stiffness in Very Old Zucker Rats. J. Gerontol. Ser. A Boil. Sci. Med. Sci. 2012, 67, 927–938. [Google Scholar] [CrossRef] [Green Version]
- Zgheel, F.; Alhosin, M.; Rashid, S.; Burban, M.; Auger, C.; Schini-Kerth, V.B. Redox-Sensitive Induction of Src/PI3-kinase/Akt and MAPKs Pathways Activate eNOS in Response to EPA: DHA 6:1. PLoS ONE 2014, 9, e105102. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Wang, S.; Luan, H.; Tuerhong, D.; Lin, Y.; Liang, J.; Xiong, Y.; Rui, L.; Wu, F. Clinopodium chinense Attenuates Palmitic Acid-Induced Vascular Endothelial Inflammation and Insulin Resistance through TLR4-Mediated NF-κB and MAPK Pathways. Am. J. Chin. Med. 2019, 47, 97–117. [Google Scholar] [CrossRef]
- Hart, C.M.; Gupta, M.P.; Evanoff, V. Oleic acid reduces oxidant stress in cultured pulmonary artery endothelial cells. Exp. Lung Res. 1997, 23, 405–425. [Google Scholar] [CrossRef] [PubMed]
- Covas, M.-I.; de la Torre, R.; Fitó, M. Virgin olive oil: A key food for cardiovascular risk protection. Br. J. Nutr. 2015, 113, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-González, M.; Martín-Calvo, N. The major European dietary patterns and metabolic syndrome. Rev. Endocr. Metab. Disord. 2013, 14, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Summerhill, V.; Karagodin, V.; Grechko, A.; Myasoedova, V.; Orekhov, A. Vasculoprotective Role of Olive Oil Compounds via Modulation of Oxidative Stress in Atherosclerosis. Front. Cardiovasc. Med. 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorzynik-Dębicka, M.; Przychodzeń, P.; Cappello, F.; Kuban-Jankowska, A.; Gammazza, A.M.; Knap, N.; Woźniak, M.; Gorska-Ponikowska, M. Potential Health Benefits of Olive Oil and Plant Polyphenols. Int. J. Mol. Sci. 2018, 19, 686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serreli, G.; Deiana, M. Extra Virgin Olive Oil Polyphenols: Modulation of Cellular Pathways Related to Oxidant Species and Inflammation in Aging. Cells 2020, 9, 478. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Young | Old | Old + Oil Mixture | |
|---|---|---|---|
| Heart (mg) | 1503.8 ± 78.8 | 1891.3 ± 55.6 ** | 1530.0 ± 235.0 # |
| Epidydimal visceral adipose tissue (mg) | 9843.1 ± 1075.2 | 20,475.4 ± 1451.2 *** | 20,377.2 ± 1231.4 *** |
| Lumbar subcutaneous adipose tissue (mg) | 4992.4 ± 787.5 | 27,905.7 ± 6193 ** | 22,955.9 ± 4816.7 *** |
| Subscapular brown adipose tissue (mg) | 496.3 ± 50.0 | 810.8 ± 119.4 * | 723.2 ± 107.8 * |
| Periaortic adipose tissue (mg) | 180.0 ± 26.0 | 307.9 ± 47.7 * | 303.3 ± 56.4 * |
| Kidneys (mg) | 2530.2 ± 50.2 | 3298.7 ± 194.2 ** | 2648.7 ± 204.2 # |
| Suprarenal glands (mg) | 61.9 ± 2.4 | 86.0 ± 12.6 * | 54.8 ± 2.3 # |
| Liver (mg) | 13,465.0 ± 425.6 | 14,886.9 ± 566.9 * | 13,481.8 ± 442.9 # |
| Spleen (mg) | 695.7 ± 20.8 | 1132.2 ± 61.5 *** | 923.2 ± 69.0 **# |
| Young | Old | Old + Oil Mixture | |
|---|---|---|---|
| Glycemia (mg/dL) | 90.7 ± 3.8 | 72.4 ± 11.1 | 81.4 ± 4.4 * |
| Total Lipids (mg/dL) | 853 ± 63 | 1051 ± 37 * | 812 ± 97.7 # |
| Triglycerides (mg/dL) | 97.6 ± 13.5 | 158.5 ± 36.4 * | 73.2 ± 11.8 # |
| Total Cholesterol (mg/dL) | 135.1 ± 15.3 | 199.3 ± 13.9 * | 113.8 ± 9.8 ### |
| LDL-cholesterol (mg/dL) | 28.8 ± 2.7 | 47.8 ± 2.6 ** | 21.3 ± 1.8 *### |
| HDL-cholesterol (mg/dL) | 15.7 ± 0.6 | 13.4 ± 2.2 | 10.3 ± 1.2 *** |
| Insulin (ng/mL) | 17.7 ± 5.2 | 81.3 ± 32.2 ** | 20.6 ± 2.8 |
| HOMA-Index | 1.75 ± 0.3 | 13.1 ± 5.4 * | 3.9 ± 0.5 |
| Leptin (ng/mL) | 11.82 ± 1.4 | 30.2 ± 5.9 ** | 37.2 ± 1.7 *** |
| Adiponectin (mg/dL) | 67.1 ± 6.7 | 108.9 ± 4.3 *** | 101.7 ± 11.6 ** |
| Interleukin-6 (pg/mL) | 135.6 ± 7.1 | 188.7 ± 24.1 * | 127.0 ± 19.0 # |
| TNFα (pg/mL) | 0.1 ± 0.1 | 1.8 ± 0.9 * | 0.1 ± 0.1 |
| Young | Old | Old + Oil Mixture | |
|---|---|---|---|
| SFA | 25.0 ± 0.8 | 28.4 ± 0.4 ** | 26.9 ± 0.5 # |
| MUFA | 49.6 ± 0.6 | 42.0 ± 2.2 ** | 46.3 ± 0.5 **# |
| PUFA | 15.2 ± 2.5 | 16.1 ± 1.2 | 20.9 ± 1.4 *# |
| Palmitic Acid (C16:0) | 8.28 ± 0.3 | 9.92 ± 0.7 * | 8.84 ± 0.7 |
| Palmitoleic Acid (C16:1) | 3.52 ± 0.5 | 5.47 ± 0.5 * | 5.74 ± 0.5 ** |
| Stearic Acid (C18:0) | 13.2 ± 0.6 | 13.0 ± 0.5 | 12.3 ± 0.2 |
| Oleic Acid (C18:1) | 47.7 ± 0.7 | 40.8 ± 1.8 ** | 45.1 ± 0.7 *# |
| Linoleic Acid (C18:2) | 1.8 ± 0.2 | 1.1 ± 0.4 | 1.5 ± 0.2 |
| ALA (C18:3) | 3.42 ± 0.2 | 2.7 ± 0.1 ** | 3.3 ± 0.4 |
| DHA (C22:6n-3) | 7.1 ± 1.5 | 6.9 ± 0.7 | 9.5 ± 0.7 # |
| EPA (C20:5n-3) | 4.7 ± 1.2 | 6.5 ± 0.5 | 8.2 ± 0.7 *# |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Hedström, D.; Amor, S.; de la Fuente-Fernández, M.; Tejera-Muñoz, A.; Priego, T.; Martín, A.I.; López-Calderón, A.; Inarejos-García, A.M.; García-Villalón, Á.L.; Granado, M. A Mixture of Algae and Extra Virgin Olive Oils Attenuates the Cardiometabolic Alterations Associated with Aging in Male Wistar Rats. Antioxidants 2020, 9, 483. https://doi.org/10.3390/antiox9060483
González-Hedström D, Amor S, de la Fuente-Fernández M, Tejera-Muñoz A, Priego T, Martín AI, López-Calderón A, Inarejos-García AM, García-Villalón ÁL, Granado M. A Mixture of Algae and Extra Virgin Olive Oils Attenuates the Cardiometabolic Alterations Associated with Aging in Male Wistar Rats. Antioxidants. 2020; 9(6):483. https://doi.org/10.3390/antiox9060483
Chicago/Turabian StyleGonzález-Hedström, Daniel, Sara Amor, María de la Fuente-Fernández, Antonio Tejera-Muñoz, Teresa Priego, Ana Isabel Martín, Asunción López-Calderón, Antonio Manuel Inarejos-García, Ángel Luís García-Villalón, and Miriam Granado. 2020. "A Mixture of Algae and Extra Virgin Olive Oils Attenuates the Cardiometabolic Alterations Associated with Aging in Male Wistar Rats" Antioxidants 9, no. 6: 483. https://doi.org/10.3390/antiox9060483