Combined Application of Bacteriophages and Carvacrol in the Control of Pseudomonas syringae pv. actinidiae Planktonic and Biofilm Forms


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Preparation of Phages
2.3. Preparation of Carvacrol
2.4. Evaluation of Phage Sensitivity to Carvacrol
2.5. Evaluation of Phage Bacteriolytic Activity In Vitro
2.6. Determination of Antimicrobial Activity of Carvacrol
2.7. Evaluation of the Efficacy of Combined Phages and Carvacrol against Psa
2.8. Biofilm Inhibition Assay
2.9. Pre-Formed Biofilm Destruction Assay
2.10. Statistical Analysis
3. Results
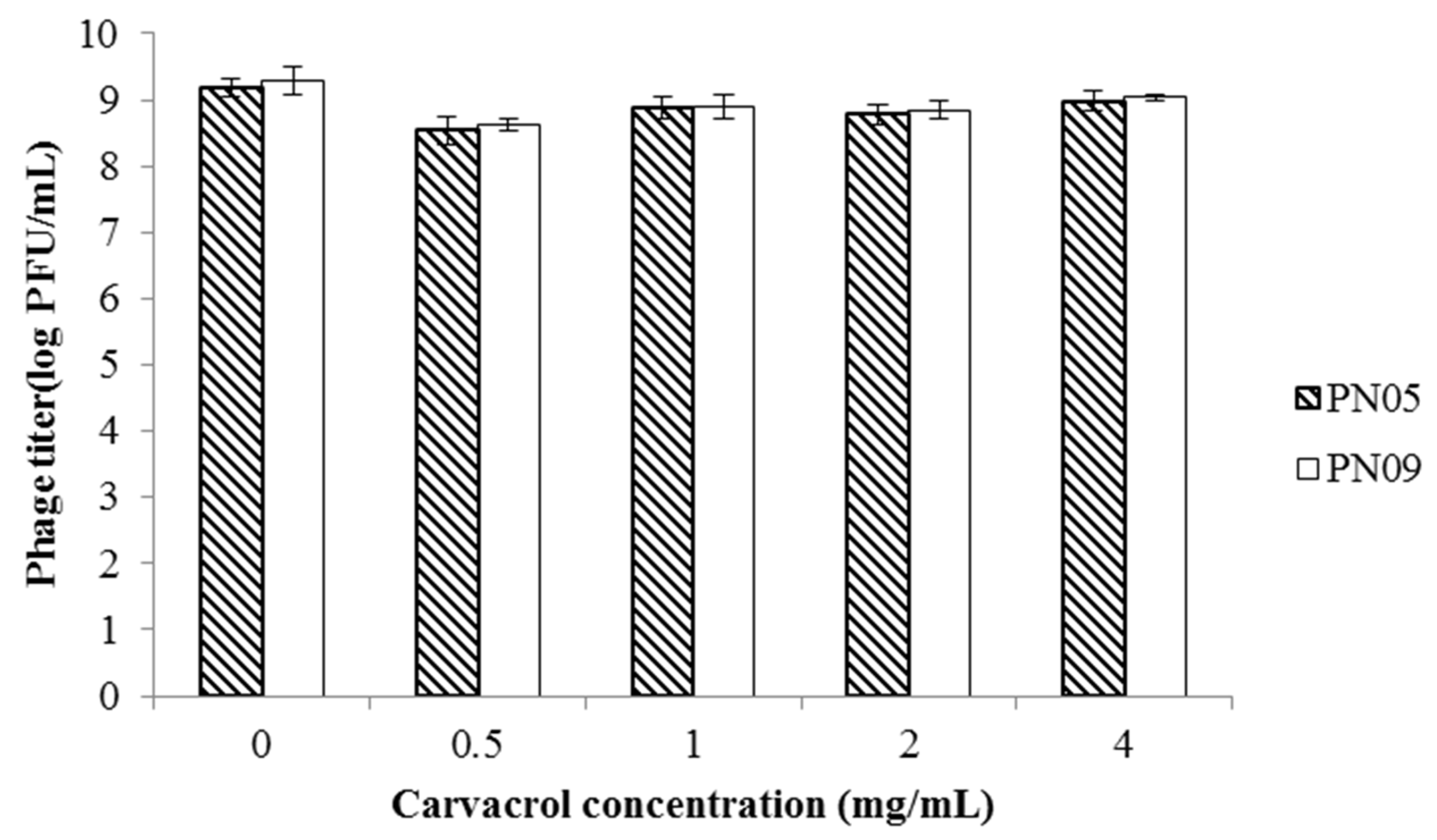
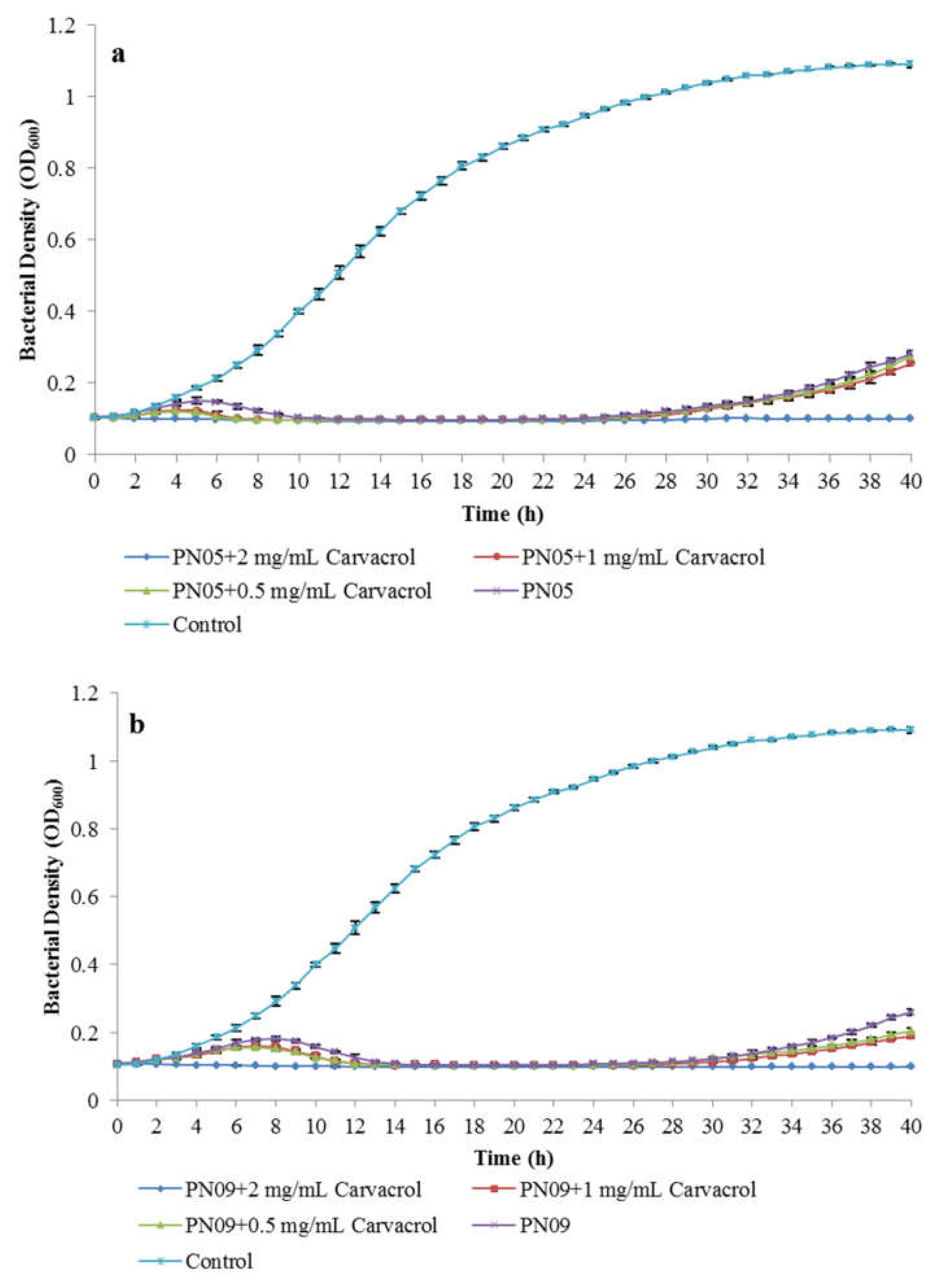
3.1. Phage Sensitivity to Carvacrol
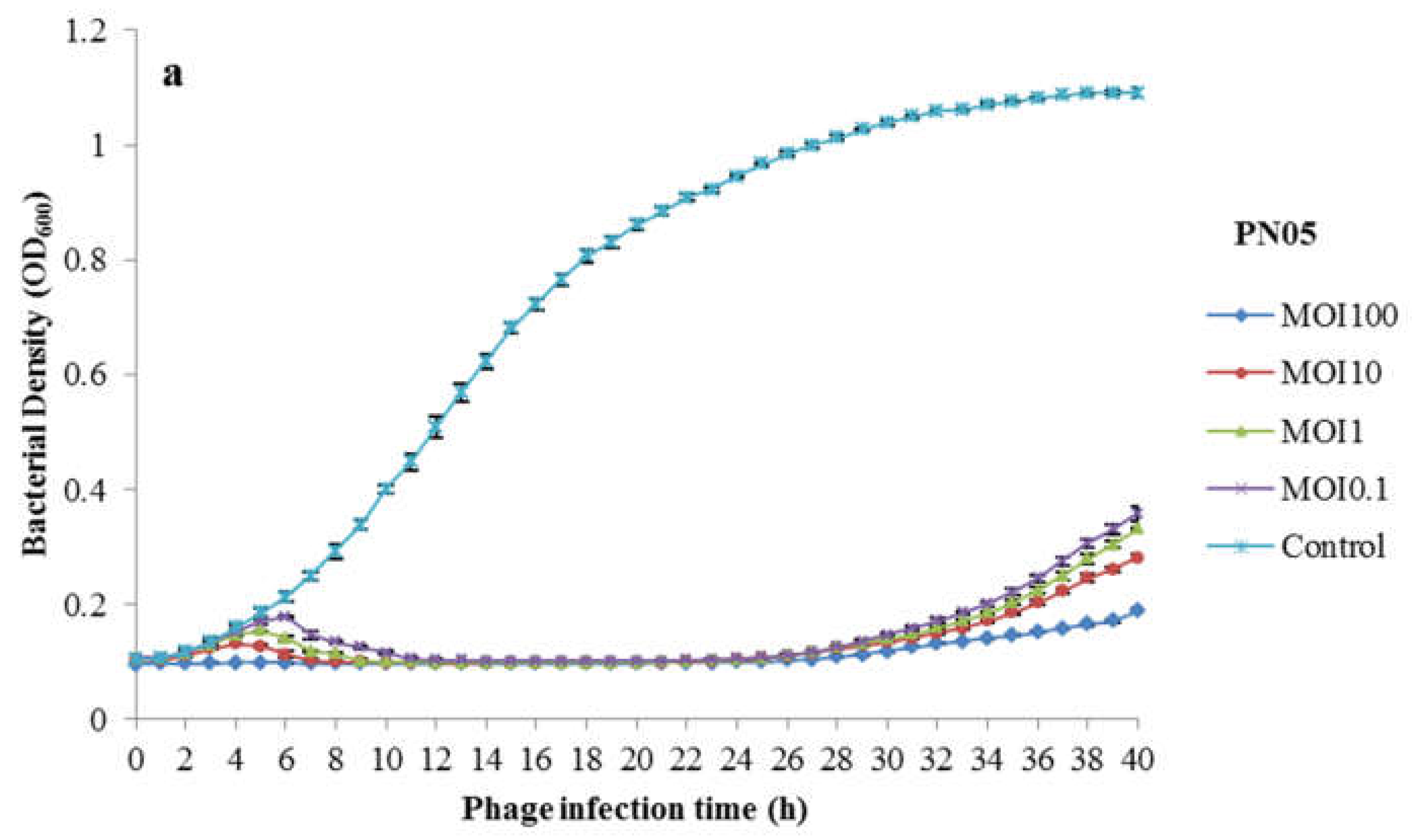
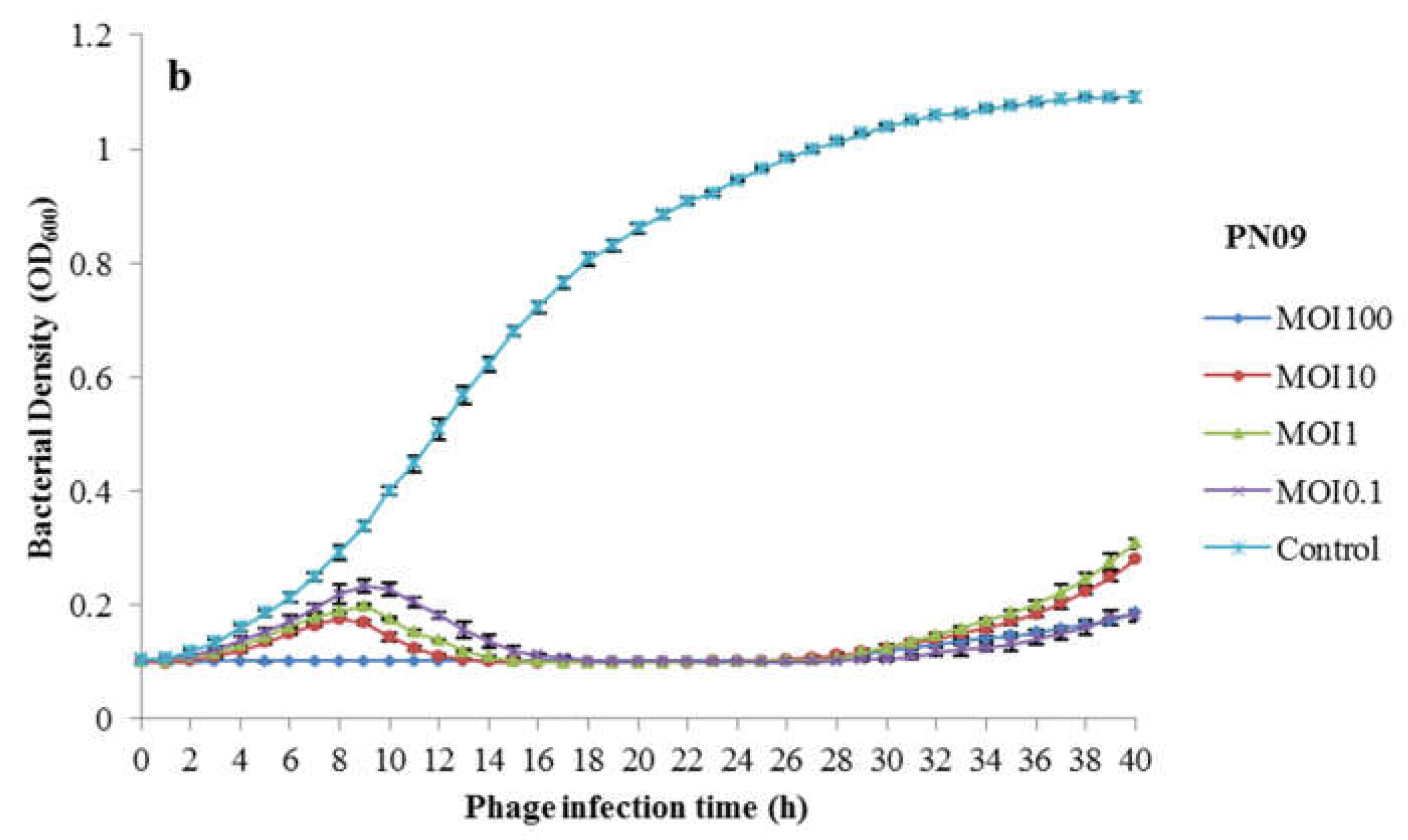
3.2. Phage Bacteriolytic Activity In Vitro
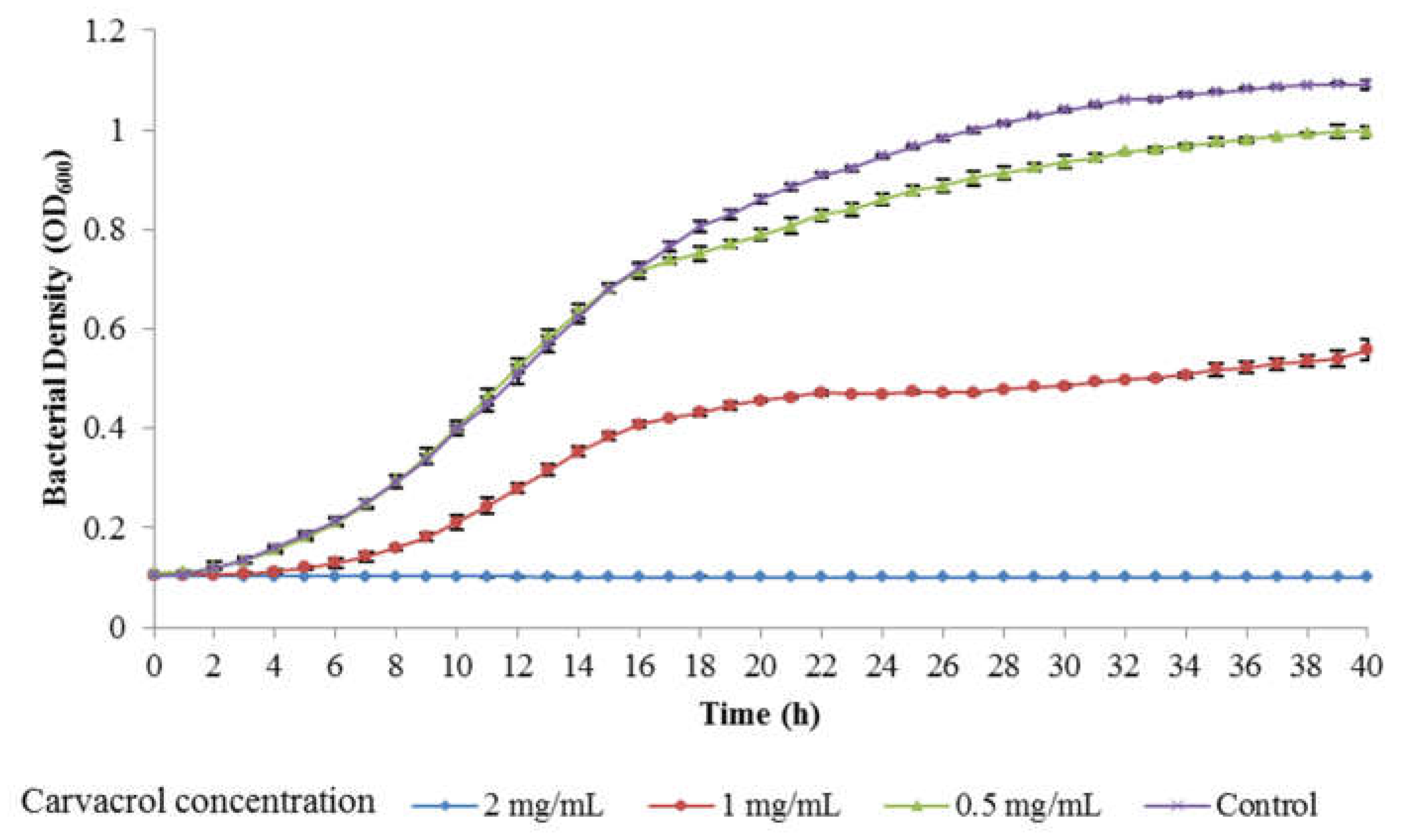
3.3. Antimicrobial Activity of Carvacrol
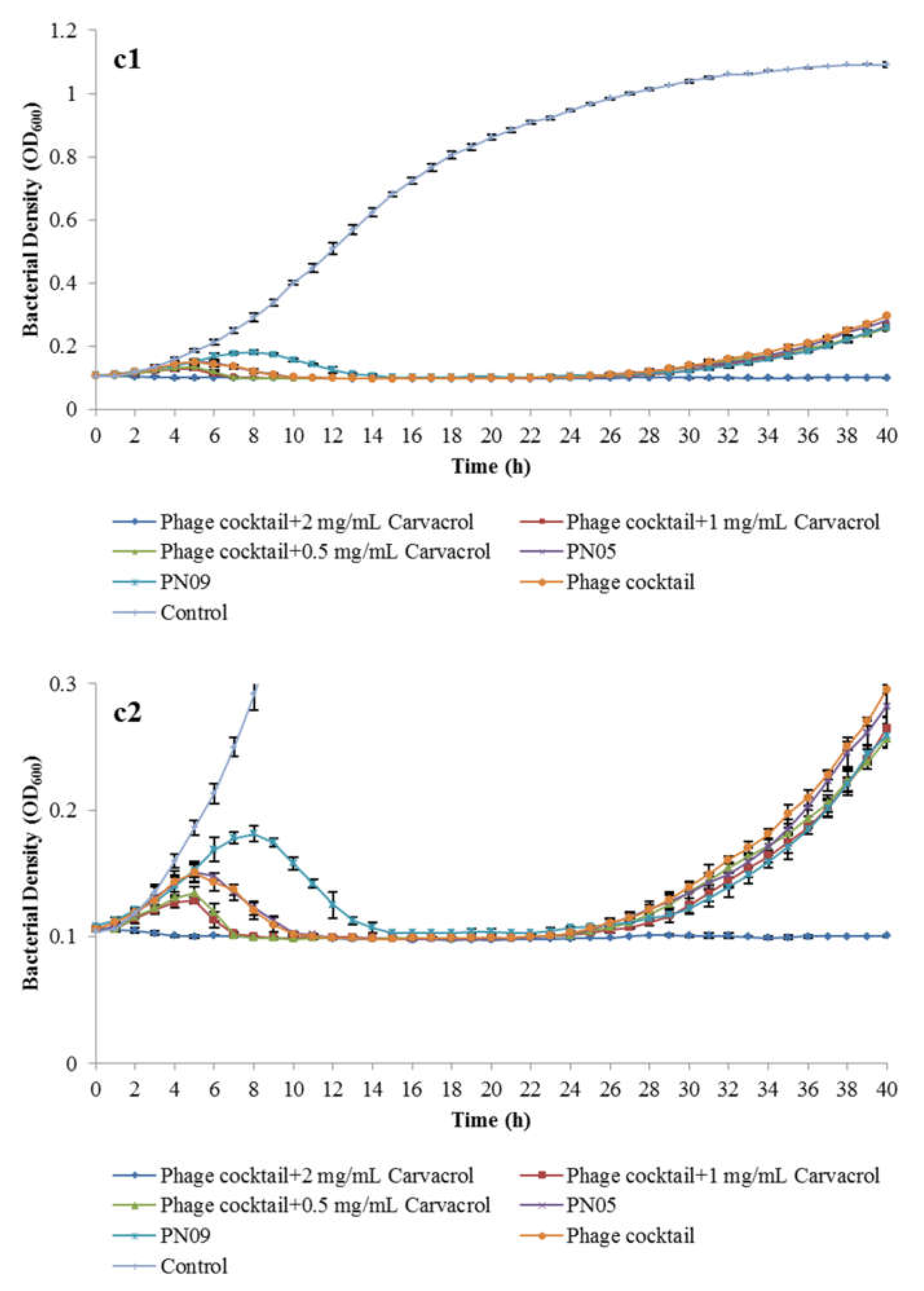
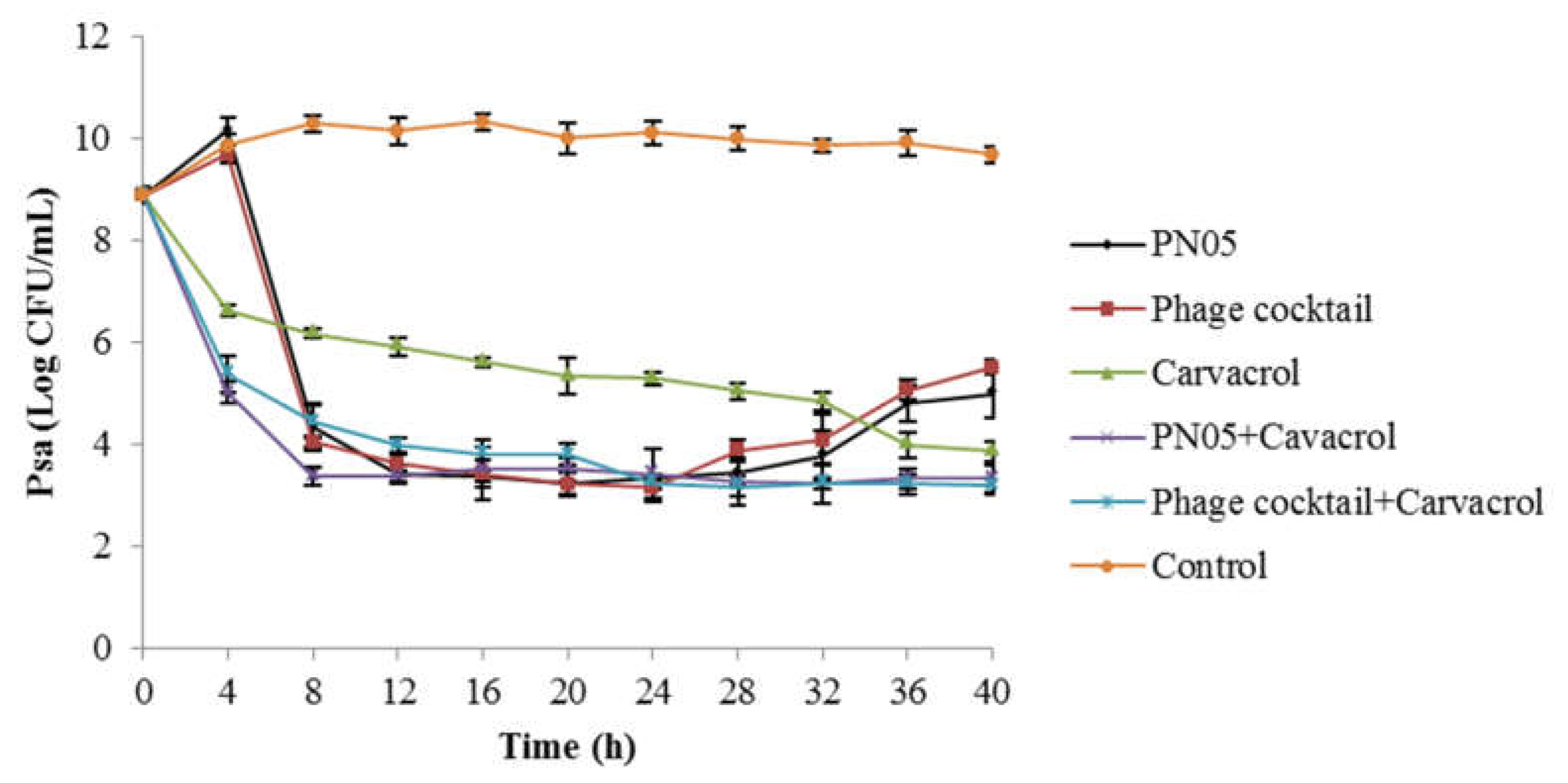
3.4. Efficacy of Combined Phage and Carvacrol against Psa
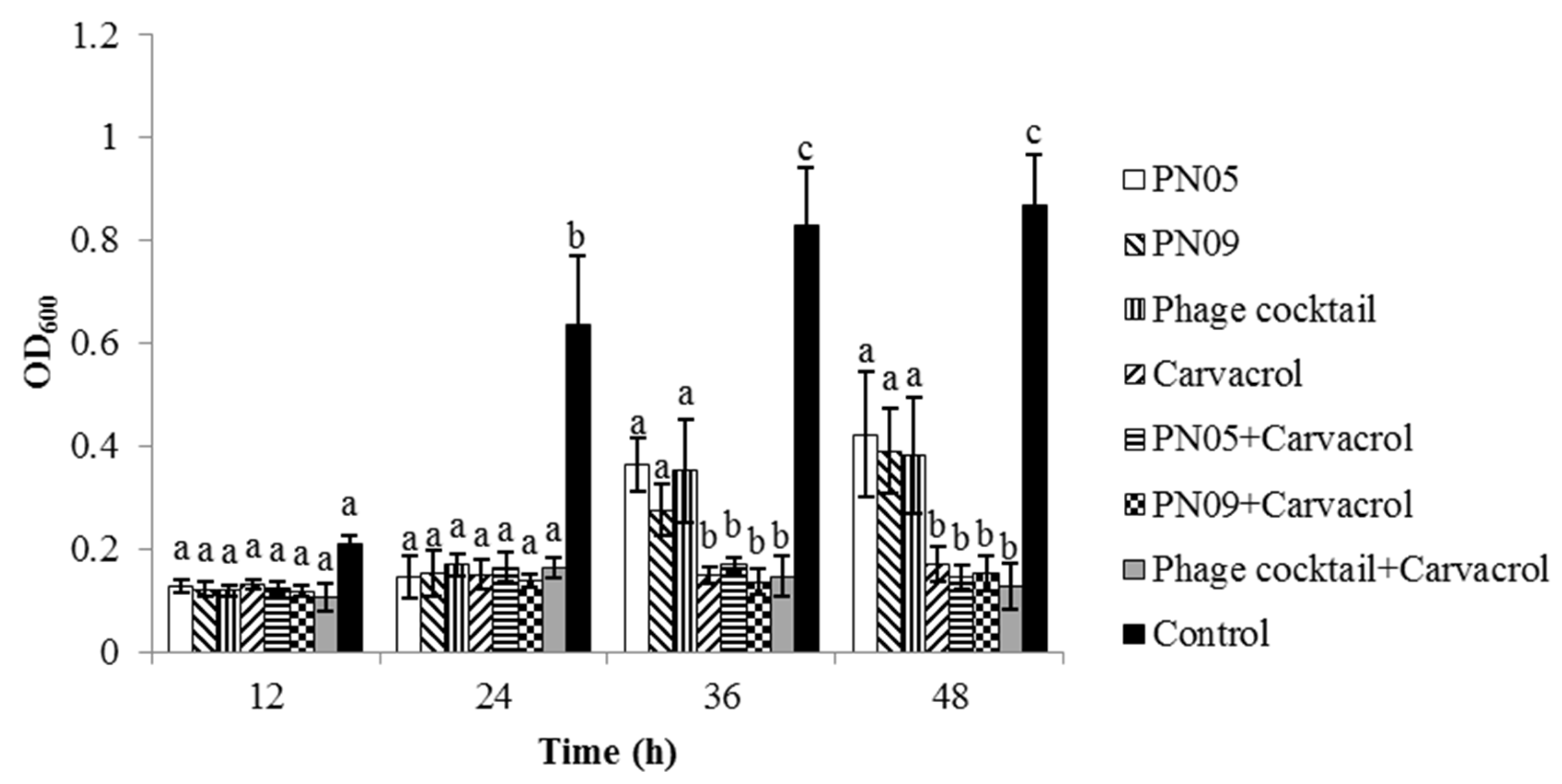
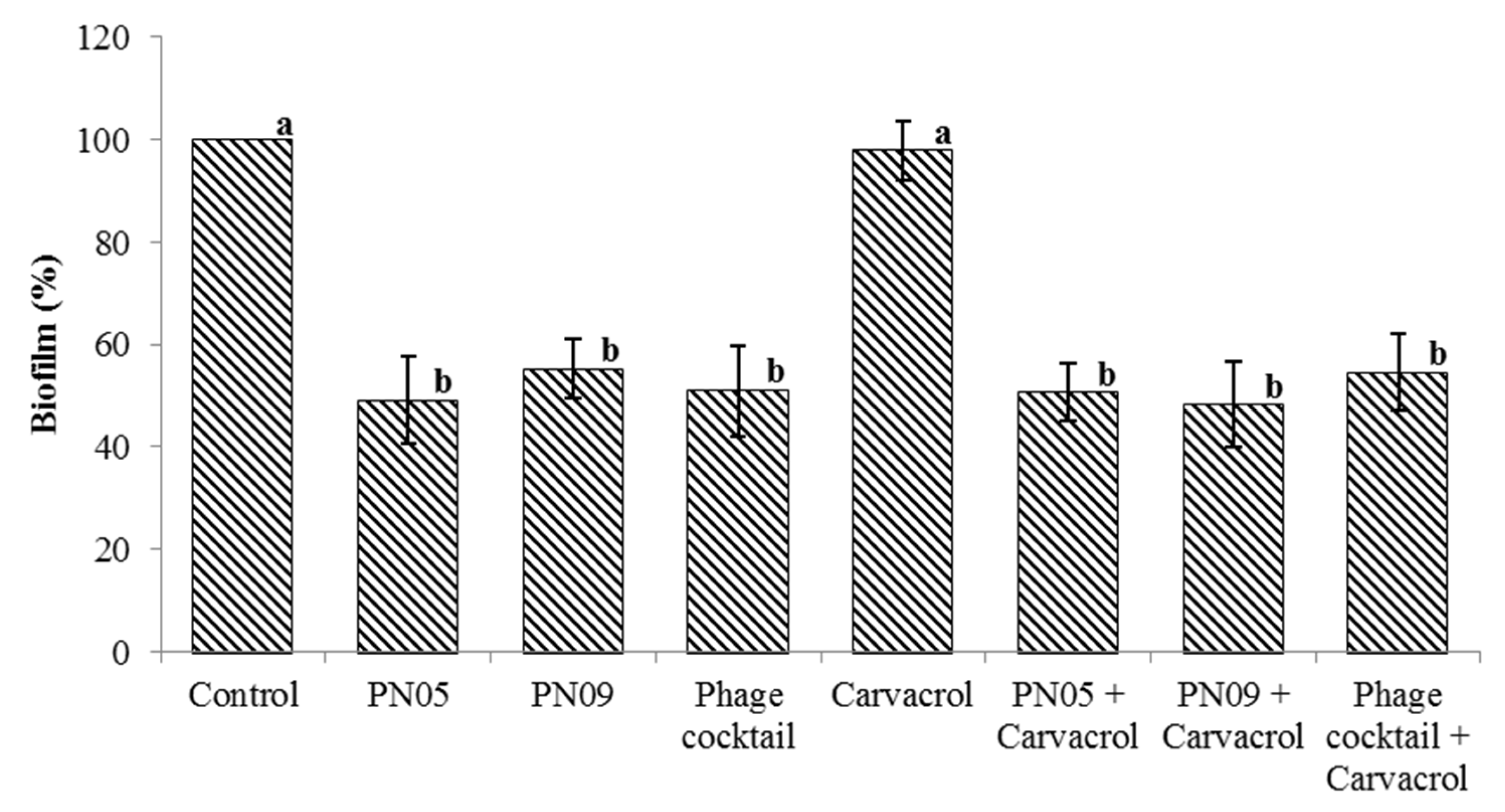
3.5. Biofilm Inhibition
3.6. Pre-Formed Biofilm Destruction
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Donati, I.; Cellini, A.; Buriani, G.; Mauri, S.; Kay, C.; Tacconi, G.; Spinelli, F. Pathways of flower infection and pollen-mediated dispersion of Pseudomonas syringae pv. actinidiae, the causal agent of kiwifruit bacterial canker. Hortic. Res. 2018, 5, 56. [Google Scholar] [PubMed] [Green Version]
- Butler, M.; Stockwell, P.; Black, M.; Day, R.; Lamont, I.; Poulter, R. Pseudomonas syringae pv. actinidiae from recent outbreaks of kiwifruit bacterial canker belong to different clones that originated in China. PLoS ONE 2013, 8, e57464. [Google Scholar]
- Ghods, S.; Sims, I.; Moradali, M.; Rehm, B. Bactericidal compounds controlling growth of the plant pathogen Pseudomonas syringae pv. actinidiae, which forms biofilms composed of a novel exopolysaccharide. Appl. Microbiol. Biotechnol. 2015, 81, 4026–4036. [Google Scholar]
- Altimira, F.; Yanez, C.; Bravo, G.; Gonzalez, M.; Rojas, L.; Seeger, M. Characterization of copper-resistant bacteria and bacterial communities from copper-polluted agricultural soils of central Chile. BMC Microbiol. 2012, 12, 193. [Google Scholar] [CrossRef] [Green Version]
- Bao, H.; Zhang, P.; Zhang, H.; Zhou, Y.; Zhang, L.; Wang, R. Bio-control of Salmonella enteritidis in foods using bacteriophages. Viruses 2015, 7, 4836–4853. [Google Scholar] [CrossRef]
- Yin, Y.; Ni, P.; Deng, B.; Wang, S.; Xu, W.; Wang, D. Isolation and characterisation of phages against Pseudomonas syringae pv. Actinidiae. Acta Agric. Scand. Sect. B Soil Plant Sci. 2019, 69, 199–208. [Google Scholar] [CrossRef]
- Frampton, R.; Taylor, C.; Holguín Moreno, A.; Visnovsky, S.; Petty, N.; Pitman, A.; Fineran, P. Identification of bacteriophages for biocontrol of the kiwifruit canker phytopathogen Pseudomonas syringae pv. actinidiae. Appl. Environ. Microbiol. 2014, 80, 2216–2228. [Google Scholar] [CrossRef] [Green Version]
- Di Lallo, G.; Evangelisti, M.; Mancuso, F.; Ferrante, P.; Marcelletti, S.; Tinari, A.; Superti, F.; Migliore, L.; D’Addabbo, P.; Frezza, D.; et al. Isolation and partial characterization of bacteriophages infecting Pseudomonas syringae pv. actinidiae, causal agent of kiwifruit bacterial canker. J. Basic Microbiol. 2014, 54, 1210–1221. [Google Scholar]
- Pinheiro, L.; Pereira, C.; Frazão, C.; Balcão, V.; Almeida, A. Efficiency of phage φ6 for biocontrol of Pseudomonas syringae pv. syringae: An in vitro preliminary study. Microorganisms 2019, 7, 286. [Google Scholar]
- Pinheiro, L.; Pereira, C.; Barreal, M.; Gallego, P.; Balcão, V.; Almeida, A. Use of phage ϕ6 to inactivate Pseudomonas syringae pv. actinidiae in kiwifruit plants: In vitro and ex vivo experiments. Appl. Microbiol. Biotechnol. 2020, 104, 1319–1330. [Google Scholar]
- Mateus, C.; Costa, L.; Pereira, C.; Silva, Y.; Cunha, A.; Almeida, A. Efficiency of phage cocktails in the inactivation of Vibrio in aquaculture. Aquaculture 2014, 424, 167–173. [Google Scholar] [CrossRef]
- Mateus, C.; Costa, L.; Pereira, C.; Silva, Y.; Almeida, A. Effect of lysozyme addition on the activity of phages against Vibrio parahaemolyticus. Aquaculture 2014, 432, 125–129. [Google Scholar] [CrossRef]
- Lopes, A.; Pereira, C.; Almeida, A. Sequential combined effect of phages and antibiotics on the inactivation of Escherichia coli. Microorganisms 2018, 6, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, P.; Pereira, C.; Gomes, G.; Almeida, A. Efficiency of single phage suspensions and phage cocktail in the inactivation of Escherichia coli and Salmonella Typhimurium: An in vitro preliminary study. Micoorganisms 2019, 7, 94. [Google Scholar] [CrossRef] [Green Version]
- Nazzaro, F.; Fratianni, F.; Martino, L.; Coppola, R.; Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Zhang, D.; Gan, R.; Ge, Y.; Yang, Q.; Ge, J.; Li, H.; Corke, H. Research progress on the antibacterial mechanisms of carvacrol: A mini review. Bioact. Compd. Health Dis. 2018, 1, 71–81. [Google Scholar] [CrossRef]
- Stratakos, A.; Sima, F.; Ward, P.; Linton, M.; Kelly, C.; Pinkerton, L.; Stef, L.; Pet, I.; Corcionivoschi, N. The in vitro, effect of carvacrol, a food additive, on the pathogenicity of O157 and non-O157 Shiga-toxin producing Escherichia coli. Food Control 2017, 84, 290–296. [Google Scholar] [CrossRef]
- Engel, J.; Heckler, C.; Tondo, E.; Daroit, D.; Malheiros, P. Antimicrobial activity of free and liposome-encapsulated thymol and carvacrol against Salmonella, and Staphylococcus aureus, adhered to stainless steel. Int. J. Food Microbiol. 2017, 252, 18–23. [Google Scholar] [CrossRef]
- Chung, D.; Cho, T.; Rhee, M. Citrus fruit extracts with carvacrol and thymol eliminated 7-log acid-adapted Escherichia coli O157:H7, Salmonella typhimurium, and Listeria monocytogenes: A potential of effective natural antibacterial agents. Food Res. Int. 2018, 107, 578. [Google Scholar] [CrossRef]
- Wang, L.; Wang, M.; Zeng, X.; Zhang, Z.; Gong, D.; Huang, Y. Membrane destruction and DNA binding of Staphylococcus aureus cells induced by carvacrol and its combined effect with a pulsed electric field. J. Agric. Food Chem. 2016, 64, 6355–6363. [Google Scholar] [CrossRef]
- Suntres, Z.; Coccimiglio, J.; Alipour, M. The bioactivity and toxicological actions of carvacrol. Crit. Rev. Food Sci. Nutr. 2015, 55, 304. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Bahuguna, A.; Kumar, P.; Bajpai, V.; Kang, S. Antimicrobial potential of carvacrol against uropathogenic Escherichia coli via membrane disruption, depolarization, and reactive oxygen species generation. Front. Microbiol. 2017, 8, 2421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miladi, H.; Zmantar, T.; Chaabouni, Y.; Fedhila, K.; Bakhrouf, A.; Mandouani, K.; Chaieb, K. Antibacterial and efflux pump inhibitors of thymol and carvacrol against food-borne pathogens. Microb. Pathog. 2016, 99, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Burt, S.; Ojofakunle, V.; Woertman, J.; Veldhuizen, E. The natural antimicrobial carvacrol inhibits quorum sensing in Chromobacterium violaceum and reduces bacterial biofilm formation at sub-lethal concentrations. PLoS ONE 2014, 9, e93414. [Google Scholar] [CrossRef] [Green Version]
- Tapia-Rodriguez, M.; Hernandez-Mendoza, A.; Gonzalez-Aguilar, G.; Martinez-Tellez, M.; Martins, C.; Ayala-Zavala, J. Carvacrol as potential quorum sensing, inhibitor of Pseudomonas aeruginosa, and biofilm production on stainless steel surfaces. Food Control 2017, 75, 255–261. [Google Scholar] [CrossRef]
- Moon, S.; Waite-Cusicb, J.; Huang, E. Control of Salmonella in chicken meat using a combination of a commercial bacteriophage and plant-based essential oils. Food Control 2020, 110, 106984. [Google Scholar] [CrossRef]
- Chang, Y.; Yoon, H.; Kang, D.; Chang, P.; Ryu, S. Endolysin LysSA97 is synergistic with carvacrol in controlling Staphylococcus aureus in foods. Int. J. Food Microbiol. 2017, 244, 19–26. [Google Scholar] [CrossRef]
- CLSI, W. Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. Approv. Stand. M7-A7. 2006. Available online: https://infostore.saiglobal.com/en-us/Standards/CLSI-M7-A7-7ED-2006-357843_SAIG_CLSI_CLSI_815015/ (accessed on 1 May 2020).
- Endersen, L.; Buttimer, C.; Nevin, E.; Coffey, A.; Neve, H.; Oliveira, H.; Lavigne, R.; O’Mahony, J. Investigating the biocontrol and anti-biofilm potential of a three phage cocktail against Cronobacter sakazakii in different brands of infant formula. Int. J. Food Microbiol. 2017, 253, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Svircev, A.; Roach, D.; Castle, A. Framing the Future with bacteriophages in agriculture. Viruses 2018, 10, 218. [Google Scholar] [CrossRef] [Green Version]
- Stalin, N.; Srinivasan, P. Characterization of Vibrio parahaemolyticus and its specific phage from shrimp pond in Palk Strait, South East coast of India. Biologicals 2016, 44, 526–533. [Google Scholar] [CrossRef]
- Yin, Y.; Ni, P.; Liu, D.; Yang, S.; Almeidab, A.; Guo, Q.; Zhang, Z.; Deng, L.; Wang, D. Bacteriophage potential against Vibrio parahaemolyticus biofilms. Food Control 2019, 98, 156–163. [Google Scholar] [CrossRef]
- Bae, Y.; Wu, J.; Lee, H.; Jo, E.; Murugaiyan, S.; Chung, E.; Lee, S. Biocontrol potential of a lytic bacteriophage PE204 against bacterial wilt of tomato. J. Microbiol. Biotechnol. 2012, 22, 1613–1620. [Google Scholar] [CrossRef]
- Czajkowski, R.; Ozymko, Z.; Lojkowska, E. Isolation and characterization of novel soilborne lytic bacteriophages infecting Dickeya spp. biovar 3 (‘D. solani’). Plant Pathol. 2014, 63, 758–772. [Google Scholar] [CrossRef]
- Kortright, K.; Chan, B.; Koff, J.; Turner, P. Phage Therapy: A renewed approach to combat antibiotic-resistant bacteria. Cell Host Microbe 2019, 25, 219–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasharawipas, T.; Manopvisetcharean, J.; Flegel, T. Phage treatment of Vibrio harveyi: A general concept of protection against bacterial infection. Res. J. Microbiol. 2011, 6, 560–567. [Google Scholar] [CrossRef] [Green Version]
- Prasad, Y.; Kumar, D.; Sharma, A. Lytic bacteriophages specific to Flavobacterium columnare rescue catfish, Clarias batrachus (Linn.) from columnaris disease. J. Environ. Biol. 2011, 32, 161–168. [Google Scholar]
- Lima, R.; Del Fiol, F.; Balcão, V. Prospects for the use of new technologies to combat multidrug-resistant bacteria. Front. Pharmacol. 2019, 10, 692. [Google Scholar] [CrossRef]
- Hooton, S.; Atterbury, R.; Connerton, I. Application of a bacteriophage cocktail to reduce Salmonella Typhimurium U288 contamination on pig skin. Int. J. Food Microbiol. 2011, 151, 157–163. [Google Scholar] [CrossRef]
- Kotan, R.; Cakir, A.; Dadasoglu, F.; Aydin, T.; Cakmakci, R.; Ozer, H.; Kordali, S.; Mete, E.; Dikbas, N. Antibacterial activities of essential oils and extracts of Turkish Achillea, Satureja and Thymus species against plant pathogenic bacteria. J. Sci. Food Agric. 2010, 15, 145–160. [Google Scholar] [CrossRef]
- Muhsin, J.; Saadia, A.; Fazal, J.; Muhammad, I.; Asif, N.; Tahir, H.; Ali, M.; Rahman, U.; Das, R. Isolation, characterization and efficacy of phage MJ2 against biofilm forming multi-drug resistant Enterobacter cloacae. Folia Microbiol. 2019, 64, 11. [Google Scholar]
- Gu, Y.; Xu, Y.; Xu, J.; Yu, X.; Huang, X.; Liu, G.; Liu, X. Identification of novel bacteriophage vB_EcoP-EG1 with lytic activity against planktonic and biofilm forms of uropathogenic Escherichia coli. Appl. Microbiol. Biotechnol. 2019, 103, 315–326. [Google Scholar] [CrossRef] [PubMed]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ni, P.; Wang, L.; Deng, B.; Jiu, S.; Ma, C.; Zhang, C.; Almeida, A.; Wang, D.; Xu, W.; Wang, S. Combined Application of Bacteriophages and Carvacrol in the Control of Pseudomonas syringae pv. actinidiae Planktonic and Biofilm Forms. Microorganisms 2020, 8, 837. https://doi.org/10.3390/microorganisms8060837
Ni P, Wang L, Deng B, Jiu S, Ma C, Zhang C, Almeida A, Wang D, Xu W, Wang S. Combined Application of Bacteriophages and Carvacrol in the Control of Pseudomonas syringae pv. actinidiae Planktonic and Biofilm Forms. Microorganisms. 2020; 8(6):837. https://doi.org/10.3390/microorganisms8060837
Chicago/Turabian StyleNi, Peien, Lei Wang, Bohan Deng, Songtao Jiu, Chao Ma, Caixi Zhang, Adelaide Almeida, Dapeng Wang, Wenping Xu, and Shiping Wang. 2020. "Combined Application of Bacteriophages and Carvacrol in the Control of Pseudomonas syringae pv. actinidiae Planktonic and Biofilm Forms" Microorganisms 8, no. 6: 837. https://doi.org/10.3390/microorganisms8060837