
Abstract
Optimizing nitrogen (N) management is an important factor for sustainable perennial biomass systems. However, N application is costly, both financially and environmentally. Our objectives were to determine: (1) N rate and plant spacing effects on yield and yield components of prairie cordgrass swards and (2) fertilizer N replacement value (FNRV) of kura clover in prairie cordgrass-kura clover binary mixtures. Plots were established in Illinois, Minnesota, South Dakota, and Wisconsin, USA, in 2010. Kura clover was transplanted on 30-cm centers in all treatments in which it was a component; prairie cordgrass seedlings were transplanted within the kura clover on 60- and 90-cm centers. Monoculture prairie cordgrass stands were established at the same population densities of mixed stands and fertilized with 0, 75, 150, or 225 kg N ha-1. Biomass was harvested in the autumn from 2011 to 2013. N (urea), year, plant spacing, and year × plant spacing affected prairie cordgrass production at all locations. Prairie cordgrass yield increased with N application, but the response varied by location. N application tended to increase prairie cordgrass tiller density and consistently increased tiller mass. Prairie cordgrass yield with 0 N was equal to or less than the yield of prairie cordgrass/kura clover mixtures at all locations in 2011 and 2012; however, kura clover provided a FNRV of 25–82 kg N ha-1 to prairie cordgrass in 2013. Kura clover has potential to provide N to prairie cordgrass in binary mixtures of these two species and on land that may not be easily farmed due to wetness.
Similar content being viewed by others

Introduction
Increasing concern about depletion of non-renewable fossil fuels, and climate change driven by gas emissions has resulted in a high level of interest in renewable and sustainable energy resources such as bioethanol [1,2,3]. Bioethanol production in the USA has been primarily starch-based using corn (Zea mays) as a primary raw material [4,5,6].
Lignocellulosic biomass materials, such as agricultural residues remaining after grain harvest and dedicated energy crops (e.g., switchgrass (Panicum virgartum L.) and prairie cordgrass (Spartina pectinata Link.)), are abundant organic resources for biofuel production. Perennial energy crops have unique advantages in that they have greater environmental sustainability and are renewable [7, 8]. Prairie cordgrass has been shown to have high potential as a biomass crop in the northern Great Plains [9,10,11].
Prairie cordgrass is a perennial warm-season grass that spreads both vegetatively through rhizomes and non-vegetatively through seed. It is native to the North American Prairie and is found in most of the USA except for California, Nevada, and Arizona in the southwest and Louisiana to South Carolina in the southeast [12]. Prairie cordgrass is valued for stabilizing soil, preventing erosion on spillways and drainage channels, and revegetating wetlands. It also provides good cover for wildlife because of its thick stands, and can be used for hay for livestock if harvested when relatively immature [12]. Prairie cordgrass is a facultative wetland species, predominantly found in low floodplains and wetlands [13] where it occupies soils too wet and inadequately aerated for other species such as big bluestem (Andropogon gerardii Vitman) and switchgrass [12]. Studies have documented its potential for biomass production in short-season areas in Europe [14], southwestern Quebec [15], eastern South Dakota [9], and Kansas [11]. On agricultural soils in Minnesota, prairie cordgrass had consistently greater long-term yields than miscanthus (Miscanthus × giganteus), switchgrass, or big bluestem [16]. Prairie cordgrass has also been shown to have high tolerance for soil salinity in Canada [17] and the USA [18].
Proper N management is crucial in achieving economically viable biomass yields, reducing environmental contamination from N leaching, and maintaining stands of perennial biofuel crops [19]. For example, optimum biomass yield of switchgrass was reached with 140 kg N ha-1 in Iowa [20], but biomass yield response to N application varied with initial soil N concentration among several locations in the USA [19], and N application was not always economically viable at these same sites [21]. In Illinois, Guo et al. [22] found a yield response of prairie cordgrass to a N rate up to 84 kg N ha-1, but no additional response occurred above this rate. Otherwise, there is little information regarding biomass yield response of prairie cordgrass to N fertilization, especially across multiple diverse environments.
For successful biomass production, one of the critical phases is initial plant density. Prairie cordgrass can be propagated by seed or vegetatively by rhizomatous growth [10]. Due to significant rhizome production, it is possible that prairie cordgrass would not need high initial plant density for biomass production. This is economically advantageous considering the generally high cost of prairie cordgrass seed. For Miscanthus × giganteus, Lewanowski et al. [23] noted a slightly greater yield in the first 2–5 years when rhizomes were planted at higher densities (> 2 m-2), but this yield increase did not compensate for the increased establishment costs associated with the higher initial plant densities. Also, high initial plant density has been shown to increase tiller mortality because of competition for nutrients [24, 25]. Therefore, information on optimal planting density for any species, including prairie cordgrass, has both economic and agronomic ramifications [26].
Kura clover (Trifolium ambiguum M. Bieb) is a rhizomatous perennial legume that is a potential candidate for inclusion in mixtures with perennial grasses grown as bioenergy crops. It can withstand seasonal flooding; thus, it may grow well in areas where prairie cordgrass is also adapted. It has persisted for over 20 years in long-term grazing trials in Wisconsin and Minnesota [27]. It tolerates frequent defoliation in monoculture or in a mixture with perennial grass under continuous grazing [28, 29].
As an alternative to synthetic N fertilizer, biologically fixed N has economic and environmental advantages [30, 31]. When grown in mixture with grasses, N fixed by the legume can be utilized by grasses through direct transfer via mycorrhizal hyphae from legume to grass, degradation and decay of dead legume tissues, or nitrogen exudation by legume roots [32, 33]. Several studies have documented improvements in both forage yield and quality in grass-legume mixtures compared with monoculture grass swards [34,35,36]. Zemenchik et al. [36] estimated the fertilizer nitrogen replacement value (FNRV) of kura clover grown with orchardgrass (Dactylis glomerata L.) or smooth bromegrass (Bromus inermis L.) to range from 74 to 325 kg N ha-1. Barnett and Posler [34] also found yield advantages when various legumes were included in binary mixtures with diverse perennial grasses. However, there is no information regarding the use of kura clover, particularly as a source of N, with prairie cordgrass for sustainable biomass production.
Our hypotheses were that N fertilizer would increase biomass production in monoculture prairie cordgrass stands and that kura clover would provide a N benefit when grown in mixture with prairie cordgrass. Based on the rhizomatous growth of prairie cordgrass, we also hypothesized that initial plant density would not have long-term effects on biomass production. Therefore, the objectives of this study were to determine (1) N rate and plant spacing effects on yield and yield components of monoculture prairie cordgrass swards and (2) to determine the fertilizer N replacement value of kura clover in prairie cordgrass/kura clover binary mixtures.
Materials and Methods
Location Description
This study was conducted from 2010 to 2013 at four locations in the USA: Arlington, WI; Rosemount, MN; Brookings, SD; and Urbana, IL. Specific information (GPS coordinates and soil type) for each location is described in Table 1. Plots were established on land at each location that was either poorly drained or commonly flooded for some period during spring.
Field Management
Prairie cordgrass was grown in monoculture or in a binary mixture with kura clover. Prairie cordgrass germplasm was from a native population collected in South Dakota and the kura clover population was developed by AgResearch New Zealand. Individual seedlings of both species were grown in conetainers (Stuewe, Inc., Corvallis, OR) during late winter-early spring 2010 in the greenhouse and then transplanted to the field in late spring 2010. Kura clover was inoculated with R. leguminosarum biovar trifolii strains 162C11, 162C13, and 162C14 mixture from Liphatech (Liphatech, INC. Milwaukee, WI) in the greenhouse. Kura clover was transplanted on 30-cm centers (30 cm between and within rows for a total density of 111,111 kura clover plants ha-1) in the field in the prairie cordgrass-kura clover mixture treatments. Prairie cordgrass seedlings were transplanted within the kura clover on 60- or 90-cm centers (populations of 26,896 and 11,881 plants ha-1, respectively) to obtain initial prairie cordgrass stand percentages of approximately 25 and 10%, respectively. Monoculture prairie cordgrass stands were also established at the same population densities as mixed stands. Four levels of N fertilizer (0, 75, 150 and 225 kg N ha-1) were applied as urea to the prairie cordgrass monocultures once annually in May from 2011 to 2013 at all locations. Individual plots were 3.0 m wide and 5.7 m long. This size allowed us to overcome potential border effects since a strip (approximately 1 m wide) was harvested through the center of the length of each plot for prairie cordgrass yield determination. A broadleaf herbicide, 2, 4-D (2, 4-dichlorophenoxy acetic acid), was applied at a rate of 0.6 kg a.i. ha-1 to suppress the kura clover in WI and MN in 2011, 2012, and 2013 and in SD and IL in 2012 and 2013. Herbicide was sprayed across the entire experimental area, including monoculture prairie cordgrass plots, to maintain consistency of treatments.
Biomass Yield
Prairie cordgrass yield (Mg ha-1) was determined at all locations from 2011 to 2013. A strip (120 cm wide for 60-cm row spacing treatment or 180 cm wide for 90-cm row spacing treatment) from the middle of each plot was harvested with a sickle-bar mower at a stubble height of about 10 cm once annually in late autumn after a killing frost and significant senescence had occurred. Little to no kura clover was present in the harvested biomass because most kura clover had fallen below the cutting height after a killing frost. For this reason, reported yields include only prairie cordgrass. Fresh samples were harvested and weighed in the field and then dried in a forced air oven at 60 °C for 72 h and reweighed for dry matter determination.
Prairie Cordgrass Yield Components
Prairie cordgrass tillers were collected from each plot before harvest for yield component analysis (i.e., tiller density and tiller mass). All prairie cordgrass tillers were hand-harvested to a stubble height of about 10 cm using a rice knife from two 0.36 m2 quadrats in the 60-cm row spacing treatment and two 0.81 m2 quadrats in 90-cm row spacing treatment. The different quadrat sizes allowed sampling from the entire width of the middle row of each plot. Collected tillers were dried at 60 °C for 72 h in a forced air oven and then the number of tillers was counted and weighed to determine tiller density and tiller mass.
Nitrogen Use Efficiency
Nitrogen use efficiency (NUE) represents yield per unit of applied N and was calculated following Zemenchik and Albrecht [37]:
Fertilizer N Replacement Value of Kura Clover
Second-order linear regressions were fitted for prairie cordgrass yield on fertilizer N rate in 2013 at each location before treatment means were calculated [36]. The regression procedure of Statistix 9 (Analytical Software, Tallahassee, FL) was used for regression analysis. Mean yield of prairie cordgrass in the binary mixture treatments for 2013 were substituted for Y in the regression model and then solved for X to obtain the FRNV of kura clover.
Statistical Analysis
Analyses of variance were performed to evaluate the effect of N rate, plant spacing, and all interactions on biomass production (balanced data) and yield components (unbalanced data for tiller mass and tiller density) of the prairie cordgrass monocultures using the linear models procedure in Statistix 9 (Analytical Software, Tallahassee, FL). Fisher’s least significant difference (LSD) was used to separate treatment means at P < 0.05. Nitrogen rate, plant spacing, and year were considered to be fixed effects. Locations were analyzed separately due to significant location × treatment interactions.
Results
Weather Conditions
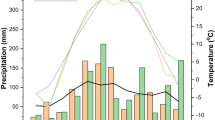
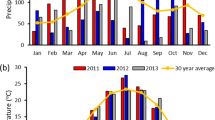
Weather conditions during the growing season varied by year and location (Tables 2 and 3). Generally, precipitation was below long-term averages from 2011 to 2013 at all locations except for WI in 2013. In 2012, a widespread drought affected much of the central US, and precipitation from May to July of 2012 was often significantly below the 30-year average at the four locations. Particularly dry months in 2012 included June (6.0 mm) in WI, July (31.0 mm) in SD, June and July (33.0 and 17.8 mm, respectively) in MN, and July (14.2 mm) in IL. Average monthly temperatures approached the 30-year mean in most years with the notable exception of higher temperatures in July 2012 in SD and May and July 2012 in IL.
Biomass Yield
Averaged across years, N rate, and plant spacing, prairie cordgrass yields varied from 6.4 Mg ha-1 in IL to 16.6 Mg ha-1 in MN (Table 4). Differences in prairie cordgrass yield occurred among years and N rates at all locations (Table 5, Fig. 1), and there was a N × year interaction in SD and IL and a plant spacing × year interaction at all locations. Biomass yields were 17.8, 13.2, 11.3, and 6.0 Mg ha-1 in WI, MN, SD, and IL, respectively in 2011. Yields of prairie cordgrass were lowest at all locations in 2012 due to abnormally low precipitation, but then increased in 2013 after the drought of 2012 (Fig. 1).

The effect of year on biomass yield of prairie cordgrass at four locations in the USA (Wisconsin, WI; Minnesota, MN; South Dakota, SD; and Illinois, IL). Data are averaged across plant spacing and N rate. Values within a location followed by different letters are significantly different at 0.05 level of probability
A significant year × plant spacing interaction was observed for biomass yield of prairie cordgrass at each location (Table 5, Fig. 2). Biomass yield was higher for the 60-cm plant spacing than the 90-cm plant spacing at all locations in 2011. Prairie cordgrass with 60-cm plant spacing yielded 18.0, 39.3, 13.0, and 39.3% more compared with the 90-cm plant spacing in WI, MN, SD, and IL, respectively in that first year (Fig. 2). In 2012, plant spacing affected yield in SD only, and by 2013 plant spacing had no effect on biomass production.

Prairie cordgrass biomass production as influenced by the plant spacing × year interaction at four locations in the USA (Wisconsin, WI; Minnesota, MN; South Dakota, SD; and Illinois, IL). Data are averaged across N rate. Values within a location followed by different letters are significantly different at 0.05 level of probability
Nitrogen rate consistently affected biomass yield of prairie cordgrass at all locations (Table 5, Fig. 3). Averaged across years and plant spacing, biomass yield was highest when 225 kg N ha-1 was applied in WI and SD, but yields peaked at the 150 kg N ha-1 rate in MN and the 75 kg N ha-1 rate in IL. Compared with the 0 N control, biomass yield increased by 8.5, 11.0, and 14.1% in WI, 3.0, 16.3, and 16.4% in MN, 35.6, 36.6, and 45.8% in SD and 30.1, 36.5, and 33.8% in IL when N was applied at rates of 75, 150, and 225 kg ha-1, respectively.

The effect of N rate on prairie cordgrass yield at four locations in the USA (Wisconsin, WI; Minnesota, MN; South Dakota, SD; and Illinois, IL). Data are averaged across year and plant spacing. Values within a location followed by different letters are significantly different at 0.05 level of probability
Nitrogen Use Efficiency
Averaged across years and plant spacing, NUE decreased in response to increasing N rates (Table 6). NUE was highest at 75 kg N ha-1 and significantly decreased at the 150 and 225 kg N ha-1 rates in SD and IL. Numerically, the highest NUE for WI and MN was also at the 75 kg N ha-1; however, there was no statistical difference among the various N rates at either of these locations (Table 6).
Averaged across N rate and year, there was no significant difference in NUE between 60- and 90-cm prairie cordgrass plant spacings, except for WI (Table 6) where the NUE in 90-cm plant spacing was higher than that in 60-cm plant spacing.
Averaged across N rates and plant spacing, NUE tended to increase over years at all locations (Table 6). The first year after establishment had the lowest NUE and the third year the highest NUE at all locations except for MN and IL which had comparable NUE in 2012 and 2013.
Yield Components
Tiller density of prairie cordgrass was negatively correlated with tiller mass at all locations (Fig. 4). Averaged across years and plant spacing, there were significant N rate effects on tiller density at SD and IL, but not at WI and MN, but there were also various 2-way interactions for these yield components (Table 7). Tiller density ranged from 110 to 134, 158 to 184, 169 to 216, and 131 to 160 tillers m-2 at WI, MN, SD, and IL, respectively (Table 8).

Relationship between tiller density (tiller m-2) and tiller mass (g tiller-1) in prairie cordgrass monocultures at four locations in the USA (Wisconsin, WI; Minnesota, MN; South Dakota, SD; and Illinois, IL). Data are averaged across years, plant spacing, and N rate
N rate affected tiller mass at all locations (Table 7). Tiller mass was greater in N-fertilized prairie cordgrass compared with the unfertilized control (Table 8). Across the 75, 150, and 225 kg N ha-1 rate, average tiller mass was 18 g at WI, 8 g at MN, 7 g at SD and 4 g at IL, while the average tiller mass of unfertilized prairie cordgrass was 14 g at WI, 6 g at MN, 5 g at SD, and 3 g at IL. There was no significant difference in tiller mass among the 75, 150, and 225 kg N ha-1 rates at any location.
Averaged across plant spacing and N rate, there was a significant effect of year on tiller density and tiller mass at all locations except for tiller mass at IL (Table 7). Tiller density and mass were not measured in 2013 in MN and in 2011 in IL. In MN, tiller density and tiller mass decreased significantly from 2011 to 2012. In IL, tiller density increased from 2012 to 2013 while tiller mass did not change (Table 8).
There was a significant plant spacing × year interaction on tiller mass and tiller density (Table 7). Overall, tiller density in 60-cm plant spacing was higher than that in 90-cm plant spacing in 2011 (Fig. 5). However, differences in tiller density between 60- and 90-cm plant spacing decreased over time at all locations except for SD where tiller density differences were noted again in 2013 between the 60- and 90-cm plant spacings. Although not consistent, tiller mass in 60-cm plant spacings tended to be lower than that in 90-cm plant spacing at all locations across the 3 years (Fig. 5).

Tiller density (tiller m-2) and tiller mass (g tiller-1) in prairie cordgrass monocultures as affected by the plant spacing x year interaction at four locations in the USA (Wisconsin, WI; Minnesota, MN; South Dakota, SD; and Illinois, IL). Data are averaged across N rates. Values within a location followed by different letters are significantly different at 0.05 level of probability
Fertilizer Nitrogen Replacement Value of Kura Clover
Averaged across plant spacing, biomass yield of prairie cordgrass monocultures to which 0 kg N ha-1 was applied annually remained relatively similar over the 3 years of this study. In contrast, biomass yield of prairie cordgrass-kura clover mixtures generally increased over time at all locations (Fig. 6). In 2011, biomass yield of prairie cordgrass monocultures with 0 kg N ha-1 was greater than that of prairie cordgrass-kura clover mixtures in WI and IL, while yields for these comparable treatments were similar in SD and MN. In 2012, yield of the prairie cordgrass monoculture control (0 kg N ha-1) was similar to that of the prairie cordgrass-kura clover mixture at MN, SD, and IL, but not WI. In 2013, prairie cordgrass yielded more when mixed with kura clover than the 0 N control at all locations except WI (Fig. 6). The FNRV varied among locations in 2013, but was highest in MN (82 kg N ha-1) and lowest in WI (25 kg N ha-1) (Table 9).

Comparison of biomass yield of prairie cordgrass grown as monocultures with 0 kg N ha-1 (PCG + 0 kg N ha-1) and prairie cordgrass in binary mixture with kura clover (PCG + KC). Data are averaged across plant spacing. Values within a location and year followed by different letters are significantly different at 0.05 level of probability. ns*, not significantly different at 0.05 level of probability (p > 0.05)
Discussion
Prairie cordgrass yield varied temporally at all locations, and differed among locations as well. Lowest overall annual yields occurred at the IL location (5–10 Mg ha-1) and the highest yields were observed at the WI location (14–20 Mg ha-1). Yields at MN and SD were intermediate to these other locations. Based on an evaluation of multiple switchgrass cultivars at several diverse environmental locations in the USA, Casler et al. [38] recommended that switchgrass not be moved more than one USDA hardiness zone north or south of their origin. However, they noted that moving cultivars across a longitudinal gradient may be possible if field tests demonstrate this type of adaptation. We suspect that the prairie cordgrass germplasm derived from SD was not well adapted in IL because of the latitudinal difference between the two locations. In addition, IL has since identified and collected IL-local prairie cordgrass germplasm that has yielded up to 20 Mg ha-1 (DoKyoung Lee, personal communication). Biomass yield in 2012 was significantly lower than in the other 2 years as a result of widespread drought throughout the USA, including the locations in this study. Despite dry conditions in 2012, prairie cordgrass yield increased in 2013 to levels above that seen in 2011, the year after establishment in the field. Perennial grass yield often increases during the first several years after establishment, and the extent to which increases occur is dependent on several factors including weather conditions, environment, and N and harvest management [26]. Thus, although our prairie cordgrass was transplanted, it was not unexpected to have prairie cordgrass yields be lower the first production year, and in the second production year because of dry conditions.
Year × plant spacing interactions affected prairie cordgrass yield at all locations (Table 5). Biomass yield differences in the first production year can likely be attributed primarily to initial plant density. Plots with narrower row spacing (60 cm) initially allowed for greater utilization of space and light based on the fact that we generally observed higher tiller density in these plots. Similarly, previous studies showed that biomass and essential oil yield of rose-scented geranium (Pelargonium species) was 132% and 99% higher for narrower spacing (60 × 30 cm) than that for wider spacing (120 × 30 cm) [39], and dry bean (Phaseolus vulgaris L.) cultivar yields were 52% higher on 250 mm compared to a 750-mm row width [40]. As we hypothesized, however, the effect of plant spacing on prairie cordgrass yield decreased over time, to the extent that no differences were noted in 2013. In addition, by the 2013 season, it was difficult to detect rows, or individual plants, in any of the plots, regardless of initial plant spacing. Prairie cordgrass is strongly rhizomatous [10], and these rhizomes give rise to new tillers that populated the inter-row and inter-plant areas over time. Initially, the wider row spacing had more inter-row and inter-plant area than the narrow plant spacing. Thus, the 90-cm plant spacing allowed for greater yield increases as tiller density increased while the high initial plant density in 60-cm plant spacing may have increased competition among plants for available nutrients. Mahmood and Honermeier [41] reported that row spacing of sorghum (Sorghum bicolor L. Meonch) had no clear effect on yield in the first experimental year, but in the second year narrower spacing yielded higher than wider row spacing. Also, other studies have demonstrated that higher initial plant density led to greater tiller mortality over time as a result of competition for nutrients and light [24, 25]. Because prairie cordgrass is a perennial, rhizomatous species, yield differences between the two plant spacings decreased over time until tiller density was similar for both the 60- and 90-cm plant spacings at three of the four locations (Fig. 5).
In previous studies in SD, the mean yield of different prairie cordgrass populations was 8.3 Mg ha-1 [9] while average yield of “Red River” and “Atkins” prairie cordgrass in Kansas was 9.5 Mg ha-1 [11]. We observed yields lower than, similar to, or higher than these at the four locations used in this study (Fig. 1).
Prairie cordgrass responded positively to N fertilizer at all locations (Fig. 3.). However, a previous 4-year study reported no yield response with N fertilization (N rates up to 168 kg ha-1) on “Red River” and “Atkins” prairie cordgrass in either South Dakota or Kansas [11]. Owens et al. [19] noted variable responses in switchgrass yield as related to N application at five locations across 2 years at diverse locations across the USA. They speculated that initial soil N concentration was at least partially responsible for the variable responses. In a follow-up to that study, that included an additional 5 years of yield data from the same locations, Lee et al. [42] found similar results with the exception that switchgrass at the Iowa site began to respond positively to N application.
Higher biomass production has often been achieved with higher N inputs. However, higher N does not always give higher NUE, and the maximum biomass produced per unit of N applied may be at lower N fertilizer rates [19]. Considering the environmental and economic impacts of N fertilizer application, determining the NUE is important when selecting biomass energy crops. Pedroso et al. [43] reported the maximum NUE was reached at the lowest N fertilizer level with switchgrass, miscanthus, and tall wheatgrass (Agropyron elongatum). Lemus et al. [44] also reported the NUE of switchgrass was highest at the lowest N rate (56 kg N ha-1) in their study. In our study, the lowest N fertilizer rate (75 kg N ha-1) had the highest NUE at all locations (Table 6).
Christian et al. [45] suggested that 1-year-old plants of switchgrass and coastal panic grass (Panicum amarum A.S. Hitchc. & Chase) for biomass production would not require a high level of N because it has relatively low biomass yield. However, more N may be necessary in succeeding years when above- and belowground mass of the crop is further developed allowing for improved nitrogen use efficiency [26]. Similar results were observed in our study when the NUE was higher for prairie cordgrass in the second or third growing season compared to the first year.
Significant N translocation to belowground storage organs occurs in perennial grasses during senescence [46,47,48], and this may contribute to higher N use efficiency. We suggest that prairie cordgrass has high N use efficiency because it is able to recycle nutrients via well-developed rhizomes. Nitrogen translocation from shoot to root biomass during senescence contributes to plant survival through the winter [49, 50], and this stored N can be re-translocated to aboveground tissues during the early growing season to help reduce the use of N fertilizer [43]. This may be particularly relevant when perennial species are harvested after a killing frost, thus maximizing N translocation to roots and rhizomes and minimizing N removal during biomass harvest, as in this study.
In our study, tiller density and tiller mass varied by N application rate, plant spacing, and year (Table 7). These variations may result from different environmental conditions among locations. Smart et al. [51] reported that switchgrass stands with low tiller number and high tiller weight yielded 25% more biomass than stands with high tiller number and low tiller weight. Similarly, in our study, WI had low tiller number, high tiller mass, and thus the highest yield. Tiller number of Miscanthus × giganteus in central Greece [52] and of two different prairie cordgrass varieties in South Dakota and Kansas [11] was not affected by N rate. Similar results in our study were found in WI and MN. However, N fertilization affected tiller density and tiller mass in SD (Table 8). Similarly, mean tiller density of tall fescue increased with N application [53], and there was high variation in tiller density of perennial ryegrass from year to year [54, 55]. Mahmood and Honermeier [41] reported that narrow row spacing caused a significant increase in tiller density compared with wide row spacing. Also, Snider et al. [56] reported that wider row spacing of sorghum produced the highest stem densities, and stem diameter declined as stem density increased at one of their experimental sites, similar to what we observed in this study (Fig. 4). Fraser and Kindscher [57] reported that stem density of transplanted prairie cordgrass plugs significantly changed over 3 years. Stem density in the small plug treatment decreased sharply while stem density in the large plug treatment increased, and at the end of 3 years the stem density between the different treatments appeared to converge.
Yield of prairie cordgrass mixed with kura clover increased annually, and by the third year had out-yielded the non-fertilized prairie cordgrass in the monoculture plots. Similarly, previous studies showed greater yields in pastures and grasslands composed of grass-legume mixtures [58,59,60,61]. As kura clover development increased over time, the transferable N from legume to grass may have also gradually increased or accumulated in the soil through degradation and decomposition of dead legume tissue [32, 62,63,64] or N exudation from the roots [33]. Nesheim and øyen [65] and Boller and Nösberger [62] reported that legumes grown in a mixture with grasses provide a large majority of their N through symbiotic N2 fixation, up to 300 kg N ha-1. Thus, N transfer from prairie cordgrass to kura clover may have boosted the prairie cordgrass yield in mixtures. Zemenchik et al. [36] estimated the FNRV of kura clover to range from 71 to 274 kg N ha-1 when grown with various cool season grasses for forage, and their results varied by location and year. In our study, the FNRV of kura clover ranged from 25 to 82 kg N ha-1 in 2013 (Table 9). This range of values is not surprising considering the variable environments utilized in this study. Regardless of the range of values, however, it demonstrates the potential benefit of the binary mixture of prairie cordgrass-kura clover.
Conclusions
Biomass of prairie cordgrass varied by location, ranging from 6.4 to 16.6 Mg ha−1, and yield was affected by N rate, year, and plant spacing. We also observed a significant influence from the interaction between year and plant spacing. Monoculture prairie cordgrass yield increased with N application; however, it had the greatest nitrogen use efficiency at the 75 kg N ha-1 rate and in later years when prairie cordgrass was well established. Prairie cordgrass tiller density tended to increase in response to increased N. Yield of prairie cordgrass with 0 N was equal to or less than the yield of prairie cordgrass grown with kura clover at all locations in 2011 and 2012; however, there was a positive N value associated with prairie cordgrass grown with kura clover in 2013. Kura clover has potential to provide N to prairie cordgrass in binary mixtures of these two species and on land that may not be easily farmed due to poor drainage or excessive wet periods.
References
Hill J, Nelson E, Tilman D, Polasky S, Tiffany D (2006) Environmental, economic and energetic costs and benefits of biodiesel and ethanol biofuels. PNAS 103:11206–11210
Ragauskas AJ, Williams CK, Davison BH, Britovsek G, Cairney J, Eckert CA, Frederick WJ Jr, Hallett JP, Leak DJ, Liotta CL, Mielenz JR, Murphy R, Templer R, Tschaplinski T (2006) The path forward for biofuels and biomaterials. Science 311:484–489
Sohrab HM, Amir HG, Meisam T, Gholamreza SJ, Gholam HN, Mehdi G, Mehdi A (2013) Lignocellulosic biomass to bioethanol, a comprehensive review with a focus on pretreatment. Renew Sust Energ Rev 27:77–93
Wheals AE, Basso LC, Alves DMG. Amorim HV (1999) Fuel ethanol after 25 years. Trends Biotechnol 17:482–487
Goldemberg J (2007) Ethanol for a sustainable energy future. Science 315:808–810
Balat M (2011) Production of bioethanol from lignocellulosic materials via the biochemical pathway: a review. Energ Convers Manag 52:858-875
Demirbas A (2003) Energy and environmental issues relating to greenhouse gas emissions in Turkey. Renew Sust Energ Rev 44:201–213
Gonzalez-Hernandez JL, Sarath G, Stein JM, Owens V, Gedye K, Boe A (2009) A multiple species approach to biomass production from native herbaceous perennial feedstocks. In Vitro Cell Dev Biol Plant 45:267–281
Boe A, Lee DK (2007) Genetic variation for biomass production in prairie cordgrass and switchgrass. Crop Sci 47:929–934
Boe A, Owens V, Gonzalez-Hernandez J, Stein J, Lee DK, Koo BC (2009) Morphology and biomass production of prairie cordgrass on marginal lands. GCB Bioenergy 3:240–250
Hernández KA, Owens VN, Boe A, González-Hernández JL, Lee DK, Aberle E (2018) Above- and belowground prairie cordgrass response to applied nitrogen on marginal land. BioEnergy Research 11:440–448. https://doi.org/10.1007/s12155-018-9908-1
Weaver JE (1954) North American prairie. Johnson Publishing Company, Lincoln, NE
Hansen PL, Chadde SW, Pfister RD (1988) Riparian dominance types of Montana (miscellaneous publication/Montana forest and conservation experiment station. No. 49. P. 411. Missoula, MT: University of Montana, School of Forestry, Montana Forest and Conservation Experiment Station
Potter L, Bingham MJ, Baker MG, Long SP (1995) The potential of two perennial C4 grasses and a perennial C4 sedge as ligno-cellulosic fuel crops in N.W. Europe. Crop establishment and yields in E. England. Ann Bot 75:513–520
Madakadze IC, Coulman BE, Mcelroy AR, Stewart KA, Smith DL (1998) Evaluation of selected warm-season grasses for biomass production in areas with a short growing season. Bioresour Technol 65:1–12
Johnson GA, Wyse DL, Sheaffer CC (2013) Yield of perennial herbaceous and woody biomass crops over time across three location. Biomass Bioenergy 58:567–574
Montemayor MB, Price JS, Rochefort L, Boudreau S (2008) Temporal variations and spatial patterns in saline and waterlogged peat fields: 1. Survival and growth of salt marsh graminoids. Environ Exp Bot 62:333–342
Anderson EK, Voigt TB, Kim SM, Lee DK (2015) Determining effects of sodicity and salinity on switchgrass and prairie cordgrass germination and plant growth. Ind Crop Prod 64:79–87
Owens VN, Viands DR, Mayton HS, Fike JH, Farris R, Heaton E, Bransby DI, Hong CO (2013) Nitrogen use in switchgrass grown for bioenergy across the USA. Biomass Bioenergy 56:286–293
Waramit N, Moore KJ, Heaton E (2014) Nitrogen and harvest date affect developmental morphology and biomass yield of warm-season grasses. GCB Bioenergy 6:534–543
Fike JH, Pease JW, Owens VN, Farris RL, Hansen JL, Heaton EA, Hong CO, Mayton HS, Mitchell RB, Viands DR (2017) Switchgrass nitrogen response and estimated production costs on diverse sites. GCB Bioenergy 9:1526–1542
Guo J, Thapa S, Voigt T, Owens V, Boe A, Lee DK (2017) Biomass yield and feedstock quality of prairie cordgrass in response to seeding rate, row spacing, and nitrogen fertilization. Agron J 109:1–12. https://doi.org/10.2134/agronj2017.03.0179
Lewandowski I, Clifton-Brown JC, Scurlock JMO, Huisman W (2000) Miscanthus: European experience with a novel energy crop. Biomass Bioenergy 19:209–227
Foti S, Cosentino SL, Patane C, Guarnaccia P (1996) Growth and yield of C4 species for biomass for energy and the environment. In: processing of the 9th European Bioenergy conference, Pergamon/Elsevier, Copenhagen, Denmark, pp 616-621
Donalatos NG, Dalianis C, Kyristis S (1998) Influence of fertilization and irrigation on the growth and biomass productivity of miscanthus sinensis x giganteus under Greek conditions. In: James and James (ed) sustainable agriculture for food energy and industry. Science Publishers, Braunschweig, Germany, pp 319–323
Miguez FE, Villamil MB, Long SP, Bollero GA (2008) Meta-analysis of the effects of management factors on Miscanthus × giganteus growth and biomass production. Agric For Meteorol 148:1280–1292
Sheaffer CC, Ehlke NJ, Albrecht KA, Peterson PR (2003) Forage legumes. Minnesota Agric Exp Stn Bull:608–2003
Peterson PR, Sheaffer CC, Jordan RM, Christians CJ (1994) Response of kura clover to sheep grazing and clipping: I. Yield and forage quality. Agron J 86:655–660
Kim BW (1996) Kura clover development and performance of kura clover-grass mixtures. Dissertation, University of Wisconsin, Madison, USA, Ph.D
Peoples MB, Herridge DE, Ladha JK (1995) Biological nitrogen fixation: an efficient source of nitrogen for sustainable agricultural production? Plant Soil 174:3–28
Graham PH, Vance CP (2000) Nitrogen fixation in perspective: an overview of research and extension needs. Field Crop Res 65:93–106
Ledgard SF, Steele KW (1992) Biological nitrogen fixation in mixed legume/grass pastures. Plant Soil 141:137–153
Paynel F, Murray PJ, Cliquet JB (2001) Root exudates: a pathway for short-term N transfer from clover to ryegrass. Plant Soil 229:235–243
Barnett FL, Posler GL (1983) Performance of cool−season perennial grasses in pure stands and in mixtures with legumes. Agron J 75:582–586
Heichel GH, Henjum KI (1991) Dinitrogen fixation, nitrogen transfer, and productivity of forage legume-grass communities. Crop Sci 31:202–208
Zemenchik RA, Albrecht KA, Schultz MK (2001) Nitrogen replacement values of kura clover and birdsfoot trefoil in mixtures with cool-season grasses. Agron J 93:451–458
Zemenchik RA, Albrecht KA (2002) Nitrogen use efficiency and apparent nitrogen recovery of Kentucky bluegrass, smooth bromegrass and orchardgrass. Agron J 94:421–428
Casler MD, Vogel KP, Taliaferro CM, Ehlke NJ, Berdahl JD, Brummer EC, Kallenbach RL, West CP, Mitchell RB (2007) Latitudinal and longitudinal adaptation of switchgrass populations. Crop Sci 47:2249–2260
Rao BRR (2002) Biomass yield, essential oil yield and essential oil composition of rose-scented geranium (Pelargonium species) as influenced by row spacings and intercropping with corn mint (Mentha arvensis L.f. piperascens Malinv. ex Holmes). Ind Crop Prod 16:133–144
Grafton KF, Schneiter AA, Nagle BJ (1988) Row spacing, plant population, and genotype × row spacing interaction effects on yield and yield components of dry bean. Agron J 80:631–634
Mahmood A, Honermeier B (2012) Effect of row spacing and cultivar on biomass yield and quality of Sorghum bicolor L. Moench J Kult Pflanzenm 64:250–257
Lee DK, Aberle E, Anderson EK, Anderson W, Baldwin BS, Baltensperger D, Barrett M, Blumenthal J, Bonos S, Bouton J, Bransby DI, Brummer C, Burks PS, Chen C, Daly C, Egenolf J, Farris RL, Fike JH, Gaussoin R, Gill JR, Gravois K, Halbleib MD, Hale A, Wanna W, Harmoney K, Heaton EA, Heiniger RW, Hoffman L, Hong CO, Kakani G, Kallenbach R, Macoon B, Medley AC, Missaoui A, Mitchell R, Moore KJ, Morrison JI, Odvody GN, Ogoshi R, Parrish JR, Quinn L, Richard E, Rooney WL, Rushing JB, Schnell R, Sousek M, Staggenborg SA, Tew T, Uehara G, Viands DR, Voigt T, Williams D, Williams L, Wilson LT, Wycislo A, Yang Y, Owens V (2018) Biomass production of herbaceous energy crops in the United States: field trial results and yield potential maps from the multiyear regional feedstock partnership. GCB Bioenergy 10:698–716. https://doi.org/10.1111/gcbb.12493
Pedroso GM, Hutmacher RB, Putnam D, Six J, Kessel CV, Linquist BA (2014) Biomass yield and nitrogen use of potential C4 and C3 dedicated energy crops in a Mediterranean climate. Field Crop Res 161:149–157
Lemus R, Brummer C, Burras CL, Moore KJ, Barker MF, Molstad NE (2008) Effects of nitrogen fertilization on biomass yield and quality in large fields of established switchgrass in southern Iowa, USA. Biomass Bioenergy 32:1187–1194
Christian DG, Riche AB, Yates NE (2002) The yield and composition of switchgrass and coastal panic grass grown as a biofuel in southern England. Bioresour Technol 83:115–124
Beale CV, Long SP (1997) Seasonal dynamics of nutrient accumulation and partitioning in the perennial C4-grass Miscanthus × giganteus and Spartina cynosuroides. Biomass Bioenergy 12:419–428
Vogel KP, Brejda JJ, Walters DT, Buxton DR (2002) Switchgrass biomass production in the Midwest USA: harvest and nitrogen management. Agron J 94:413–420
Guretzky JA, Biermacher JT, Cook BJ, Kering MK, Mosali J (2011) Switchgrass for forage and bioenergy: harvest and nitrogen rate effects on biomass yields and nutrient composition. Plant Soil 339:69–81
Clark FE (1977) Internal cycling of nitrogen in shortgrass prairie. Ecology 58:1322–1333
Parrish DJ, Fike JH (2005) The biology and agronomy switchgrass for biofuels. Crc Crit Rev Plant Sci 24:423–459
Smart AJ, Moser LE, Vogel KP (2004) Morphological characteristics of big bluestem and switchgrass plants divergently selected for seedling tiller number. Crop Sci 44:607–613
Danalatos NG, Archontoulis SV, Mitsios I (2007) Potential growth and biomass productivity of Miscanthus × giganteus as affected by plant density and N-fertilization in central Greece. Biomass Bioenergy 31:145–152
Mazzanti A, Lemaire G, Gastal F (1994) The effect of nitrogen fertilization upon the herbage production of tall fescue swards continuously grazed with sheep. 1. Herbage growth dynamics. Grass Forage Sci 49:111–120
Garwood EA (1969) Seasonal tiller populations of grass and grass/clover swards with and without irrigation. J.Br Grassl Soc 24:333-344
Davies A (1971) Changes in growth rate and morphology of perennial ryegrass swards at high and low nitrogen levels. J Agric Sci 77:123–134
Snider JL, Raper RL, Schwab EB (2012) The effect of row spacing and seeding rate on biomass production and plant stand characteristics of non-irrigated photoperiod-sensitive sorghum (Sorghum bicolor (L.) Moench). Ind Crop Prod 37:527–535
Fraser A, Kindscher K (2005) Spatial distribution of Spartina pectinata transplants to restore wet prairie. Restor Ecol 13:144–151
Jakubowski A, Casler M, Jackson R (2017) Legume addition to perennial warm-season grass swards increases harvested biomass. Crop Sci 57:3343–3351
Sanderson MA, Soder KJ, Muller LD, Klement KD, Skinner RH, Goslee SC (2005) Forage mixture productivity and botanical composition in pastures grazed by dairy cattle. Agron J 97:465–1471
Tracy B, Sanderson M (2004) Productivity and stability relationships in mowed pasture communities of varying species composition. Crop Sci 44:2180–2186
Tilman D (1996) Biodiversity: population versus ecosystem stability. Ecology 77:350–356
Boller B, Nösberger J (1994) Difference in nitrogen fixation among field-grown red clover strains at different levels of 15N fertilization. Euphytica 78:167–174
Herridge DF, Peoples MB, Boddey RM (2008) Global input of biological nitrogen fixation in agricultural systems. Plant Soil 311:1–18
Jensen EE, People MB, Boddey PM, Gresshoff H, Hauggaard-Nielsen H, Alves B, Morrison MJ (2012) Legumes for mitigation of climate change and the provision of feedstock for biofuels and biorefineries. A review. Agronomy for Sustainable Development 32:329–364
Nesheim L, Øyen J (1994) Nitrogen fixation by red clover (Trifolium pratense L.) grown in mixtures with Timothy (Phleum pratense L.) at different levels of nitrogen fertilization. Acta Agric Scand B Soil Plant Sci 44:28–24
Funding
This research was supported by funding from the North Central Regional Sun Grant Center at South Dakota State University through a grant provided by the US Department of Energy Bioenergy Technologies Office under award no. DE-FG36-08GO88073.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kim, S., Albrecht, K., Sheaffer, C. et al. Biomass Production of Prairie Cordgrass (Spartina pectinata Link.) Using Urea and Kura Clover (Trifolium ambiguum Bieb.) as a Source of Nitrogen. Bioenerg. Res. 13, 1095–1107 (2020). https://doi.org/10.1007/s12155-020-10139-2
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12155-020-10139-2