Cellular, Molecular, and Physiological Aspects of In Vitro Plant Regeneration

1
Department of Horticulture, College of Agriculture, Isfahan University of Technology, Isfahan 84156-83111, Iran
2
Department of Agricultural Sciences, University of Helsinki, PL-27 Helsinki, Finland
*
Author to whom correspondence should be addressed.
Plants 2020, 9(6), 702; https://doi.org/10.3390/plants9060702
Submission received: 14 May 2020
/
Revised: 25 May 2020
/
Accepted: 27 May 2020
/
Published: 1 June 2020
(This article belongs to the Special Issue Advances in Plant Regeneration)
{kind=link}
{kind=link}
Abstract
:Plants generally have the highest regenerative ability because they show a high degree of developmental plasticity. Although the basic principles of plant regeneration date back many years, understanding the cellular, molecular, and physiological mechanisms based on these principles is currently in progress. In addition to the significant effects of some factors such as medium components, phytohormones, explant type, and light on the regeneration ability of an explant, recent reports evidence the involvement of molecular signals in organogenesis and embryogenesis responses to explant wounding, induced plant cell death, and phytohormones interaction. However, some cellular behaviors such as the occurrence of somaclonal variations and abnormalities during the in vitro plant regeneration process may be associated with adverse effects on the efficacy of plant regeneration. A review of past studies suggests that, in some cases, regeneration in plants involves the reprogramming of distinct somatic cells, while in others, it is induced by the activation of relatively undifferentiated cells in somatic tissues. However, this review covers the most important factors involved in the process of plant regeneration and discusses the mechanisms by which plants monitor this process.
1. Introduction
The initiation of in vitro studies of plant cells and tissue culture dates back to 1902, when Gottlieb Haberland presented a “totipotency” hypothesis that each cell has all the genetic information needed to produce a perfect plant [1,2]. Differentiated cells in plants are able to re-enter the cell cycle, proliferate and regenerate tissues and organs, and even become a complete plant, according to this hypothesis. Several reports have shown the totipotent ability of plant cells through which the plant can be regenerated, which in turn is widely used in several basic studies such as in micropropagation, germplasm conservation, and formation of genetically modified plants [3,4]. Plants have powerful regenerative abilities thanks to the property of developmental plasticity of their cells [5,6]. In vitro plant regeneration is a process in which explants, after undergoing cell division and differentiation, form organs and tissues throughout their growth period [7,8]. In vitro plant regeneration can be performed via somatic embryogenesis or organogenesis [9]. Organogenesis is the process by which new organs and even whole plants are usually formed in response to wounds from other parts of the organs. In somatic embryogenesis, first, a structural cell similar to zygotic embryos is formed, and then the entire plant is regenerated [6,10,11,12,13]. The potential for plant regeneration, which has long been used to propagate clones, cuttings, and grafts, is the basis of ongoing research and agricultural applications [14]. Micropropagation has been applied commercially worldwide, although the capability of plant regeneration varies significantly in different genotypes [6,14,15,16]. During the last several years, several agents regulating plant regeneration have been studied, such as exogenously supplied phytohormones in vitro [17,18,19], explant type [5,20,21,22], physiological properties of the donor plants [23,24], mineral uptake and their distribution patterns [25,26], changes in mevalonate kinase activity [27], and reprogramming of differentiated somatic cells and activation of relatively undifferentiated cells in somatic tissues [6]. Nontraditional inducers such as some amino acids [28]; light intensity and quality [29]; weak electric current [30]; and some antibiotics, for example, cefotaxime [31], have also been reported to affect in vitro plant regeneration. Rathore and Goldsworthy [30] passed very weak electric current 1 microamp between the tissue and the culture medium and noticed a dramatic increase in tobacco callus growth. Azmi et al. [32] reported the beneficial effects of a mixed light color of LED (red and blue) on in vitro plant regeneration of Rosa kordesii. This review covers novel findings of how plants adjust regeneration in terms of the cellular, molecular and physiological aspects and discuss influence of developmental and environmental factors on plant regeneration efficiency.
2. Organogenesis
Plant shoots and roots are able to retain their apical meristem functions even after a part of their meristems is removed. However, when the whole meristems are excised, plant cells of differentiated tissues or organs have the ability to produce new shoots and lateral roots via organogenesis [6,8,16,33]. In vitro plant regeneration by organogenesis is the result of organ formation through dedifferentiation of differentiated cells and reorganization of cell division to create particular organ primordia and meristems after the vascular connection between the explant and the newly regenerating organ [34,35].
3. Somatic Embryogenesis
Somatic embryogenesis is one of the biotechnological techniques for multiplication of important economic cultivars. This process is a type of plant cell totipotency in which embryos arise from somatic or vegetative cells if no fertilization takes place [36,37]. Several factors such as the origin of the explant, culture medium, and in vitro environmental conditions affect the success or failure of the somatic embryogenesis response [36]. Somatic cells undergo embryogenesis stages by developing structures similar to zygotic embryos without merging of gametes [38,39,40]. Somatic embryogenesis could be well suited for mass propagation of endangered crop species [41] and for commercial production. When somatic embryos are formed directly without a callus intermediate stage, this process is called direct embryogenesis [6], and is useful for rapid plant regeneration and minimizing somaclonal and chimeric variations [42]. Successful clonal propagation of elite genotypes requires a high percentage of genetic homogeneity among all regenerates. Therefore, the genetic homogeneity of in vitro regenerated plants is highly noticeable at an early stage of this process. Several strategies such as morpho-physiological, biochemical, cytological, and DNA-based molecular markers approaches have been employed to maintain the genetic constancy of the in vitro regenerated plantlets. Detection of the genetic homogeneity in micropropagated plants using molecular techniques could be achieved with polymerase chain reaction (PCR)-based techniques such as random amplified polymorphic DNA (RAPD) and inter simple sequence repeat (ISSR) [43,44]. For plant crops that are difficult to breed or have a poor genetic basis, somaclonal variation can be a very useful option for breeders as a new option [45]. Indirect plant regeneration is carried out by organogenesis or embryogenesis in two steps. In the first step, callus is induced, followed by the second stage, in which the shoot meristems or somatic embryos are initiated from the callus tissues, resulting in an organ formation [6,46]. Choosing the right explant, medium, phytohormones, genotype, carbohydrate, and gelling agent, as well as some other agents such as light regime, temperature, and humidity, noticeably affects organogenesis and embryogenesis processes [29]. Shoot clumps can be regenerated from shoot tips or bud stems that have only one bud, various mature somatic tissues, pollen, and protoplasts [6,47,48]. Protoplasts possess the ability to develop new cell wall and to regenerate complete plants when grown in an appropriate culture medium. Crop improvement could be facilitated by genome editing in regeneration from protoplasts [49]. By genome editing, it is possible to modify genome sequences as well as modify the arrangement of gene expression patterns in a pre-specified area of an organism. Genome editing covers wide spectra of techniques applying either a site-specific recombinase (SSR) or site-specific nuclease (SSN) system. Genome editing is speedy with a very low hazard of unforeseen effects, and can be employed with any crop, even those that have complex genomes and are difficult to breed [50]. Modulation of phytohormone types, ratio, and concentration has been applied as an efficient approach to optimize organogenesis [51,52].
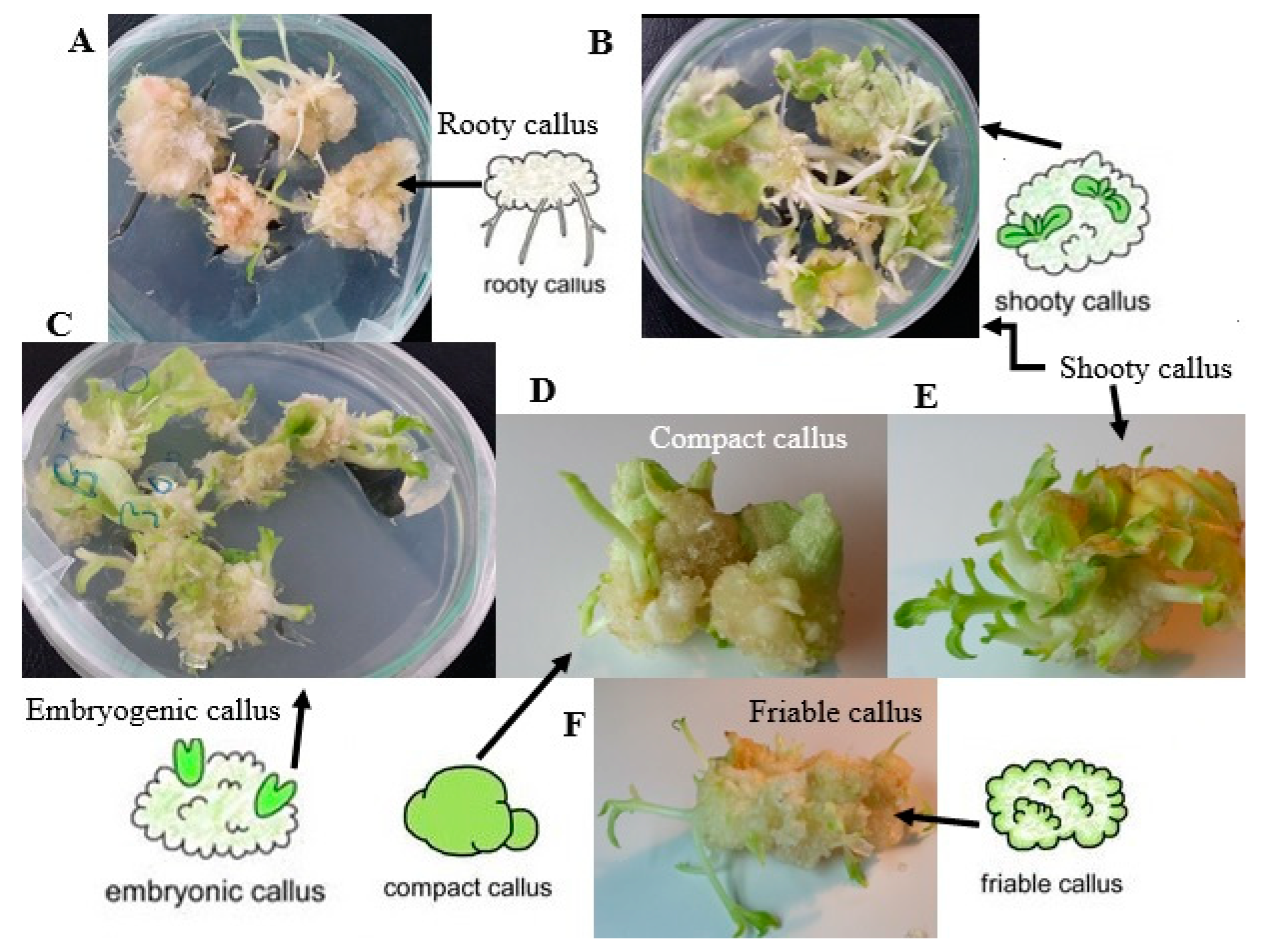
Somatic embryos have been reported to regenerate from a different type of explants, such as leaf explants [53,54], root explants [55], glandular trichomes [56], or haploid cells resulting from cell meiosis of both male and female gametophyte [57], or even fully differentiated stomatal guard cells [58]. Somatic embryogenesis is a more preferred pathway than organogenesis in mass propagation owing to the higher proliferation rate, more convenient use of liquid culture medium, the handling of a large number of embryos at a time, and more possibilities for applying bioreactors [59,60,61]. Homogeneous dispersion of nutrients and better uptake of medium constitutes by the explants is one of the advantages of liquid culture compared with solid culture medium, which subsequently causes further growth of the cultures in a suitable bioreactor system [62]. However, the most important drawback of liquid culture is overproduction of regenerations showing a higher rate of hyperhydricity [62]. Hyperhydricity can also be accelerated by exposing explants to different stresses or unsuitable growth regulator treatments [37]. The direct somatic embryogenesis process involves induction, maintenance of embryogenic cultures, embryo development and maturation, embryo germination, and plant regeneration [63,64]. Since the first report on carrot in 1958 [65], many plant species have been reported to produce somatic embryogenesis. Figure 1 shows morphology of different stages during plant organogenesis in Nicotiana rustica. Although in vitro regeneration techniques via somatic embryogenesis have been optimized for many crops, some important crop species, such as Hemerocallis sp L. [66] and Corylus avellana L. [67], are still difficult to multiply in vitro. Therefore, understanding the physiological, cellular, and molecular mechanisms of plant regeneration is important to address unanswered fundamental questions in cell and developmental biology.
4. Photoautotrophic Micro-Propagation
Most explants that have chlorophyll and photosynthesize can grow in a sugar-free medium or photoautotrophic growing condition [68]. This tissue culture technique is also known as photosynthetic micropropagation or inorganic micropropagation [69]. The high cost of production has limited the widespread use of micropropagation. Most of these limitations could be attributed to the heterotrophic characteristics of explants that require growth in a sugar-bearing culture medium. On the other hand, high relative humidity, high ethylene concentration, as well as low CO2 concentrations around explants grown in conventional tissue culture systems are other problems with heterotrophic cultures. Therefore, autotrophic culture is the best option to solve these problems [68,70]. Reduced physiologic and morphologic abnormalities, minimized biological contamination and prevented plant loss, ease of rooting, and acclimatization in vitro and in vivo are some important advantages of photoautotrophic micropropagation. Meanwhile, enhancement in cost for CO2 enrichment and lighting for an efficient photoautotrophic growth of the plantlets are some of the disadvantages of photoautotrophic micropropagation [70].
5. Application of Wide-Spectrum Light-Emitting Diodes in Micropropagation
Even in the case of heterotrophic explants that need a sugar source to grow in the culture medium, the key role of light in the activity of genes and enzymes, as well as the growth of explants, cannot be ignored [32,71]. So far, various light sources such as high-pressure sodium or fluorescent lamps have been used in in vitro cultures [72]. Because fluorescent lamps (FL) come in many varieties and provide a wide spectra of light (350–750 nm), they are used for many plant species, although their drawbacks include high power consumption, unstable radiation parameters, and noticeable heat emission [73]. LED lighting is more cost effective than fluorescent lamps [74]. Miler et al. [75] reported the ability to use several light-emitting diodes (LEDs) for the in vitro regeneration of some ornamental plant species such as Chrysanthemum × grandiflorum, Gerbera jamesonii, Heuchera × hybrida, Ficus benjamina, and Lamprocapnos spectabilis. Ramirez-Mosqueda et al. [76] tested five types of wavelengths in plant regeneration in vitro. They concluded that blue light (460 nm) caused longitudinal growth of regenerated shoots and improved chlorophyll synthesis in the explants.
6. Organogenesis in Response to Explant Wounding
Plant regeneration after tissue damage is termed as de novo organogenesis, in which organs such as shoots and roots are regenerated from the wound location and detached organs [77]. Organogenesis via tissue wounding includes three successive steps: (1) in the first step, signals are provided to stimulate the regeneration process, (2) then phytohormone accumulation is performed, leading to (3) cell fat transition [2,77]. Given that the regeneration process begins more than the cut end of the explants, the main induction stimulus for the regeneration phenomenon could be attributed to wound stimuli [2,6,78]. Wounding causes an enhanced cytokinin biosynthesis, which in turn increases cell proliferation and callus formation [79]. The wound created in the explants not only triggers the production of auxins to induce cells to regenerate, but also activates signaling pathways that are responsible for the emergence of root tips [77,80,81]. Recent reports have demonstrated that transcription factor genes are involved in the wounding process and promotion of root rip emergence. Any factor that interferes with the signal pathway negatively affects the emergence of adventitious root tips. Finally, it should be noted that the wound has complex roles in the de novo organogenesis process and that genes regulate the cellular environment for organ emergence [81].
7. Embryogenesis in Response to Explant Wounding
Somatic embryogenesis is the process in which ectopic embryos is arisen from asexual cells, without gamete formation, fertilization, or seed development [82]. Phytohormones or abiotic stresses usually affect embryogenesis from somatic tissues [83]. In addition to hormone-mediated somatic embryogenesis-induction, somatic embryos can also be accelerated by overexpression of specific transcription factor (TF) genes, such as the homeodomain TF WUSCHEL (WUS), the AP2 TFs PLETHORA 4/BABY BOOM (PLT4/BBM), PLT5/EMBRYO MAKER (PLT5/EMK), the MADS box TF AGAMOUS-LIKE 15 (AGL15), the LEAFY COTYLEDON genes LEC1, LEC2, and overexpression of the SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASE 1 (SERK1) [84,85,86,87,88,89,90,91]. Recently, Mozgova et al. [92] showed that polycomb repressive complex 2 (PRC2)-activity, known as an epigenetic processor of developmental phase transitions in plants [93], inflicted an obstacle to hormone-mediated transcriptional reprogramming to embryogenesis in vegetative tissue of Arabidopsis thaliana. Discovering molecular mechanisms for controlling the somatic embryogenesis process will be one of the most important approaches to identify the factors that control in vitro embryogenesis.
8. Cellular Origins and Plant Regeneration
The cellular behaviour studies are very important in plants to differentiate between embryogenic and nonembryogenic calli [94]. Taha and Wafa [94] investigated cellular behaviour to detect the somaclonal variations in vitro. However, cellular behaviour in regenerates and intact plants needs to be evaluated to determine the occurrence of somaclonal variation in the plant regeneration process.
8.1. Changes in Cellular Behaviour during In Vitro Plant Regeneration
Plants possess a greater cellular plasticity than those observed in the other organisms, which dramatically guarantees the cell’s ability to regenerate [6]. Recent findings on plant tissue and organ regeneration indicate that a cell may commence follow four regeneration process including cell death, division, dedifferentiation, and trans-differentiation. These studies have outlined comprehensive perspectives of regeneration at the cellular level and help a lot to know the regenerative capacity of plant cells [95].
8.2. Programmed Cell Death in Plants
Programmed cell death (PCD) in plants often occurs as a result of DNA damage, showing autolytic features, and has a noticeable role in the induction of tissue and organ regeneration [96]. However, the underlying mechanisms responsible for these mechanisms remain largely unknown. Induction of PCD takes place by some plant-specific transcription factors such as SUPPRESSOR OF GAMMA RESPONSE 1 (SOG1) and ETHYLENE RESPONSE FACTOR115 (ERF115)-PHYTOCHROME A SIGNAL TRANSDUCTION1 (PAT1), respectively [97,98]. The induced plant cell death accelerates regeneration responses, which in turn changes the expression of genes involved in cell division process, resulting in enhanced cell division [98].
Although it is not clear yet how regenerative cells are induced in response to the cell death, mechanical disarray caused by cell death, affecting orientation in cell division of appending cells, reinforces the possibility of mechanical regulation in regeneration process [99,100]. Any cellular modifications to reduce specialization are called dedifferentiation [101], whereas transdifferentiation is defined as the jump from one type of specialized cell to another type [102]. Nguyen and McCurdy [103] asserted that dedifferentiation could be part of transdifferentiation. Because of the property of callus as proliferating mass of dedifferentiated cells, dedifferentiation is strongly associated with callus formation. Thus, the formation of callus can be considered as a kind of transdifferentiation [4]. However, identifying dedifferentiation and transdifferentiation remains unexplored, and must be clarified in future. Besides, the question of whether differentiation and transdifferentiation contribute to callus formation should be addressed at the cellular level on plant regeneration in the future [2].
8.3. Cell Fate Reprogramming and Pluripotency Acquisition
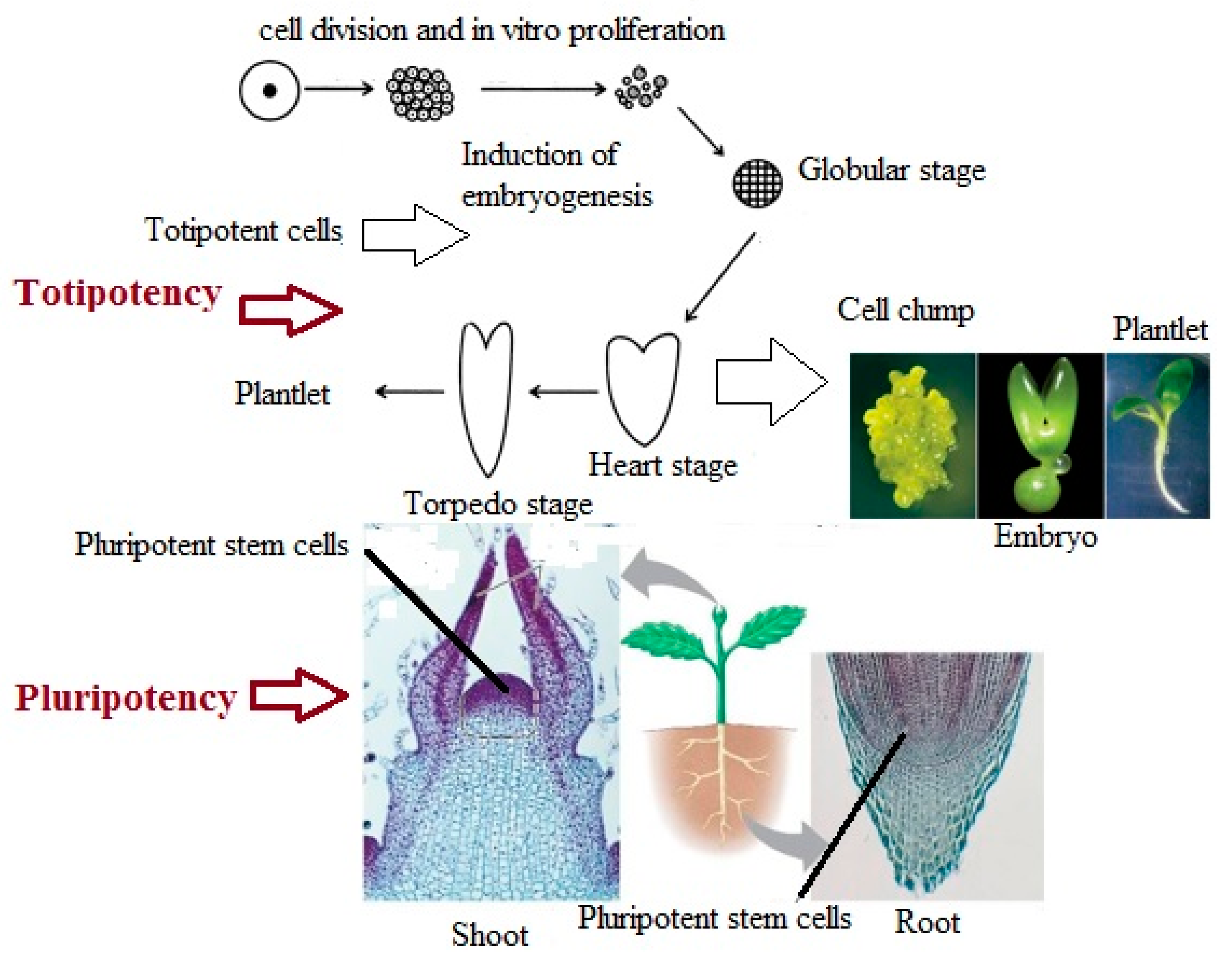
Pluripotency is defined as the ability of unique cells in the plant’s meristems to become an adult organism in response to environmental agents [104]. As can be seen in Figure 2, pluripotent cells are present in the root and shoot apices, where they create cells and tissues, but do not have the capability to create an embryo. Vice versa, under different circumstances, a somatic plant cell can dedifferentiate to generate a totipotent embryogenic cell that has the capability to produce an embryo [105]. According to Ikeuchi et al. [6], plants’ regeneration process is performed throughout two distinct cellular strategies. One is by reactivating cells that are not sufficiently differentiated, and the other is by reprogramming them into somatic cells. In both cases, regeneration relies on the phenomenon of cellular flexibility, which can be widely specified as the capability to redefine cell fate. Recent findings have demonstrated that finally differentiated cells can be reprogrammed into pluripotent cells, which corroborate the reversibility of cell differentiation [104]. Therefore, modulation of signaling pathways may enhance somatic cell reprogramming. However, the mechanisms by which somatic cells dedifferentiate into pluripotency are still unknown and need to be addressed.
8.4. Wound Responses and Signaling during Plant Regeneration
Wounding in the explant is the first incident in plant regeneration [79]. Wound signals such as electric current, hydraulic pressure, Ca2+, reactive oxygen species (ROS), oligopeptide system, oligosaccharides, jasmonic acid, salicylic acid, ethylene, abscisic acid, and changes in various metabolic processes of plant metabolism play a very important role in the regeneration process [106,107]. The results of analysis of the genes downstream of wound signaling indicated that wounding significantly affects plant regeneration [108,109]. However, information on how the wound signals affect in vitro plant regeneration is still insufficient [77,109,110]. Ikeuchi et al. [79], using transcriptome analysis and quantitative hormonal analysis, investigated how wounding causes callus formation in Arabidopsis (Arabidopsis thaliana). They concluded that wounding changes the gene expression involved in hormone biosynthesis, resulting in an enhanced accumulation of cytokinin, which is vital for wound-induced callus formation. Chen et al. [109] reported the involvement of short-term and long-term wound signaling in plant regeneration. Short- term wound signaling has three stages of signal delivery. Stage (1) lasts for a few hours and the wound signal diffuses swiftly from the wound location to the mesophyll cells and activates YUCCA1 (YUC1) and YUC4 expression in there. In stage (2), expression of YUCs transcription factors causes the production of auxin within 4 h, and then polar transmission to regenerating cells around the wound location, which lasts for about 12 h after the wounding. In stage 3, when auxin reaches regenerating cells, the expression of WUSCHEL RELATED HOMEOBOX11 (WOX11) and WOX12 causes the cell fate transition to regenerate organs at around 1 to 2 days after wounding [109,111]. During the long-term wound signaling, YUC4 as well as a group of NAC (NAM, ATAF1 and 2, and CUC2) transcription factor genes including NAC1, which are present in the cells near to the wound location, are activated. The task of NAC1 is to control the proliferation process through cell wall metabolism [109]. Expression of YUC4 transcription factors results in a high auxin accumulation in the regenerating cells [109]. The correlation between activation of NAC1 and YUC4 to produce auxin remains a mystery that needs to be clarified in the future [109]. Recently, Rymen et al. [112] also showed the main mechanism of epigenetics, which is based on wound induced cell reprograming of wound healing in plants. They asserted the expression of some wound-induced transcriptional factors such as WIND1, H3K9/14ac, and H3K27ac immediately after wounding of explant. However, wounding possesses intricate biological impact and has multiple tasks in plant regeneration, but how the wound re-activates cell proliferation and accelerates cellular reprogramming is not very clear yet and needs to be addressed more than ever to clarify all aspects of these process [79,109].
9. Molecular Basis of Plant Regeneration
9.1. Molecular Mechanisms Involved in Plant Regeneration
Recent progress in molecular techniques has led to a major perception of the underlying processes of plant regeneration. Depending on the direct or indirect shoot regeneration, the plant cells do not readily regenerate shoots, unless transferred to a callus or shoot induction medium. In terms of indirect organogenesis, induced callus simulates lateral root meristem followed by shoot regeneration upon being transferred to shoot induction medium. However, in direct plant regeneration, in the process of regenerating a shoot in vitro, somatic cells first respond to phytohormones, then the responsive cells begin to divide, and eventually new shoots appear [4,33,113]. Cytokines and auxins are the most pervasive phytohormones, and either directly or indirectly accelerate the shoot regeneration process. Molecular studies have also shown the presence of important genes in the pathway of cytokinin signaling in plants [114]. Of the most important receptors for cytokinin are the histidine protein kinases (AHKs), while the histidine phosphotransfer proteins (AHPs) are responsible for the transfer of the signal from AHKs, the result of which may be activation or suppression of shoot regeneration. Several regulatory genes, such as KNOTTED1 (KN1), SHOOT MERISTEMLESS (STM), WUSCHEL (WUS), and CLAVATA 1-3 (CLV1-3), have been identified in shoot regeneration [115,116,117,118]. The shoot regeneration process can be considered as the result of interconnections among cytokinin receptors, cell cycles, and development of shoot meristem [119,120]. Some transcription factors such as CUPSHAPED COTYLEDON1 (CUC1) and CUC2 have been reported to be responsible for shoot meristem regeneration during embryogenesis and are activated through expression of transcription factors PLT3, PLT5, and PLT7 [121]. The expression of PLT1, PLT2, CUC, CUC2, and WIND1-4 is caused by wounding results in the attainment of pluripotency at the wounding location of explants [122,123]. When the explants are transferred to the shoot regeneration medium, which is also enriched with cytokines, the expression of transcription factor SHOOT MERISTEM REGULATOR WUSCHEL (WUS) results in enhanced shoot regeneration [122,124]. Upon transferring on shoot-inducing medium, some other transcription factors, such as ENHANCER OF SHOOT REGENERATION1 (ESR1) and ESR2, which stimulate embryogenesis process and shoot regeneration, are induced by enhancing CUC1 expression [125]. Root regeneration from explant has been reported to be the result of the expression of some transcription factors such as WUSCHEL RELATED HOMEOBOX11 (WOX11) and WOX12. These transcription factors responsible for rooting induce the expression of LATERAL ORGAN BOUNDARIES DOMAIN16 (LBD16), LBD29, and then WOX5 in response to auxin-supplemented medium [6,111]. The family of LBDs and WOX5 also has a role in lateral root development [126]. Some families of the transcription factor AUXIN RESPONSE FACTOR (ARF) have been reported to activate the expression of WOX11 in root regeneration of leaf samples [111].
9.2. Biochemical Changes during Plant Regeneration
Previous reports demonstrated the involvement of oxidative stress in plant regeneration process [127,128]. Some of the important events in the plant regeneration process such as programmed cell death, phytohormone signaling pathways, and differentiation of cells have been reported to be influenced by reactive oxygen species (ROS) [129]. Although previous studies have pointed to the dual role of ROS in regenerating plants that are both toxic and accelerating, very little has been reported on the ROS effects on in vitro plant regeneration [130]. Employing antioxidants in plant tissues that scavenge ROS negatively affects metabolic pathways in plant cells that are critical for organ differentiation [130]. Overproduction of ROS has been found to be linked with shoot regeneration and is needed in the early stages of shoot regeneration [130].
9.3. Somaclonal Variation during Plant Regeneration Process
Natural variation in vitro plant regeneration is a matter of concern to plant breeders. The uniformity of obtained plants within clone propagation is desired in commercial plant propagation [131]. However, induction of genetic variability in undifferentiated cells, isolated protoplasts, callus, and tissues of in vitro obtained plants should not be overlooked [132,133]. Wide spectra of variation in regenerated plants have been shown in banana tissue culture [134,135]. The origin of variation arising from in vitro plant regeneration may be both genetic and non-genetic [136]. Genetic variation induced in plant regeneration, called somaclonal variation, is undesirable to propagate true-to-type plants from a selected genotype [45,137]. An understanding of genetic variability for in vitro plant regeneration process is very beneficial for identifying novel factors that improve the efficiency of regeneration [138,139]. The most important factors involved in variations in tissue culture are wounding, explant sterilization, misbalance of media components such as sources of phytohormones used, sugar source, illumination, and humidity [43,140,141,142]. Oxidative stress damage during in vitro plant regeneration may also result in variation [143,144]. Type of tissue source is another important factor that can cause many variations in the plant regeneration process [135]. The tissues with higher differentiation properties such as leaf and root explants produce more variations than explants having meristem such as shoot tips [145]. The existence of somatic mutations in donor plants (existence of chimera in explants) is another source of variation in plant regeneration [146]. When regenerating via axillary branching, plants show variation, while the cultures that pass through the callus stage have a greater chance of variation [147]. Fast proliferation of a tissue in micropropagation process by shortening the subculture period is one of the items contributing to reducing the rate of variations in in vitro plant regeneration [148,149]. Meanwhile, the prolong subcultures of in vitro tissues enhances changes in DNA methylation [150]. Tanurdzic et al. [143] showed that tissue culture may reactivate silent involved genes, resulting in somaclonal variations. The occurrence of somaclonal variation is also affected by external agents such as phytohormones (both concentration and the ratio of different plant growth regulators), temperature, and light intensity of culture media [149,150,151,152,153,154,155,156]. It has been reported that 2, 4- Dichlorophenoxyacetic acid (2, 4- d) resulted in enhanced DNA ploidy levels and methylation events in in vitro plant regeneration cultures [157]. Among all factors affecting somaclonal variation, plant genotype influence can undoubtedly be one of the most important factors influencing variation arisen from in vitro plant regeneration [154]. Phenotypic, cytological, biochemical, and genetic or epigenetic observations have been used to characterize somaclonal variation in plant regeneration systems [133].
The incidence of somaclonal variation in plant regeneration process has been reported to be the result of alterations in chromosome number [158], point mutations [159], chromosome breakage and rearrangement [160], DNA amplification [161], epigenetic variation [142], and separation of pre-existing chimeras in tissue [141].
9.4. Regeneration Capacity in Response to Epigenetic Mechanisms
Epigenetic regulation has an essential role in cell differentiation, which is a characteristic process involved in plant regeneration. DNA methylation has been reported as one of the most important factors in phenotypic changes and is considered as one of the most important epigenetic mechanisms that play a role in the plant regeneration process [162,163]. Obtaining true-to-type plants is the main target of large-scale clonal multiplication. Therefore, evaluation of genetic stability using molecular markers such as RAPD and ISSR must be included [164]. A recent literature review in the field of epigenetics by Miguel and Marum [164] has shown highly transformative mechanisms of chromatin remodeling in cell dedifferentiation and differentiation processes. The question to be addressed is whether epigenetic mechanisms are capable of disrupting cellular programming, which is necessary for plant regeneration [163]. Epigenetic events cause the expression of specific transcription factors in the plant regeneration process. The results of several studies showed that cell re-programming is associated with noticeable modifications in DNA methylation [163,165].
10. Physiological Responses of Plant Regeneration
Although the capability of plant cells to regenerate a perfect plant has long since been known, the question that needs to be addressed is how a somatic cell can become a whole plant [166]. Regeneration, which involves a wide range of healing from a small cut in the plant to the formation of an organ, or even a complete plantlet, results widely from physiological responses in plants. However, the mode of recovery of these missing organs varies considerably among plant species [5,7]. When the proper physiological triggers phytohormones, some stresses including wounding or pathogen infection are received by the plant, and somatic plant cells begin to create adventitious embryos, roots, and shoots [78]. The regeneration of plants takes place in three stages. In the first stage, plant tissue responds to embryogenesis or organogenesis stimuli in a process called dedifferentiation. It then (second step) enters the induction phase, during which cells are identified to produce shoot, root, or embryo. The last third step enters the realization stage, which results in the appearance of shoots, roots, and embryos [167]. Various factors such as environmental constrains, biotic stresses, and abnormalities affect the occurrence of in vitro plant regeneration, the most important of which will be described later in this review.
10.1. Developmental and Environmental Constraints on Plant Regeneration
Success in plant regeneration depends on several items such as explant type, nutrients, phytohormones, temperature, and illumination [168,169,170]. The regeneration capacity of an explant varies according to the stage of growth in which the donor plant is located and generally decreases with the aging of the plants [171]. The regeneration capacity of explants, obtained from juvenile plants, is much more than mature plants [172,173]. However, decreased regeneration capacity with age may be attributed to decreased responsiveness to phytohormones [174]. A significant decrease in expression of a microRNA that controls transferring from the juvenile to adult phase (miR156) in old plants results in the loss of regeneration capacity of the explants [173,174]. Light is one of the important environmental agents controlling regeneration in plant cell, tissue, and organ cultures [175]. Light intensity and quality are important factors that affect shoot regeneration in micropropagation of several crops [176,177,178]. LED lighting has been shown to be a suitable option for in vitro culture of various plant species. In addition to low costs, it provides better growth and physiological development for plantlets [76]. The efficacy of phytohormones such as their regulation and metabolism as well as adjustment of endogenous hormone levels is also affected by light [170,179]. The regulation of endogenous hormonal levels and phytohormones efficacy are also influenced by light [179]. Explant type, the constituents of media and genotype are important items affecting the success of in vitro plant regeneration. Several other agents in in vitro regeneration are sugar sources, gelling agent, and growth ingredients [180,181,182]. The process of in vitro plant regeneration begins with the formation of callus and then ends with the induction of shoot formation. During these two stages of the micropropagation process, the levels required by exogenous phytohormones in the culture medium may vary. Therefore, the success of in vitro plant regeneration is dependent on explant response by supplementation of phytohormones in the culture medium [183].
10.2. Biotic and Abiotic Stress Associated with Defense Responses in Regeneration Process
In vitro shoot induction has been reported to be influenced by abiotic stress [184]. Puijalon et al. [185] evaluated the regenerative capacity of rhizome explants in response to some abiotic stresses such as warm water, hot air, cold water, and sodium chloride (NaCl). On the basis of their own experiments, they reported positive effects of these stresses. Jose and Thomas [186] also showed the positive role of abiotic stresses in shoot induction from rhizome segments of Curcuma caesia in cultures. The methods of gene-profiling have detected some genes that act as molecular signatures of in vitro plant regeneration. One of these genes discovered in the early step of shoot induction and regeneration is RAP2.6L (At5g13330), a member of the ERF (ethylene response factor) subfamily B-4 of the ERF/APETALA2 transcription factor gene family. RAP2.6L apparently controls the expression of many other genes involved in the process of shoot regeneration [113]. Ravindran et al. [187] also reported the entity of a significant variability between different plant cultivars in terms of plant regeneration and resistance to biotic and abiotic stresses. The role of nanoparticles (NPs) in plant tissue culture has also been reviewed by Kim et al. [188]. Callus induction, organogenesis, somatic embryogenesis, somaclonal variation, genetic transformation, and secondary metabolite production are positively affected by NPs [188]. Although the benefit of NPs in the omission of microbial contaminants in plant tissue cultures has been reported to depend on their dimensions, size, distribution, and type [188], the mechanisms underlying acceleration or inhibition effects of NPs on each above mentioned factor remain unclear and need to be clarified in the future.
10.3. The Stimulatory Effect of Antibiotics on Plant Regeneration in Tissue Culture
The medium supplemented with antibiotics such as β-lactam antibiotics and tetracycline has been reported to result in higher regeneration capacity as compared with untreated explants [189,190]. Despite the successful application of antibiotics to suppress bacterial growth in vitro, excessive and regular use of them is detrimental to the regeneration process and results in a reduced regeneration ability of tissue cultures [191]. Antibiotics were reported either to hamper [192] or promote explant growth and development [193,194]. The mechanism of action of antibiotics and their effects on the regeneration process in plants is not yet clear. However, some antibiotics are thought to follow the activities of phytohormones, and some even have structures similar to auxin. [192]. The sensitivity of the explant being cultured in vitro may be specialized for any plant species and generally depends on the growing conditions, type of culture, and culture system [192]. Therefore, before using any antibiotic to prevent or minimize the growth of harmful microbes, the type and concentration of antibiotics with a thorough knowledge of their microbiological activities as well as with the least toxic effects on the regenerative process should be evaluated [195]. A stimulatory impact of some antibiotics such as β-Lactam (carbenicillin), cefotaxime, and timentin on in vitro plant regeneration has been previously reported [195,196,197].
10.4. Abnormalities in Plant Regeneration Process
In vitro plant propagation (micropropagation) is usually performed based on organogenesis or somatic embryogenesis. However, the main difficulty in plant regeneration is the large number of abnormalities of regenerated shoots and embryos that cannot be converted to normal plantlets [134,198,199]. The special conditions during in vitro culture result in the formation of physiologically, morphologically, and anatomically abnormal plantlets, and the understanding of these in vitro malformations is a precondition to expand an efficient plant regeneration protocol [198]. The signs of abnormalities are often characterized by a weak photosynthetic efficiency, poor stomata performance, and a significant decrease in reduction of cuticle wax [198]. Genetic or epigenetic modifications in the DNA are the main cause of abnormalities in somatic embryos (SE). Some external agents such as phytohormones application, mutagens, or even stressful factors significantly cause DNA modification. If the abnormality is the result of DNA modifications, it will be very difficult to reverse. Meanwhile, abnormalities caused by epigenetic modification may be reversible and the resulting abnormal embryos can regenerate to a perfect plantlet [43,134,199,200].
11. Cytokinin and Auxin Response to Enhance In Vitro Plant Regeneration
Despite the widespread use of micropropagation in many plant species that have great economic value, the ability of explant used to regenerate an entire plantlet depends greatly on the type of plant species used. Therefore, a comprehensive knowledge of the mechanisms by which the plantlet is regenerated is very important. The ratio of auxins to cytokines, depending on the range of plant species used for micropropagation, plays a very important role in the success of plant regeneration [14]. Signaling pathways in the plant in relation to the effect of phytohormones are one of the important goals in the direction of genetic manipulation to increase the regenerative capacity of explants in tissue culture conditions. Histidine-containing phosphotransfer (HPt) protein has been reported to be responsible for transferring the phosphate signal that regulates the expression of the genes responsible for cytokine regulation [201]. The enhanced expression of the genes responsible for cytokine regulation causes hypersensitivity to cytokinin in the multiplication process. By increasing the capacity of explants to respond to cytokinin, the genes responsible for cytokine regulation could be expressed as a tool to dominate the recalcitrance of in vitro plant regeneration in some crop species [14,201].
12. Conclusions and Future Perspectives
What we can understand from reviewing past studies is that many of the events that occur during the plant regeneration process can be controlled by manipulating signaling pathways related to the interaction of phytohormones, explant wounding, and programmed cell death. Although the key regulators of hormone signaling pathways have been previously discovered, more work is needed to understand how they retrieve cell proliferative capacity. We need to address a few questions: how explants understand and transmit endogenous and environmental signals, and how they induce or maintain cell differentiation. Moreover, it would be useful to study different mechanisms at both the molecular and physiological levels by which the explants regulate in vitro regeneration. The prospects of gene editing in differentiation of recalcitrant plants are as follows: we are still faced with a challenge of genetic dependence on in vitro plant regeneration via organogenesis, somatic embryogenesis, androgenesis, and protoplast regeneration. A big question is, can we have a common culture medium for most of the genotypes of different plants? The application of innovative tools with a multidisciplinary approach to address issues of in vitro plant regeneration for wider applications in crop improvement, commercial applications, and secondary metabolites should be investigated.
Author Contributions
S.S.B. prepared the figures and revised manuscript; S.M.J. supervised, corrected and reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Haberlandt, G. Culturversuehe mit isolierten Pflanzenzellen. Sitzungsber. Akad. Wiss. Wien Math. Naturwiss. 1902, 111, 69–92. [Google Scholar]
- Feher, A. Callus, dedifferentiation, totipotency, somatic embryogenesis: What these terms mean in the era of molecular plant biology? Front. Plant Sci. 2019, 10, 536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorpe, T.A. History of plant tissue culture. Mol. Biotechnol. 2007, 37, 169–180. [Google Scholar] [CrossRef]
- Sugimoto, K.; Gordon, S.P.; Meyerowitz, E.M. Regeneration in plants and animals: Dedifferentiation, transdifferentiation, or just differentiation? Trends Cell Biol. 2011, 21, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Pulianmackal, A.J.; Kareem, A.V.; Durgaprasad, K.; Trivedi, Z.B.; Prasad, K. Competence and regulatory interactions during regeneration in plants. Front Plant Sci. 2014, 5, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant regeneration: Cellular origins and molecular mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef] [Green Version]
- Birnbaum, K.D.; Sánchez Alvarado, A. Slicing across kingdoms: Regeneration in plants and animals. Cell 2008, 132, 697–710. [Google Scholar] [CrossRef] [Green Version]
- Dinneny, J.R.; Benfey, P.N. Plant stem cell niches: Standing the test of time. Cell 2008, 132, 553–557. [Google Scholar] [CrossRef] [Green Version]
- Trick, H.N.; Dinkins, R.D.; Santarem, E.R.; Di, R.; Samoylov, V.; Meurer, C.A.; Walker, D.R.; Parrott, W.A.; Finer, J.J.; Collins, G.B. Recent advances in soybean transformation. Plant Tissue Cult. Biotechnol. 1997, 3, 9–24. [Google Scholar]
- Jain, S.M.; Soltes, E.J.; Newton, R.J. Enhancement of somatic embryogenesis in Norway spruce (Picea abies L.). Theor. Appl. Genet. 1988, 76, 501–506. [Google Scholar] [CrossRef]
- Jain, S.M.; Dong, N.; Newton, R.J. Somatic embryogenesis in slash pine (Pinus elliottii) from immature embryo cultures in vitro. Plant Sci. 1989, 65, 233–241. [Google Scholar] [CrossRef]
- Jain, S.M. An Overview of Progress on Somatic Embryogenesis in Forest Trees. In Plant Biotechnology and In Vitro Biology in the 21st Century; Altman, A., Ziv, M., Izhar, S., Eds.; Current Plant Science and Biotechnology in Agriculture; Springer: Dordrecht, The Netherlands, 1999; Volume 36, pp. 57–63. [Google Scholar]
- Jain, S.M.; Gupta, P.K. Stepwise Protocols for Somatic Embryogenesis in Woody Plants; Springer International Publishing: Cham, Switzerland, 2018; Volume 1–2. [Google Scholar]
- Hill, K.; Schaller, G.E. Enhancing plant regeneration in tissue culture. Plant Signal Behav. 2013, 8, e25709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnyk, C.W.; Meyerowitz, E.M. Plant grafting. Curr. Biol. 2015, 25, 183–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, T.T.; Liu, H.; Shi, D.Y.; Wang, M.; Bie, X.M.; Li, X.G.; Zhang, X.S. Thioredoxin mediated ROS homeostasis explains natural variation in plant regeneration. Plant Physiol. 2018, 176, 2231–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auer, C.A.; Motyka, V.; Brezinova, A.; Kaminek, M. Endogenous cytokinin accumulation and cytokinin oxidase activity during shoot organogenesis of Petunia hybrida. Physiol. Plant. 1999, 105, 141–147. [Google Scholar] [CrossRef]
- Mercier, H.; Souza, B.M.; Kraus, J.E.; Hamasaki, R.M.; Sotta, B. Endogenous auxin and cytokinin contents associated with shoot formation in leaves of pineapple cultured in vitro. Braz. J. Plant Physiol. 2003, 15, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, M.; Novak, O.; Strnad, M.; Van Staden, J. Endogenous cytokinin in shoots of Aloe polyphylla cultured in vitro in relation to hyperhydricity, exogenous cytokinins and gelling agents. Plant Growth Regul. 2006, 50, 219–230. [Google Scholar] [CrossRef]
- Brand, M.H.; Lineberger, R.D. Micropropagation of American sweetgum (Liquidambar styraciflua L.). In Biotechnology in Agriculture and Forestry; Bajaj, Y.P.S., Ed.; High Tech and Micropropagation; Springer: Berlin, Germany, 1999; Volume 18, pp. 3–24. [Google Scholar]
- George, M.W.; Tripepi, R.R. Cytokinins, donor plants and time in culture affect shoot regenerative capacity of American elm leaves. Plant Cell Tiss Org. 1994, 39, 27–36. [Google Scholar] [CrossRef]
- Kareem, A.; Radhakrishnan, D.; Sondhi, Y.; Aiyaz, M.; Roy, M.V.; Sugimoto, K.; Prasad, K. De novo assembly of plant body plan: A step ahead of Dead pool. Regeneration 2016, 3, 182–197. [Google Scholar] [CrossRef]
- Bonga, J.M.; Von Aderkas, P. In vitro Culture of Trees; Kluwer: Dordrecht, The Netherlands, 1992; 236 p. [Google Scholar]
- Kozlowski, T.T. Physiological ecology of natural regeneration of harvested and disturbed forest stands: Implications for forest management. For. Ecol. Manag. 2002, 158, 195–221. [Google Scholar] [CrossRef]
- Mengel, K.; Kirkby, E.A. Principles of Plant Nutrition, 5th ed.; Kluwer: Dordrecht, The Netherlands, 2001; 849 p. [Google Scholar]
- Ramage, C.M.; Williams, R.R. Mineral uptake in tobacco leaf discs during different developmental stages of shoot organogenesis. Plant Cell Rep. 2003, 21, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Newton, R.J. Mevalonate kinase activity during different stage of plant regeneration from nodular callus cultures in white pine (Pinus strobus). Tree Physiol. 2005, 26, 195–200. [Google Scholar]
- Duncan, D.R.; Williams, M.T.; Zehr, B.T.; Widholm, J.M. The Production of callus capable of plant regeneration from immature embryos of numerous Zea mays genotypes. Planta 1985, 165, 322–332. [Google Scholar] [PubMed]
- Kothari, S.L.; Joshi, A.; Kachhwaha, S.; Ochoa-Alejo, N. Chilli peppers-A review on tissue culture and transgenesis. Biotechnol. Adv. 2010, 28, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Rathore, K.S.; Goldsworthy, A. Electrical control of shoot regeneration in plant tissue cultures. Bio/Technol. 1985, 3, 1107–1109. [Google Scholar]
- Danilova, S.A.; Dolgikh, Y.I. The stimulatory effect of the antibiotic cefotaxime on plant regeneration in maize tissue culture. Russ. J. Plant Physiol. 2004, 51, 559–562. [Google Scholar] [CrossRef]
- Azmi, N.S.; Ahmad, R.; Ibrahim, R. Effects of red and blue (RB) LED on the in vitro growth of Rosa kordesii in multiplication phase. 2nd International Conference on Agriculture and Biotechnology. IPCBEE 2014, 79, 20–24. [Google Scholar]
- Duclercq, J.; Sangwan-Norreel, B.; Catterou, M.; Sangwan, R.S. De novo shoot organogenesis: From art to science. Trends Plant Sci. 2011, 16, 597–606. [Google Scholar] [CrossRef]
- Terzi, M.; Lo Schiavo, F. Somatic embryogenesis. In Plant Tissue Culture: Applications and Limitations; Bhajwani, S.S., Ed.; Elsevier: Amsterdam, The Netherlands, 1990; pp. 54–66. [Google Scholar]
- Sugiyama, M. Organogenesis in vitro. Curr. Opin. Plant Biol. 1999, 2, 61–64. [Google Scholar] [CrossRef]
- Nic-Can, G.I.; Galaz-Avalos, R.M.; De-La-Pena, C.; Alcazar-Magana, A.; Wrobel, K.; Loyola-Vargas, V.M. Somatic embryogenesis: Identified factors that lead to embryogenic repression. A case of species of the same genus. PLoS ONE 2015, 10, e0126414. [Google Scholar] [CrossRef]
- Horstman, A.; Bemer, M.; Boutilier, K. A transcriptional view on somatic embryogenesis. Regeneration 2017, 4, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.G.; Maheswaran, G. Somatic embryogenesis: Factors influencing coordinated behaviour of cells as an embryogenic group. Ann. Bot. 1986, 57, 443–462. [Google Scholar]
- Raemakers, C.J.J.M.; Jacobsen, E.; Visser, R.G.F. Secondary somatic embryogenesis and applications in plant breeding. Euphytica 1995, 81, 93–107. [Google Scholar]
- Sobri, H.; Rusli, I.; Kiong, A.L.P. Somatic embryogenesis: An alternative method for in vitro micropropagation. Iran. J. Biotechnol. 2006, 4, 156–161. [Google Scholar]
- Baskaran, P.; Kumari, A.; Naidoo, D.; Van Staden, J. In vitro propagation and ultrastructural studies of somatic embryogenesis of Ledebouria ovatifolia. In Vitro Cell Dev. Biol. Plant. 2016, 52, 283–292. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Phillips, G.C. Plant cell, tissue and organ culture. In Fundam. Methods; Springer: Berlin/Heidelberg, Germany, 1995; p. 359. [Google Scholar]
- Shirani Bidabadi, S.; Sariah, M.; Zakaria, W.; Maziah, M. Study of genetic and phenotypic variability among somaclones induced by BAP and TDZ in micropropagated shoot tips of banana (Musa spp.) cultivars using RAPD markers. J. Agric. Sci. 2010, 2, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Vijayanand, K.G.; Reddy, M.P. Plant regeneration in non-toxic Jatropha curcas -impacts of plant growth regulators, source and type of explants. J. Plant Biochem. Biotechnol. 2011, 20, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Krishna, H.; Alizadeh, M.; Singh, D.; Singh, U.; Chauhan, N.; Eftekhari, M.; Sadh, R.K. Somaclonal variations and their applications in horticultural crops improvement. 3 Biotech 2016, 6, 54. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.J.; Wang, L.; Sun, W.; Zhang, Y.; Zhou, C.; Su, Y.H.; Li, W.; Sun, T.T.; Zhao, X.Y.; Li, X.G.; et al. Pattern of auxin and cytokinin responses for shoot meristem induction results from the regulation of cytokinin biosynthesis by auxin response factor. Plant Physiol. 2013, 161, 240–251. [Google Scholar]
- Maraschin, S.F.; de Priester, W.; Spaink, H.P.; Wang, M. Androgenic switch: An example of plant embryogenesis from the male gametophyte perspective. J. Exp. Bot. 2005, 56, 1711–1726. [Google Scholar] [CrossRef]
- Chupeau, M.C.; Granier, F.; Pichon, O.; Renou, J.P.; Gaudin, V.; Chupeau, Y. Characterization of the early events leading to totipotency in Arabidopsis protoplast liquid culture by temporal transcript profiling. Plant Cell. 2013, 25, 2444–2463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roest, S.; Gilissen, L.J.W. Plant regeneration from protoplasts: A literature review. Acta Bot. Neerl. 1989, 38, 1–23. [Google Scholar] [CrossRef]
- Abdallah, N.A.; Prakash, C.S.; McHughen, A.G. Genome editing for crop improvement: Challenges and opportunities. GM Crops Food 2015, 6, 183–205. [Google Scholar] [PubMed] [Green Version]
- Ochatt, S.J.; Pontecaille, C.; Rancillac, M. The growth regulators used for bud regeneration and shoot rooting affect the competence for flowering and seed set in regenerated plants of protein peas. In Vitro Cell. Dev. Biol. Plant 2000, 36, 188–193. [Google Scholar]
- Pan, Z.G.; Liu, C.Z.; Murch, S.J.; El-Demerdash, M.; Saxena, P.K. Plant regeneration from mesophyll protoplasts of the Egyptian medicinal plants Artemisia judaica L. and Echinops spinosissimus Turra. Plant Sci. 2003, 165, 681–687. [Google Scholar] [CrossRef]
- Wang, Y.H.; Bhalla, P.L. Somatic embryogenesis from leaf explants of Australian fan flower, Scaevola aemula R. Br. Plant Cell Rep. 2004, 22, 408–414. [Google Scholar] [CrossRef]
- Chung, H.H.; Chen, J.T.; Chang, W.C. Plant regeneration through direct somatic embryogenesis from leaf explants of Dendrobium. Biol. Plant. 2007, 51, 346–350. [Google Scholar] [CrossRef]
- Iantcheva, A.; Slavov, S.; Prinsen, E.; Vlahova, M.; van Onckelen, H.; Atan-assov, A. Embryo induction and regeneration from root explants of Medicago truncatula after osmotic pre-treatment. Plant Cell Tissue Organ Cult. 2005, 81, 37–43. [Google Scholar]
- Kim, T.D.; Lee, B.S.; Kim, T.S.; Choi, Y.E. Developmental plasticity of glandular trichomes into somatic embryogenesis in Tilia amurensis. Ann. Bot. 2007, 100, 177–183. [Google Scholar]
- Soriano, M.; Li, H.; Boutilier, K. Microspore embryogenesis: Establishment of embryo identity and pattern in culture. Plant Reprod. 2013, 26, 181–196. [Google Scholar]
- Chen, J.T.; Hong, P.I. Cellular origin and development of secondary somatic embryos in Oncidium leaf cultures. Biol. Plant. 2012, 56, 215–220. [Google Scholar] [CrossRef]
- Hamidah, M.; Abdul Karim, A.G.; Debergh, P. Somatic embryogenesis and plant regeneration in Anthurium scherzerianum. Plant Cell Tissue Organ Cult. 1997, 48, 189–193. [Google Scholar] [CrossRef]
- Yantcheva, A.; Vlahova, M.; Antanassov, A. Direct somatic embryogenesis and plant regeneration of carnation (Dianthus caryophyllus L.). Plant Cell Rep. 1998, 18, 148–153. [Google Scholar] [CrossRef]
- Sobri, H.; Rusli, I.; Anna, L.P.K. Adventitious shoots regeneration from root and stem explants of Eurycoma longifolia Jack-An important tropical medicinal plant. Int. J. Agric. Res. 2006, 1, 183–193. [Google Scholar]
- Prasad, V.S.S.; Gupta, S.D. In vitro shoot regeneration of gladiolus in semi-solid agar versus liquid cultures with support systems. Plant Cell Tissue Organ Cult. 2006, 87, 263–271. [Google Scholar] [CrossRef]
- Ibaraki, Y.; Kurata, K. Automation of somatic embryo production. Plant Cell Tissue Organ Cult. 2001, 65, 179–199. [Google Scholar] [CrossRef]
- Korban, S.S. Somatic embryogenesis in Rose: Gene expression and genetic transformation. In Somatic Embryogenesis: Plant Cell Monographs; Mujib, A., Smaaj, J., Eds.; Springer: New York, NY, USA; Berlin, Germany, 2005; pp. 247–257. [Google Scholar]
- Steward, F.C.; Mapes, M.O.; Mears, K. Growth and organized development of cultured cells. II. Organization in cultures grown from freshly suspended cells. Am. J. Bot. 1958, 45, 705–708. [Google Scholar] [CrossRef]
- Matand, K.; Shoemake, M.; Li, C. High frequency in vitro regeneration of adventitious shoots in daylilies (Hemerocallis sp) stem tissue using thidiazuron. BMC Plant Biol. 2020, 20, 31. [Google Scholar] [CrossRef] [Green Version]
- Thomson, G.E.; Deering, T.D. Effect of cytokinin type and concentration on in vitro shoot proliferation of hazelnut (Corylus avellana L.). N. Z. J. Crop Hort. 2011, 39, 209–213. [Google Scholar] [CrossRef]
- Xiao, Y.; Niu, G.; Kozai, T. Development and application of photoautotrophic micropropagation plant system. Plant Cell Tissue Org. 2010, 105, 149–158. [Google Scholar] [CrossRef]
- Kozai, T.; Afreen, F.; Zobayed, S.M.A. Photoautotrophic (Sugar-Free Medium) Micropropagation as a New Propagation and Transplant Production System; Springer: Dordrecht, The Netherlands; Cham, Switzerland, 2005; p. 316. [Google Scholar]
- Kozai, T. Photoautotrophic micropropagation. In Vitro Cell Dev. Biol. 1991, 27, 47–51. [Google Scholar]
- Manivannan, A.; Soundararajan, P.; Halimah, N.; Ko, C.H. Blue LED light enhances growth, phytochemical contents, and antioxidant enzyme activities of Rehmannia glutinosa cultured in vitro. Hort. Environ. Biotechnol. 2015, 56, 105–113. [Google Scholar] [CrossRef]
- Gupta, D.S.; Jatothu, B. Fundamentals and applications of light-emitting diodes (LEDs) in in vitro plant growth and morphogenesis. Plant Biotechnol. Rep. 2013, 7, 211–220. [Google Scholar] [CrossRef]
- Bello-Bello, J.J.; Perez-Sato, J.A.; Cruz-Cruz, C.A.; Martinez-Estrada, E. Light-emitting diodes: Progress in plant micropropagation. InTech 2017, 6, 93–103. [Google Scholar]
- Swanepoel, J.; Chikuni, E.; De Merwe, H.V. Light emitting diode vs fluorescent considerations inpublic and commercial lighting applications. In Proceedings of the ACRID’17: EAI International Conference on Research, Innovation and Development for Africa, Victoria Falls, Zimbabwe, 20–21 June 2017; pp. 182–190. [Google Scholar]
- Miler, N.; Kulus, D.; Wozny, A.; Rymarz, D.; Hajzer, M.; Wierzbowski, K.; Nelke, R.; Szeffs, L. Application of wide-spectrum light-emitting diodes in micropropagation of popular ornamental plant species: A study on plant quality and cost reduction. In Vitro Cell. Dev. Biol. Plant. 2019, 55, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Mosqueda, M.A.; Iglesias-Andreu, L.G.; Luna-Sanchez, I.J. Light quality affects growth and development of in vitro plantlet of Vanilla planifolia Jacks. S. Afr. J. Bot. 2017, 109, 288–293. [Google Scholar]
- Xu, L.; Huang, H. Genetic and epigenetic controls of plant regeneration. Curr. Top Dev. Biol. 2014, 108, 1–33. [Google Scholar]
- Ikeuchi, M.; Sugimoto, K.; Iwase, A. Plant Callus: Mechanisms of induction and repression. Plant Cell 2013, 25, 3159–3173. [Google Scholar] [CrossRef] [Green Version]
- Ikeuchi, M.; Iwase, A.; Rymen, B.; Lambolez, A.; Kojima, M.; Takebayashi, Y.; Heyman, J.; Watanabe, S.; Seo, M.; De Veylder, L.; et al. Wounding Triggers Callus Formation via Dynamic Hormonal and Transcriptional Changes. Plant Physiol. 2017, 175, 1158–1174. [Google Scholar]
- Nuruzzaman, M.; Sharoni, A.; Kikuchi, S. Roles of NAC transcription factors in the regulation of biotic and abiotic stress responses in plants. Front Microbiol. 2013, 4, 248. [Google Scholar]
- Chen, L.; Sun, B.; Xu, L.; Liu, W. Wound signaling: The missing link in plant regeneration. Plant Signal Behav. 2016, 11, e1238548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.; Schmid, M.W.; Grossniklaus, U. Plant germline formation: Common concepts and developmental flexibility in sexual and asexual reproduction. Development 2015, 142, 229–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feher, A. Somatic embryogenesis—Stress-induced remodeling of plant cell fate. Biochim. Biophys. Acta 2015, 1849, 385–402. [Google Scholar] [CrossRef] [PubMed]
- Lotan, T.; Ohto, M.; Yee, K.M.; West, M.A.; Lo, R.; Kwong, R.W.; Yamagishi, K.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. Arabidopsis LEAFY COTYLEDON1 is sufficient to induce embryo development in vegetative cells. Cell 1998, 93, 1195–1205. [Google Scholar] [CrossRef] [Green Version]
- Hecht, V.; Vielle-Calzada, J.P.; Hartog, M.V.; Schmidt, E.D.; Boutilier, K.; Grossniklaus, U.; de Vries, S.C. The Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR KINASE 1 gene is expressed in developing ovules and embryos and enhances embryogenic competence in culture. Plant Physiol. 2001, 127, 803–816. [Google Scholar] [CrossRef]
- Stone, S.L.; Kwong, L.W.; Yee, K.M.; Pelletier, J.; Lepiniec, L.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. LEAFY COTYLEDON2 encodes a B3 domain transcription factor that induces embryo development. Proc. Natl. Acad. Sci. USA 2001, 98, 11806–11811. [Google Scholar] [CrossRef] [Green Version]
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.; Hattori, J.; Liu, C.M.; van Lammeren, A.A.M.; Miki, B.L.A.; et al. Ectopic expression of BABY BOOM triggers a conversion from vegetative to embryonic growth. Plant Cell 2002, 14, 1737–1749. [Google Scholar]
- Zuo, J.; Niu, Q.W.; Frugis, G.; Chua, N.H. The WUSCHEL gene promotes vegetative-to-embryonic transition in Arabidopsis. Plant J. 2002, 30, 349–359. [Google Scholar]
- Harding, E.W.; Tang, W.; Nichols, K.W.; Fernandez, D.E.; Perry, S.E. Expression and maintenance of embryogenic potential is enhanced through constitutive expression of AGAMOUS-Like 15. Plant Physiol. 2003, 133, 653–663. [Google Scholar] [CrossRef] [Green Version]
- Thakare, D.; Tang, W.; Hill, K.; Perry, S.E. The MADS-domain transcriptional regulator AGAMOUS-LIKE15 promotes somatic embryo development in Arabidopsis and soybean. Plant Physiol. 2008, 146, 1663–1672. [Google Scholar] [CrossRef] [Green Version]
- Tsuwamoto, R.; Yokoi, S.; Takahata, Y. Arabidopsis EMBRYOMAKER encoding an AP2 domain transcription factor plays a key role in developmental change from vegetative to embryonic phase. Plant Mol. Biol. 2010, 73, 481–492. [Google Scholar] [PubMed]
- Mozgova, I.; Munoz-Viana, R.; Hennig, L. PRC2 represses hormone-induced somatic embryogenesis in vegetative tissue of Arabidopsis thaliana. PLoS Genet. 2017, 13, e1006562. [Google Scholar]
- Xiao, J.; Wagner, D. Polycomb repression in the regulation of growth and development in Arabidopsis. Curr. Opin. Plant. Biol. 2015, 23, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Taha, R.M.; Wafa, S.N. Plant regeneration and cellular behaviour studies in Celosia cristata grown in vivo and in vitro. Sci. World J. 2012, 359413, 1–8. [Google Scholar]
- Mironova, V.; Xu, J. A single-cell view of tissue regeneration in plants. Curr. Opin. Plant. Biol. 2019, 52, 149–154. [Google Scholar] [PubMed]
- Vriz, S.; Reiter, S.; Galliot, B. Cell death: A program to regenerate. Curr. Top Dev. Biol. 2014, 108, 121–151. [Google Scholar]
- Johnson, R.A.; Conklin, P.A.; Tjahjadi, M.; Missirian, V.; Toal, T.; Brady, S.M.; Britt, A.B. suppressor of gamma response1 links DNA damage response to organ regeneration. Plant Physiol. 2018, 176, 1665–1675. [Google Scholar] [CrossRef] [Green Version]
- Heyman, J.; Cools, T.; Canher, B.; Shavialenka, S.; Traas, J.; Vercauteren, I.; Van den Daele, H.; Persiau, G.; De Jaeger, G.; Sugimoto, K.; et al. The heterodimeric transcription factor complex ERF115-PAT1 grants regeneration competence. Nat. Plants. 2016, 2, 16165. [Google Scholar]
- Marhava, P.; Hoermayer, L.; Yoshida, S.; Marhavy, P.; Benkova, E.; Friml, J. Re-activation of stem cell pathways for pattern restoration in plant wound healing. Cell 2019, 177, 957–969. [Google Scholar] [CrossRef] [Green Version]
- Louveaux, M.; Julien, J.D.; Mirabet, V.; Boudaoud, A.; Hamant, O. Cell division plane orientation based on tensile stress in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2016, 113, E4294–E4303. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, M. Historical review of research on plant cell dedifferentiation. J. Plant Res. 2015, 128, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.N.; Burke, Z.D.; Tosh, D. Transdifferentiation, metaplasia and tissue regeneration. Organogenesis 2004, 1, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.; McCurdy, D.W. Transdifferentiation–a plant perspective. In Molecular Cell Biology of the Growth and Differentiation of Plant Cells; CRC Press: Boca Raton, FL, USA, 2016; pp. 298–319. [Google Scholar]
- Sanges, D.; Cosma, M.P. Reprogramming cell fate to pluripotency: The decision-making signalling pathways. Int. J. Dev. Biol. 2010, 54, 1575–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdeil, J.L.; Alemanno, L.; Niemenak, N.; Tranbarger, T.J. Pluripotent versus totipotent plant stem cells: Dependence versus autonomy? Trends Plant Sci. 2007, 12, 245–252. [Google Scholar] [PubMed]
- Leon, J.; Rojo, E.; Sanchez-Serrano, J.J. Wound signaling in plants. J. Exp. Bot. 2001, 52, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E.; Mithofer, A.; Boland, W. Before gene expression: Early events in plant-insect interaction. Trends Plant Sci. 2007, 12, 310–316. [Google Scholar]
- Iwase, A.; Mitsuda, N.; Koyama, T.; Hiratsu, K.; Kojima, M.; Arai, T.; Inoue, Y.; Seki, M.; Sakakibara, H.; Sugimoto, K.; et al. The AP2/ERF transcription factor WIND1 controls cell dedifferentiation in Arabidopsis. Curr. Biol. 2011, 21, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Tong, J.; Xiao, L.; Ruan, Y.; Liu, J.; Zeng, M.; Huang, H.; Wang, J.W.; Xu, L. YUCCA-mediated auxin biogenesis is required for cell fate transition occurring during de novo root organogenesis in Arabidopsis. J. Exp. Bot. 2016, 67, 4273–4284. [Google Scholar] [CrossRef] [Green Version]
- Lup, S.D.; Tian, X.; Xu, J.; Perez-Perez, J.M. Wound signaling of regenerative cell reprogramming. Plant Sci. 2016, 250, 178–187. [Google Scholar]
- Liu, J.; Sheng, L.; Xu, Y.; Li, J.; Yang, Z.; Huang, H.; Xu, L. WOX11 and 12 are involved in the first-step cell fate transition during de novo root organogenesis in Arabidopsis. Plant Cell 2014, 26, 1081–1093. [Google Scholar] [CrossRef] [Green Version]
- Rymen, B.; Kawamura, A.; Lambolez, A.; Inagaki, S.; Takebayashi, A.; Iwase, A.; Sakamoto, Y.; Sko, K.; Favero, D.S.; Ikeuchi, M.; et al. Histone acetylation orchestrates wound-induced transcriptional activation and cellular reprogramming in Arabidopsis. Commun. Biol. 2019, 2, 404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, P.; Lall, S.; Nettelton, D.; Howell, S.H. Gene Expression Programs during Shoot, Root, and Callus Development in Arabidopsis Tissue Culture. Plant Physiol. 2006, 141, 620–637. [Google Scholar] [CrossRef] [Green Version]
- Haberer, G.; Kieber, j.j. Cytokinins. New insights into a classic phytohormone. Plant Physiol. 2002, 128, 354–362. [Google Scholar] [PubMed]
- Vollbrecht, E.; Veit, B.; Sinha, N.; Hake, S. The developmental gene Knotted-1 is a member of a maize homeobox gene family. Nature 1991, 350, 241–243. [Google Scholar] [PubMed]
- Mayer, K.F.; Schoof, H.; Haecker, A.; Lenhard, M.; Jurgens, G.; Laux, T. Role of WUSCHEL in Regulating Stem Cell Fate in the Arabidopsis Shoot Meristem. Cell 1998, 95, 805–815. [Google Scholar] [PubMed] [Green Version]
- Fletcher, J.C.; Brand, U.; Running, M.P.; Simon, R.; Meyerowitz, E.M. Communication of cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science 1999, 283, 1911–1914. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Lemaux, P.G. Molecular analysis of in vitro shoot organogenesis. Crit. Rev. Plant Sci. 2004, 23, 325–335. [Google Scholar] [CrossRef]
- Riou-Khamlichi, C.; Huntley, R.; Jacqmard, A.; Murray, J.A.H. Cytokinin activation of Arabidopsis cell division through a D-type cyclin. Science 1999, 283, 1541–1544. [Google Scholar] [CrossRef] [Green Version]
- Rupp, H.M.; Frank, M.; Werner, T.; Strnad, M.; Schmuelling, T. Increased steady state mRNA levels of the STM and KNAT1 homeobox genes in cytokinin overproducing Arabidopsis thaliana indicate a role for cytokinins in the shoot apical meristem. Plant J. 1999, 18, 557–563. [Google Scholar] [CrossRef]
- Kareem, A.; Durgaprasad, K.; Sugimoto, K.; Du, Y.; Pulianmackal, A.J.; Trivedi, Z.B.; Abhayadev, P.V.; Pinon, V.; Meyerowitz, E.M.; Sceres, B.; et al. PLETHORA genes control regeneration by a two-step mechanism. Curr. Biol. 2015, 25, 1017–1030. [Google Scholar]
- Gordon, S.P.; Heisler, M.G.; Reddy, G.V.; Ohno, C.; Das, P.; Meyerowitz, E.M. Pattern formation during de novo assembly of the Arabidopsis shoot meristem. Development 2007, 134, 3539–3548. [Google Scholar] [PubMed] [Green Version]
- Motte, H.; Vercauteren, A.; Depuydt, S.; Landschoot, S.; Geelen, D.; Werbrouck, S.; Goormachtig, S.; Vuylsteke, M.; Vereecke, D. Combining linkage and association mapping identifies RECEPTOR-LIKE PROTEIN KINASE1 as an essential Arabidopsis shoot regeneration gene. Proc. Natl. Acad. Sci. USA 2014, 111, 8305–8310. [Google Scholar] [PubMed] [Green Version]
- Chatfield, S.P.; Capron, R.; Severino, A.; Penttila, P.A.; Alfred, S.; Nahal, H.; Provart, N.J. Incipient stem cell niche conversion in tissue culture: Using a systems approach to probe early events in WUSCHEL-dependent conversion of lateral root primordia into shoot meristems. Plant J. 2013, 73, 798–813. [Google Scholar] [CrossRef] [PubMed]
- Kirch, T.; Simon, R.; Werr, W. The DORNRÖSCHEN/ENHANCER OF SHOOTREGENERATION1 gene of arabidopsis acts in the control of meristem cell fate and lateral organ development. Plant Cell 2003, 15, 694–705. [Google Scholar] [CrossRef] [Green Version]
- Goh, T.; Joi, S.; Mimura, T.; Fukaki, H. The establishment of asymmetry in Arabidopsis lateral root founder cells is regulated by LBD16/ASL18 and related LBD/ASL proteins. Development 2012, 139, 883–893. [Google Scholar] [CrossRef] [Green Version]
- Libik, M.; Konieczny, R.; Pater, B.; Slesak, I.; Miszalski, Z. Differences in the activities of some antioxidant enzymes and in H2O2 content during rhizogenesis and somatic embryogenesis in callus cultures of the ice plant. Plant Cell Rep. 2005, 23, 834–841. [Google Scholar] [CrossRef]
- VijendrA, P.D.; Jayanna, S.G.; Kumar, V.; Gajula, H.; Rajashekar, J.; Sannabommaji, T.; Basappa, G. Antioxidant status during in vitro plant regeneration in Lucas aspera Spreng. Int. J. Pure App. Biosci. 2017, 5, 1529–1537. [Google Scholar] [CrossRef]
- Gechev, T.S.; Van Breusegem, F.; Stone, J.M.; Denev, I.; Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays 2006, 28, 1091–1101. [Google Scholar]
- Faltin, Z.; Holland, D.; Velcheva, M.; Tsapovetsky, M.; Roeckel-Drevet, P.; Handa, A.K.; Abu-Abied, M.; Friedman-Einat, M.; Eshdat, Y.; Perl, A. Glutathione peroxidase regulation of reactive oxygen species level is crucial for in vitro plant differentiation. Plant Cell Physiol. 2010, 51, 1151–1162. [Google Scholar] [CrossRef] [Green Version]
- Krishna, H.; Singh, D. Micropropagation of lasora (Cordia myxa Roxb.). Indian J. Hortic. 2013, 70, 323–327. [Google Scholar]
- Bairu, M.W.; Aremu, A.O.; VanStaden, J. Somaclonal variation in plants: Causes and detection methods. Plant Growth Regul. 2011, 63, 147–173. [Google Scholar] [CrossRef]
- Currais, L.; Loureiro, J.; Santos, C.; Canhoto, J.M. Ploidy stability in embryogenic cultures and regenerated plantlets of tamarillo. Plant Cell Tiss Org. 2013, 114, 149–159. [Google Scholar] [CrossRef]
- Shirani, S.; Mahdavi, F.; Maziah, M. Morphological abnormality among regenerated shoots of banana and plantain (Musa spp.) after in vitro multiplication with TDZ and BAP from excised shoot tips. Afr. J. Biotechnol. 2009, 8, 5755–5761. [Google Scholar]
- Sahijram, L.; Soneji, J.; Bollamma, K. Analyzing somaclonal variation in micropropagated bananas (Musa spp.). In Vitro Cell Dev. Biol. Plant 2003, 39, 551–556. [Google Scholar] [CrossRef]
- Chen, Z.; Ahuja, M.R. Regeneration and genetic variation in plant tissue cultures. In Clonal Forestry; Ahuja, M.R., Libby, W.J., Eds.; Genetics and Biotechnology; Springer: Berlin/Heidelberg, Germany, 1993; Volume 1, pp. 87–100. [Google Scholar]
- Larkin, P.J.; Scowcroft, W.R. SomacIonal variation -a novel source of variability from cell cultures for plant improvement. Theor. Appl. Genet. 1981, 60, 197–214. [Google Scholar] [CrossRef]
- Mano, Y.; Komatsuda, T. Identification of QTLs controlling tissue-culture traits in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2002, 105, 708–715. [Google Scholar] [CrossRef]
- Trujillo-Moya, C.; Gisbert, C.; Vilanova, S.; Nuez, F. Localization of QLTs for in vitro plant regeneration in tomato. BMC Plant Biol. 2011, 11, 140. [Google Scholar] [CrossRef] [Green Version]
- Joyce, S.M.; Cassells, A.C.; Jain, S.M. Stress and aberrant phenotypes in vitro culture. Plant Cell Tissue Organ Cult. 2003, 74, 103–121. [Google Scholar] [CrossRef]
- Sato, K.; Motoi, Y.; Yamaji, N.; Yoshida, H. 454 sequencing of pooled BAC clones on chromosome 3H of barley. BMC Genom. 2011, 12, 246–252. [Google Scholar] [CrossRef] [Green Version]
- Smulders, M.J.M.; De Klerk, G.J. Epigenetics in plant tissue culture. Plant Growth Regul. 2011, 63, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Tanurdzic, M.; Vaughn, M.W.; Jiang, H.; Lee, T.J.; Slotkin, R.K.; Sosinski, B.; Thompson, W.F.; Doerge, R.W.; Martienssen, R.A. Epigenomic consequences of immortalized plant cell suspension culture. PLoS Biol. 2008, 6, 2880–2895. [Google Scholar] [PubMed]
- Nivas, S.K.; Dsouza, L. Genetic fidelity in micropropagated plantlets of Anacardium occidentale L. (Cashew) an important fruit tree. Int. J. Sci. Res. 2014, 3, 2142–2146. [Google Scholar]
- Duncan, R.R. Tissue culture-induced variation in crop improvement. Adv. Agron. 1997, 58, 201–240. [Google Scholar]
- Karp, A. Origins, Causes and Uses of Variation in Plant Tissue Cultures. In Plant Cell and Tissue Culture; Vasil, I.K., Thorpe, T.A., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 139–151. [Google Scholar]
- Zayova, E.; Vassilevska, I.R.; Kraptchev, B.; Stoeva, D. Somaclonal variations through indirect organogenesis in eggplant (Solanum melongena L.). Biol. Divers Conserv. 2010, 3, 1–5. [Google Scholar]
- Farahani, F.; Yari, R.; Masoud, S. Somaclonal variation in Dezful cultivar of olive (Olea europaea subsp. europaea). Gene Conserve. 2011, 10, 216–221. [Google Scholar]
- Sun, L.; Yang, H.; Chen, M.; Ma, D.; Lin, C. RNA-seq reveals dynamic changes of gene expression in key stages of intestine regeneration in the sea cucumber Apostichopus japonicus. PLoS ONE 2013, 8, e69441. [Google Scholar]
- Rival, A.; Ilbert, P.; Labeyrie, A.; Torres, E.; Doulbeau, S.; Personne, A.; Dussert, S.; Beulé, T.; Durand-Gasselin, T.; Tregear, J.; et al. Variations in genomic DNA methylation during the long-term in vitro proliferation of oil palm embryogenic suspension cultures. Plant Cell Rep. 2013, 32, 359–368. [Google Scholar] [CrossRef]
- Mohan Jain, S. Creation of variability by mutation and tissue culture for improving plants. Acta Hortic. 1997, 447, 69–78. [Google Scholar]
- Jain, S.M.; Brar, D.S.; Ahloowalia, B.S. Somaclonal Variations and Induced Mutations in Crop Improvement; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; Volume 32, p. 620. [Google Scholar]
- Jain, S.M. Tissue culture-derived variation in crop improvement. Euphytica 2001, 118, 153–166. [Google Scholar]
- Nwauzoma, A.B.; Jaja, E.T. A review of somaclonal variation in plantain (Musa spp.): Mechanisms and applications. J. Appl. Biosci. 2013, 67, 5252–5260. [Google Scholar] [CrossRef] [Green Version]
- Bello-Bello, J.J.; Iglesias-Andreu, L.G.; Avilés-Viñas, S.A.; Gómez-Uc, E.; Canto-Flick, A.; Santana-Buzzy, N. Somaclonal variation in habanero pepper (Capsicum chinense Jacq.) as assessed ISSR molecular markers. HortScience 2014, 49, 481–485. [Google Scholar]
- Matsuda, S.; Sato, M.; Ohno, S. Cutting leaves and plant growth regulator application enhance somaclonal variation induced by transposition of VGs1 of Saintpaulia. J. Jpn. Soc. Hortic. Sci. 2014, 83, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, T.C.R.; Carvalho, C.R. Vertical heterogeneity of DNA ploidy level assessed by flow cytometry in calli of Passiflora cincinnata. In Vitro Cell Dev. Biol. Plant. 2014, 50, 158–165. [Google Scholar] [CrossRef]
- Leva, A.R.; Petruccelli, R.; Rinaldi, L.M.R. Somaclonal variation in tissue culture: A case study with olive. In Recent Advances in Plant in Vitro Culture; Leva, A.R., Rinaldi, L.M.R., Eds.; INTECH Open Access Publisher: Croatia, Balkans, 2012; pp. 123–150. [Google Scholar]
- Gady, A.L.F.; Herman, F.W.K.; Van de Wal, M.H.B.J.; Visser, R.G.F.; Bachem, C.W.B. Implementation of two high through-put techniques in a novel application: Detecting point mutations in large EMS mutated plant populations. Plant Methods 2009, 5, 13. [Google Scholar] [PubMed] [Green Version]
- Alvarez, M.E.; Nota, F.; Cambiagno, D.A. Epigenetic control of plant immunity. Mol. Plant Pathol. 2010, 11, 563–576. [Google Scholar]
- Tiwari, J.K.; Poonam Sarkar, D.; Pandey, S.K.; Gopal, J.; Kumar, S.R. Molecular and morphological characterization of somatic hybrids between Solanum tuberosum L. and S. etuberosum Lindl. Plant Cell Tissue Organ 2010, 103, 175–187. [Google Scholar]
- Gornikiewicz, B.; Ronowicz, A.; Podolak, J.; Modanecki, P.; Stanislawska-Sachadyn, A.; Sachadyn, P. Epigenetic basis of regeneration: Analysis of genomic DNA methylation profiles in the MRL/MpJ Mouse. DNA Res. 2013, 20, 605–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birnbaum, K.D.; Roudier, F. Epigenetic memory and cell fate reprogramming in plants. Regeneration 2017, 4, 15–20. [Google Scholar] [PubMed]
- Miguel, C.; Marum, L. An epigenetic view of plant cells cultured in vitro: Somaclonal variation and beyond. J. Exp. Bot. 2011, 62, 3713–3725. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Seo, P.J. Dynamic epigenetic changes during plant regeneration. Trends Plant Sci. 2018, 23, 235–247. [Google Scholar]
- Vogel, G. How does a single somatic cell become a whole plant? Science 2005, 309, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Klerk, G.J.; Arnholdt-Schmit, B.; Lieberei, R. Regeneration of roots, shoots and embryo: Physiological, biochemical and molecular aspects. Biol. Plantarum. 1997, 39, 53–66. [Google Scholar] [CrossRef]
- Kaul, K.; Sabharwal, P.S. Morphogenetic studies on Haworthia: Establishment of tissue culture and control of differentiation. Am. J. Bot. 1972, 59, 377–385. [Google Scholar] [CrossRef]
- Ogihara, Y.T.K. Tissue culture in Haworthia. I. Effects of auxins and kinetin on callus growth. Bot. Mag. Jpn. 1978, 91, 83–91. [Google Scholar] [CrossRef]
- Chen, Y.M.; Huang, J.Z.; Hou, T.W.; Pan, I.C. Effects of light intensity and plant growth regulators on callus proliferation and shoot regeneration in the ornamental succulent Haworthia. Bot. Stud. 2019, 60, 10. [Google Scholar] [CrossRef]
- Vidal, N.; Arellano, G.; San-Jose, M.C.; Vieitez, A.M.; Ballester, A. Developmental stages during the rooting of in-vitro-cultured Quercus robur shoots from material of juvenile and mature origin. Tree Physiol. 2003, 23, 1247–1254. [Google Scholar] [CrossRef]
- Becerra, D.C.; Forero, A.P.; Gongora, G.A. Age and physiological condition of donor plants affect in vitro morphogenesis in leaf explants of Passiflora edulis f. flavicarpa. Plant Cell Tissue Organ 2004, 79, 87–90. [Google Scholar] [CrossRef]
- Zhang, T.Q.; Lian, H.; Tang, H.; Dolezal, K.; Zhou, C.M.; Yu, S.; Chen, J.H.; Chen, Q.; Liu, H.; Ljung, K.; et al. An intrinsic microRNA timer regulates progressive decline in shoot regenerative capacity in plants. Plant Cell 2015, 27, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.W.; Weigel, D.; Poethig, R.S. The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef] [Green Version]
- Dou, H.; Niu, G.; Gu, M.; Masabni, J.G. Effects of light quality on growth and phytonutrient accumulation of herbs under controlled Environments. Horticulturae 2017, 3, 36. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, J.; Wang, X.; Zhao, B.; Wang, Y. Light intensity and spectral quality influencing the callus growth of Cistanche deserticola and biosynthesis of phenylethanoid glycosides. Plant Sci. 2003, 165, 657–661. [Google Scholar] [CrossRef]
- Meziani, R.; Jaiti, F.; Mazri, M.A.; Anjarne, M.; Chitt, M.A.; El Fadile, J.; Alem, C. Effects of plant growth regulators and light intensity on the micropropagation of date palm (Phoenix dactylifera L.) cv. Mejhoul. J. Crop Sci. Biotechnol. 2015, 18, 325–331. [Google Scholar] [CrossRef]
- Farhadi, N.; Panahandeh, J.; Azar, A.M.; Salte, S.A. Effects of explant type, growth regulators and light intensity on callus induction and plant regeneration in four ecotypes of Persian shallot (Allium hirtifolium). Sci. Hortic. 2017, 218, 80–86. [Google Scholar] [CrossRef]
- Stavang, J.A.; Moe, R.; Olsen, J.E.; Junttila, O. Differential temperature regulation of GA metabolism in light and darkness in pea. J. Exp. Bot. 2007, 58, 3061–3069. [Google Scholar] [CrossRef] [PubMed]
- Delporte, F.; Pretova, A.; Du Jardin, P.; Watillon, B. Morpho-histology and genotype dependence of in vitro morphogenesis in mature embryo cultures of wheat. Protoplasma 2014, 251, 1455–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miroshnichenko, D.; Chaban, I.; Chernobrovkina, M.; Dolgov, S. Protocol for efficient regulation of in vitro morphogenesis in einkorn (Triticum monococcum L.), a recalcitrant diploid wheat species. PLoS ONE 2017, 12, 1–23. [Google Scholar] [CrossRef]
- Malik, K.; Birla, D.; Yadav, H.; Sainger, M.; Chaudhary, D.; Jaiwal, P.K. Evaluation of carbon sources, gelling agents, growth hormones and additives for efficient callus induction and plant regeneration in indian wheat (Triticum aestivum L.) genotypes using mature embryos. J. Crop Sci. Biotech. 2017, 20, 185–192. [Google Scholar] [CrossRef]
- Singh, C.K.; Raj, S.R.; Jaiswar, P.S.; Patil, V.R.; Punwar, B.S.; Chavda, J.C.; Subhash, N. Effect of plant growth regulators on in vitro plant regeneration of sandalwood (Santalum album L.) via organogenesis. Agroforest Syst. 2015, 90, 281–288. [Google Scholar] [CrossRef]
- Pumisutapon, P.; Visser, R.G.F.; Jan de Klerk, G. Moderate abiotic stresses increase rhizome growth and outgrowth of axillary buds in Alstroemeria cultured in vitro. Plant Cell Tissue Organ 2012, 110, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Puijalon, S.; Piola, F.; Bornette, G. Abiotic stresses increase plant regeneration ability. Evol. Ecol. 2008, 22, 493–506. [Google Scholar] [CrossRef]
- Jose, S.; Thomas, T.D. Abiotic stresses increase plant regeneration ability of rhizome explants of Curcuma caesia Roxb. Plant Cell Tissue Organ 2015, 122, 767–772. [Google Scholar] [CrossRef]
- Ravindran, P.N.; Nirmal Babu, K.; Shiva, K.N. Botany and crop improvement of turmeric. In Turmeric: The Genus Curcuma; Ravindran, P.N., Nirmal Babu, K., Sivaraman, K., Eds.; CRC press: London, UK, 2007; pp. 15–70. [Google Scholar]
- Kim, D.H.; Gopal, J.; Iyyakkannu Sivanesan, I. Nanomaterials in plant tissue culture: The disclosed and undisclosed. RSC Adv. 2017, 7, 36492–36505. [Google Scholar] [CrossRef] [Green Version]
- Stanisic, M.; Ninkovic, S.; Savic, J.; Cosic, T.; Mitic, N. The effects of β-lactam antibiotics and hygromycin B on de novo shoot organogenesis in apple cv. Golden Delicious. Arch. Biol. Sci. 2018, 70, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.; Wu, R.; Han, Y.; Wan, T.; Cai, Y. Optimizing suitable antibiotics for bacterium control in micropropagation of cherry rootstock using a modified leaf disk diffusion method and E test. Plants 2019, 8, 66. [Google Scholar] [CrossRef] [Green Version]
- Abdi, G.; Salehi, H.; Khosh-Khui, M. Nano silver: A novel nanomaterial for removal of bacterial contaminants in valerian (Valeriana officinalis L.) tissue cultures. Acta Physiol. Plant 2008, 30, 709–714. [Google Scholar] [CrossRef]
- Qin, Y.H.; Teixeira da Silva, J.A.; Bi, J.H.; Zhang, S.L.; Hu, G.B. Response of in vitro strawberry to antibiotics. Plant Growth Regul. 2011, 65, 183–193. [Google Scholar] [CrossRef]
- Mittal, P.; Gosal, S.S.; Senger, A.; Kumar, P. Impact of cefotaxime on somatic embryogenesis and shoot regeneration in sugarcane. Physiol. Mol. Biol. Plant 2009, 15, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Shehata, A.M.; Wannarat, W.; Skirvin, R.M.; Norton, M.A. The dual role of carbenicillin in shot regeneration and somatic embryogenesis of horseradish (Armoracia rusticana) in vitro. Plant Cell Tissue Organ 2010, 102, 397–402. [Google Scholar] [CrossRef]
- Grzebelus, E.; Skop, L. Effect of β-lactam antibiotics on plant regeneration in carrot protoplast cultures. In Vitro Cell Dev. Biol. Plant 2014, 50, 568–575. [Google Scholar] [CrossRef] [Green Version]
- Tambarussi, E.V.; Rogalski, M.; Nogueira, F.T.S.; Brondani, G.E.; De Martin, V.F.; Carrer, H. Influence of antibiotics on indirect organogenesis of Teak. Ann. For. Res. 2015, 58, 177–183. [Google Scholar] [CrossRef]
- Ramanjaneyulu, P.; Rao, A.V.B. Studies on effect of certain antibiotics on callus growth in Terminalia chebula. Int. J. Curr. Res. Rev. 2016, 8, 41–45. [Google Scholar]
- Hazarika, B.N. Morpho-physiological disorders in in vitro culture of plants. Sci. Horticult. 2006, 108, 105–120. [Google Scholar] [CrossRef]
- Garcia, C.; Furtado de Almeida, A.; Costa, M.; Britto, D.; Valle, R.; Royaert, S.; Marelli, J.P. Abnormalities in somatic embryogenesis caused by 2,4-D: An overview. Plant Cell Tissue Organ. 2019, 137, 193–212. [Google Scholar]
- Shirani Bidabadi, S.; Sariah, M.; Zakaria, W.; Subramaniam, S.; Maziah, M. Induced mutations for enhancing variability of banana (Musa spp.) shoot tip cultures using ethyl methanesulphonate (EMS). Aust. J. Crop Sci. 2012, 6, 391–401. [Google Scholar]
- To, J.P.; Haberer, G.; Ferreira, F.J.; Deruère, J.; Mason, M.G.; Schaller, G.E.; Alonso, J.M.; Ecker, J.R.; Kieber, J.J. Type-A Arabidopsis response regulators are partially redundant negative regulators of cytokinin signaling. Plant Cell 2014, 16, 658–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Morphology of different stages during plant organogenesis in Nicotiana rustica. (A) Root regeneration from callus tissue. (B) Shooty callus. (C) Embryo regeneration from callus tissue. (D) Compact callus (E) Shoot regeneration from callus tissue (F) Shoot clumps regenerated from a friable callus.
Figure 1.
Morphology of different stages during plant organogenesis in Nicotiana rustica. (A) Root regeneration from callus tissue. (B) Shooty callus. (C) Embryo regeneration from callus tissue. (D) Compact callus (E) Shoot regeneration from callus tissue (F) Shoot clumps regenerated from a friable callus.

Figure 2.
Totipotency and pluripotency in plant regeneration.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bidabadi, S.S.; Jain, S.M. Cellular, Molecular, and Physiological Aspects of In Vitro Plant Regeneration. Plants 2020, 9, 702. https://doi.org/10.3390/plants9060702
AMA Style
Bidabadi SS, Jain SM. Cellular, Molecular, and Physiological Aspects of In Vitro Plant Regeneration. Plants. 2020; 9(6):702. https://doi.org/10.3390/plants9060702
Chicago/Turabian StyleBidabadi, Siamak Shirani, and S. Mohan Jain. 2020. "Cellular, Molecular, and Physiological Aspects of In Vitro Plant Regeneration" Plants 9, no. 6: 702. https://doi.org/10.3390/plants9060702
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.