Screening of Natural Stilbene Oligomers from Vitis vinifera for Anticancer Activity on Human Hepatocellular Carcinoma Cells

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
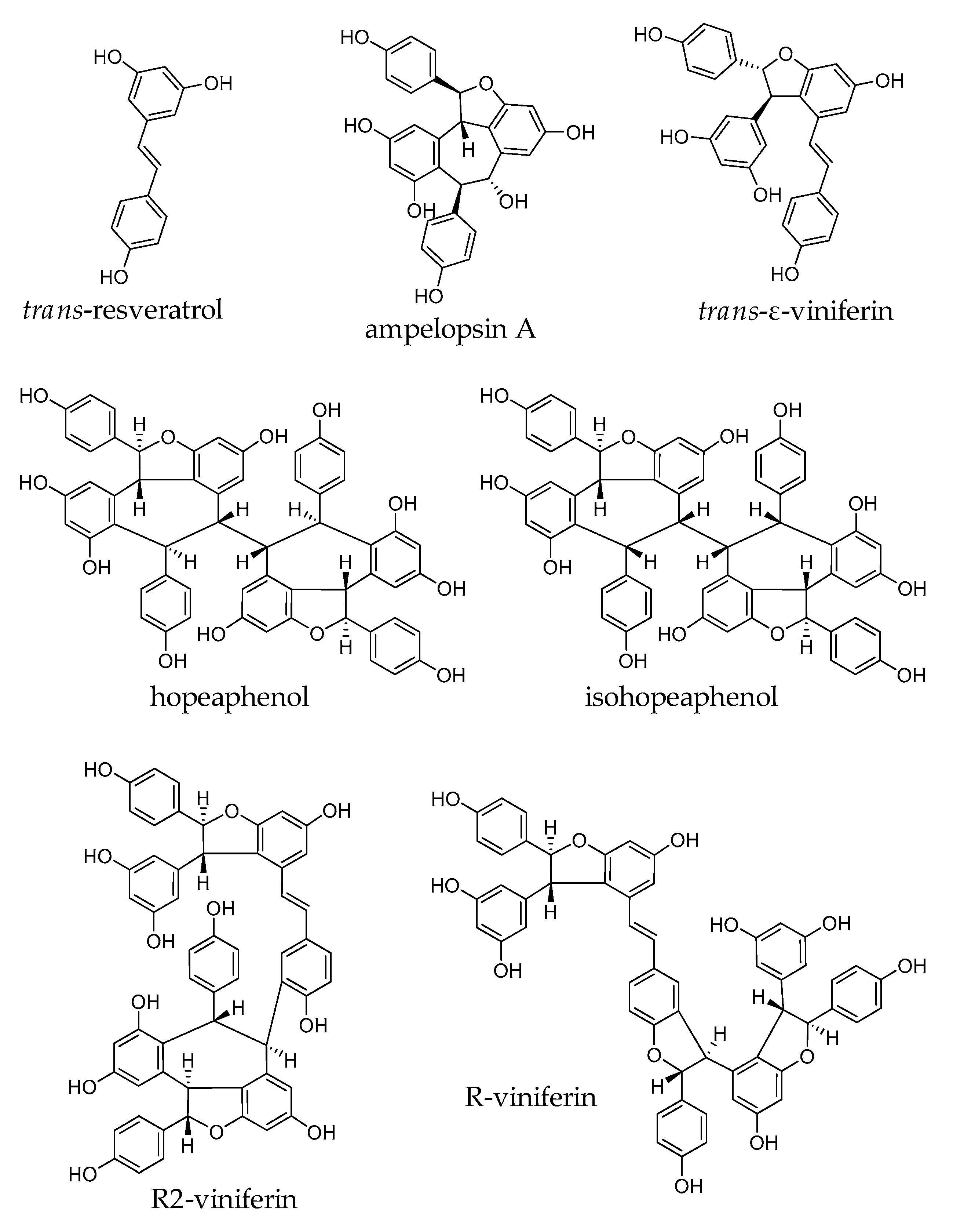
2.1. Stilbenes from Vitis vinifera
2.2. Cell Culture
2.3. Cell Viability Assay
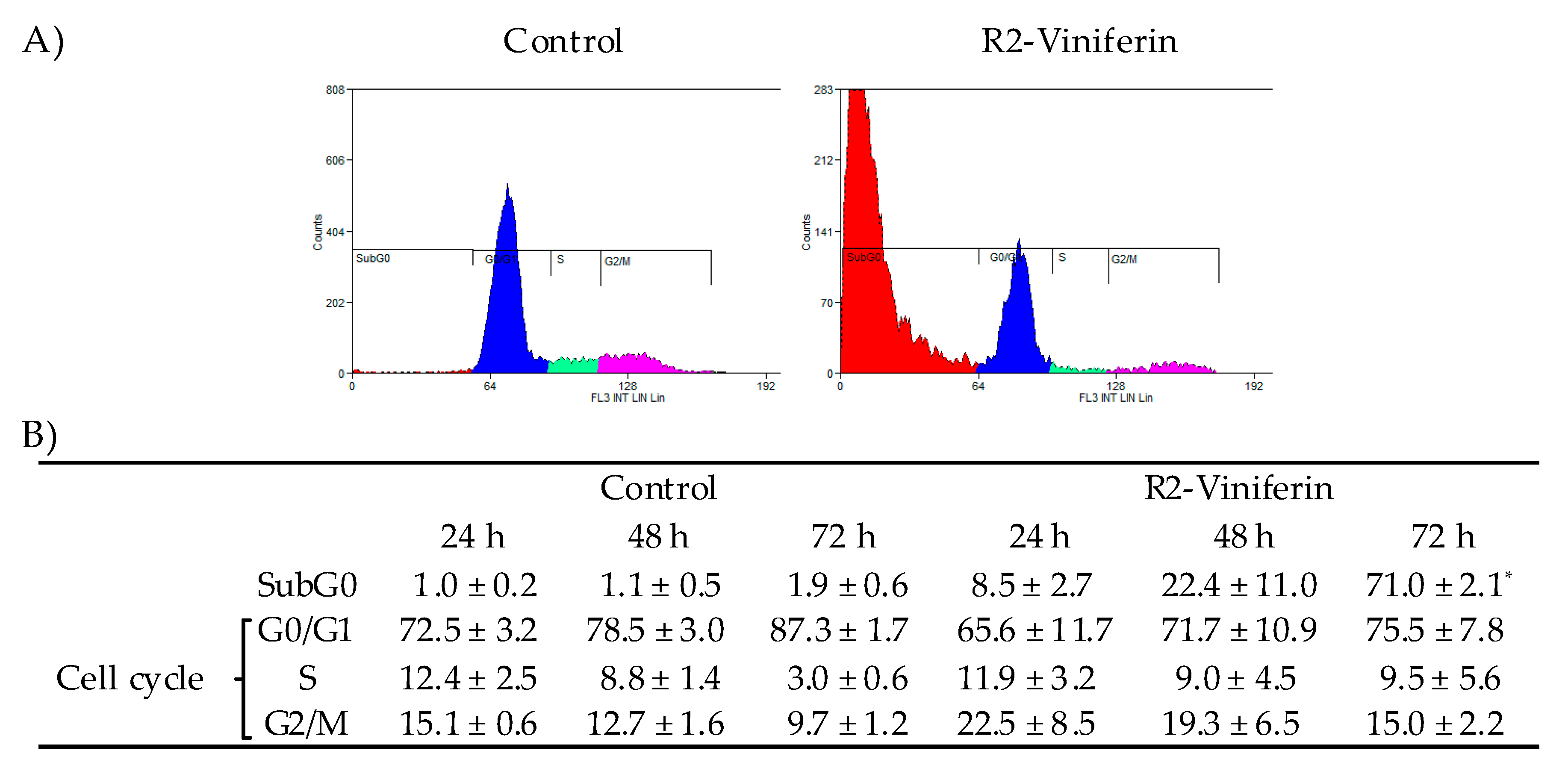
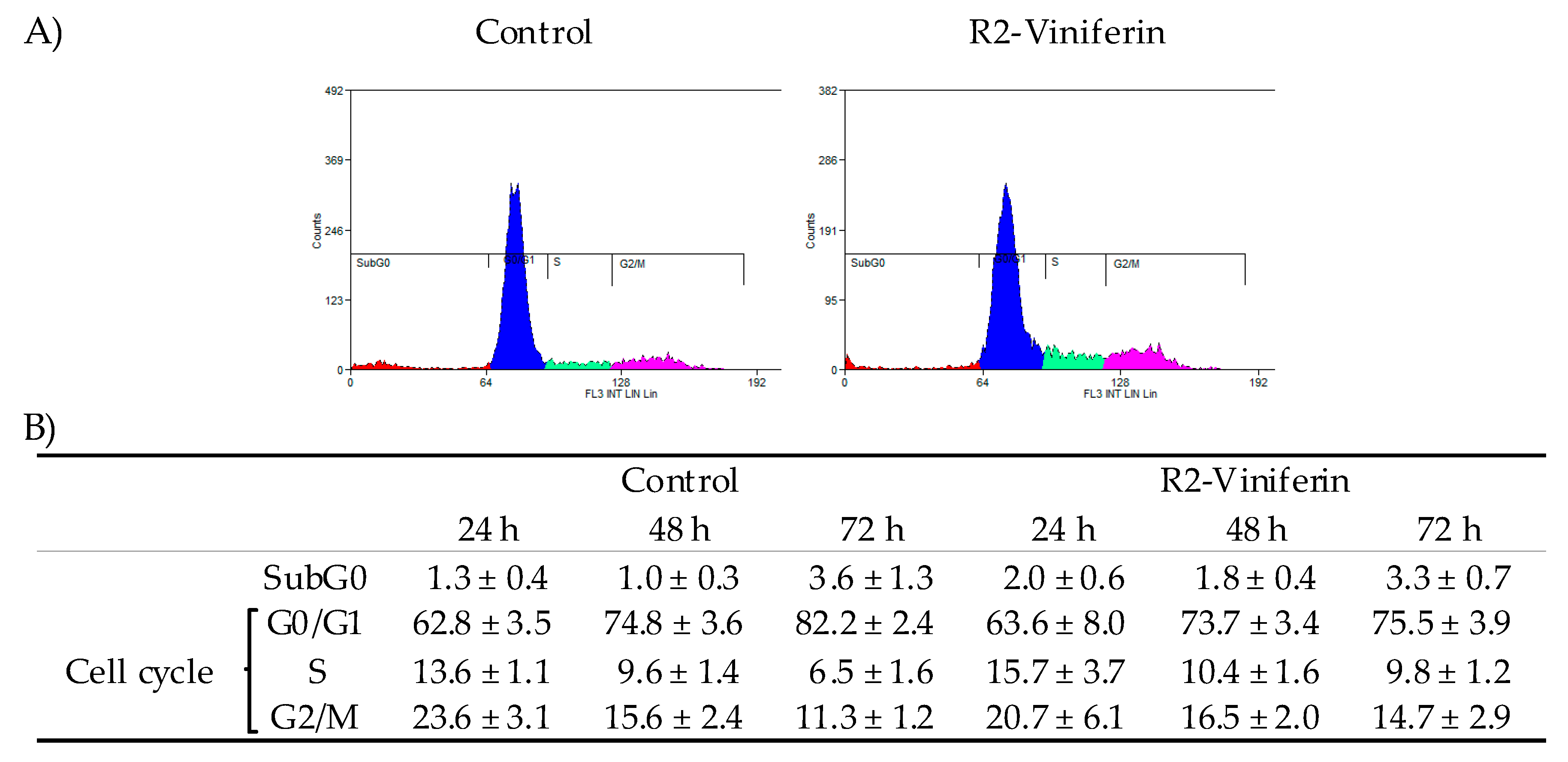
2.4. Cell Cycle Analysis
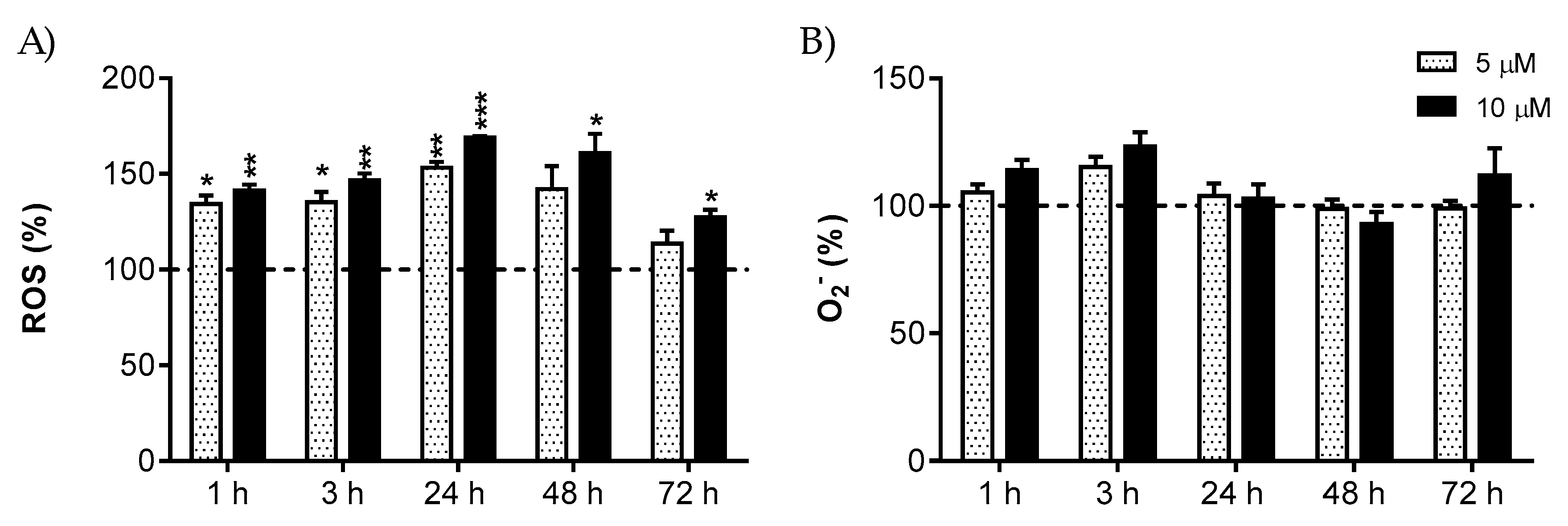
2.5. Intracellular ROS and Mitochondrial O2− Measurement
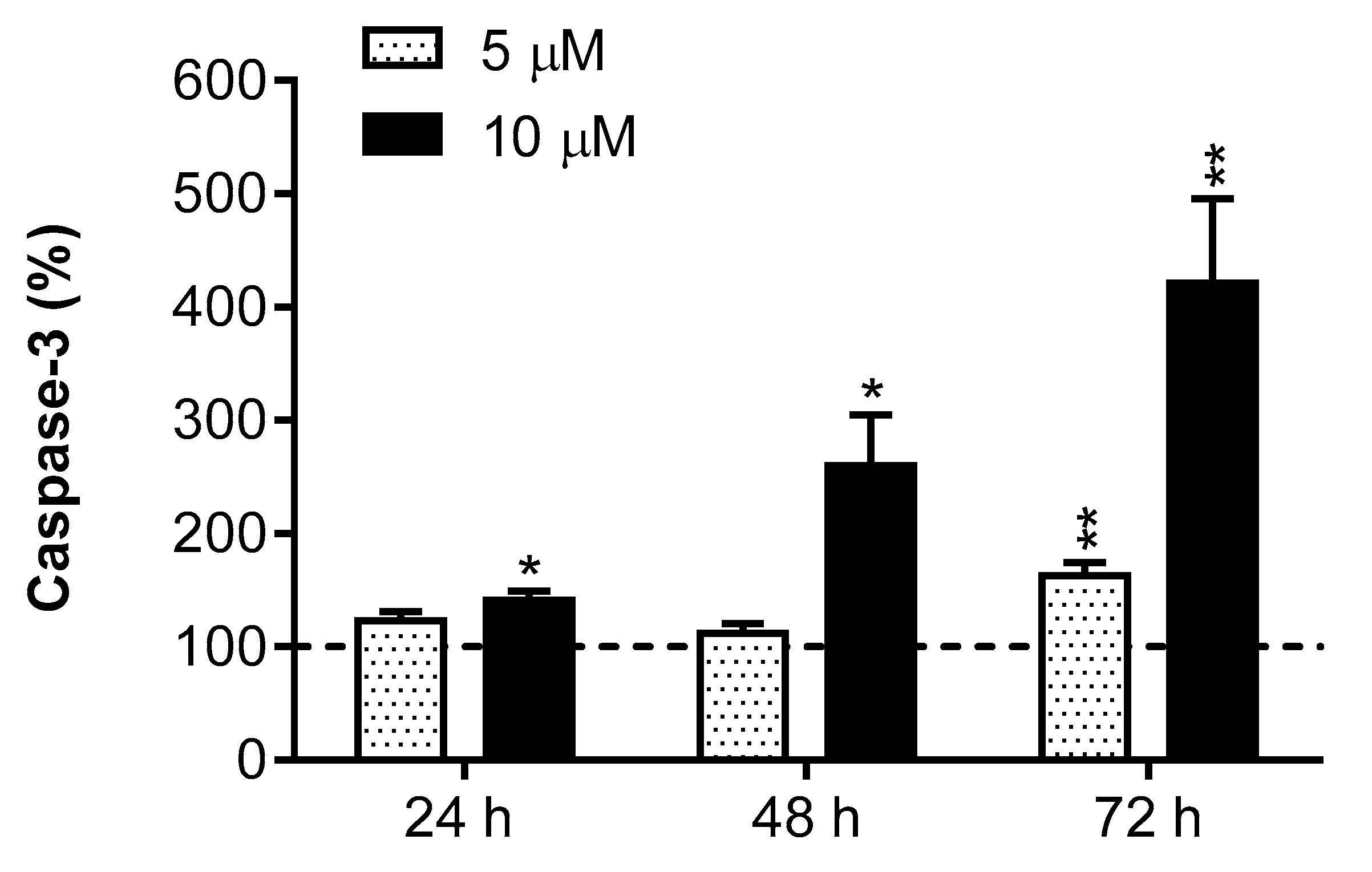
2.6. Caspase-3 Activity
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BSA | Bovine serum albumin |
| CDKs | Cyclin-dependent kinases |
| DCF | 2´,7´-Dichlorofluorescein |
| DMSO | Dimethyl sulfoxide |
| DTT | Dithiothreitol |
| ECL | Enhanced chemiluminescence |
| EDTA | Ethylenediaminetetraacetic acid |
| EGTA | Ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid |
| ERK | Extracellular-signal-regulated kinase |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| H2DCF-DA | 2′,7′ Dichlorodihydrofluorescein diacetate |
| HCC | Hepatocellular carcinoma |
| HEPES | 4-(2-Hydroxyethyl)-1-piperazineethanesulfonic acid |
| NF-kB | Nuclear factor kappa B |
| O2− | Superoxide anion |
| PBS | Phosphate buffer saline |
| ROS | Reactive oxygen species |
References
- Mittal, S.; El-Serag, H.B. Epidemiology of hepatocellular carcinoma: Consider the population. J. Clin. Gastroenterol. 2013, 47, S2–S6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.D.; Hainaut, P.; Gores, G.J.; Amadou, A.; Plymoth, A.; Roberts, L.R. A global view of hepatocellular carcinoma: Trends, risk, prevention and management. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 589–604. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.P.; Harris, C.C. Molecular epidemiology of human cancer: Contribution of mutation spectra studies of tumor suppressor genes. Cancer Res. 1998, 58, 4023–4037. [Google Scholar] [PubMed]
- Link, T.; Iwakuma, T. Roles of p53 in extrinsic factor-induced liver carcinogenesis. Hepatoma Res. 2017, 3, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.P.; Schwank, J.; Staib, F.; Wang, X.W.; Harris, C.C. TP53 mutations and hepatocellular carcinoma: Insights into the etiology and pathogenesis of liver cancer. Oncogene 2007, 26, 2166–2176. [Google Scholar] [CrossRef] [Green Version]
- Kunst, C.; Haderer, M.; Heckel, S.; Schlosser, S.; Müller, M. The p53 family in hepatocellular carcinoma. Transl. Cancer Res. 2016, 5, 632–638. [Google Scholar] [CrossRef]
- Khoury, L.; Zalko, D.; Audebert, M. Evaluation of four human cell lines with distinct biotransformation properties for genotoxic screening. Mutagenesis 2016, 31, 83–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- El Khawand, T.; Courtois, A.; Valls, J.; Richard, T.; Krisa, S. A review of dietary stilbenes: Sources and bioavailability. Phytochem. Rev. 2018, 17, 1007–1029. [Google Scholar] [CrossRef]
- Iveta Pugajeva, I.; Perkonsa, I.; Górnaś, P. Identification and determination of stilbenes by Q-TOF in grape skins, seeds, juice and stems. J. Food Compost. Anal. 2018, 74, 44–52. [Google Scholar] [CrossRef]
- Loupit, G.; Prigent, S.; Franc, C.; De Revel, G.; Richard, T.; Cookson, S.Y.; Valls, J. Polyphenol profiles of just pruned grapevine canes from wild Vitis accessions and Vitis vinifera cultivars. J. Agric. Food Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, R.F.; Valls-Fonayet, J.; Richard, T.; Cantos-Villar, E. A rapid quantification of stilbene content in wine by ultra-high pressure liquid chromatography – Mass spectrometry. Food Control 2020, 108, 06821. [Google Scholar] [CrossRef]
- Zhu, X.; Wu, C.; Qiu, S.; Yuan, X.; Li, L. Effects of Resveratrol on Glucose Control and Insulin Sensitivity in Subjects with Type 2 Diabetes: Systematic Review and Meta-Analysis. Nutr. Metab. 2017, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Vervandier-Fasseur, D.; Latruffe, N. The Potential Use of Resveratrol for Cancer Prevention. Molecules 2019, 24, 4506. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.K.; Luo, J.Y.; Lau, C.W.; Chen, Z.Y.; Tian, X.Y.; Huang, Y. Pharmacological basis and new insights of resveratrol action in the cardiovascular system. Br. J. Pharmacol. 2019, 2019, 1–20. [Google Scholar] [CrossRef]
- Sun, A.Y.; Wang, Q.; Simonyi, A.; Sun, G.Y. Resveratrol as a therapeutic agent for neurodegenerative diseases. Mol. Neurobiol. 2010, 41, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Gabaston, J.; El Khawand, T.; Waffo-Teguo, P.; Decendit, A.; Richard, T.; Mérillon, J.M.; Pavela, R. Stilbenes from grapevine root: A promising natural insecticide against Leptinotarsa decemlineata. J. Pest. Sci. 2018, 91, 897–906. [Google Scholar] [CrossRef]
- Trepiana, J.; Meijide, S.; Navarro, R.; Hernandez, M.L.; Ruiz-Sanz, J.I.; Ruiz-Larrea, M.B. Influence of oxygen partial pressure on the characteristics of human hepatocarcinoma cells. Redox Biol. 2017, 12, 103–113. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Willenberg, I.; Brauer, W.; Empl, M.T.; Schebb, N.H. Development of a rapid LC-UV method for the investigation of chemical and metabolic stability of resveratrol oligomers. J. Agric. Food Chem. 2012, 60, 7844–7850. [Google Scholar] [CrossRef]
- Espinoza, J.L.; Inaoka, P.T. Gnetin-C and other resveratrol oligomers with cancer chemopreventive potential. Ann. N. Y. Acad. Sci. 2017, 1403, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Baechler, S.A.; Schroeter, A.; Dicker, M.; Steinberg, P.; Marko, D. Topoisomerase II-targeting properties of a grapevine-shoot extract and resveratrol oligomers. J. Agric. Food Chem. 2014, 62, 780–788. [Google Scholar] [CrossRef]
- Colin, D.; Lancon, A.; Delmas, D.; Lizard, G.; Abrossinow, J.; Kahn, E.; Jannin, B.; Latruffe, N. Antiproliferative activities of resveratrol and related compounds in human hepatocyte derived HepG2 cells are associated with biochemical cell disturbance revealed by fluorescence analyses. Biochimie 2008, 90, 1674–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, D.; Zhelev, Z.; Semkova, S.; Aoki, I.; Bakalova, R. Resveratrol Modulates the Redox-status and Cytotoxicity of Anticancer Drugs by Sensitizing Leukemic Lymphocytes and Protecting Normal Lymphocytes. Anticancer Res. 2019, 39, 3745–3755. [Google Scholar] [CrossRef] [PubMed]
- Yen, C.M.; Tsai, C.W.; Chang, W.S.; Yang, Y.C.; Hung, Y.W.; Lee, H.T.; Shen, C.C.; Sheu, M.L.; Wang, J.Y.; Gong, C.L.; et al. Novel Combination of Arsenic Trioxide (As2O3) Plus Resveratrol in Inducing Programmed Cell Death of Human Neuroblastoma SK-N-SH Cells. Cancer Genom. Proteom. 2018, 15, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Redondo-Blanco, S.; Fernández, J.; Gutiérrez-Del-Río, I.; Villar, C.J.; Lombó, F. New Insights toward Colorectal Cancer Chemotherapy Using Natural Bioactive Compounds. Front. Pharmacol. 2017, 8, 109. [Google Scholar] [CrossRef] [Green Version]
- Loisruangsin, A.; Hikita, K.; Seto, N.; Niwa, M.; Takaya, Y.; Kaneda, N. Structural analysis of the inhibitory effects of polyphenols, (+)-hopeaphenol and (−)-isohopeaphenol, on human SIRT1. Biofactors 2018, 45, 253–258. [Google Scholar] [CrossRef]
- Asenstrfer, R.E.; Markides, A.J.; Iland, P.G.; Jones, G.P. Formation of R2-viniferin during red wine fermentation and maturation. Aust. J. Grape Wine Res. 2003, 9, 40–46. [Google Scholar] [CrossRef]
- Shin, D.; Kwon, H.Y.; Sohn, E.J.; Nam, M.S.; Kim, J.H.; Lee, J.C.; Ryu, S.Y.; Park, B.; Kim, S.H. Upregulation of death receptor 5 and production of reactive oxygen species mediate sensitization of PC-3 prostate cancer cells to TRAIL induced apoptosis by R2-viniferin. Cell Physiol. Biochem. 2015, 36, 1151–1162. [Google Scholar] [CrossRef]
- Sung, M.J.; Davaatseren, M.; Kim, W.; Park, S.K.; Kim, S.H.; Hur, H.J.; Kim, M.S.; Kim, Y.S.; Kwon, D.Y. Vitisin A suppresses LPS-induced NO production by inhibiting ERK, p38, and NF-kappaB activation in RAW 264.7 cells. Int. Immunopharmacol. 2009, 9, 319–323. [Google Scholar] [CrossRef]
- Liao, W.; Liu, J.; Liu, B.; Huang, X.; Yin, Y.; Cai, D.; Li, M.; Zhu, R. JIB-04 induces cell apoptosis via activation of the p53/Bcl-2/caspase pathway in MHCC97H and HepG2 cells. Oncol. Rep. 2018, 40, 3812–3820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, F.; Yang, Z.; Yu, D.; Wang, J.; Li, R.; Ding, G. Sepia ink oligopeptide induces apoptosis in prostate cancer cell lines via caspase-3 activation and elevation of Bax/Bcl-2 ratio. Mar. Drugs 2012, 10, 2153–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delmas, D.; Jannin, B.; Cherkaoui, M.; Latruffe, N. Inhibitory effect of resveratrol on the proliferation of human and rat hepatic derived cell lines. Oncol. Rep. 2000, 7, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.; Persson, J.L. Cancer therapy: Targeting cell cycle regulators. Anticancer Agents Med. Chem. 2008, 8, 723–731. [Google Scholar] [CrossRef]
- Hongmei Luo, H.; Yang, A.; Schulte, B.A.; Wargovich, M.J.; Wang, G.Y. Resveratrol Induces Premature Senescence in Lung Cancer Cells via ROS-Mediated DNA Damage. PLoS ONE 2013, 8, e60065. [Google Scholar] [CrossRef] [Green Version]
- Kahn, M.A.; Chen, H.; Wan, X.; Tania, M.; Xu, A.; Chen, F.; Zhanget, D. Regulatory effects of resveratrol on antioxidant enzymes: A mechanism of growth inhibition and apoptosis induction in cancer cells. Mol. Cells 2013, 35, 219–225. [Google Scholar] [CrossRef]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef]
- Walle, T. Bioavailability of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef]
- Snyder, S.A.; Gollner, A.; Chiriac, M.I. Regioselective reactions for programmable resveratrol oligomer synthesis. Nature 2011, 474, 461–466. [Google Scholar] [CrossRef]
- Matsuura, B.S.; Keylor, M.H.; Li, B.; Lin, Y.; Allison, S.; Pratt, D.A.; Stephenson, C.R. A scalable biomimetic synthesis of resveratrol dimers and systematic evaluation of their antioxidant activities. Angew. Chem. Int. Ed. Engl. 2015, 54, 3754–3757. [Google Scholar] [CrossRef] [Green Version]
- Keylor, M.H.; Matsuura, B.S.; Griesser, M.; Chauvin, J.R.; Harding, R.A.; Kirillova, M.S.; Zhu, X.; Fischer, O.J.; Pratt, D.A.; Stephenson, C.R. Synthesis of resveratrol tetramers via a stereoconvergent radical equilibrium. Science 2016, 354, 1260–1265. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Min, J.S.; Kim, D.; Zheng, Y.F.; Mailar, K.; Choi, W.J.; Lee, C.; Bae, S.K. A simple and sensitive liquid chromatography-tandem mass spectrometry method for trans-ε-viniferin quantification in mouse plasma and its application to a pharmacokinetic study in mice. J. Pharm. Biomed. Anal. 2017, 134, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Courtois, A.; Atgié, C.; Marchal, A.; Hornedo-Ortega, R.; Lapèze, C.; Faure, C.; Richard, T.; Krisa, S. Tissular distribution and metabolism of trans-ε-viniferin after intraperitoneal injection in rat. Nutrients 2018, 10, 1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Hu, M. Bioavailability challenges associated with development of anti-cancer phenolics. Mini-Rev. Med. Chem. 2010, 10, 550–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courtois, A.; Garcia, M.; Krisa, S.; Atgié, C.; Sauvant, P.; Richard, T.; Faure, C. Encapsulation of ε-viniferin in onion-type multi-lamellar liposomes increases its solubility and its photo-stability and decreases its cytotoxicity on Caco-2 intestinal cells. Food Funct. 2019, 10, 2573–2582. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
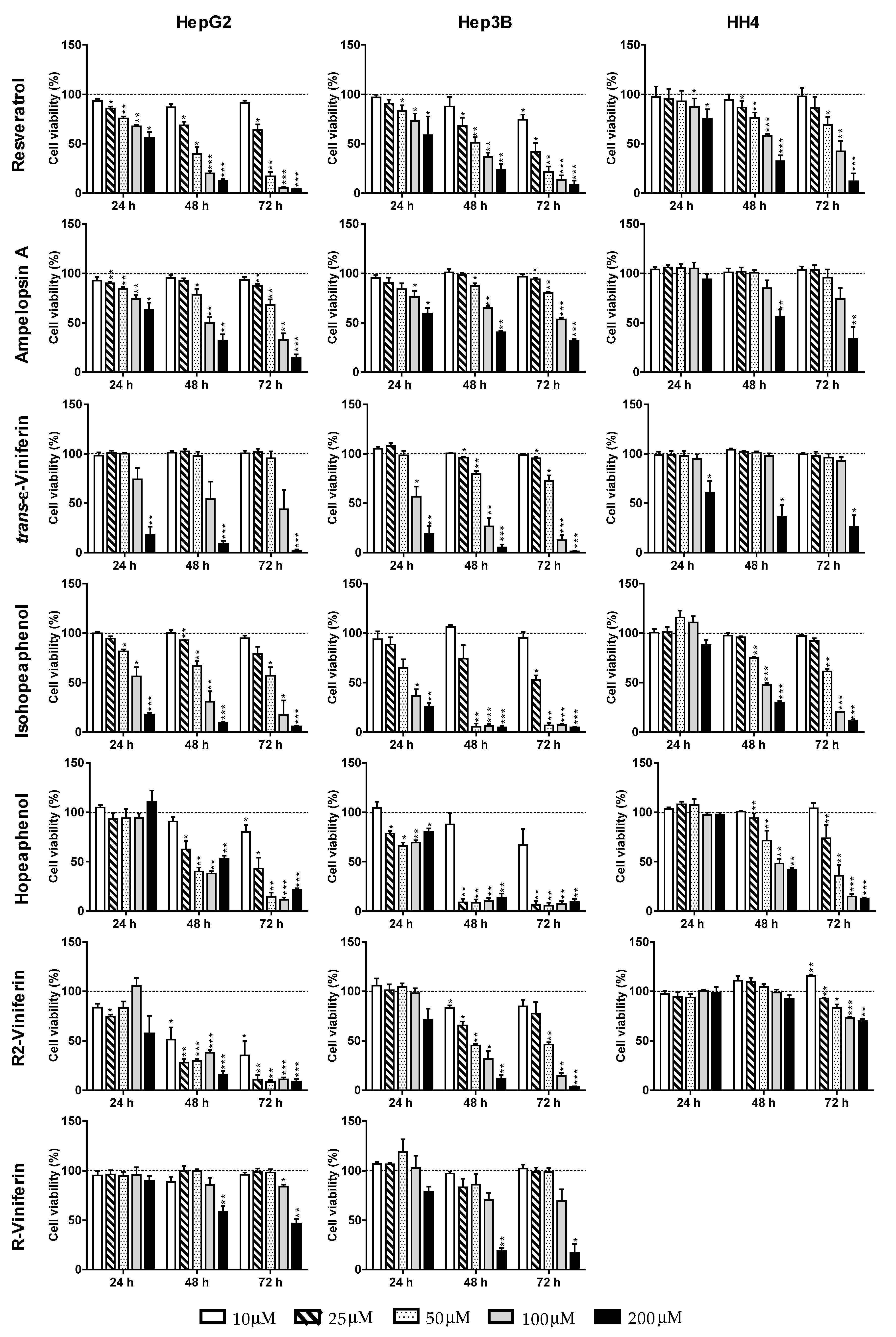
| Compound | Time (h) | HepG2 | Hep3B | HH4 |
|---|---|---|---|---|
| Monomer | ||||
| trans-Resveratrol | 24 h | >200 | >200 | >200 |
| 48 h | 40.3 ± 9.3 | 42.0 ± 11.6 | 135.0 ± 9.0 | |
| 72 h | 30.3 ± 4.4 | 21.0 ± 16.8 | 93.0 ± 16.1 | |
| Dimer | ||||
| Ampelopsin A | 24 h | >200 | >200 | >200 |
| 48 h | 98.6 ± 24.9 | 147.8 ± 14.4 | 178.3 ± 67.8 | |
| 72 h | 75.5 ± 21.5 | 109.1 ± 7.3 | 133.8 ± 34.7 | |
| trans-ε-Viniferin | 24 h | 140.0 ± 39.7 | 108.1 ± 31.8 | >200 |
| 48 h | 103.7 ± 19.2 | 73.9 ± 17.3 | 192.7 ± 21.1 | |
| 72 h | 94.8 ± 28.3 | 63.1 ± 10.8 | 177.9 ± 20.5 | |
| Tetramer | ||||
| Hopeaphenol | 24 h | >200 | >200 | >200 |
| 48 h | 27.0 ± 3.3 | 16.8 ± 2.3 | 92.0 ± 38.0 | |
| 72 h | 24.4 ± 2.0 | 13.1 ± 4.1 | 37.6 ± 13.0 | |
| Isohopeaphenol | 24 h | 113.0 ± 33.0 | 86.6 ± 11.7 | >200 |
| 48 h | 68.8 ± 31.0 | 37.0 ± 4.5 | 96.0 ± 5.5 | |
| 72 h | 54.1 ± 34.0 | 26.0 ± 3.0 | 63.7 ± 3.7 | |
| R2-Viniferin | 24 h | >200 | >200 | >200 |
| 48 h | 10.2 ± 8.2 | 43.9 ± 3.6 | >200 | |
| 72 h | 9.7 ± 0.4 | 47.8 ± 2.8 | >200 | |
| R-Viniferin | 24 h | >200 | >200 | n.d b |
| 48 h | >200 | 137.2 ± 19.8 | n.d b | |
| 72 h | 192.0 ± 27.1 | 134.9 ± 35.7 | n.d b | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aja, I.; Ruiz-Larrea, M.B.; Courtois, A.; Krisa, S.; Richard, T.; Ruiz-Sanz, J.-I. Screening of Natural Stilbene Oligomers from Vitis vinifera for Anticancer Activity on Human Hepatocellular Carcinoma Cells. Antioxidants 2020, 9, 469. https://doi.org/10.3390/antiox9060469
Aja I, Ruiz-Larrea MB, Courtois A, Krisa S, Richard T, Ruiz-Sanz J-I. Screening of Natural Stilbene Oligomers from Vitis vinifera for Anticancer Activity on Human Hepatocellular Carcinoma Cells. Antioxidants. 2020; 9(6):469. https://doi.org/10.3390/antiox9060469
Chicago/Turabian StyleAja, Iris, M. Begoña Ruiz-Larrea, Arnaud Courtois, Stéphanie Krisa, Tristan Richard, and José-Ignacio Ruiz-Sanz. 2020. "Screening of Natural Stilbene Oligomers from Vitis vinifera for Anticancer Activity on Human Hepatocellular Carcinoma Cells" Antioxidants 9, no. 6: 469. https://doi.org/10.3390/antiox9060469