Isolation, Identification, and Optimization of γ-Aminobutyric Acid (GABA)-Producing Bacillus cereus Strain KBC from a Commercial Soy Sauce moromi in Submerged-Liquid Fermentation

 ,
, 
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
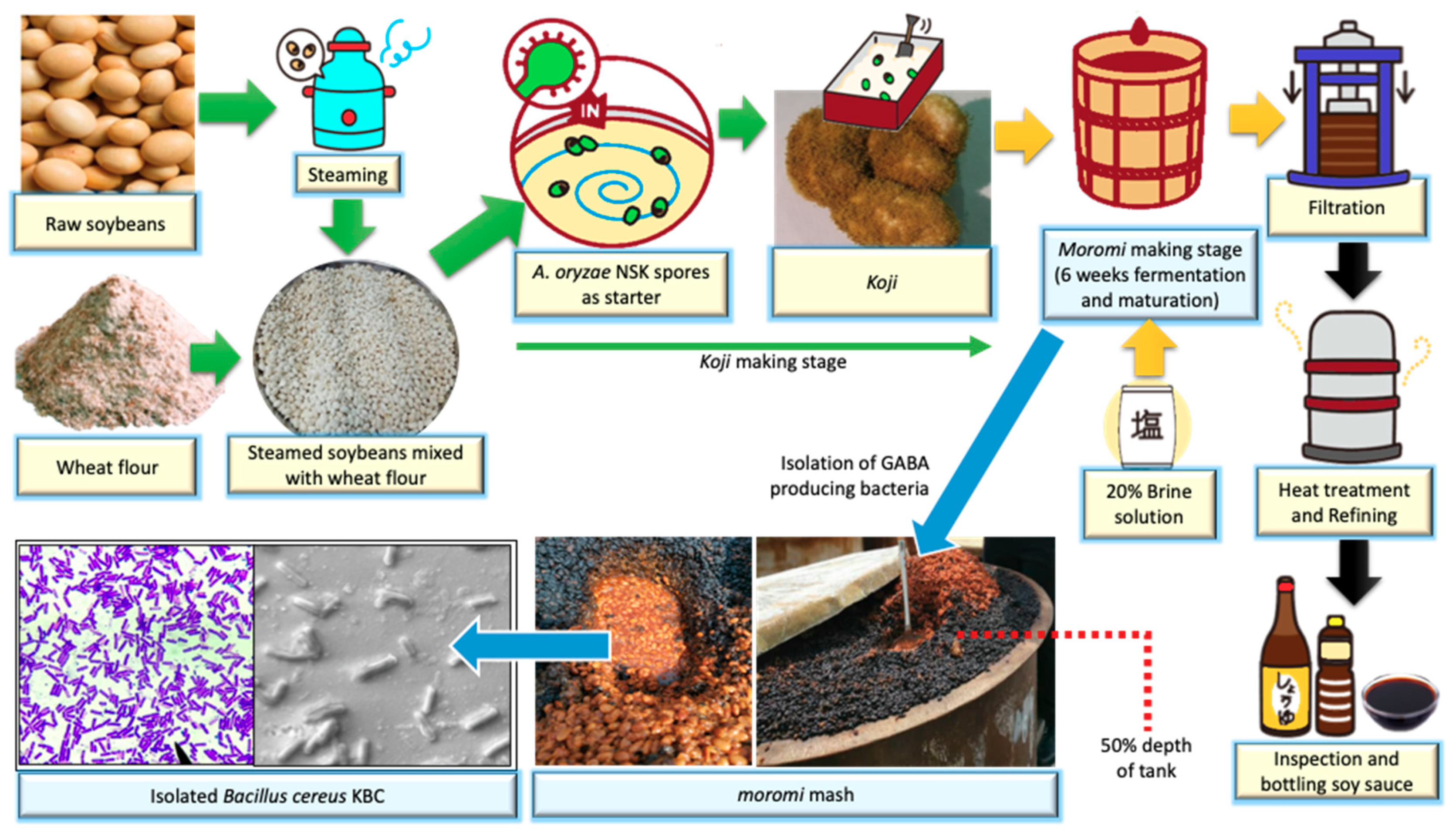
2.1. Isolation of B. cereus KBC
2.2. Morphological Analysis
2.3. Identification Using 16S rRNA
2.4. Inoculum Preparation
2.5. Medium Preparation
2.6. Production of GABA by an Isolated Strain
2.7. Optimisation of GABA Production Using RSM
2.8. Analytical Procedures
2.8.1. Determination of Dry Cell Weight
2.8.2. Determination of GABA
2.9. Statistical Analysis
3. Results and Discussion
3.1. Identification of B. cereus KBC
3.1.1. Morphological Characteristics of B. cereus KBC
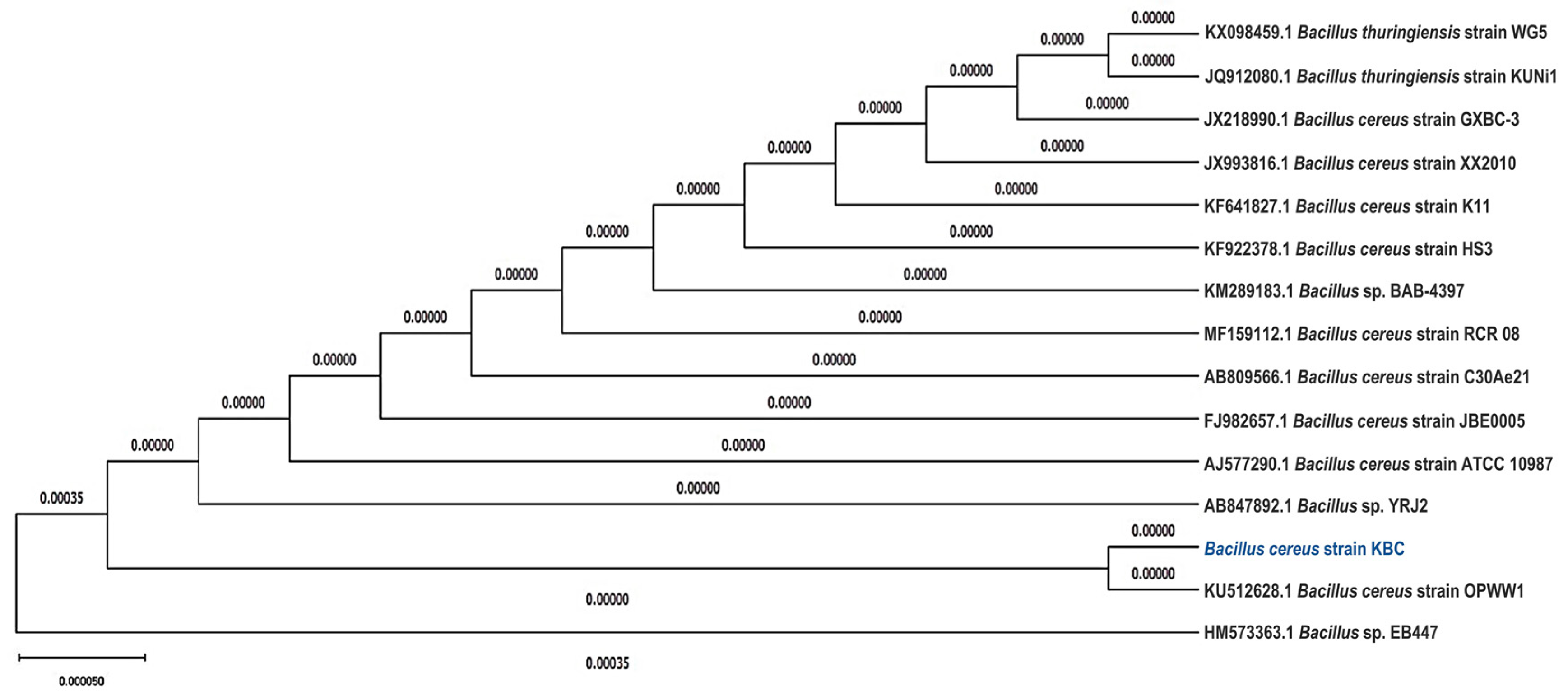
3.1.2. 16S rRNA Identification of B. cereus KBC
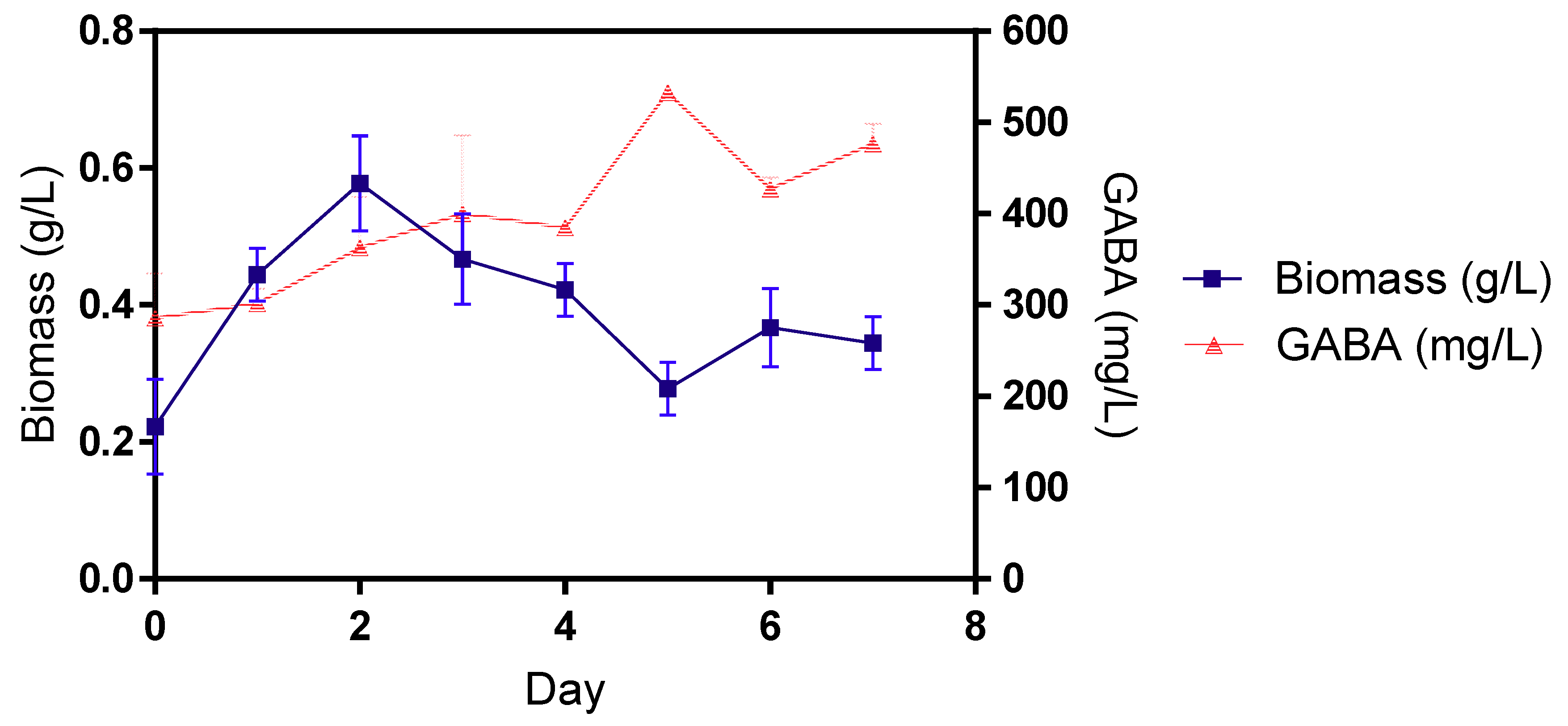
3.2. GABA Production by B. cereus KBC
3.2.1. Optimisation of GABA Production by B. cereus KBC
3.2.2. Validation of the Optimised Conditions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pizzarello, S. Non-Protein Amino Acids. In Encyclopedia of Astrobiology; Gargaud, M., Amils, R., Cleaves, H.J., Eds.; Springer: Berlin/Heidelber, Germany, 2015. [Google Scholar]
- Mander, L.; Liu, H. Comprehensive Natural Products II: Chemistry and Biology; Elsevier: Amsterdam, The Netherlands, 2010. [Google Scholar] [CrossRef]
- Vranova, V.; Rejsek, K.; Skene, K.R.; Formanek, P. Non-protein amino acids: Plant, soil and ecosystem interactions. Plant Soil 2011, 342, 31–48. [Google Scholar] [CrossRef]
- Petroff, O.A.C. GABA and glutamate in the human brain. Neuroscientist 2002, 8, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Komatsuzaki, N.; Shima, J.; Kawamoto, S.; Momose, H.; Kimura, T. Production of γ-aminobutyric acid (GABA) by Lactobacillus paracasei isolated from traditional fermented foods. Food Microbiol. 2005. [Google Scholar] [CrossRef]
- Lee, B.J.; Kim, J.S.; Kang, Y.M.; Lim, J.H.; Kim, Y.M.; Lee, M.S.; Jeong, M.H.; Ahn, C.B.; Je, J.Y. Antioxidant activity and γ-aminobutyric acid (GABA) content in sea tangle fermented by Lactobacillus brevis BJ20 isolated from traditional fermented foods. Food Chem. 2010. [Google Scholar] [CrossRef]
- Aoki, H.; Furuya, Y.; Endo, Y.; Fujimoto, K. Effect of γ-aminobutyric acid-enriched tempeh-like fermented soybean (gaba-tempeh) on the blood pressure of spontaneously hypertensive rats. Biosci. Biotechnol. Biochem. 2003. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Shirai, T.; Ochiai, H.; Kasao, M.; Hayakawa, K.; Kimura, M.; Sansawa, H. Blood-pressure-lowering effect of a novel fermented milk containing γ-aminobutyric acid (GABA) in mild hypertensives. Eur. J. Clin. Nutr. 2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, W.C.; Wang, C.Y.; Shyu, Y.T.; Yu, R.C.; Ho, K.C. Influence of preprocessing methods and fermentation of adzuki beans on γ-aminobutyric acid (GABA) accumulation by lactic acid bacteria. J. Funct. Foods 2013. [Google Scholar] [CrossRef]
- Wan-Mohtar, W.A.A.Q.I.; Ab Kadir, S.; Halim-Lim, S.A.; Ilham, Z.; Hajar-Azhari, S.; Saari, N. Vital parameters for high gamma-aminobutyric acid (GABA) production by an industrial soy sauce koji Aspergillus oryzae NSK in submerged-liquid fermentation. Food Sci. Biotechnol. 2019. [Google Scholar] [CrossRef]
- Wong, C.G.T.; Bottiglieri, T.; Snead, O.C. GABA, γ-hydroxybutyric acid, and neurological disease. Ann. Neurol. 2003. [Google Scholar] [CrossRef]
- Capitani, G.; De Biase, D.; Aurizi, C.; Gut, H.; Bossa, F.; Grütter, M.G. Crystal structure and functional analysis of Escherichia coli glutamate decarboxylase. EMBO J. 2003. [Google Scholar] [CrossRef] [Green Version]
- Adeghate, E.; Ponery, A.S. GABA in the endocrine pancreas: Cellular localization and function in normal and diabetic rats. Tissue Cell 2002. [Google Scholar] [CrossRef] [PubMed]
- Cavagnini, F.; Pinto, M.; Dubini, A.; Invitti, C.; Cappelletti, G.; Polli, E.E. Effects of gamma aminobutyric acid (GABA) and muscimol on endocrine pancreatic function in man. Metabolism 1982. [Google Scholar] [CrossRef]
- Dhakal, R.; Bajpai, V.K.; Baek, K.H. Production of GABA (γ-aminobutyric acid) by microorganisms: A review. Braz. J. Microbiol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Cao, Y. Lactic acid bacterial cell factories for gamma-aminobutyric acid. Amino Acids 2010, 39, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Brower, P.L.; Winkle, D.D. Chiral HPLC separations for process development of S-(+)-isobutyl GABA, a potential anti-epileptic agent. J. Liq. Chromatogr. Relat. Technol. 1996. [Google Scholar] [CrossRef]
- Venkatesh, T.; Mainkar, P.S.; Chandrasekhar, S. Total synthesis of (±)-galanthamine from GABA through regioselective aryne insertion. Org. Biomol. Chem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Román, S.; Sánchez-Siles, L.M.; Siegrist, M. The importance of food naturalness for consumers: Results of a systematic review. Trends Food Sci. Technol. 2017, 67, 44–57. [Google Scholar] [CrossRef]
- Holzapfel, W.H.; Geisen, R.; Schillinger, U. Biological preservation of foods with reference to protective cultures, bacteriocins and food-grade enzymes. Int. J. Food Microbiol. 1995, 24, 343–362. [Google Scholar] [CrossRef]
- Cui, Y.; Miao, K.; Niyaphorn, S.; Qu, X. Production of gamma-aminobutyric acid from lactic acid bacteria: A systematic review. Int. J. Mol. Sci. 2020, 21, 995. [Google Scholar] [CrossRef] [Green Version]
- Murooka, Y.; Yamshita, M. Traditional healthful fermented products of Japan. J. Ind. Microbiol. Biotechnol. 2008, 35, 791. [Google Scholar] [CrossRef]
- Sarasa, S.B.; Mahendran, R.; Muthusamy, G.; Thankappan, B.; Selta, D.R.F.; Angayarkanni, J. A Brief Review on the Non-protein Amino Acid, Gamma-amino Butyric Acid (GABA): Its Production and Role in Microbes. Curr. Microbiol. 2020, 77, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Ab Kadir, S.; Wan-Mohtar, W.A.A.Q.I.; Mohammad, R.; Abdul Halim Lim, S.; Sabo Mohammed, A.; Saari, N. Evaluation of commercial soy sauce koji strains of Aspergillus oryzae for γ-aminobutyric acid (GABA) production. J. Ind. Microbiol. Biotechnol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, D. Industrialization of Fermented Soy Sauce Production Centering around Japanese Shoyu; Food Science and Technology (Marcel Dekker, Inc.): New York, NY, USA, 2004; pp. 1–88. [Google Scholar]
- Liu, W.; Bao, Q.; Qing, M.; Chen, X.; Sun, T.; Li, M.; Zhang, J.; Yu, J.; Bilige, M.; Sun, T.; et al. Isolation and identification of lactic acid bacteria from Tarag in Eastern Inner Mongolia of China by 16S rRNA sequences and DGGE analysis. Microbiol. Res. 2012. [Google Scholar] [CrossRef] [PubMed]
- Sohedein, M.N.A.; Wan-Mohtar, W.A.A.Q.I.; Hui-Yin, Y.; Ilham, Z.; Chang, J.S.; Supramani, S.; Siew-Moi, P. Optimisation of biomass and lipid production of a tropical thraustochytrid Aurantiochytrium sp. UMACC-T023 in submerged-liquid fermentation for large-scale biodiesel production. Biocatal. Agric. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Mohd Hanafiah, Z.; Wan Mohtar, W.H.M.; Abu Hasan, H.; Jensen, H.S.; Klaus, A.; Wan-Mohtar, W.A.A.Q.I. Performance of wild-Serbian Ganoderma lucidum mycelium in treating synthetic sewage loading using batch bioreactor. Sci. Rep. 2019. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jawan, R.; Kasimin, M.E.; Jalal, S.N.; Mohd Faik, A.A.; Abbasiliasi, S.; Ariff, A. Isolation, characterisation and in vitro evaluation of bacteriocins-producing lactic acid bacteria from fermented products of Northern Borneo for their beneficial roles in food industry. J. Phys. Conf. Ser. 2019, 1358, 012020. [Google Scholar] [CrossRef]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-modulating bacteria of the human gut microbiota. Nat. Microbiol. 2019. [Google Scholar] [CrossRef]
- Zhuang, K.; Jiang, Y.; Feng, X.; Li, L.; Dang, F.; Zhang, W.; Man, C. Transcriptomic response to GABA-producing Lactobacillus plantarum CGMCC 1.2437T induced by L-MSG. PLoS ONE 2018. [Google Scholar] [CrossRef]
- Yang, S.Y.; Lü, F.X.; Lu, Z.X.; Bie, X.M.; Jiao, Y.; Sun, L.J.; Yu, B. Production of γ-aminobutyric acid by Streptococcus salivarius subsp. thermophilus Y2 under submerged fermentation. Amino Acids 2008. [Google Scholar] [CrossRef]
- Lu, X.; Chen, Z.; Gu, Z.; Han, Y. Isolation of γ-aminobutyric acid-producing bacteria and optimization of fermentative medium. Biochem. Eng. J. 2008. [Google Scholar] [CrossRef]
- Di Cagno, R.; Mazzacane, F.; Rizzello, C.G.; De Angelis, M.; Giuliani, G.; Meloni, M.; De Servi, B.; Gobbetti, M. Synthesis of γ-aminobutyric acid (GABA) by Lactobacillus plantarum DSM19463: Functional grape must beverage and dermatological applications. Appl. Microbiol. Biotechnol. 2010. [Google Scholar] [CrossRef] [PubMed]
- Hajar-Azhari, S.; Wan-Mohtar, W.A.A.Q.I.; Ab Kadir, S.; Rahim, M.H.A.; Saari, N. Evaluation of a Malaysian soy sauce koji strain Aspergillus oryzae NSK for γ-aminobutyric acid (GABA) production using different native sugars. Food Sci. Biotechnol. 2018. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
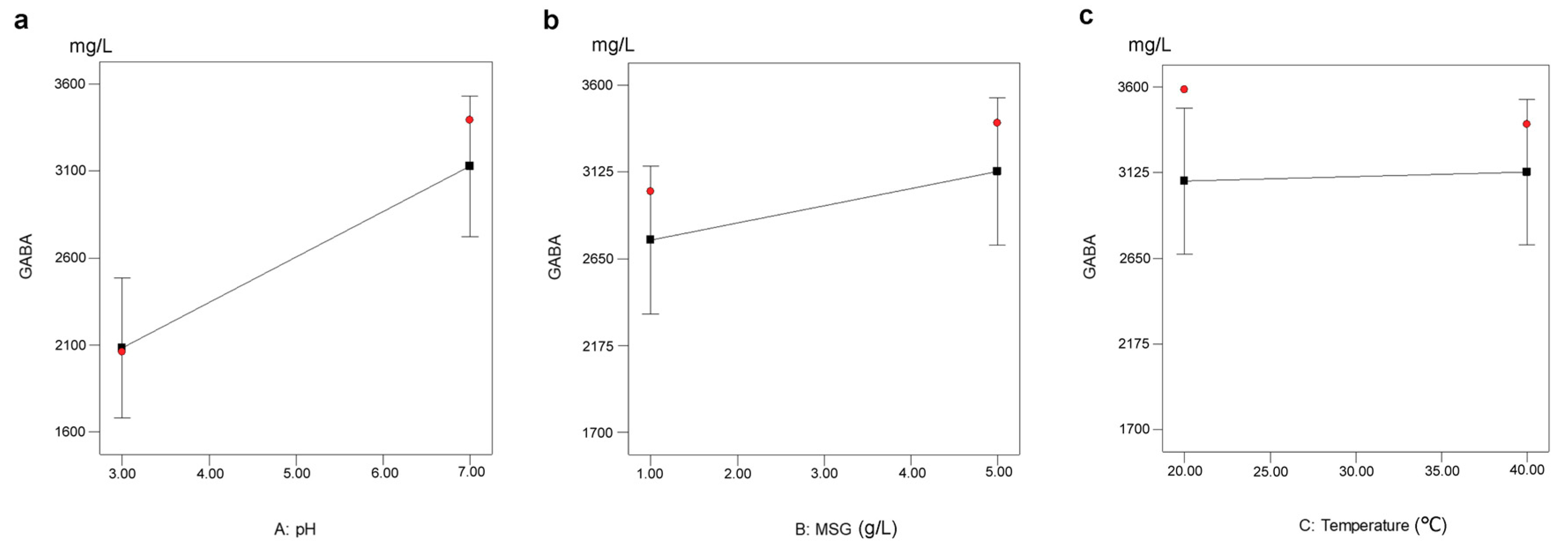{kind=link}
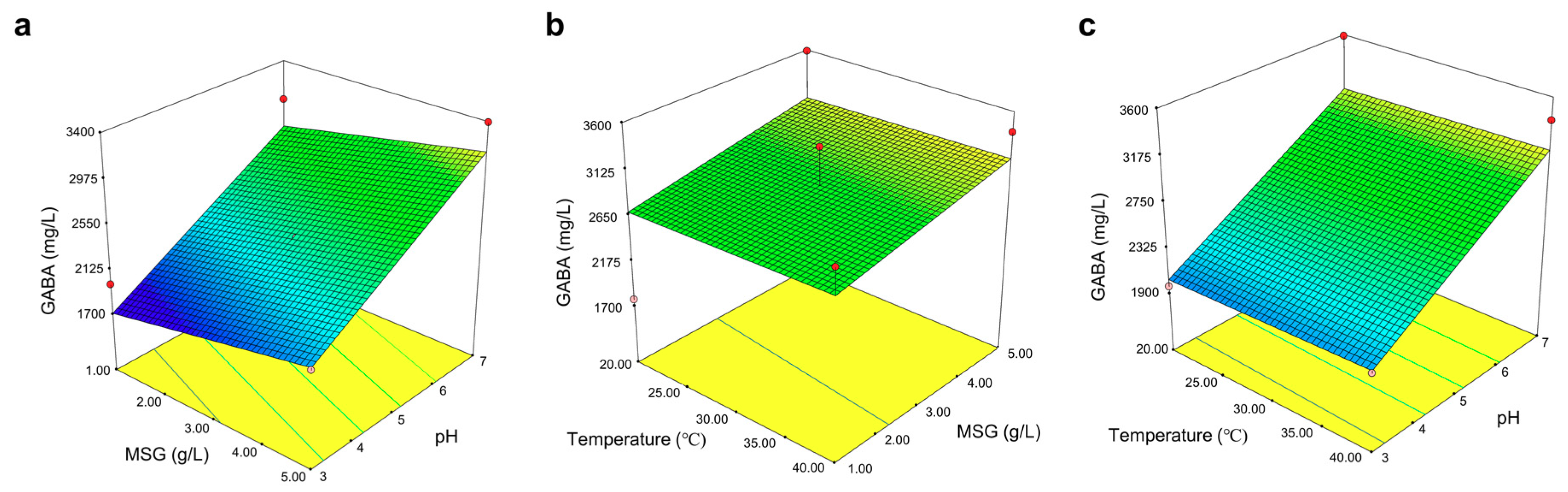{kind=link}
| Variables | Range and Levels | ||
|---|---|---|---|
| −1 | 0 | 1 | |
| pH | 3.0 | 5.0 | 7.0 |
| MSG (g L−1) | 1.0 | 3.0 | 5.0 |
| Temperature (°C) | 20 | 30 | 40 |
| Run No. | Variables | Actual Response | ||
|---|---|---|---|---|
| pH | MSG (g L−1) | Temperature (°C) | GABA (mg L−1) | |
| 1 | 3 | 1 | 40 | 1989.54 |
| 2 | 5 | 3 | 20 | 3303.81 |
| 3 | 5 | 3 | 30 | 2178.9 |
| 4 | 7 | 5 | 40 | 3393.02 |
| 5 | 5 | 3 | 30 | 1990.2 |
| 6 | 5 | 5 | 30 | 2141.45 |
| 7 | 3 | 3 | 30 | 1857.36 |
| 8 | 5 | 3 | 30 | 2181.97 |
| 9 | 3 | 5 | 20 | 1965.97 |
| 10 | 5 | 3 | 30 | 2063.18 |
| 11 | 3 | 5 | 40 | 2060.37 |
| 12 | 5 | 3 | 30 | 2146.49 |
| 13 | 7 | 5 | 20 | 3585.29 |
| 14 | 7 | 1 | 40 | 3018.52 |
| 15 | 5 | 3 | 40 | 2379.37 |
| 16 | 5 | 1 | 30 | 2520.9 |
| 17 | 7 | 3 | 30 | 3303.19 |
| 18 | 3 | 1 | 20 | 1969.95 |
| 19 | 7 | 1 | 20 | 1766.99 |
| 20 | 5 | 3 | 30 | 2047.25 |
| Source | Sum of Squares | Mean Square | DF | F Value | Prob > F | |
|---|---|---|---|---|---|---|
| Model | 3,088,531.300 | 1,029,510.433 | 3 | 4.965 | 0.0127 * | significant |
| A: pH | 2,728,832.702 | 2,728,832.702 | 1 | 13.161 | 0.0023 * | significant |
| B: MSG | 353,508.535 | 353,508.535 | 1 | 1.705 | 0.2101 | |
| C: Temperature | 6190.063 | 6190.063 | 1 | 0.030 | 0.8650 | |
| Residual | 3,317,449.610 | 207,340.600 | 16 | |||
| Pure Error | 31,289.248 | 6257.850 | 5 | |||
| Lack of Fit | 3,286,160.362 | 11 | 47.739 | 0.0002 * | significant | |
| Cor Total | 6,405,980.910 | 298,741.851 | 19 | |||
| Standard Deviation = 455.35 | Mean = 2393.19 | Adequate Precision = 7.221 | ||||
| R2 = 0.4821 | Adjusted R2 = 0.3850 | |||||
| Isolated Strain | Source | GABA Concentration (mg L−1) | Reference |
|---|---|---|---|
| Lactobacillus paracasei NFRI 7415 | Fermented fish | 31,145.30 | [5] |
| Lactobacillus brevis BJ20 | Kimchi | 2465.00 | [6] |
| Lactococcus lactis subsp. lactis B | Kimchi and yoghurt | 6410.00 | [34] |
| Lactobacillus plantarum DSM19463 | Cheeses | 498.10 | [35] |
| Aspergillus oryzae NSK | Soy sauce koji | 73.13 | [24] |
| Aspergillus oryzae NSK | Soy sauce koji | 354.08 | [36] |
| Aspergillus oryzae NSK (unoptimized) | Soy sauce koji | 3278.31 | [10] |
| Bacillus cereus strain KBC (unoptimized) | Soy sauce moromi | 532.74 | This study |
| Bacillus cereus strain KBC (optimized) | Soy sauce moromi | 3393.02 | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan-Mohtar, W.A.A.Q.I.; Sohedein, M.N.A.; Ibrahim, M.F.; Ab Kadir, S.; Suan, O.P.; Weng Loen, A.W.; Sassi, S.; Ilham, Z. Isolation, Identification, and Optimization of γ-Aminobutyric Acid (GABA)-Producing Bacillus cereus Strain KBC from a Commercial Soy Sauce moromi in Submerged-Liquid Fermentation. Processes 2020, 8, 652. https://doi.org/10.3390/pr8060652
Wan-Mohtar WAAQI, Sohedein MNA, Ibrahim MF, Ab Kadir S, Suan OP, Weng Loen AW, Sassi S, Ilham Z. Isolation, Identification, and Optimization of γ-Aminobutyric Acid (GABA)-Producing Bacillus cereus Strain KBC from a Commercial Soy Sauce moromi in Submerged-Liquid Fermentation. Processes. 2020; 8(6):652. https://doi.org/10.3390/pr8060652
Chicago/Turabian StyleWan-Mohtar, Wan Abd Al Qadr Imad, Mohamad Nor Azzimi Sohedein, Mohamad Faizal Ibrahim, Safuan Ab Kadir, Ooi Poh Suan, Alan Wong Weng Loen, Soumaya Sassi, and Zul Ilham. 2020. "Isolation, Identification, and Optimization of γ-Aminobutyric Acid (GABA)-Producing Bacillus cereus Strain KBC from a Commercial Soy Sauce moromi in Submerged-Liquid Fermentation" Processes 8, no. 6: 652. https://doi.org/10.3390/pr8060652