The Evaluation of Egg-Parasitic Fungi Paraboeremia taiwanensis and Samsoniella sp. for the Biological Control of Meloidogyne enterolobii on Chinese Cabbage



Abstract
:1. Introduction
2. Materials and Methods
2.1. Nematode Culture
2.2. Fungal Culture
2.3. In Vitro Infection Experiments
2.4. Scanning Electron Microscopy (SEM)
2.5. Greenhouse Trials
2.6. Data Analysis
3. Results
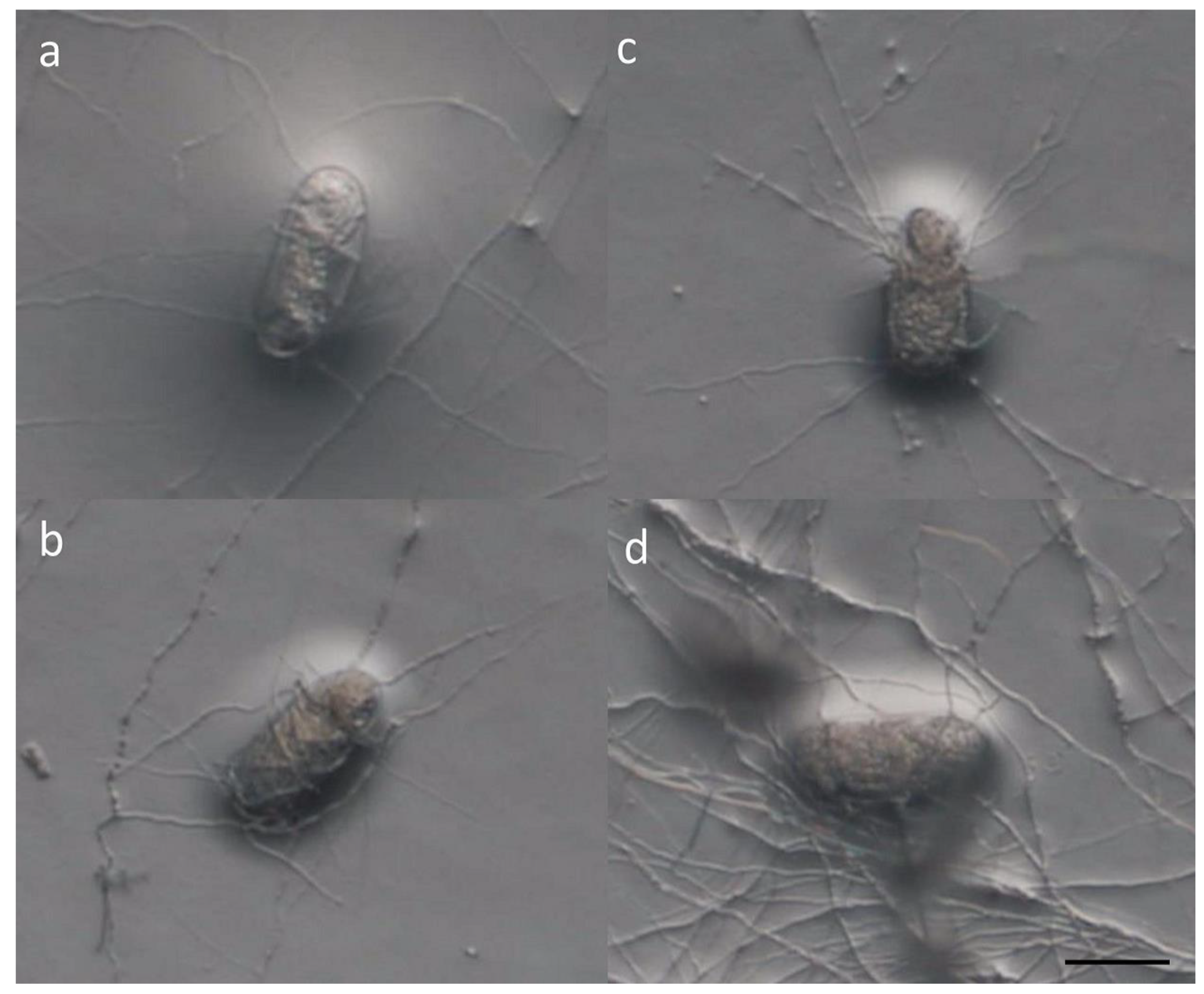
3.1. In Vitro Experiments
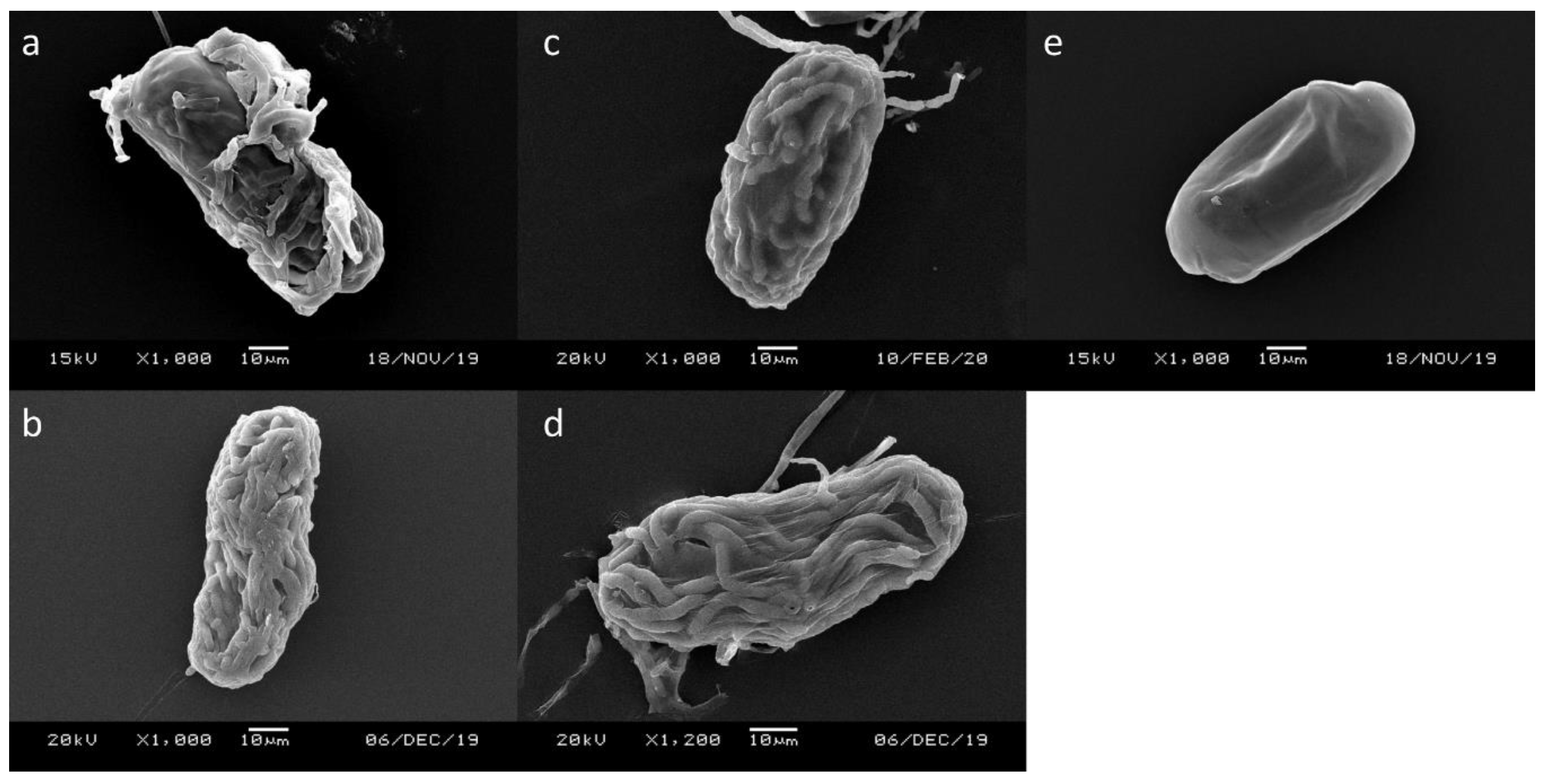
3.2. Scanning Electronic Microscope (SEM)
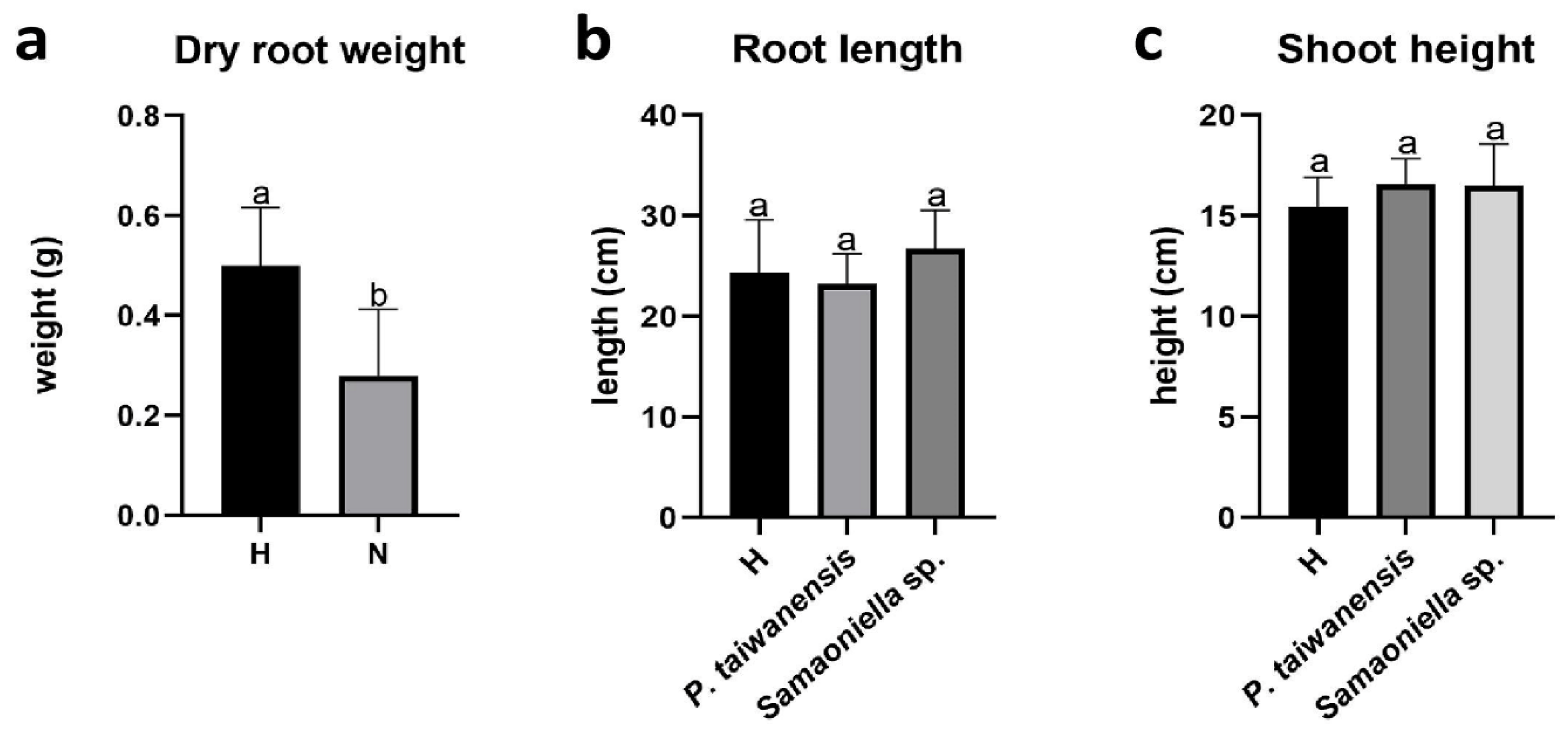
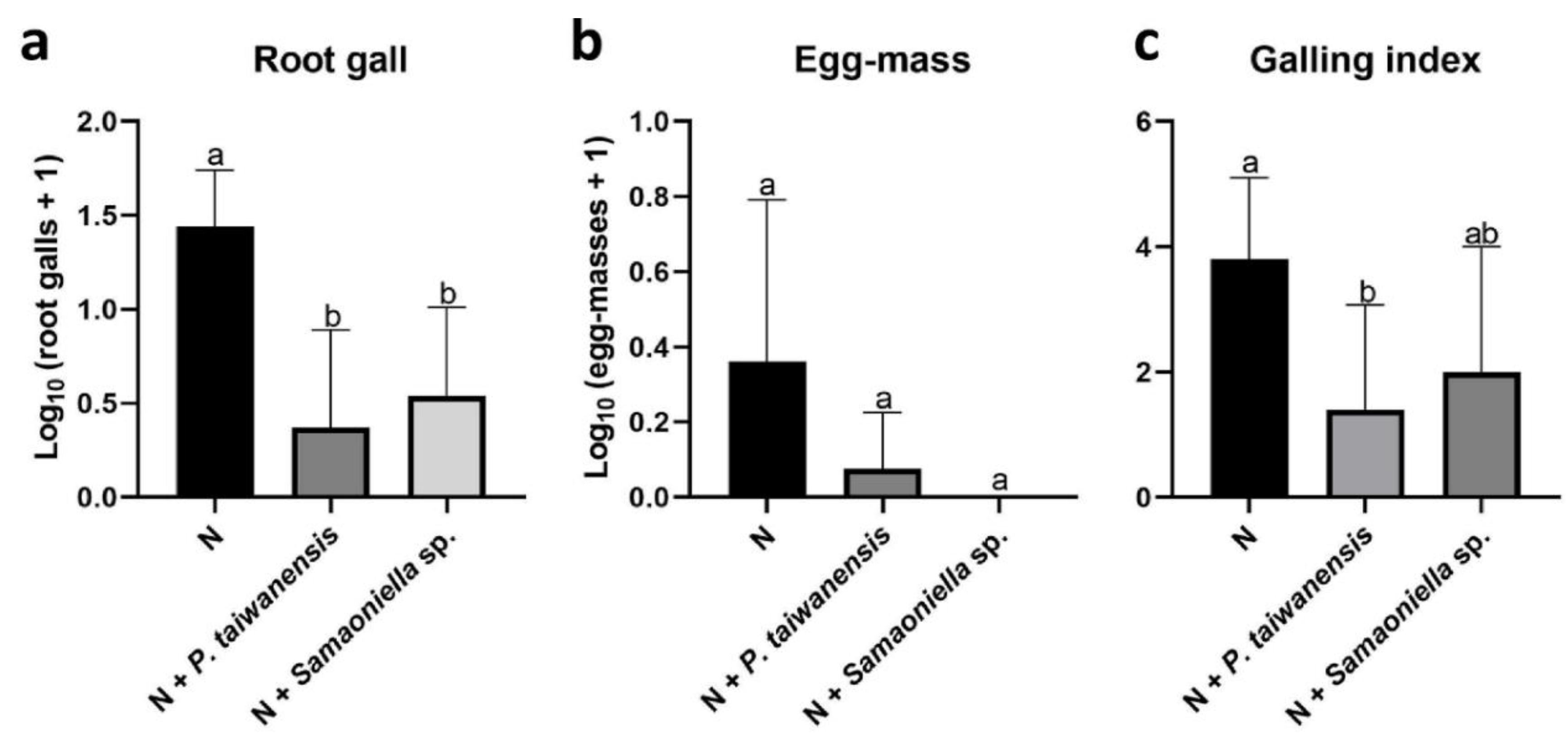
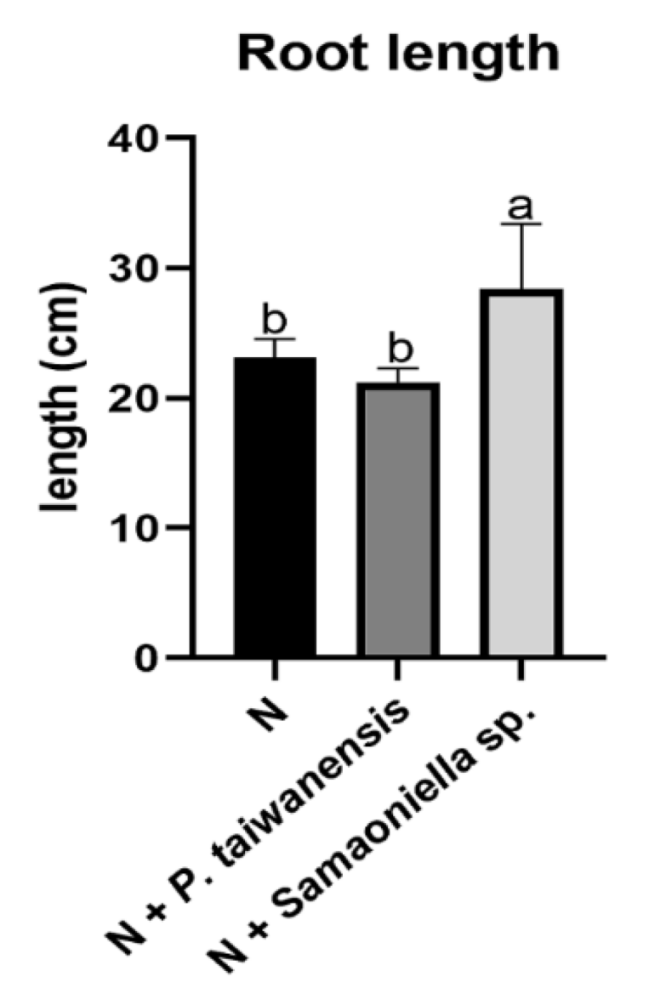
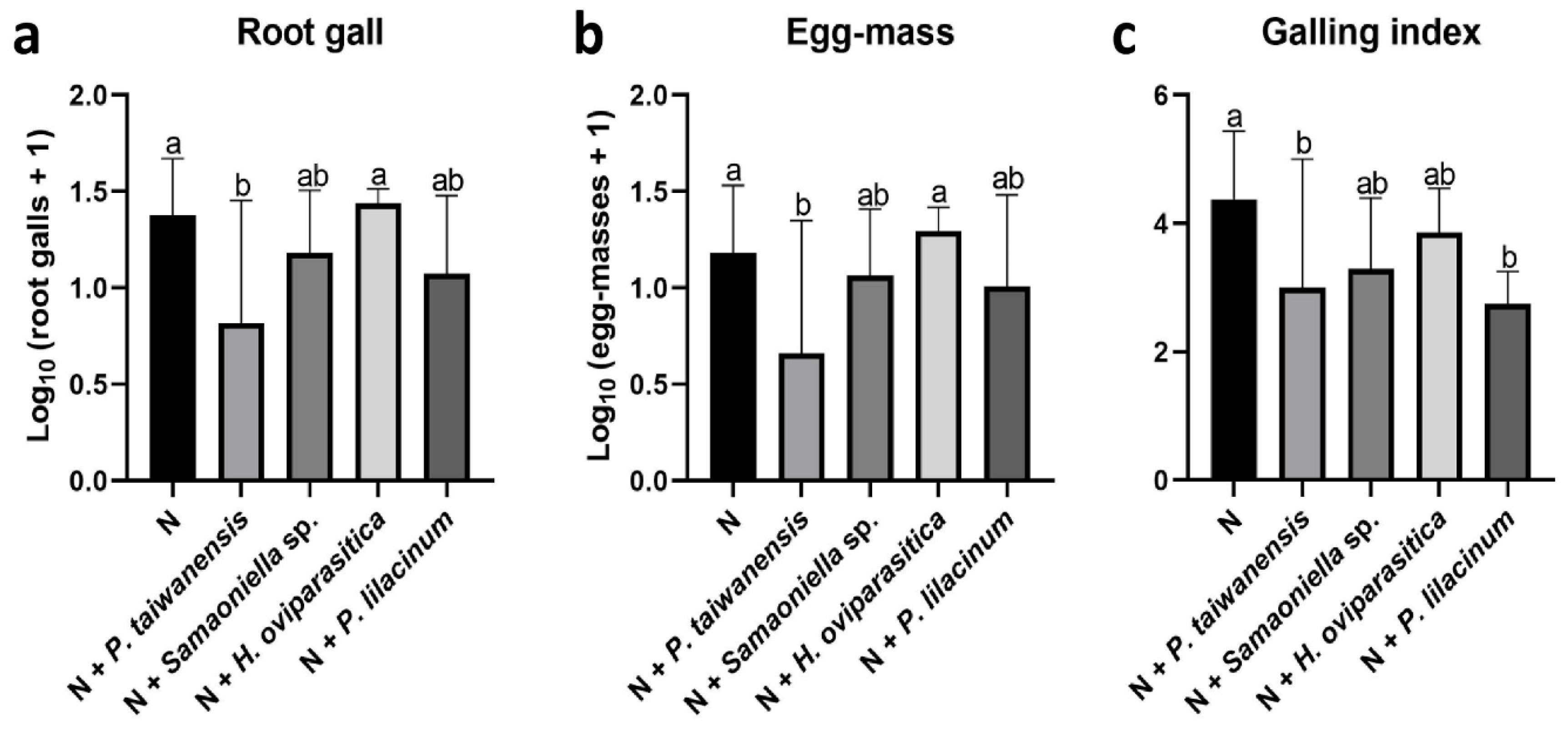
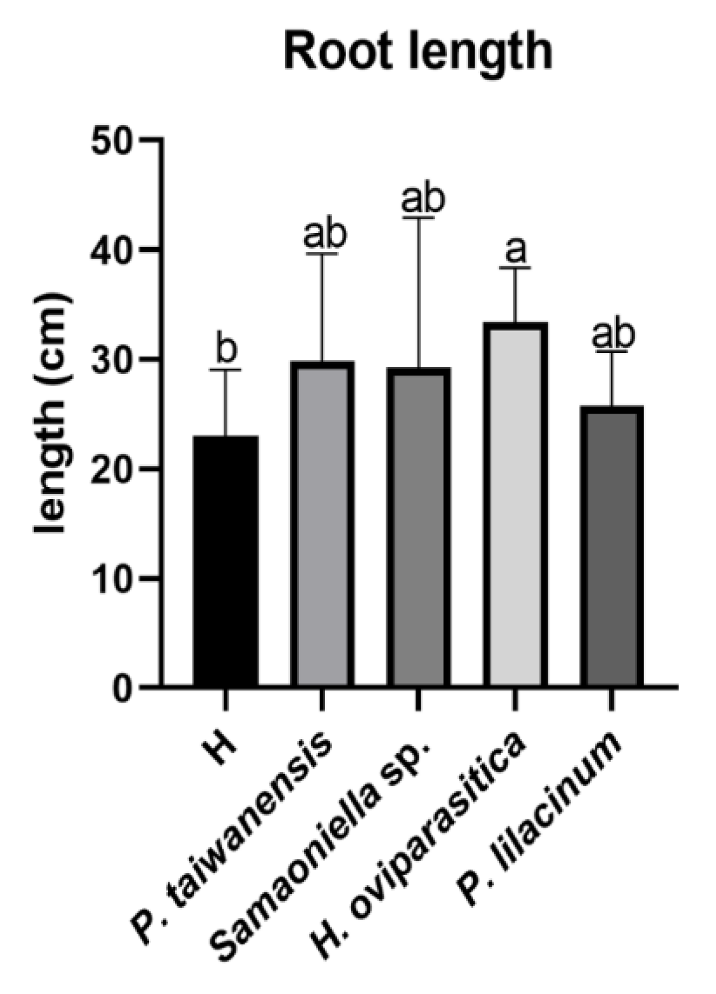
3.3. Greenhouse Trials
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rich, J.R.; Brito, J.A.; Kaur, R.; Ferrell, J.A. Weed species as hosts of Meloidogyne: A review. Nematropica 2009, 39, 157–185. [Google Scholar]
- Wesemael, W.; Viaene, N.; Moens, M. Root-knot nematodes (Meloidogyne spp.) in Europe. Nematology 2011, 13, 3–16. [Google Scholar] [CrossRef]
- Singh, S.; Singh, B.; Singh, A.P. Nematodes: A Threat to Sustainability of Agriculture. Procedia Environ. Sci. 2015, 29, 215–216. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, W.R.; Taylor, D.P. Plant Nematology; Reinhold Publishing Corporation: New York, NY, USA, 1967. [Google Scholar]
- EPPO. An Emerging Root-Knot Nematode, Meloidogyne enterolobii: Addition to the EPPO Alert List. EPPO Reporting Service. 2008, p. 5. Available online: Gd.eppo.int/reporting/article-690 (accessed on 31 March 2020).
- Japan Plant Protection Station. Japan Plant Protection Act. List of the Plants Subject to Field Inspection in Exporting Countries. 2019. Available online: www.maff.go.jp/pps (accessed on 29 January 2020).
- Jindapunnapat, K.; Chinnasri, B.; Kwankuae, S. Biological control of root-knot nematodes Meloidogyne enterolobii in guava by the fungus Trichoderma harzianum. J. Dev. Sustain. Agric. 2013, 8, 110–118. [Google Scholar]
- Ye, W.M.; Koenning, S.R.; Zhuo, K.; Liao, J.L. First report of Meloidogyne enterolobii on cotton and soybean in North Carolina, United States. Plant Dis. 2013, 97, 1262. [Google Scholar] [CrossRef] [PubMed]
- Rutter, W.B.; Skantar, A.M.; Handoo, Z.A.; Mueller, J.D.; Aultman, S.P.; Agudelo, P. Meloidogyne enterolobii found infecting root-knot nematode resistant sweetpotato in South Carolina, United States. Plant Dis. 2018, 103, 775. [Google Scholar] [CrossRef]
- Carneiro, R.; Cirotto, P.; Quintanilha, A.; Silva, D.; Carneiro, R. Resistance to Meloidogyne mayaguensis in Psidium spp. accessions and their grafting compatibility with P. guajava cv. Paluma. Fitopatol. Bras. 2007, 32, 281–284. [Google Scholar] [CrossRef]
- Freitas, V.M.; Silva, J.G.P.; Gomes, C.B.; Castro, J.M.C.; Correa, V.R.; Carneiro, R.M.D.G. Host status of selected cultivated fruit crops to Meloidogyne enterolobii. Eur. J. Plant Pathol. 2017, 148, 307–319. [Google Scholar] [CrossRef]
- Kiewnick, S.; Karssen, G.; Brito, J.A.; Oggenfuss, M.; Frey, J.E. First report of root-knot nematode Meloidogyne enterolobii on tomato and cucumber in Switzerland. Plant Dis. 2008, 92, 1370. [Google Scholar] [CrossRef]
- Minuto, A.; Gullino, M.; Lamberti, F.; D’Addabbo, T.; Tescari, E.; Ajwa, H.; Garibaldi, A. Application of an emulsifiable mixture of 1,3-dichloropropene and chloropicrin against root knot nematodes and soilborne fungi for greenhouse tomatoes in Italy. Crop Prot. 2006, 25, 1244–1252. [Google Scholar] [CrossRef]
- Ibekwe, A.M.; Papiernik, S.K.; Gan, J.; Yates, S.R.; Yang, C.H.; Crowley, D.E. Impact of fumigants on soil microbial communities. Appl. Environ. Microbiol. 2001, 67, 3245–3257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, J.A.; Stanley, J.D.; Kaur, R.; Cetintas, R.; Di Vito, M.; Thies, J.A.; Dickson, D.W. Effects of the Mi-1, N and Tabasco genes on infection and reproduction of Meloidogyne mayaguensis on tomato and pepper genotypes. J. Nematol. 2007, 39, 327–332. [Google Scholar] [PubMed]
- Stirling, G.R. Biological Control of Plant Parasitic Nematodes: SOIL Ecosystem Management in Sustainable Agriculture, 2nd ed.; CAB International: Wallingford, UK, 2014. [Google Scholar]
- Lopez-Llorca, L.V.; Maciá-Vicente, J.G.; Jansson, H.B. Mode of action and interactions of nematophagous fungi. In Integrated Management of Plant Pests and Diseases; Ciancio, A., Mukerji, K.G., Eds.; Springer: Dordrecht, The Netherlands, 2008; Volume 2, pp. 51–76. [Google Scholar]
- Silva, S.D.; Carneiro, R.M.D.G.; Faria, M.; Souza, D.A.; Monnerat, R.G.; Lopes, R.B. Evaluation of Pochonia chlamydosporia and Purpureocillium lilacinum for suppression of Meloidogyne enterolobii on tomato and banana. J. Nematol. 2017, 49, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thongkaewyuan, A.; Chairin, T. Biocontrol of Meloidogyne incognita by Metarhizium guizhouense and its protease. Biol. Control 2018, 126, 142–146. [Google Scholar] [CrossRef]
- Kepenekci, I.; Saglam, H.D.; Oksal, E.; Yanar, D.; Yanar, Y. Nematicidal activity of Beauveria bassiana (Bals.-Criv.) Vuill. against root-knot nematodes on tomato grown under natural conditions. Egypt. J. Biol. Pest Control 2017, 27, 117–120. [Google Scholar]
- Yang, J.-I.; Stadler, M.; Chuang, W.-Y.; Wu, S.; Ariyawansa, H.A. In vitro inferred interactions of selected entomopathogenic fungi from Taiwan and eggs of Meloidogyne graminicola. Mycol. Prog. 2020, 19, 97–109. [Google Scholar] [CrossRef]
- Bidochka, M.J.; Khachatourians, G.G. Purification and properties of an extracellular protease produced by the entomopathogenic fungus Beauveria bassiana. Appl. Environ. Microbiol. 1987, 53, 1679–1684. [Google Scholar] [CrossRef] [Green Version]
- Leger, R.J.S.; Charnley, A.K.; Cooper, R.M. Characterization of cuticle-degrading proteases produced by the entomopathogen Metarhizium anisopliae. Arch. Biochem. Biophys. 1987, 253, 221–232. [Google Scholar] [CrossRef]
- Lei, Y.; Lü, L.; He, Y. Study on induction of enzyme activity of entomopathogenic fungus Isaria fumosorosea SCAU-IFCF01. Huan Jing Kun Chong Xue Bao 2017, 39, 1000–1006. (In Chinese) [Google Scholar]
- Díaz-Godínez, G.; Candia-Sanchez, L.F.; Díaz, R.; Sánchez, C.; Villegas, E.; Monroy, G.; Hernández Velázquez, V.M. Activity of lipases, chitinases and proteases of the entomopathogenic fungus, Metarhizium anisopliae developed in different culture media. In Biotechnology Summit 2016, 3rd ed.; Fernández-Luqueño, F., López-Valdez, F., Santos Villalobos, S.D.L., Eds.; Cinvestav: San Pedro Zacatenco, Mexico, 2016; pp. 155–159. [Google Scholar]
- Morton, O.; Hirsch, P.; Kerry, B. Infection of plant-parasitic nematodes by nematophagous fungi—A review of the application of molecular biology to understand infection processes and to improve biological control. Nematology 2004, 6, 161–170. [Google Scholar] [CrossRef]
- Bird, A.F.; McClure, M.A. The tylenchid (Nematoda) eggshell: Structure, composition and permeability. Parasitology 1976, 72, 19–28. [Google Scholar] [CrossRef]
- Baral, H.-O.; Weber, E.; Gams, W.; Hagedorn, G.; Liu, B.; Liu, X.; Marson, G.; Marvanová, L.; Stadler, M.; Weiß, M. Generic names in the Orbiliaceae (Orbiliomycetes) and recommendations on which names should be protected or suppressed. Mycol. Prog. 2018, 17, 5–31. [Google Scholar] [CrossRef] [Green Version]
- Kiewnick, S.; Sikora, R.A. Biological control of the root-knot nematode Meloidogyne incognita by Paecilomyces lilacinus strain 251. Biol. Control 2006, 38, 179–187. [Google Scholar] [CrossRef]
- Olatinwo, R.; Yin, B.; Becker, J.O.; Borneman, J. Suppression of the plant-parasitic nematode Heterodera schachtii by the fungus Dactylella oviparasitica. Phytopathology 2006, 96, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith Becker, J.; Borneman, J.; Becker, J.O. Dactylella oviparasitica parasitism of the sugar beet cyst nematode observed in trixenic culture plates. Biol. Control 2013, 64, 51–56. [Google Scholar] [CrossRef]
- Hu, M.X.; Zhuo, K.; Liao, J.L. Multiplex PCR for the simultaneous identification and detection of Meloidogyne incognita, M. enterolobii, and M. javanica using DNA extracted directly from individual galls. Phytopathology 2011, 101, 1270–1277. [Google Scholar] [CrossRef] [Green Version]
- Thies, J.; Mueller, J.D.; Fery, R. Effectiveness of resistance to southern root-knot nematode in ‘Carolina Cayenne’ pepper in greenhouse, microplot, and field tests. J. Am. Soc. Hortic. Sci. 1997, 122, 200–204. [Google Scholar] [CrossRef]
- Yang, J.-I.; Benecke, S.; Jeske, D.R.; Rocha, F.S.; Smith Becker, J.; Timper, P.; Becker, J.O.; Borneman, J. Population dynamics of Dactylella oviparasitica and Heterodera schachtii: Toward a decision model for sugar beet planting. J. Nematol. 2012, 44, 237–244. [Google Scholar]
- Stirling, G.R.; Mankau, R. Mode of parasitism of Meloidogyne and other nematode eggs by Dactylella oviparasitica. J. Nematol. 1979, 11, 282–288. [Google Scholar]
- Stirling, G.R.; Mankau, R. Parasitism of Meloidogyne eggs by a new fungal parasite. J. Nematol. 1978, 10, 236–240. [Google Scholar]
- Kepenekci, I.; Oksal, E.; Saglam, H.D.; Atay, T.; Tulek, A.; Evlice, E. Identification of Turkish isolate of the entomopathogenic fungi, Purpureocillium lilacinum (syn: Paecilomyces lilacinus) and its effect on potato pests, Phthorimaea operculella (Zeller) (Lepidoptera: Gelechiidae) and Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae). Egypt. J. Biol. Pest Control 2015, 25, 121–127. [Google Scholar]
- Toledo-Hernández, R.A.; Holguín-Meléndez, F.; Huerta-Palacios, G.; Liedo, P.; Toledo, J.; Valle-Mora, J. Pathogenicity and virulence of Purpureocillium lilacinum (Hypocreales: Ophiocordycipitaceae) on Mexican fruit fly adults. Fla. Entomol. 2019, 102, 309–314. [Google Scholar] [CrossRef] [Green Version]
- Yüksel, E.; Açıkgöz, C.; Demirci, F.; Muștu, M. Effects of the entomopathogenic fungi, Beauveria bassiana, Isaria farinosa and Purpureocillium lilacinum, on eggs of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). /Entomopatojen funguslar Beauveria bassiana, Isaria farinosa ve Purpureocillium lilacinum’un Domates Yaprak galeri Güvesi Tuta absoluta’ nın yumurta dönemine etkileri. Türkiye Biyolojik Mücadele Dergisi 2017, 8, 39–47. [Google Scholar]
- Cabanillas, E.; Barker, K.R. Impact of Paecilomyces lilacinus inoculum level and application time on control of Meloidogyne incognita on tomato. J. Nematol. 1989, 21, 115–120. [Google Scholar]
- Kiewnick, S.; Sikora, R. Efficacy of Paecilomyces lilacinus (strain 251) for the control of root-knot nematodes. Commun. Agric. Appl. Biol. Sci. 2003, 68, 123–128. [Google Scholar]
- Heydari, A.; Pessarakli, M. A review on biological control of fungal plant pathogens using microbial antagonists. J. Biol. Sci. 2010, 10, 273–290. [Google Scholar] [CrossRef] [Green Version]
- Luambano, N.D.; Manzanilla-López, R.H.; Kimenju, J.W.; Powers, S.J.; Narla, R.D.; Wanjohi, W.J.; Kerry, B.R. Effect of temperature, pH, carbon and nitrogen ratios on the parasitic activity of Pochonia chlamydosporia on Meloidogyne incognita. Biol. Control 2015, 80, 23–29. [Google Scholar] [CrossRef]
- Takayuki, M.; Yuji, O. Tomato culture filtrate stimulates hatching and activity of Meloidogyne incognita juveniles. Nematology 2009, 11, 51–61. [Google Scholar]
- Wesemael, W.; Perry, R.; Moens, M.J.N.V. The influence of root diffusate and host age on hatching of the root-knot nematodes, Meloidogyne chitwoodi and M. fallax. Nematology 2006, 8, 895–902. [Google Scholar] [CrossRef]
- Yankova, V.; Markova, D.; Naidenov, M.; Arnaoudov, B. Management of root-knot nematodes (Meloidogyne spp.) in greenhouse cucumbers using microbial products. Türk Tarım Doğa Bilimleri Derg. 2014, 1 (Özel Sayı-2), 1569–1573. [Google Scholar]
- Monfort, E.; Lopez-Llorca, L.V.; Jansson, H.-B.; Salinas, J. In vitro soil receptivity assays to egg-parasitic nematophagous fungi. Mycol. Prog. 2006, 5, 18–23. [Google Scholar] [CrossRef]
- Yang, J.-I.; Loffredo, A.; Borneman, J.; Becker, J.O. Biocontrol efficacy among strains of Pochonia chlamydosporia obtained from a root-knot nematode suppressive soil. J. Nematol. 2012, 44, 67–71. [Google Scholar] [PubMed]
- Carneiro, R.M.D.G.; Hidalgo-Díaz, L.; Martins, I.; Silva, K.F.A.d.S.; Sousa, M.G.d.; Tigano, M.S. Effect of nematophagous fungi on reproduction of Meloidogyne enterolobii on guava (Psidium guajava) plants. Nematology 2011, 13, 721–728. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungi | Egg Hatching Rate (%) 1 | Egg Infection Rate (%) 1 |
|---|---|---|
| Paraboeremia taiwanensis NTUCC 17–013 | 46.33 ± 8.10 a | 47.83 ± 9.93 a |
| Samsoniella sp. NTUCC 18–159 | 47.16 ± 8.13 a | 47.50 ± 9.18 a |
| Hyalorbilia oviparasitica strain DoUCR50 | 41.83 ± 5.69 a | 51.17 ± 7.15 a |
| Purpureocillium lilacinum stain PL251 | 40.33 ± 3.01 a | 55.83 ± 1.26 a |
| No fungi control | 86.67 ± 1.04 b | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Y.-J.; Ariyawansa, H.A.; Becker, J.O.; Yang, J.-i. The Evaluation of Egg-Parasitic Fungi Paraboeremia taiwanensis and Samsoniella sp. for the Biological Control of Meloidogyne enterolobii on Chinese Cabbage. Microorganisms 2020, 8, 828. https://doi.org/10.3390/microorganisms8060828
Liang Y-J, Ariyawansa HA, Becker JO, Yang J-i. The Evaluation of Egg-Parasitic Fungi Paraboeremia taiwanensis and Samsoniella sp. for the Biological Control of Meloidogyne enterolobii on Chinese Cabbage. Microorganisms. 2020; 8(6):828. https://doi.org/10.3390/microorganisms8060828
Chicago/Turabian StyleLiang, Yu-Jie, Hiran A. Ariyawansa, J. Ole Becker, and Jiue-in Yang. 2020. "The Evaluation of Egg-Parasitic Fungi Paraboeremia taiwanensis and Samsoniella sp. for the Biological Control of Meloidogyne enterolobii on Chinese Cabbage" Microorganisms 8, no. 6: 828. https://doi.org/10.3390/microorganisms8060828