Virulence Potential of a Multidrug-Resistant Escherichia coli Strain Belonging to the Emerging Clonal Group ST101-B1 Isolated from Bloodstream Infection

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain
2.2. Total DNA Extraction, Whole-Genome Sequencing (WGS), and Genome Assembly
2.3. Genomic Analyses and Annotation
2.4. Data Availability
2.5. Serum Agglutination Assay for Typing the O and H Antigens
2.6. Antimicrobial Susceptibility Testing
2.7. Plasmid DNA Extraction and Analysis
2.8. In Silico Plasmid Analysis
2.9. Conjugation Assay
2.10. Characterization of Conjugative Plasmids
2.11. Determination of the Lowest Bacterial Inoculum Which Was Resistant to Human Serum Complement
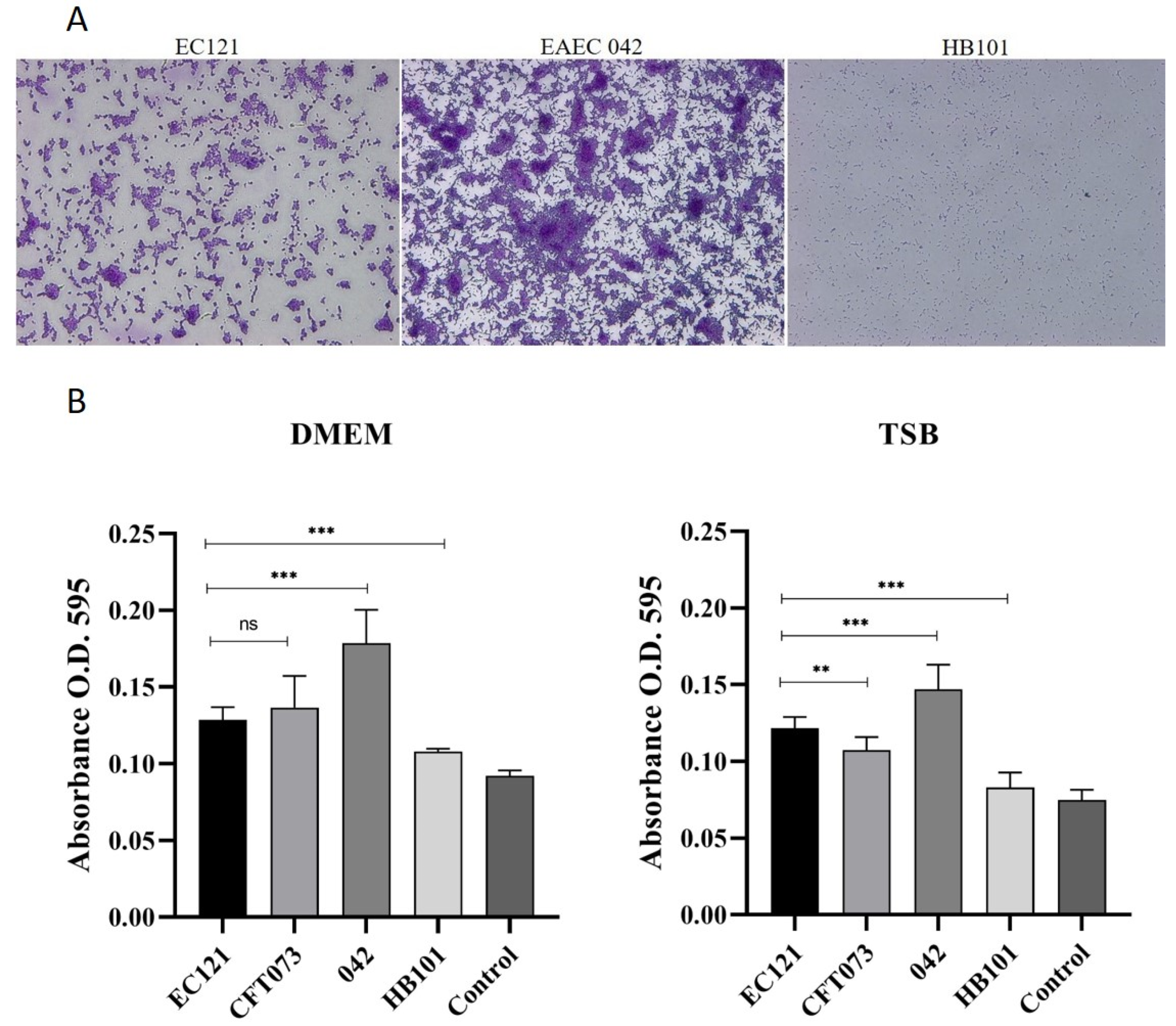
2.12. Biofilm Formation on Abiotic Surfaces
2.13. Cell Culture and Maintenance
2.14. Adherence Assays in HeLa, Caco-2, and T24 Cells
2.15. Short Period Interaction and Invasion Assay in Caco-2 and T24 Cells
2.16. In Vivo Assay in Galleria Mellonella Virulence Model
2.17. Statistical Analyses
2.18. Ethics Statements
3. Results
3.1. Genetic Characterization and In Silico Analysis
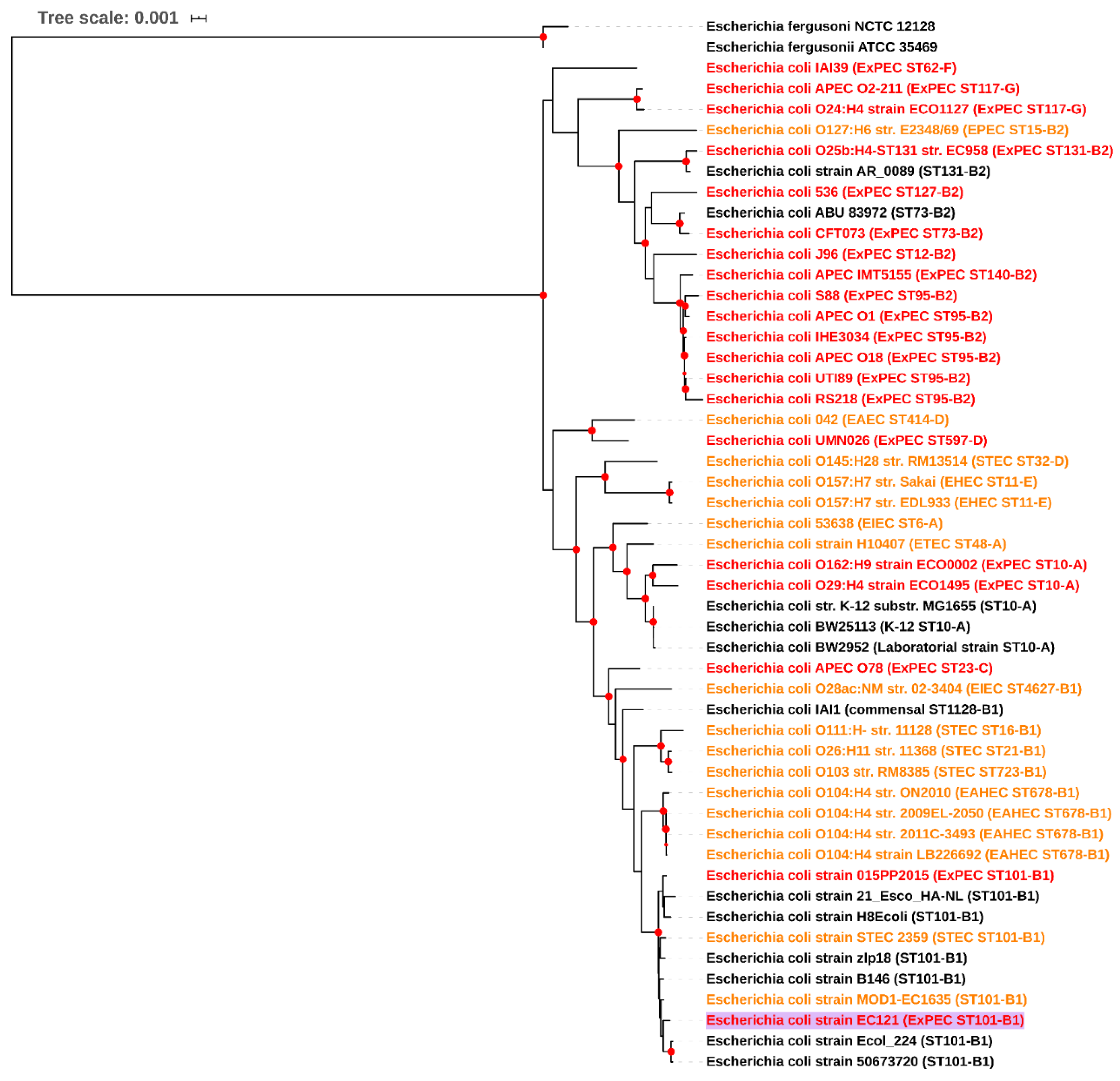
3.1.1. EC121 Belongs to Serotype O154:H25, ST101-B1, and is Related to Diarrheagenic E. coli
3.1.2. EC121 Harbors Genes Involved in Virulence and Stress Response
3.1.3. No Complete Phage Sequences Were Detected in the EC121 Strain
3.1.4. EC121 Has Multiple Resistance Genes, and Several Efflux-Pumps Compatible with its Antimicrobial Susceptibility Profile
3.1.5. The EC121 Strain Harbors Multiple Plasmids
3.2. Virulence Phenotype
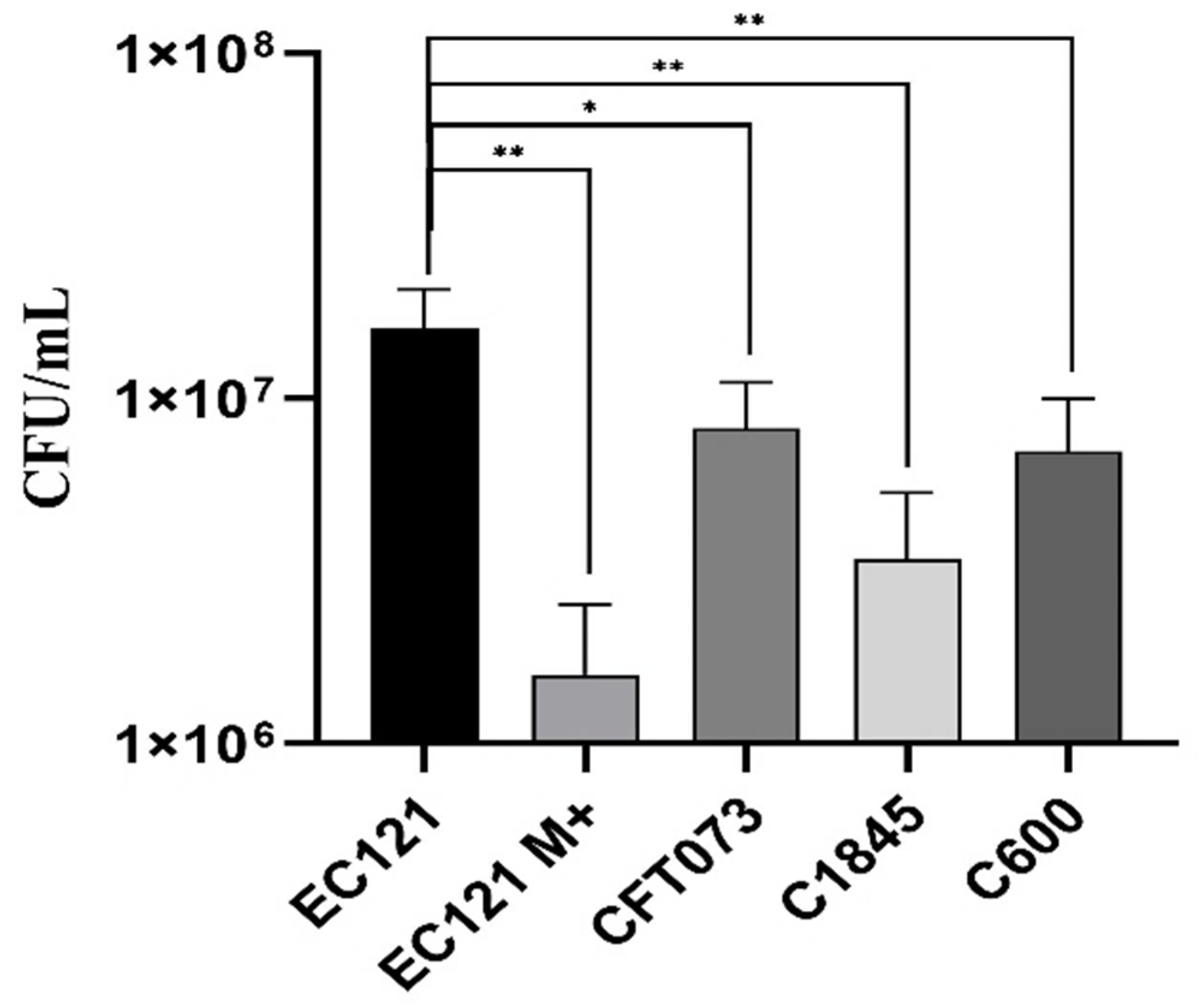
3.2.1. The EC121 Strain Resists the Bactericidal Activity of the Human Complement System and Adheres to Abiotic Surfaces
3.2.2. The EC121 Strain Adheres to and Invades Epithelial Cell Lineages
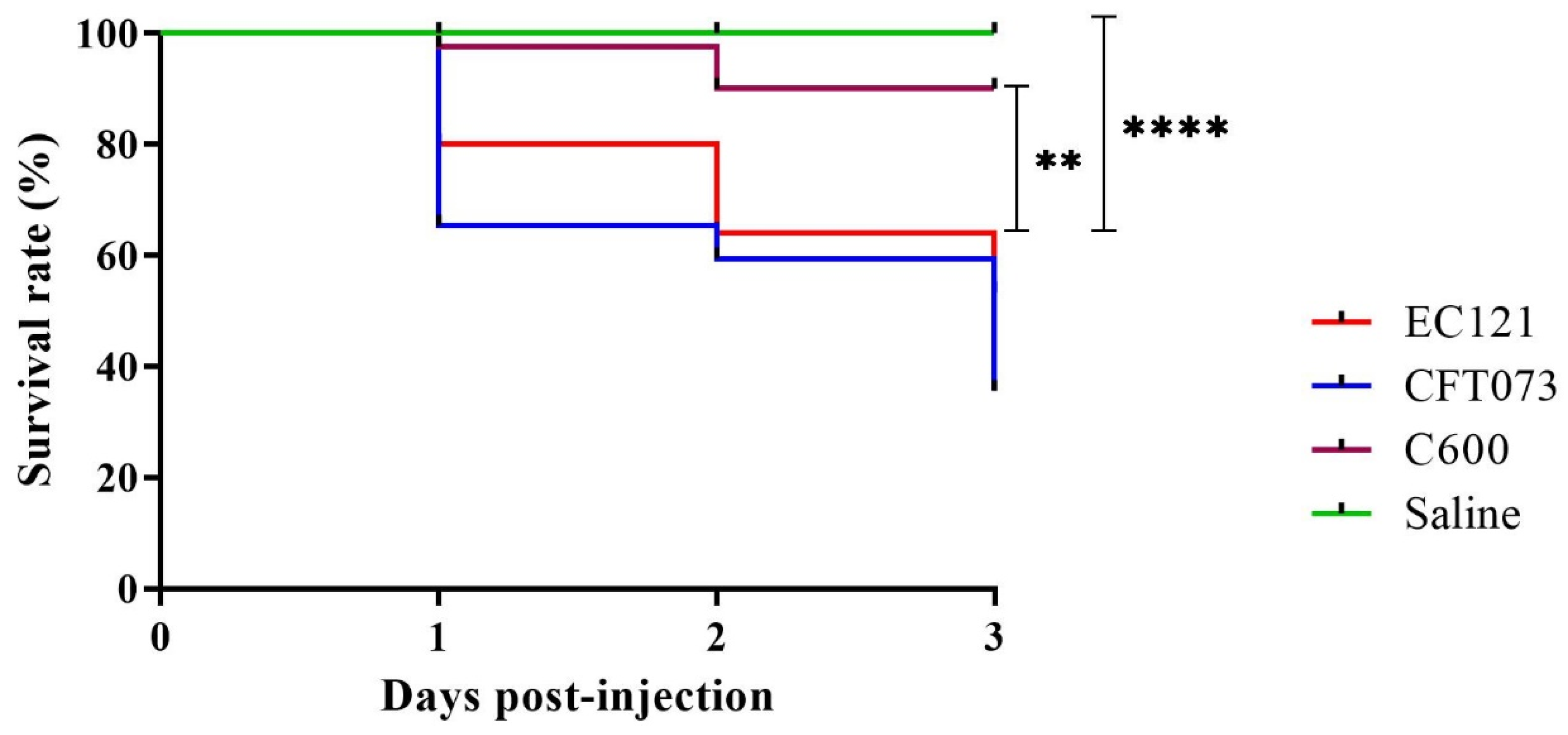
3.2.3. EC121 Strain Is Virulent in the Galleria mellonella Virulence Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Biedenbach, D.J.; Moet, G.J.; Jones, R.N. Occurrence and antimicrobial resistance pattern comparisons among bloodstream infection isolates from the SENTRY Antimicrobial Surveillance Program (1997–2002). Diagn. Microbiol. Infect. Dis. 2004, 50, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Temkin, E.; Fallach, N.; Almagor, J.; Gladstone, B.P.; Tacconelli, E.; Carmeli, Y. DRIVE-AB Consortium Estimating the number of infections caused by antibiotic-resistant Escherichia coli and Klebsiella pneumoniae in 2014: A modelling study. Lancet. Glob. Heal. 2018, 6, e969–e979. [Google Scholar] [CrossRef] [Green Version]
- Tu, B.; Liu, S.X.; Wu, D.; Zhang, X.; Shi, L.; Xie, Y.X.; Zhao, P.; Qin, E.Q. Clinical features of community-acquired bloodstream infection due to Gram-negative bacilli in patients with liver cirrhosis. Zhonghua Gan Zang Bing Za Zhi 2018, 26, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Satwani, P.; Freedman, J.L.; Chaudhury, S.; Jin, Z.; Levinson, A.; Foca, M.D.; Krajewski, J.; Sahdev, I.; Talekar, M.K.; Gardenswartz, A.; et al. A Multicenter Study of Bacterial Blood Stream Infections in Pediatric Allogeneic Hematopoietic Cell Transplantation Recipients: The Role of Acute Gastrointestinal Graft-versus-Host Disease. Biol. Blood Marrow Transplant. 2017, 23, 642–647. [Google Scholar] [CrossRef] [Green Version]
- Nivesvivat, T.; Piyaraj, P.; Thunyaharn, S.; Watanaveeradej, V.; Suwanpakdee, D. Clinical epidemiology, risk factors and treatment outcomes of extended-spectrum beta-lactamase producing Enterobacteriaceae bacteremia among children in a Tertiary Care Hospital, Bangkok, Thailand. BMC Res. Notes 2018, 11, 624. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization GLASS. Global Antimicrobial Resistance Surveillance System (GLASS) Report; WHO: Geneva, Switzerland, 2018; Available online: https://www.who.int/glass/resources/publications/early-implementation-report/en/ (accessed on 17 June 2019).
- Landman, W.J.M.; van Eck, J.H.H. The incidence and economic impact of the Escherichia coli peritonitis syndrome in Dutch poultry farming. Avian. Pathol. 2015, 44, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Kaye, K.S.; Marchaim, D.; Chen, T.-Y.; Baures, T.; Anderson, D.J.; Choi, Y.; Sloane, R.; Schmader, K.E. Effect of Nosocomial Bloodstream Infections on Mortality, Length of Stay, and Hospital Costs in Older Adults. J. Am. Geriatr. Soc. 2014, 62, 306–311. [Google Scholar] [CrossRef]
- Barriere, S.L. Clinical, economic and societal impact of antibiotic resistance. Expert Opin. Pharmacother. 2015, 16, 151–153. [Google Scholar] [CrossRef] [Green Version]
- Ciorba, V.; Odone, A.; Veronesi, L.; Pasquarella, C.; Signorelli, C. Antibiotic resistance as a major public health concern: Epidemiology and economic impact. Ann. Ig. 2015, 27, 562–579. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Russo, T.A. Extraintestinal pathogenic Escherichia coli: “The other bad E. coli.”. J. Lab. Clin. Med. 2002, 139, 155–162. [Google Scholar] [CrossRef]
- Johnson, J.R.; Russo, T.A. Molecular Epidemiology of Extraintestinal Pathogenic Escherichia coli. EcoSal Plus 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Murray, A.C.; Gajewski, A.; Sullivan, M.; Snippes, P.; Kuskowski, M.A.; Smith, K.E. Isolation and molecular characterization of nalidixic acid-resistant extraintestinal pathogenic Escherichia coli from retail chicken products. Antimicrob. Agents Chemother. 2003, 47, 2161–2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spurbeck, R.R.; Dinh, P.C.; Walk, S.T.; Stapleton, A.E.; Hooton, T.M.; Nolan, L.K.; Kim, K.S.; Johnson, J.R.; Mobley, H.L.T. Escherichia coli Isolates That Carry vat, fyuA, chuA, and yfcV Efficiently Colonize the Urinary Tract. Infect. Immun. 2012, 80, 4115–4122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, D.A.; Mills, G.; Johnson, J.R.; Porter, S.; Wiles, S. In vivo correlates of molecularly inferred virulence among extraintestinal pathogenic Escherichia coli (ExPEC) in the wax moth Galleria mellonella model system. Virulence 2014, 5, 388–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.C.M.; Zidko, A.C.M.; Pignatari, A.C.; Silva, R.M. Assessing the diversity of the virulence potential of Escherichia coli isolated from bacteremia in São Paulo, Brazil. Brazilian J. Med. Biol. Res. 2013, 46, 968–973. [Google Scholar] [CrossRef] [Green Version]
- Leimbach, A.; Hacker, J.; Dobrindt, U.E. E. coli as an all-rounder: The thin line between commensalism and pathogenicity. Curr. Top. Microbiol. Immunol. 2013, 358, 3–32. [Google Scholar] [CrossRef]
- Wang, Y.; Kim, K.S. Role of OmpA and IbeB in Escherichia coli K1 Invasion of Brain Microvascular Endothelial Cells In Vitro and In Vivo. Pediatr. Res. 2002, 51, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.H.; Chen, Y.H.; Fu, Q.; Stins, M.; Wang, Y.; Wass, C.; Kim, K.S. Identification and characterization of an Escherichia coli invasion gene locus, ibeB, required for penetration of brain microvascular endothelial cells. Infect. Immun. 1999, 67, 2103–2109. [Google Scholar] [CrossRef] [Green Version]
- Mitra, A.; Palaniyandi, S.; Herren, C.D.; Zhu, X.; Mukhopadhyay, S. Pleiotropic Roles of uvrY on Biofilm Formation, Motility and Virulence in Uropathogenic Escherichia coli CFT073. PLoS ONE 2013, 8, e55492. [Google Scholar] [CrossRef]
- Kim, W.J.; Shea, A.E.; Kim, J.H.; Daaka, Y. Uropathogenic Escherichia coli invades bladder epithelial cells by activating kinase networks in host cells. J. Biol. Chem. 2018, 293, 16518–16527. [Google Scholar] [CrossRef] [Green Version]
- Kondratyeva, K.; Wollman, A.; Gerlitz, G.; Navon-Venezia, S. Adhesion and invasion to epithelial cells and motility of extended-spectrum β-lactamase-producing Escherichia coli reveal st131 superiority: A comparative in vitro study of extraintestinal pathogenic E. coli lineages. J. Med. Microbiol. 2017, 66, 1350–1357. [Google Scholar] [CrossRef] [PubMed]
- Saldaña, Z.; De La Cruz, M.A.; Carrillo-Casas, E.M.; Durán, L.; Zhang, Y.; Hernández-Castro, R.; Puente, J.L.; Daaka, Y.; Girón, J.A. Production of the Escherichia coli common pilus by uropathogenic E. coli is associated with adherence to HeLa and HTB-4 cells and invasion of mouse bladder urothelium. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R. Microbial virulence determinants and the pathogenesis of urinary tract infection. Infect. Dis. Clin. N. Am. 2003, 17, 261–278. [Google Scholar] [CrossRef]
- Buckles, E.L.; Wang, X.; Lane, M.C.; Lockatell, C.V.; Johnson, D.E.; Rasko, D.A.; Mobley, H.L.T.; Donnenberg, M.S. Role of the K2 Capsule in Escherichia coli Urinary Tract Infection and Serum Resistance. J. Infect. Dis. 2009, 199, 1689–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, J.; Ba-Thein, W.; Kumao, T.; Obata Yasuoka, M.; Akaza, H.; Hayshi, H. Type 1, P and S fimbriae, and afimbrial adhesin I are not essential for uropathogenic Escherichia coli to adhere to and invade bladder epithelial cells. FEMS Immunol. Med. Microbiol. 2002, 33, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Mittal, S.; Sharma, M.; Chaudhary, U. Biofilm and multidrug resistance in uropathogenic Escherichia coli. Pathog. Glob. Health 2015, 109, 26–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.C.M. Potencial de virulência e suscetibilidade a antimicrobianos de amostras de Escherichia coli isoladas de bacteremia. Sua relação com o estado de imunocompetência do paciente e a origem da infecção. Ph.D Thesis, Universidade Federal de São Paulo (UNIFESP), São Paulo, Brazil, 2013. [Google Scholar]
- Landraud, L.; Jauréguy, F.; Frapy, E.; Guigon, G.; Gouriou, S.; Carbonnelle, E.; Clermont, O.; Denamur, E.; Picard, B.; Lemichez, E.; et al. Severity of Escherichia coli bacteraemia is independent of the intrinsic virulence of the strains assessed in a mouse model. Clin. Microbiol. Infect. 2013, 19, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhang, J.; Zheng, B.; Wei, Z.; Shen, P.; Li, S.; Li, L.; Xiao, Y. Molecular epidemiology and genetic diversity of fluoroquinolone-resistant Escherichia coli isolates from patients with community-onset infections in 30 Chinese county hospitals. J. Clin. Microbiol. 2015, 53, 766–770. [Google Scholar] [CrossRef] [Green Version]
- Izdebski, R.; Baraniak, A.; Herda, M.; Fiett, J.; Bonten, M.J.M.; Carmeli, Y.; Goossens, H.; Hryniewicz, W.; Brun-buisson, C.; Gniadkowski, M.; et al. MLST reveals potentially high-risk international clones of Enterobacter cloacae. J. Antimicrob. Chemother. 2015, 70, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Brauner, J.; Hallin, M.; Deplano, A.; Mendonça, R.; Nonhoff, C.; Ryck, R.; Roisin, S.; Struelens, M.J.; Denis, O. Community-acquired methicillin-resistant Staphylococcus aureus clones circulating in Belgium from 2005 to 2009: Changing epidemiology. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 613–620. [Google Scholar] [CrossRef]
- Riley, L.W. Pandemic lineages of extraintestinal pathogenic Escherichia coli. Clin. Microbiol. Infect. 2014, 20, 380–390. [Google Scholar] [CrossRef] [Green Version]
- Roer, L.; Overballe-Petersen, S.; Hansen, F.; Schønning, K.; Wang, M.; Røder, B.L.; Hansen, D.S.; Justesen, U.S.; Andersen, L.P.; Fulgsang-Damgaard, D.; et al. Escherichia coli Sequence Type 410 Is Causing New International High-Risk Clones. mSphere 2018, 3, e00337-18. [Google Scholar] [CrossRef] [Green Version]
- Reineke, K.; Sevenich, R.; Hertwig, C.; Janßen, T.; Fröhling, A.; Knorr, D.; Wieler, L.H.; Schlüter, O. Comparative study on the high pressure inactivation behavior of the Shiga toxin-producing Escherichia coli O104:H4 and O157:H7 outbreak strains and a non-pathogenic surrogate. Food Microbiol. 2015, 46, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, C.A.; Walsh, C.; Karczmarczyk, M.; O’Brien, S.; Akasheh, N.; Quirke, M.; Farrell-Ward, S.; Buckley, T.; Fogherty, U.; Kavanagh, K.; et al. Multi-drug resistant Escherichia coli in diarrhoeagenic foals: Pulsotyping, phylotyping, serotyping, antibiotic resistance and virulence profiling. Vet. Microbiol. 2018, 223, 144–152. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization WHO. Global Action Plan on Antimicrobial Resistance; WHO: Geneva, Switzerland, 2017 20 June; Available online: https://www.who.int/antimicrobial-resistance/publications/global-action-plan/en/ (accessed on 17 June 2019).
- Shaik, S.; Ranjan, A.; Tiwari, S.K.; Hussain, A.; Nandanwar, N.; Kumar, N.; Jadhav, S.; Semmler, T.; Baddam, R.; Islam, M.A.; et al. Comparative Genomic Analysis of Globally Dominant ST131 Clone with Other Epidemiologically Successful Extraintestinal Pathogenic Escherichia coli (ExPEC) Lineages. MBio 2017, 8, e01596-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjan, A.; Shaik, S.; Mondal, A.; Nandanwar, N.; Hussain, A.; Semmler, T.; Kumar, N.; Tiwari, S.K.; Jadhav, S.; Wieler, L.H.; et al. Molecular Epidemiology and Genome Dynamics of New Delhi Metallo-β-Lactamase-Producing Extraintestinal Pathogenic Escherichia coli Strains from India. Antimicrob. Agents Chemother. 2016, 60, 6795–6805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Savov, E.; Nazli, A.; Trifonova, A.; Todorova, I.; Gergova, I.; Nordmann, P. Outbreak caused by NDM-1- and RmtB-producing Escherichia coli in Bulgaria. Antimicrob. Agents Chemother. 2014, 58, 2472–2474. [Google Scholar] [CrossRef] [Green Version]
- Mora, A.; Blanco, M.; López, C.; Mamani, R.; Blanco, J.E.J.; Alonso, M.P.; García-Garrote, F.; Dahbi, G.; Herrera, A.; Fernández, A.; et al. Emergence of clonal groups O1:HNM-D-ST59, O15:H1-D-ST393, O20:H34/HNM-D-ST354, O25b:H4-B2-ST131 and ONT:H21,42-B1-ST101 among CTX-M-14-producing Escherichia coli clinical isolates in Galicia, northwest Spain. Int. J. Antimicrob. Agents 2011, 37, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global extraintestinal pathogenic Escherichia coli (Expec) lineages. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef]
- World Health Organization WHO. No Time to Wait: Securing the Future from Drug-Resistant Infections; WHO: Geneva, Switzerland, 2019. Available online: https://www.who.int/antimicrobial-resistance/interagency-coordination-group/final-report/en/ (accessed on 27 June 2019).
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Calvert, M.B.; Jumde, V.R.; Titz, A. Pathoblockers or antivirulence drugs as a new option for the treatment of bacterial infections. Beilstein J. Org. Chem. 2018, 14, 2607–2617. [Google Scholar] [CrossRef] [PubMed]
- Picard, B.; Garcia, J.S.; Gouriou, S.; Duriez, P.; Brahimi, N.; Bingen, E.; Elion, J.; Denamur, E. The link between phylogeny and virulence in Escherichia coli extraintestinal infection. Infect. Immun. 1999, 67, 546–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Clermont, O.; Menard, M.; Kuskowski, M.A.; Picard, B.; Denamur, E. Experimental Mouse Lethality of Escherichia coli Isolates, in Relation to Accessory Traits, Phylogenetic Group, and Ecological Source. J. Infect. Dis. 2006, 194, 1141–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massot, M.; Daubié, A.-S.; Clermont, O.; Jauréguy, F.; Couffignal, C.; Dahbi, G.; Mora, A.; Blanco, J.; Branger, C.; Mentré, F.; et al. Phylogenetic, virulence and antibiotic resistance characteristics of commensal strain populations of Escherichia coli from community subjects in the Paris area in 2010 and evolution over 30 years. Microbiology 2016, 162, 642–650. [Google Scholar] [CrossRef]
- Sabaté, M.; Moreno, E.; Pérez, T.; Andreu, A.; Prats, G. Pathogenicity island markers in commensal and uropathogenic Escherichia coli isolates. Clin. Microbiol. Infect. 2006, 12, 880–886. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, P.S.; Lipinski, L.; Dziembowski, A. PlasFlow: Predicting plasmid sequences in metagenomic data using genome signatures. Nucleic Acids Res. 2018, 46, e35. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Ponten, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Joensen, K.G.; Tetzschner, A.M.M.; Iguchi, A.; Aarestrup, F.M.; Scheutz, F. Rapid and easy in silico serotyping of Escherichia coli isolates by use of whole-genome sequencing data. J. Clin. Microbiol. 2015, 53, 2410–2426. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico Detection and Typing of Plasmids using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Liang, Y.; Lynch, K.H.; Dennis, J.J.; Wishart, D.S. PHAST: A Fast Phage Search Tool. Nucleic Acids Res. 2011, 39, W347–W352. [Google Scholar] [CrossRef]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef]
- Altschul, S.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Bateman, A. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Mareuil, F.; Doppelt-Azeroual, O.; Ménager, H.; Mareuil, F.; Doppelt-Azeroual, O.; Ménager, H. A public Galaxy platform at Pasteur used as an execution engine for web services. F1000Research 2017, 6. [Google Scholar] [CrossRef]
- Abby, S.S.; Néron, B.; Ménager, H.; Touchon, M.; Rocha, E.P.C. MacSyFinder: A Program to Mine Genomes for Molecular Systems with an Application to CRISPR-Cas Systems. PLoS ONE 2014, 9, e110726. [Google Scholar] [CrossRef]
- Abby, S.S.; Cury, J.; Guglielmini, J.; Néron, B.; Touchon, M.; Rocha, E.P.C. Identification of protein secretion systems in bacterial genomes. Sci. Rep. 2016, 6, 23080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orskov, F.; Orskov, I. Escherichia coli O:H serotypes isolated from human blood. Prevalence of the K1 antigen with technical details of O and H antigenic determination. Acta Pathol. Microbiol. Scand. Suppl. 1975, 83, 595–600. [Google Scholar] [PubMed]
- EUCAST The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters Version 9.0. 2019. Available online: http://www.eucast.org (accessed on 20 June 2019).
- Birnboim, H.C.; Doly, J. A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res. 1979, 7, 1513–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, R.A.; Anantham, S.; Hall, R.M. An improved plasmid size standard, 39R861+. Plasmid 2019, 102, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Meyers, J.A.; Sanchez, D.; Elwell, L.P.; Falkow, S. Simple Agarose Gel Electrophoretic Method for the Identification and Characterization of Plasmid Deoxyribonucleic Acid. J. Bacteriol. 1976, 127, 1529–1537. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, Y.; Peirano, G.; Pitout, J.D.D. Complete genome sequence of Escherichia coli J53, an azide-resistant laboratory strain used for conjugation experiments. Genome Announc. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Allué-Guardia, A.; Nyong, E.C.; Koenig, S.S.K.; Vargas, S.M.; Bono, J.L.; Eppinger, M. Closed Genome Sequence of Escherichia coli K-12 Group Strain C600. Microbiol. Resour. Announc. 2019, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, M.P.; Yamamoto, D.; de Mello Santos, A.C.; Ooka, T.; Hernandes, R.T.; Vieira, M.A.M.; Santos, F.F.; Silva, R.M.; Hayashi, T.; Gomes, T.A.T. Phenotypic characterization and virulence-related properties of Escherichia albertii strains isolated from children with diarrhea in Brazil. Pathog. Dis. 2019, 77. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Houdouin, V.; Bonacorsi, S.; Brahimi, N.; Clermont, O.; Nassif, X.; Bingen, E. A uropathogenicity island contributes to the pathogenicity of Escherichia coli strains that cause neonatal meningitis. Infect. Immun. 2002, 70, 5865–5869. [Google Scholar] [CrossRef] [Green Version]
- Garcia, B.G.; Ooka, T.; Gotoh, Y.; Vieira, M.A.M.; Yamamoto, D.; Ogura, Y.; Girão, D.M.; Sampaio, S.C.F.; Melo, A.B.; Irino, K.; et al. Genetic relatedness and virulence properties of enteropathogenic Escherichia coli strains of serotype O119:H6 expressing localized adherence or localized and aggregative adherence-like patterns on HeLa cells. Int. J. Med. Microbiol. 2016, 306, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.F.; Yamamoto, D.; Abe, C.M.; Bryant, J.A.; Hernandes, R.T.; Kitamura, F.C.; Castro, F.S.; Valiatti, T.B.; Piazza, R.M.F.; Elias, W.P.; et al. The Type III Secretion System (T3SS)-Translocon of Atypical Enteropathogenic Escherichia coli (aEPEC) Can Mediate Adherence. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.J.; Mulvey, M.A.; Schilling, J.D.; Pinkner, J.S.; Hultgren, S.J. Type 1 pilus-mediated bacterial invasion of bladder epithelial cells. EMBO J. 2000, 19, 2803–2812. [Google Scholar] [CrossRef] [PubMed]
- Hernandes, R.T.; Silva, R.M.; Carneiro, S.M.; Salvador, F.A.; Fernandes, M.C.D.C.; Padovan, A.C.B.; Yamamoto, D.; Mortara, R.A.; Elias, W.P.; da Silva Briones, M.R.; et al. The localized adherence pattern of an atypical enteropathogenic Escherichia coli is mediated by intimin omicron and unexpectedly promotes HeLa cell invasion. Cell. Microbiol. 2008, 0. [Google Scholar] [CrossRef] [PubMed]
- Alghoribi, M.F.; Gibreel, T.M.; Dodgson, A.R.; Beatson, S.A.; Upton, M. Galleria mellonella Infection Model Demonstrates High Lethality of ST69 and ST127 Uropathogenic E. coli. PLoS ONE 2014, 9, e101547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrey, D.O.; Pereira Dantas, P.; Martins, W.B.S.; Marques De Carvalho, F.; Gonzaga, L.; Almeida, P.; Sands, K.; Portal, E.; Sauser, J.; Cayô, R.; et al. An Emerging Clone, Klebsiella pneumoniae Carbapenemase 2–Producing K. pneumoniae Sequence Type 16, Associated With High Mortality Rates in a CC258-Endemic Setting. Clin. Infect. Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Beghain, J.; Bridier-Nahmias, A.; Nagard, H.L.; Denamur, E.; Clermont, O. ClermonTyping: An easy-to-use and accurate in silico method for Escherichia genus strain phylotyping. Microb. Genom. 2018, 4, e000192. [Google Scholar] [CrossRef]
- Pfeifer, Y.; Trifonova, A.; Pietsch, M.; Brunner, M.; Todorova, I.; Gergova, I.; Wilharm, G.; Werner, G.; Savov, E. Clonal Transmission of Gram-Negative Bacteria with Carbapenemases NDM-1, VIM-1, and OXA-23/72 in a Bulgarian Hospital. Microb. Drug Resist. 2017, 23, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Lagrutta, E.; Taylor, P.; Pham, J.; Nordmann, P. Emergence of metallo-β-lactamase NDM-1-producing multidrug-resistant Escherichia coli in Australia. Antimicrob. Agents Chemother. 2010, 54, 4914–4916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, J.S.; Kim, H.M.; Koo, H.S.; Yang, J.W.; Yoo, J.I.; Kim, H.S.; Park, H.K.; Lee, Y.S. Nosocomial transmission of NDM-1-producing Escherichia coli ST101 in a Korean hospital. J. Antimicrob. Chemother. 2013, 68, 2170–2172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaji, R.; Friedman, C.R.; Rubin, J.; Suh, J.; Thys, E.; McDermott, P.; Hung-Fan, M.; Riley, L.W. A Population-Based Surveillance Study of Shared Genotypes of Escherichia coli Isolates from Retail Meat and Suspected Cases of Urinary Tract Infections. mSphere 2018, 3, e00179-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, M.R.; McCulloch, J.A.; Vianello, M.A.; Moura, Q.; Pérez-Chaparro, P.J.; Esposito, F.; Sartori, L.; Dropa, M.; Matté, M.H.; Lira, D.P.A.; et al. First Report of the Globally Disseminated IncX4 Plasmid Carrying the mcr-1 Gene in a Colistin-Resistant Escherichia coli Sequence Type 101 Isolate from a Human Infection in Brazil. Antimicrob. Agents Chemother. 2016, 60, 6415–6417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerhold, G.; Schulze, M.H.; Gross, U.; Bohne, W. Multilocus sequence typing and CTX-M characterization of ESBL-producing E. coli: A prospective single-centre study in Lower Saxony, Germany. Epidemiol. Infect. 2016, 144, 3300–3304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, E.R.; Aung, M.S.; Paul, S.K.; Ahmed, S.; Haque, N.; Ahamed, F.; Sarkar, S.R.; Roy, S.; Rahman, M.M.; Mahmud, M.C.; et al. Prevalence and Molecular Epidemiology of Clinical Isolates of Escherichia coli and Klebsiella pneumoniae Harboring Extended-Spectrum Beta-Lactamase and Carbapenemase Genes in Bangladesh. Microb. Drug Resist. 2018, 24, 1568–1579. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, Y.; Witte, W.; Holfelder, M.; Busch, J.; Nordmann, P.; Poirel, L. NDM-1-producing Escherichia coli in Germany. Antimicrob. Agents Chemother. 2011, 55, 1318–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, D.A.; Freeman, J.T.; Roberts, S.A.; Heffernan, H.; Dyet, K.; Paterson, D.L.; Rogers, B.A.; Sidjabat, H.E.; Masters, J. Rectal colonization with New Delhi metallo-lactamase-1-producing Escherichia coli prior to transrectal ultrasound (TRUS)-guided prostate biopsy. J. Antimicrob. Chemother. 2013, 68, 2957–2959. [Google Scholar] [CrossRef] [Green Version]
- Potron, A.; Poirel, L.; Rondinaud, E.; Nordmann, P. Intercontinental spread of OXA-48 beta-lactamase-producing Enterobacteriaceae over a 11-year period, 2001 to 2011. Eurosurveillance 2013, 18, 20549. [Google Scholar] [CrossRef]
- Samuelsen, Ø.; Overballe-Petersen, S.; Bjørnholt, J.V.; Brisse, S.; Doumith, M.; Woodford, N.; Hopkins, K.L.; Aasnæs, B.; Haldorsen, B.; Sundsfjord, A.; et al. Molecular and epidemiological characterization of carbapenemase-producing Enterobacteriaceae in Norway, 2007 to 2014. PLoS ONE 2017, 12, e0187832. [Google Scholar] [CrossRef] [Green Version]
- Mantilla-Calderon, D.; Jumat, M.R.; Wang, T.; Ganesan, P.; Al-Jassim, N.; Hong, P.-Y. Isolation and Characterization of NDM-Positive Escherichia coli from Municipal Wastewater in Jeddah, Saudi Arabia. Antimicrob. Agents Chemother. 2016, 60, 5223–5231. [Google Scholar] [CrossRef] [Green Version]
- Braga, J.F.V.; Chanteloup, N.K.; Trotereau, A.; Baucheron, S.; Guabiraba, R.; Ecco, R.; Schouler, C. Diversity of Escherichia coli strains involved in vertebral osteomyelitis and arthritis in broilers in Brazil. BMC Vet. Res. 2016, 12, 140. [Google Scholar] [CrossRef]
- Koo, H.-J.; Kwak, H.-S.; Yoon, S.-H.; Woo, G.-J. Phylogenetic group distribution and prevalence of virulence genes in Escherichia coli isolates from food samples in South Korea. World J. Microbiol. Biotechnol. 2012, 28, 1813–1816. [Google Scholar] [CrossRef]
- Müller, A.; Jansen, W.; Grabowski, N.T.; Monecke, S.; Ehricht, R.; Kehrenberg, C. ESBL- and AmpC-producing Escherichia coli from legally and illegally imported meat: Characterization of isolates brought into the EU from third countries. Int. J. Food Microbiol. 2018, 283, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Lin, D.; Chen, K.; Chan, E.W.C.; Yao, W.; Chen, S. Molecular Characterization of Escherichia coli Strains Isolated from Retail Meat That Harbor blaCTX-M and fosA3 Genes. Antimicrob. Agents Chemother. 2016, 60, 2450–2455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Wang, Y.; Shi, X.; Wang, S.; Ren, H.; Shen, Z.; Wang, Y.; Lin, J.; Wang, S. Rapid rise of the ESBL and mcr-1 genes in Escherichia coli of chicken origin in China, 2008–2014. Emerg. Microbes Infect. 2018, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehmer, T.; Vogler, A.J.; Thomas, A.; Sauer, S.; Hergenroether, M.; Straubinger, R.K.; Birdsell, D.; Keim, P.; Sahl, J.W.; Williamson, C.H.D.; et al. Phenotypic characterization and whole genome analysis of extended-spectrum beta-lactamase-producing bacteria isolated from dogs in Germany. PLoS ONE 2018, 13, e0206252. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; García-Cobos, S.; Ruijs, G.J.H.M.; Kampinga, G.A.; Arends, J.P.; Borst, D.M.; Möller, L.V.; Holman, N.D.; Schuurs, T.A.; Bruijnesteijn van Coppenraet, L.E.; et al. Epidemiology of Extended-Spectrum β-Lactamase-Producing E. coli and Vancomycin-Resistant Enterococci in the Northern Dutch–German Cross-Border Region. Front. Microbiol. 2017, 8, 1914. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tian, G.-B.; Zhang, R.; Shen, Y.; Tyrrell, J.M.; Huang, X.; Zhou, H.; Lei, L.; Li, H.-Y.; Doi, Y.; et al. Prevalence, risk factors, outcomes, and molecular epidemiology of mcr-1-positive Enterobacteriaceae in patients and healthy adults from China: An epidemiological and clinical study. Lancet Infect. Dis. 2017, 17, 390–399. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, B.; Tada, T.; Miyoshi-Akiyama, T.; Shimada, K.; Ohara, H.; Kirikae, T.; Pokhrel, B.M. Identification of a novel NDM variant, NDM-13, from a multidrug-resistant Escherichia coli clinical isolate in Nepal. Antimicrob. Agents Chemother. 2015, 59, 5847–5850. [Google Scholar] [CrossRef] [Green Version]
- Herrero-Fresno, A.; Zachariasen, C.; Hansen, M.H.; Nielsen, A.; Hendriksen, R.S.; Nielsen, S.S.; Olsen, J.E. Apramycin treatment affects selection and spread of a multidrug-resistant Escherichia coli strain able to colonize the human gut in the intestinal microbiota of pigs. Vet. Res. 2016, 47, 12. [Google Scholar] [CrossRef] [Green Version]
- Trobos, M.; Lester, C.H.; Olsen, J.E.; Frimodt-Moller, N.; Hammerum, A.M. Natural transfer of sulphonamide and ampicillin resistance between Escherichia coli residing in the human intestine. J. Antimicrob. Chemother. 2009, 63, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Duriez, P.; Clermont, O.; Picard, B.; Denamur, E.; Bonacorsi, S.; Bingen, E.; Chaventré, A.; Elion, J. Commensal Escherichia coli isolates are phylogenetically distributed among geographically distinct human populations. Microbiology 2001, 147, 1671–1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallström, T.; Resman, F.; Ristovski, M.; Riesbeck, K. Binding of complement regulators to invasive nontypeable Haemophilus influenzae isolates is not increased compared to nasopharyngeal isolates, but serum resistance is linked to disease severity. J. Clin. Microbiol. 2010, 48, 921–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, C.; Phillips, R.; Roberts, A.P. Serum resistance among Escherichia coli strains causing urinary tract infection in relation to O type and the carriage of hemolysin, colicin, and antibiotic resistance determinants. Infect. Immun. 1982, 35, 270–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, N.R.L.; Stevens, M.P.; Dean, R.E.; Saint, R.J.; Pankhania, D.; Prior, J.L.; Atkins, T.P.; Kessler, B.; Nithichanon, A.; Lertmemongkolchai, G.; et al. Systematic Mutagenesis of Genes Encoding Predicted Autotransported Proteins of Burkholderia pseudomallei Identifies Factors Mediating Virulence in Mice, Net Intracellular Replication and a Novel Protein Conferring Serum Resistance. PLoS ONE 2015, 10, e0121271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desroches, M.; Clermont, O.; Lafeuillade, B.; Rodriguez, C.; Darty, M.; Royer, G.; Bouvet, O.; Ounnoughene, N.; Noizat-Pirenne, F.; Denamur, E.; et al. Genotypic and phenotypic characteristics of Escherichia coli involved in transfusion-transmitted bacterial infections: Implications for preventive strategies. Transfusion 2018, 58, 1940–1950. [Google Scholar] [CrossRef]
- Peirano, G.; Mulvey, G.L.; Armstrong, G.D.; Pitout, J.D.D. Virulence potential and adherence properties of Escherichia coli that produce CTX-M and NDM β-lactamases. J. Med. Microbiol. 2013, 62, 525–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, V.C.S.; Haake, D.A.; Churchill, B.M.; Justice, S.S.; Kim, J.H. Intracellular Bacterial Communities: A Potential Etiology for Chronic Lower Urinary Tract Symptoms. Urology 2015, 86, 425–431. [Google Scholar] [CrossRef] [Green Version]
- Rivardo, F.; Martinotti, M.G.; Turner, R.J.; Ceri, H. Synergistic effect of lipopeptide biosurfactant with antibiotics against Escherichia coli CFT073 biofilm. Int. J. Antimicrob. Agents 2011, 37, 324–331. [Google Scholar] [CrossRef]
- Wu, H.; Moser, C.; Wang, H.Z.; Høiby, N.; Song, Z.J. Strategies for combating bacterial biofilm infections. Int. J. Oral Sci. 2015, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kjelstrup, C.K.; Barber, A.E.; Norton, J.P.; Mulvey, M.A.; L’Abée-Lund, T.M. Escherichia coli O78 isolated from septicemic lambs shows high pathogenicity in a zebrafish model. Vet. Res. 2017, 48. [Google Scholar] [CrossRef] [Green Version]
- Hauser, E.; Mellmann, A.; Semmler, T.; Stoeber, H.; Wieler, L.H.; Karch, H.; Kuebler, N.; Fruth, A.; Harmsen, D.; Weniger, T.; et al. Phylogenetic and molecular analysis of food-borne shiga toxin-producing Escherichia coli. Appl. Environ. Microbiol. 2013, 79, 2731–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellmann, A.; Bielaszewska, M.; Köck, R.; Friedrich, A.W.; Fruth, A.; Middendorf, B.; Harmsen, D.; Schmidt, M.A.; Karch, H. Analysis of Collection of Hemolytic Uremic Syndrome–associated Enterohemorrhagic Escherichia coli. Emerg. Infect. Dis. 2008, 14, 1287–1290. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Mernelius, S.; Jernberg, C.; Einemo, I.M.; Monecke, S.; Ehricht, R.; Löfgren, S.; Matussek, A. Shiga toxin-producing Escherichia coli infection in Jönköping county, Sweden: Occurrence and molecular characteristics in correlation with clinical symptoms and duration of stx shedding. Front. Cell. Infect. Microbiol. 2018, 8, 125. [Google Scholar] [CrossRef] [PubMed]
- Ferdous, M.; Friedrich, A.W.; Grundmann, H.; de Boer, R.F.; Croughs, P.D.; Islam, M.A.; Kluytmans-van den Bergh, M.F.Q.; Kooistra-Smid, A.M.D.; Rossen, J.W.A. Molecular characterization and phylogeny of Shiga toxin–producing Escherichia coli isolates obtained from two Dutch regions using whole genome sequencing. Clin. Microbiol. Infect. 2016, 22, 642. [Google Scholar] [CrossRef] [Green Version]
- Jønsson, R.; Struve, C.; Jenssen, H.; Krogfelt, K.A. The wax moth Galleria mellonella as a novel model system to study Enteroaggregative Escherichia coli pathogenesis. Virulence 2017, 8, 1894–1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrieri, C.G.; Pereira, M.F.; Galdino, A.C.M.; dos Santos, A.L.S.; Elias, W.P.; Schuenck, R.P.; Spano, L.C. Typical and Atypical Enteroaggregative Escherichia coli Are Both Virulent in the Galleria mellonella Model. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Heitmueller, M.; Billion, A.; Dobrindt, U.; Vilcinskas, A.; Mukherjee, K. Epigenetic mechanisms regulate innate immunity against uropathogenic and commensal-like Escherichia coli in the surrogate insect model Galleria mellonella. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [Green Version]
- Ciesielczuk, H.; Betts, J.; Phee, L.; Doumith, M.; Hope, R.; Woodford, N.; Wareham, D.W. Comparative virulence of urinary and bloodstream isolates of extra-intestinal pathogenic Escherichia coli in a Galleria mellonella model. Virulence 2015, 6, 145–151. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Features | Occurrence |
|---|---|
| Hypothetical proteins | 702 |
| Proteins with functional assignments | 4473 |
| Proteins with E.C. b number assignments | 1300 |
| Proteins with G.O. b assignments | 1076 |
| Proteins with pathway assignments | 911 |
| Proteins with PLfam b assignments | 5063 |
| Proteins with PGfam b assignments | 5064 |
| Virulence Traits | Associated with Virulence In | |||
|---|---|---|---|---|
| Other Genera | DEC | ExPEC | Various | |
| Colonization and invasion | MlaA, MsbB, Sfm, MisL, IcsP | Lpf-1O26, EhaG, Hcp, Elf, EhaA, EhaB, CFA-I | FdeC, Pix, Ygi, Yad, Yeh, Yra, Yfc, YchO, IbeB, IbeC, EptC, OmpA | Type 1 Fimbriae (H191), Ecp, Curli |
| Immune system evasion | SodB, TrxA, SirA, FpkA | Iss, OmpTpa, OmpTca, Mig-14, HlyF | RelA | |
| Iron acquisition system | Sit, Iro, Fhu, Enterobactin | |||
| Regulators | DsbA, DegP, SlyA, CpxAR | EvgAS | DsbAB, PhoPQ | QseBC, RcsAB |
| Toxins and Bacteriocins | Microcin V, Colicin B, Colicin M | ClyA, Hly III | ||
| Non-LEE effectors | EspL1, EspL4, EspX1, EspX4, EspX5, EspR1 | |||
| Region | Length | Completeness a | Most Common Phage/Accession Number | GC % |
|---|---|---|---|---|
| 1 | 6.3 kb | Incomplete | Bacillus phage G/NC_023719 | 51.27 |
| 2 | 30.9 kb | Incomplete | Salmonella phage 118970_sal3/NC_031940 | 49.82 |
| 3 | 14.2 kb | Questionable | Enterobacteria phage P88/NC_026014 | 49.75 |
| 4 | 20 kb | Incomplete | Shigella phage POCJ13/NC_025434 | 47.95 |
| 5 | 28.5 kb | Incomplete | Shigella phage SfII/NC_021857 | 45.37 |
| 6 | 14.4 kb | Intact | Enterobacteria phage Fels-2/NC_010463 | 49.56 |
| 7 | 9.4 kb | Incomplete | Bacillus phage Shanette/NC_028983 | 49.50 |
| 8 | 7.5 kb | Incomplete | Enterobacteria phage phi92/NC_023693 | 44.44 |
| 9 | 10.2 kb | Questionable | Enterobacteria phage P2/NC_001895 | 55.39 |
| 10 | 3.8 kb | Incomplete | Bacteriophage WPhi/NC_005056 | 50.16 |
| 11 | 8.4 kb | Questionable | Phage cdtI/NC_009514 | 46.05 |
| 12 | 6.7 kb | Incomplete | Shewanella sp. phage¼/NC_025436 | 47.92 |
| Antimicrobial Class | Genotype | Phenotype | |
|---|---|---|---|
| Antimicrobial Agent | MIC b (μg/mL) | ||
| Aminoglycosides | aph(3″)-Ib, aph(6)-Id, aph(4)-Ia, aac(3)-Iva | amikacin | 2 |
| gentamicin | 32 | ||
| β-lactams | blaTEM-1B, blaCTX-M-2 | ampicillin | ≥256 |
| piperacillin/tazobactam | >256/4 | ||
| ceftazidime | 64 | ||
| ceftriaxone | ≥512 | ||
| cefepime | 16 | ||
| ertapenem | ≤0.5 | ||
| imipenem | ≤0.5 | ||
| meropenem | ≤0.5 | ||
| aztreonam | >32 | ||
| Sulfonamides/Trimethoprim | sul2, sul1, sul1′, dfrA14, dfrA7 | trimethoprim/sulfamethoxazole | >128/2432 |
| Phenicols | catA1 | chloramphenicol | >64 |
| Tetracyclines | tet(A) | minocycline | 16 |
| tigecycline | 1 | ||
| Fluoroquinolones | parE(S458A); parC (S80I); gyrA (S83L and D87Y) | ciprofloxacin | >64 |
| Polymyxins | - | colistin | ≤0.25 |
| polymyxin B | ≤0.25 | ||
| Fosfomycin | - | fosfomycin | 0.25 |
| Macrolides | mph(A), mdf(A) | azithromycin | NT |
| Traits | EC121 | J53 | C600 | ACG04 | ACC09 |
|---|---|---|---|---|---|
| Antimicrobial a | |||||
| ceftazidime | 64 | 0.25 | 0.25 | 8 | 2 |
| ceftriaxone | ≥64 | ≤0.125 | ≤0.125 | >64 | >64 |
| cefepime | 16 | 0.25 | ≤0.125 | 16 | 2 |
| piperacillin/tazobactam | >256 | 2 | 2 | 8 | 2 |
| aztreonam | >32 | ≤0.125 | ≤0.125 | 16 | 4 |
| amikacin | 4 | 2 | 2 | 16 | 2 |
| gentamycin | 16 | 0.5 | 0.5 | 32 | 0.5 |
| trimethoprim/sulfamethoxazole | >128 | ≤0.25 | ≤0.25 | >128 | >128 |
| ciprofloxacin | >64 | ≤0.125 | ≤0.125 | ≤0.125 | ≤0.125 |
| minocycline | 16 | 1 | 1 | 4 | 1 |
| tigecycline | 1 | 0.25 | 0.125 | 0.25 | 0.125 |
| Replicon typeb | |||||
| IncHI2 | + | - | - | + | + |
| IncL/M | + | - | - | + | - |
| IncFIB | + | - | - | + | - |
| IncN | + | - | - | + | - |
| Virulencec | |||||
| hlyF | + | - | - | + | - |
| iroN | + | - | - | + | - |
| sitA | + | - | - | + | - |
| Resistance gene | |||||
| blaCTX-M | + | - | - | + | + |
| E. coli Strains | ||||||
|---|---|---|---|---|---|---|
| J96 | EC121 | C600 | ||||
| Challenge Period | NHS | inHS | NHS | inHS | NHS | inHS |
| 30 min | 101 | 101 | 102 | 101 | 108 | 101 |
| 1 h | 102 | 101 | 102 | 101 | NG | 101 |
| 2 h | 102 | 101 | 102 | 101 | NG | 101 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, A.C.M.; Silva, R.M.; Valiatti, T.B.; Santos, F.F.; Santos-Neto, J.F.; Cayô, R.; Streling, A.P.; Nodari, C.S.; Gales, A.C.; Nishiyama-Jr, M.Y.; et al. Virulence Potential of a Multidrug-Resistant Escherichia coli Strain Belonging to the Emerging Clonal Group ST101-B1 Isolated from Bloodstream Infection. Microorganisms 2020, 8, 827. https://doi.org/10.3390/microorganisms8060827
Santos ACM, Silva RM, Valiatti TB, Santos FF, Santos-Neto JF, Cayô R, Streling AP, Nodari CS, Gales AC, Nishiyama-Jr MY, et al. Virulence Potential of a Multidrug-Resistant Escherichia coli Strain Belonging to the Emerging Clonal Group ST101-B1 Isolated from Bloodstream Infection. Microorganisms. 2020; 8(6):827. https://doi.org/10.3390/microorganisms8060827
Chicago/Turabian StyleSantos, Ana Carolina M., Rosa M. Silva, Tiago B. Valiatti, Fernanda F. Santos, José F. Santos-Neto, Rodrigo Cayô, Ana P. Streling, Carolina S. Nodari, Ana C. Gales, Milton Y. Nishiyama-Jr, and et al. 2020. "Virulence Potential of a Multidrug-Resistant Escherichia coli Strain Belonging to the Emerging Clonal Group ST101-B1 Isolated from Bloodstream Infection" Microorganisms 8, no. 6: 827. https://doi.org/10.3390/microorganisms8060827