Heme-Oxygenase and Kidney Transplantation: A Potential for Target Therapy?

 , ,
, ,  , , ,
, , , 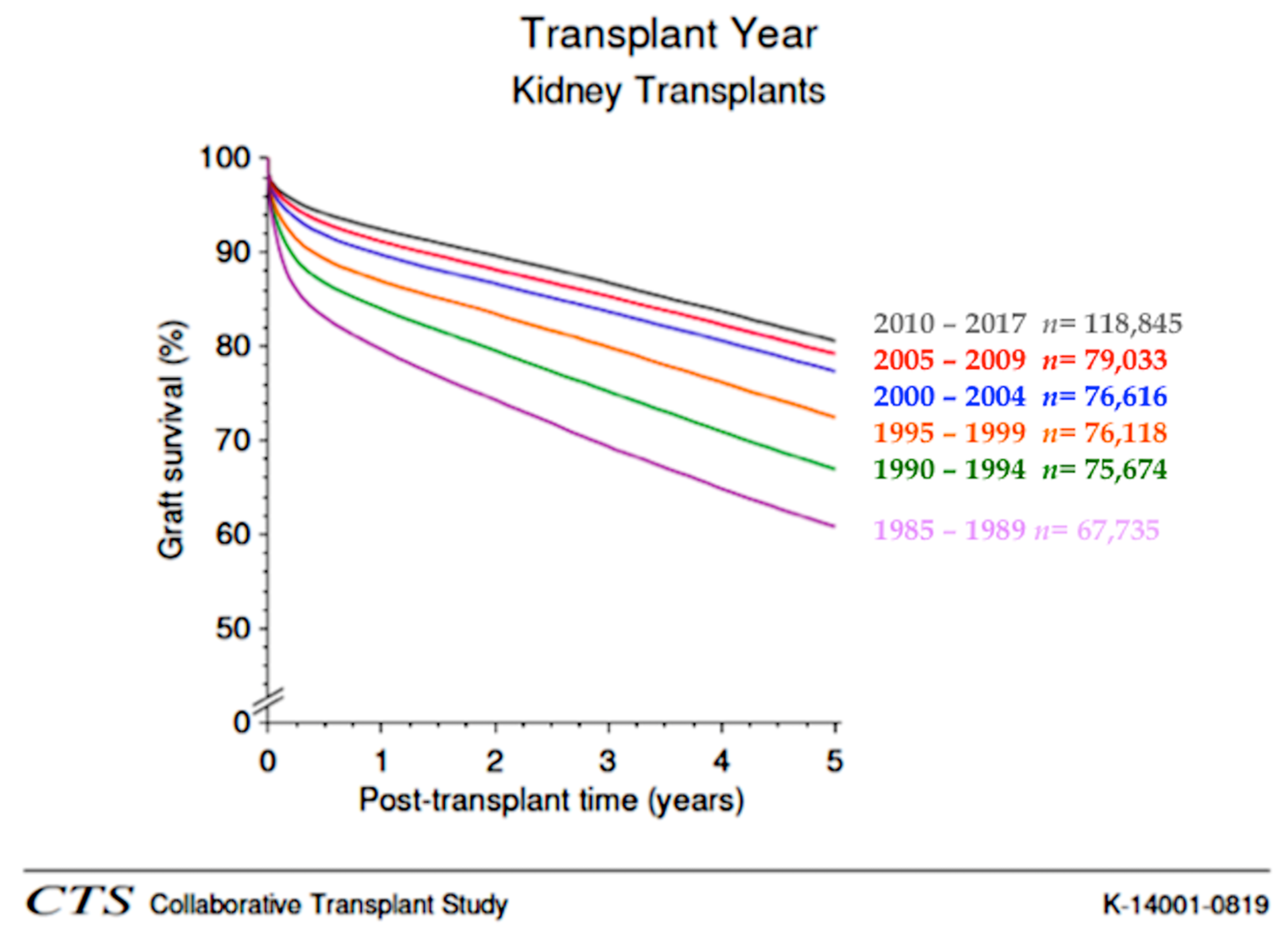{kind=link}
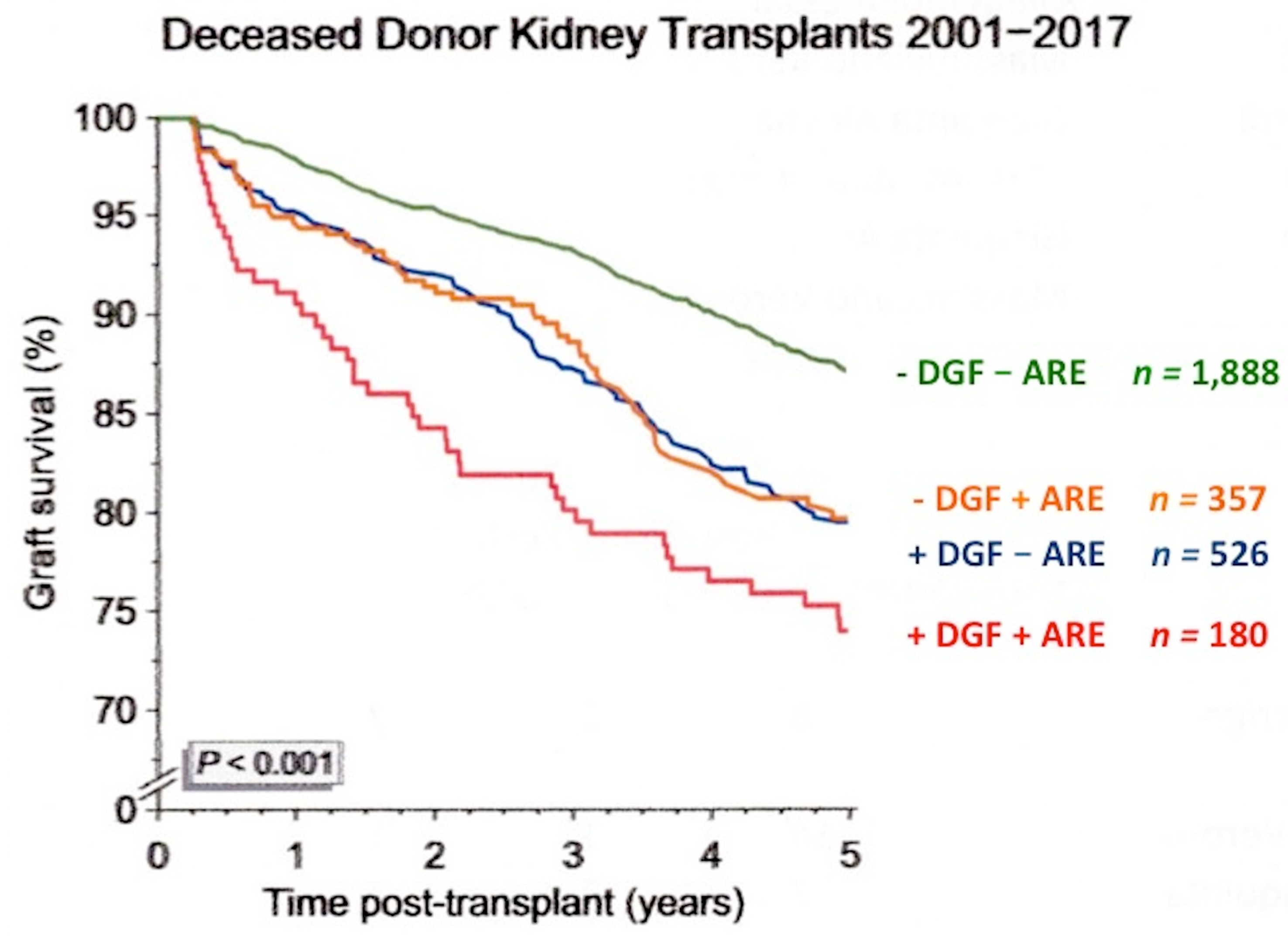{kind=link}
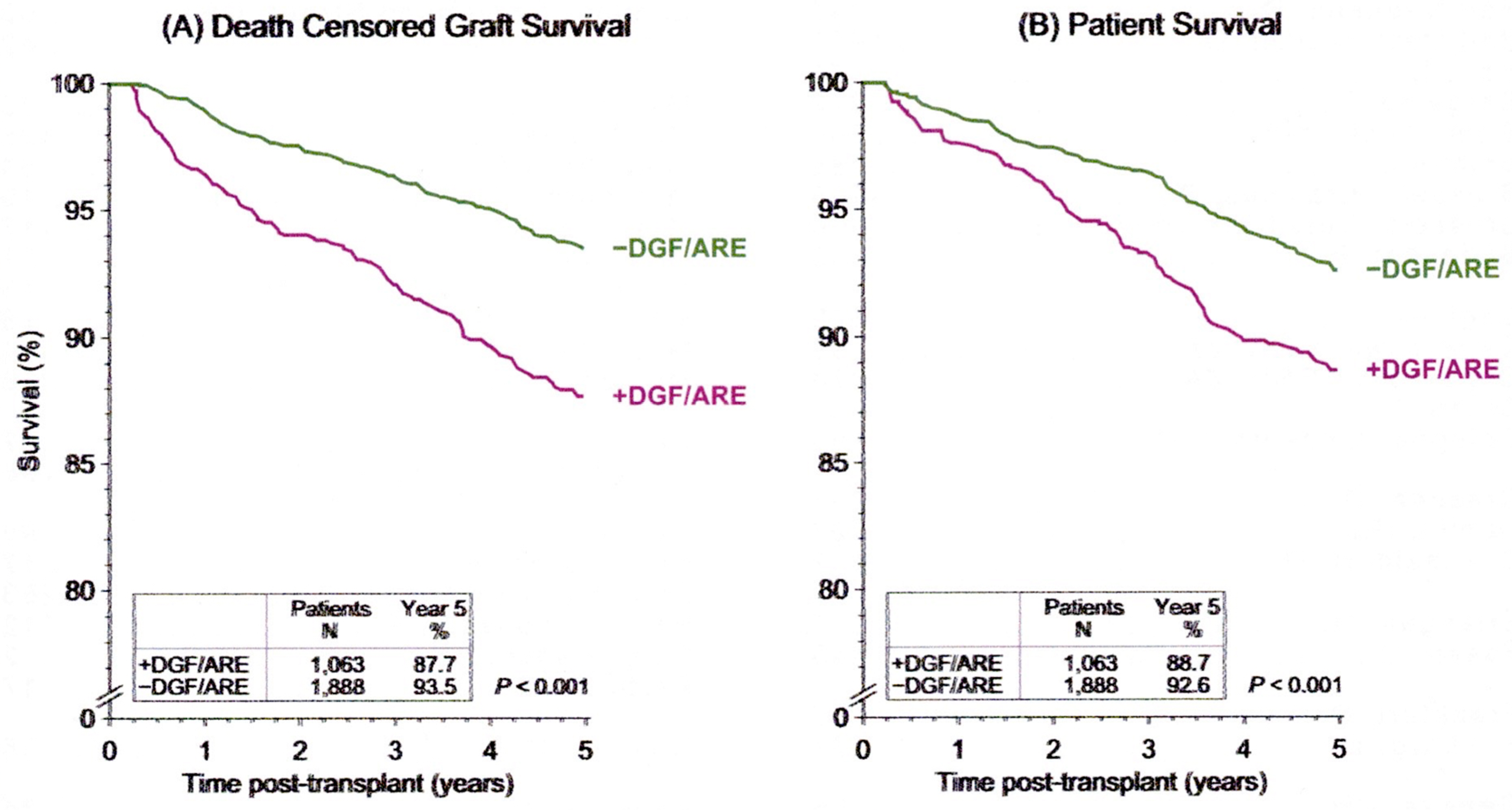{kind=link}
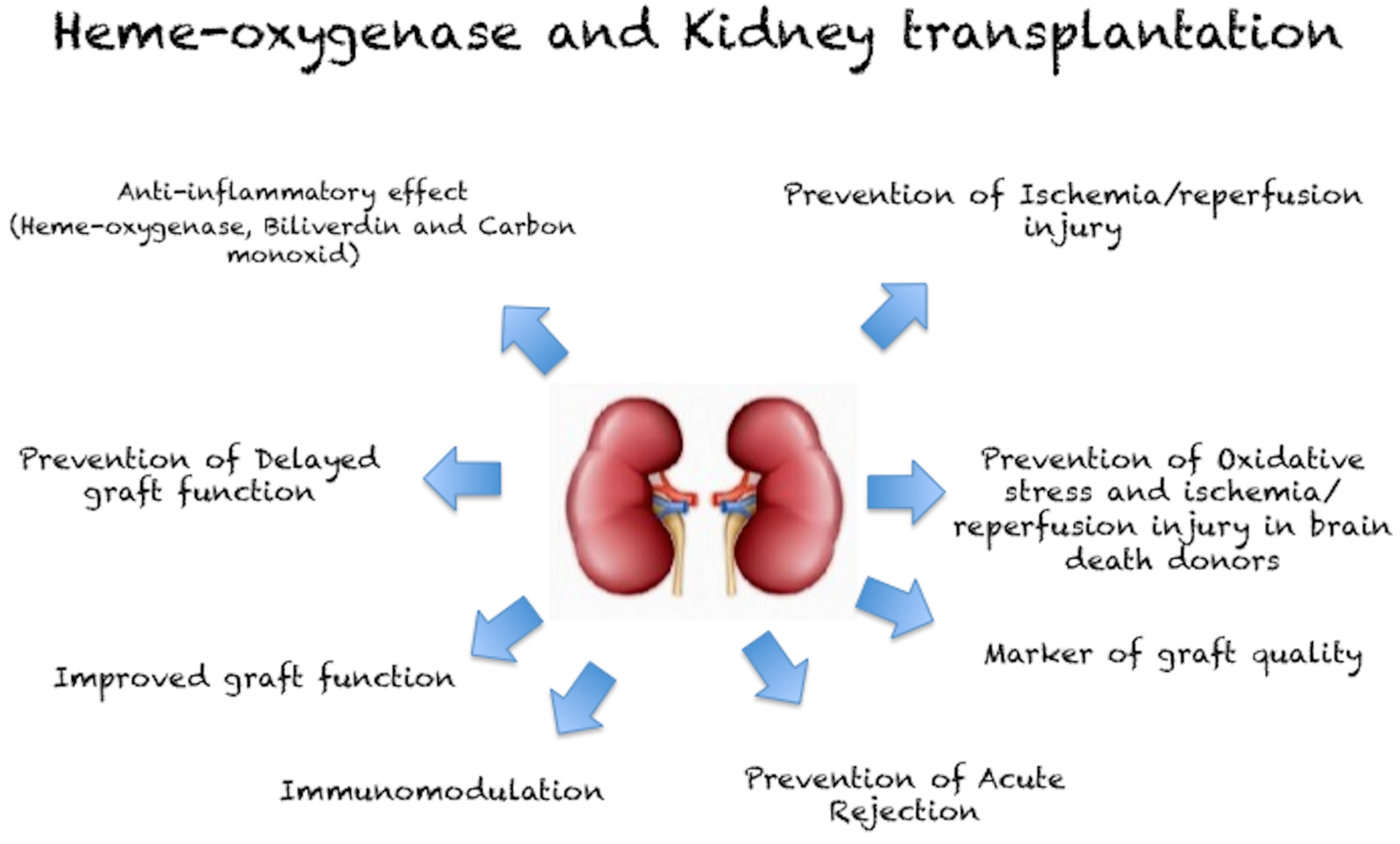{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Literature Research and Study Selection
Study Selection
3. Ischemia-Reperfusion Injury (IRI)
3.1. Ischemic Injury and Hypoxic Adaption
3.2. Type of Donor
4. Acute and Chronic Rejection and Immunosuppression
5. Potential Therapeutic Targets and Final Considerations
Author Contributions
Funding
Conflicts of Interest
References
- Veroux, M.; Corona, D.; Veroux, P. Kidney transplantation: Future challenges. Minerva Chir. 2009, 64, 75–100. [Google Scholar] [PubMed]
- Hariharan, S.; McBride, M.A.; Cherikh, W.S.; Tolleris, C.B.; Bresnahan, B.A.; Johnson, C.P. Post-transplant renal function in the first year predicts long-term kidney transplant survival. Kidney Int. 2002, 62, 311–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, W.H.; Johnson, D.W.; Teixeira-Pinto, A.; Wong, G. Association Between Duration of Delayed Graft Function, Acute Rejection, and Allograft Outcome After Deceased Donor Kidney Transplantation. Transplantation 2019, 103, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Correa-Costa, M.; Amano, M.T.; Câmara, N.O.S. Cytoprotection behind heme oxygenase-1 in renal diseases. World J. Nephrol. 2012, 1, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Cavaleri, M.; Veroux, M.; Palermo, F.; Vasile, F.; Mineri, M.; Palumbo, J.; Salemi, L.; Astuto, M.; Murabito, P. Perioperative Goal-Directed Therapy during Kidney Transplantation: An Impact Evaluation on the Major Postoperative Complications. J. Clin. Med. 2019, 8, 80. [Google Scholar] [CrossRef] [Green Version]
- Grosso, G.; Corona, D.; Mistretta, A.; Zerbo, D.; Sinagra, N.; Giaquinta, A.; Cimino, S.; Ekser, B.; Giuffrida, G.; Leonardi, A.; et al. Delayed Graft Function and Long-Term Outcome in Kidney Transplantation. Transplant. Proc. 2012, 44, 1879–1883. [Google Scholar] [CrossRef]
- Sorbello, M.; Morello, G.; Parrinello, L.; Molino, C.; Rinzivillo, D.; Pappalardo, R.; Cutuli, M.; Corona, D.; Veroux, P.; Veroux, M. Effect of N-Acetyl-Cysteine (NAC) Added to Fenoldopam or Dopamine on End-Tidal Carbon Dioxide and Mean Arterial Pressure at Time of Renal Artery Declamping During Cadaveric Kidney Transplantation. Transplant. Proc. 2010, 42, 1056–1060. [Google Scholar] [CrossRef]
- Perico, N.; Cattaneo, D.; Sayegh, M.H.; Remuzzi, G. Delayed graft function in kidney transplantation. Lancet 2004, 364, 1814–1827. [Google Scholar] [CrossRef]
- Ojo, A.O.; Wolfe, R.A.; Held, P.J.; Port, F.K.; Schmouder, R.L. Delayed graft function: Risk factors and implications for renal allograft survival. Transplantation 1997, 63, 968–974. [Google Scholar] [CrossRef]
- Yarlagadda, S.G.; Coca, S.; Formica, R.N.; Poggio, E.D.; Parikh, C.R. Association between delayed graft function and allograft and patient survival: A systematic review and meta-analysis. Nephrol. Dial. Transplant. 2008, 24, 1039–1047. [Google Scholar] [CrossRef] [Green Version]
- Sikorski, E.M.; Hock, T.; Hill-Kapturczak, N.; Agarwal, A. The story so far: Molecular regulation of the heme oxygenase-1 gene in renal injury. Am. J. Physiol. Physiol. 2004, 286, F425–F441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nath, K.A.; Garovic, V.D.; Grande, J.P.; Croatt, A.J.; Ackerman, A.W.; Farrugia, G.; Katusic, Z.S.; Belcher, J.D.; Vercellotti, G.M. Heme oxygenase-2 protects against ischemic acute kidney injury: Influence of age and sex. Am. J. Physiol. Physiol. 2019, 317, F695–F704. [Google Scholar] [CrossRef] [PubMed]
- Nath, K.A. Heme oxygenase-1 and acute kidney injury. Curr. Opin. Nephrol. Hypertens. 2014, 23, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nath, K. Heme oxygenase-1: A provenance for cytoprotective pathways in the kidney and other tissues. Kidney Int. 2006, 70, 432–443. [Google Scholar] [CrossRef] [Green Version]
- Bolisetty, S.; Zarjou, A.; Agarwal, A. Heme Oxygenase 1 as a Therapeutic Target in Acute Kidney Injury. Am. J. Kidney Dis. 2017, 69, 531–545. [Google Scholar] [CrossRef] [Green Version]
- Jarmi, T.; Agarwal, A. Heme oxygenase and renal disease. Curr. Hypertens. Rep. 2009, 11, 56–62. [Google Scholar] [CrossRef]
- Stocker, R. Antioxidant activities of bile pigments. Antioxid. Redox Signal 2004, 6, 841–849. [Google Scholar]
- Siedlecki, A.; Irish, W.; Brennan, D.C. Delayed graft function in the kidney transplant. Arab. Archaeol. Epigr. 2011, 11, 2279–2796. [Google Scholar] [CrossRef] [Green Version]
- Hanto, U.W. Ischemia-reperfusion injury in kidney transplantation. Front. Biosci. 2015, 7, 134–154. [Google Scholar] [CrossRef]
- Merion, R.M.; Ashby, V.B.; Wolfe, R.A.; Distant, D.A.; Hulbert-Shearon, T.E.; Metzger, R.A.; Ojo, A.; Port, F.K. Deceased-Donor Characteristics and the Survival Benefit of Kidney Transplantation. JAMA 2005, 294, 2726–2733. [Google Scholar] [CrossRef] [Green Version]
- Loutzenhiser, R.; Bidani, A.; Chilton, L. Renal myogenic response: Kinetic attributes and physiological role. Circ. Res. 2002, 90, 1316–1324. [Google Scholar] [CrossRef] [Green Version]
- Salahudeen, A.K. Cold ischemic injury of transplanted kidneys: New insights from experimental studies. Am. J. Physiol. Physiol. 2004, 287, F181–F187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanlaya, R.; Thongboonkerd, V. Molecular Mechanisms of Epigallocatechin-3-Gallate for Prevention of Chronic Kidney Disease and Renal Fibrosis: Preclinical Evidence. Curr. Dev. Nutr. 2019, 3, nzz101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganz, T. Systemic Iron Homeostasis. Physiol. Rev. 2013, 93, 1721–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trujillo, J.; Sanchez-Lozada, L.G.; Osorio-Alonso, H.; Tapia, E.; Scholze, A. New Pathogenic Concepts and Therapeutic Approaches to Oxidative Stress in Chronic Kidney Disease. Oxidative Med. Cell. Longev. 2016, 2016, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Na, H.-K.; Surh, Y.-J. Modulation of Nrf2-mediated antioxidant and detoxifying enzyme induction by the green tea polyphenol EGCG. Food Chem. Toxicol. 2008, 46, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Conde, E.; Alegre, L.; Blanco-Sanchez, I.; Sáenz-Morales, D.; Aguado-Fraile, E.; Ponte, B.; Ramos, E.; Saiz, A.; Jimenez, C.; Ordoñez, A.; et al. Hypoxia Inducible Factor 1-Alpha (HIF-1 Alpha) Is Induced during Reperfusion after Renal Ischemia and Is Critical for Proximal Tubule Cell Survival. PLoS ONE 2012, 7, e33258. [Google Scholar] [CrossRef]
- Delpech, P.O.; Thuillier, R.; Le Pape, S.; Rossard, L.; Jayle, C.; Billault, C.; Goujon, J.M.; Hauet, T. Effects of warm ischaemia combined with cold preservation on the hypoxia-inducible factor 1α pathway in an experimental renal autotransplantation model. BJS 2014, 101, 1739–1750. [Google Scholar] [CrossRef]
- Damman, J.; Bloks, V.W.; Daha, M.R.; Van Der Most, P.J.; Sanjabi, B.; Van Der Vlies, P.; Snieder, H.; Ploeg, R.J.; Krikke, C.; Leuvenink, H.G.; et al. Hypoxia and Complement-and-Coagulation Pathways in the Deceased Organ Donor as the Major Target for Intervention to Improve Renal Allograft Outcome. Transplantation 2015, 99, 1293–1300. [Google Scholar] [CrossRef]
- Rosenberger, C.; Eckardt, K.-U. Oxygenation of the Transplanted Kidney. Semin. Nephrol. 2019, 39, 554–566. [Google Scholar] [CrossRef]
- Lemos, F.B.; Ijzermans, J.N.; Zondervan, P.E.; Peeters, A.M.; Engel, S.V.D.; Mol, W.M.; Weimar, W.; Baan, C. Differential expression of heme oxygenase-1 and vascular endothelial growth factor in cadaveric and living donor kidneys after ischemia-reperfusion. J. Am. Soc. Nephrol. 2003, 14, 3278–3287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wszoła, M.; Kwiatkowski, A.; Domagala, P.; Wirkowska, A.; Bieniasz, M.; Diuwe, P.; Kieszek, R.; Durlik, M.; Chmura, A. Preservation of Kidneys by Machine Perfusion Influences Gene Expression and May Limit Ischemia/Reperfusion Injury. Prog. Transplant. 2014, 24, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Redaelli, C.A.; Wagner, M.; Kulli, C.; Tian, Y.H.; Schilling, M.K.; Wagner, A.C.C.; Mazzucchelli, L.; Kubulus, D. Hyperthermia-induced HSP expression correlates with improved rat renal isograft viability and survival in kidneys harvested from non-heart-beating donors. Transpl. Internat. 2001, 14, 351–360. [Google Scholar] [CrossRef]
- Wagner, M.; Cadetg, P.; Ruf, R.; Mazzucchelli, L.; Ferrari, P.; Redaelli, C.A. Heme oxygenase-1 attenuates ischemia/reperfusion-induced apoptosis and improves survival in rat renal allografts. Kidney Int. 2003, 63, 1564–1573. [Google Scholar] [CrossRef] [Green Version]
- Volti, G.L.; Sorrenti, V.; Murabito, P.; Galvano, F.; Veroux, M.; Gullo, A.; Acquaviva, R.; Stacchiotti, A.; Bonomini, F.; Vanella, L.; et al. Pharmacological Induction of Heme Oxygenase-1 Inhibits iNOS and Oxidative Stress in Renal Ischemia-Reperfusion Injury. Transplant. Proc. 2007, 39, 2986–2991. [Google Scholar] [CrossRef]
- Rund, K.M.; Peng, S.; Greite, R.; Claaßen, C.; Nolte, F.; Oger, C.; Galano, J.-M.; Balas, L.; Durand, T.; Chen, R.; et al. Dietary omega-3 PUFA improved tubular function after ischemia induced acute kidney injury in mice but did not attenuate impairment of renal function. Prostaglandins Other Lipid Mediat. 2019, 146, 106386. [Google Scholar] [CrossRef]
- Yi, X.; Zhang, G.; Yuan, J. Renoprotective Role of Fenoldopam Pretreatment Through Hypoxia-Inducible Factor-1alpha and Heme Oxygenase-1 Expressions in Rat Kidney Transplantation. Transplant. Proc. 2013, 45, 517–522. [Google Scholar] [CrossRef]
- Salahudeen, A.K.; Yang, M.; Huang, H.; Doré, S.; Stec, D.E. Fenoldopam Preconditioning: Role of Heme Oxygenase-1 in Protecting Human Tubular Cells and Rodent Kidneys Against Cold-Hypoxic Injury. Transplantation 2011, 91, 176–182. [Google Scholar] [CrossRef] [Green Version]
- Rosenberger, C.; Pratschke, J.; Rudolph, B.; Heyman, S.N.; Schindler, R.; Babel, N.; Eckardt, K.-U.; Frei, U.; Rosen, S.; Reinke, P. Immunohistochemical Detection of Hypoxia-Inducible Factor-1α in Human Renal Allograft Biopsies. J. Am. Soc. Nephrol. 2006, 18, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, K.S.; Kimura, S.; Murase, N. Use of carbon monoxide in minimizing ischemia/reperfusion injury in transplantation. Transplant. Rev. 2011, 26, 125–139. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, J.; Ozaki, K.S.; Nalesnik, M.A.; Ueki, S.; Castillo-Rama, M.; Faleo, G.; Ezzelarab, M.B.; Nakao, A.; Ekser, B.; Echeverri, G.; et al. Ex vivo application of carbon monoxide in UW solution prevents transplant-induced renal ischemia/reperfusion injury in pigs. Arab. Archaeol. Epigr. 2010, 10, 763–772. [Google Scholar] [CrossRef] [Green Version]
- Nijboer, W.N.; Schuurs, T.; Van Der Hoeven, J.; Leuvenink, H.G.; Van Der Heide, J.H.; Van Goor, H.; Ploeg, R. Effects of brain death on stress and inflammatory response in the human donor kidney. Transplant. Proc. 2005, 37, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Schuurs, T.A.; Morariu, A.; Ottens, P.J.; ’t Hart, N.A.; Popma, S.H.; Leuvenink, H.G.; Ploeg, R.J. Time-Dependent Changes in Donor Brain Death Related Processes. Arab. Archaeol. Epigr. 2006, 6, 2903–2911. [Google Scholar] [CrossRef] [PubMed]
- Nijboer, W.N.; Schuurs, T.A.; Van Der Hoeven, J.A.B.; Fekken, S.; Wiersema-Buist, J.; Leuvenink, H.G.; Hofker, S.; Van Der Heide, J.J.H.; Van Son, W.J.; Ploeg, R.J. Effect of Brain Death on Gene Expression and Tissue Activation in Human Donor Kidneys. Transplantation 2004, 78, 978–986. [Google Scholar] [CrossRef] [Green Version]
- Van Dullemen, L.F.; Bos, E.; Schuurs, T.A.; Kampinga, H.H.; Ploeg, R.J.; Van Goor, H.; Leuvenink, H.G. Brain death induces renal expression of heme oxygenase-1 and heat shock protein 70. J. Transl. Med. 2013, 11, 22. [Google Scholar] [CrossRef] [Green Version]
- Tullius, S.G.; Nieminen-Kelhä, M.; Bachmann, U.; Reutzel-Selke, A.; Jonas, S.; Pratschke, J.; Bechstein, W.O.; Reinke, P.; Buelow, R.; Neuhaus, P.; et al. Induction of heme-oxygenase-1 prevents ischemia/reperfusion injury and improves long-term graft outcome in rat renal allografts. Transplant. Proc. 2001, 33, 1286–1287. [Google Scholar] [CrossRef]
- Hölzen, J.P.; August, C.; Bahde, R.; Minin, E.; Lang, D.; Heidenreich, S.; Dietl, K.-H.; Spiegel, H.-U. Influence of Heme Oxygenase-1 on Microcirculation After Kidney Transplantation. J. Surg. Res. 2008, 148, 126–135. [Google Scholar] [CrossRef]
- Rossi, M.; Delbauve, S.; Wespes, E.; Roumeguère, T.; Leo, O.; Flamand, V.; Le Moine, A.; Hougardy, J.-M. Dual effect of hemin on renal ischemia-reperfusion injury. Biochem. Biophys. Res. Commun. 2018, 503, 2820–2825. [Google Scholar] [CrossRef]
- Nijboer, W.N.; Ottens, P.J.; Van Dijk, A.; Van Goor, H.; Ploeg, R.J.; Leuvenink, H.G. Donor pretreatment with carbamylated erythropoietin in a brain death model reduces inflammation more effectively than erythropoietin while preserving renal function. Crit. Care Med. 2010, 38, 1155–1161. [Google Scholar] [CrossRef]
- Kotsch, K.; Francuski, M.; Pascher, A.; Klemz, R.; Seifert, M.; Mittler, J.; Schumacher, G.; Buelow, R.; Volk, H.-D.; Tullius, S.; et al. Improved Long-Term Graft Survival after HO-1 Induction in Brain-Dead Donors. Arab. Archaeol. Epigr. 2006, 6, 477–486. [Google Scholar] [CrossRef]
- Schnuelle, P.; Yard, B.A.; Braun, C.; Dominguez-Fernandez, E.; Schaub, M.; Birck, R.; Sturm, J.; Post, S.; Van Der Woude, F.J. Impact of donor dopamine on immediate graft function after kidney transplantation. Arab. Archaeol. Epigr. 2004, 4, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Schnuelle, P.; Gottmann, U.; Hoeger, S.; Boesebeck, D.; Lauchart, W.; Weiss, C.; Fischereder, M.; Jauch, K.-W.; Heemann, U.; Zeier, M.; et al. Effects of donor pre-reatment with dopamine on graft function after kidney transplantation: A randomized controlled trial. JAMA 2009, 302, 1067–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florim, G.M.; Caldas, H.C.; Gonçalves, N.N.; Bueno, G.O.; Baptista, M.A.; Fernandes-Charpiot, I.M.; Abbud-Filho, M. Activation of HMGB1–TLR4 Pathway and Inflammasome Contribute to Enhanced Inflammatory Response in Extended Criteria and Kidneys with KDPI ≥85%. Transplantation 2020, 104, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Pena, A.; Reoma, J.L.; Krause, E.; Boothman, E.L.; Padiyar, N.P.; Cook, K.E.; Bartlett, R.H.; Punch, J.D.; Pena, R. Extracorporeal support: Improves donor renal graft function after cardiac death. Arab. Archaeol. Epigr. 2010, 10, 1365–1374. [Google Scholar] [CrossRef] [Green Version]
- Summers, D.M.; Johnson, R.J.; Allen, J.; Fuggle, S.V.; Collett, D.; Watson, C.J.; Bradley, J.A. Analysis of factors that affect outcome after transplantation of kidneys donated after cardiac death in the UK: A cohort study. Lancet 2010, 376, 1303–1311. [Google Scholar] [CrossRef]
- Wang, L.; Vijayan, V.; Jang, M.-S.; Thorenz, A.; Greite, R.; Rong, S.; Chen, R.; Shushakova, N.; Tudorache, I.; Derlin, K.; et al. Labile Heme Aggravates Renal Inflammation and Complement Activation After Ischemia Reperfusion Injury. Front. Immunol. 2019, 10, 2975. [Google Scholar] [CrossRef]
- Öllinger, R.; Kogler, P.; Biebl, M.; Sieb, M.; Sucher, R.; Bösmüller, C.; Troppmair, J.; Mark, W.; Weiss, H.; Margreiter, R. Protein levels of heme oxygenase-1 during reperfusion in human kidney transplants with delayed graft function. Clin. Transplant. 2008, 22, 418–423. [Google Scholar] [CrossRef]
- Wever, K.E.; Menting, T.P.; Rovers, M.; van der Vliet, A.J.; Rongen, G.A.; Masereeuw, R.; Ritskes-Hoitinga, M.; Hooijmans, C.R.; Warle, M. Ischemic preconditioning in the animal kidney, a systematic review and meta-analysis. PLoS ONE 2012, 7, e32296. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.-C.; Lin, L.-C.; Wu, M.-S.; Chien, C.-T.; Lai, M.-K. Repetitive Hypoxic Preconditioning Attenuates Renal Ischemia/Reperfusion Induced Oxidative Injury via Upregulating HIF-1α–Dependent bcl-2 Signaling. Transplantation 2009, 88, 1251–1260. [Google Scholar] [CrossRef]
- Soendergaard, P.; Krogstrup, N.V.; Secher, N.; Ravlo, K.; Keller, A.K.; Toennesen, E.; Bibby, B.M.; Moldrup, U.; Ostraat, E.O.; Pedersen, M.; et al. Improved GFR and renal plasma perfusion following remote ischaemic conditioning in a porcine kidney transplantation model. Transpl. Int. 2012, 25, 1002–1012. [Google Scholar] [CrossRef]
- Qiu, T.; Wang, Z.; Liu, X.; Chen, H.; Zhou, J.; Chen, Z.-Y.; Wang, M.; Jiang, G.; Wang, L.; Yu, G.; et al. Effect of ozone oxidative preconditioning on oxidative stress injury in a rat model of kidney transplantation. Exp. Ther. Med. 2017, 13, 1948–1955. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Kim, Y.; Matas, A.J.; Alam, J.; A Nath, K. Gas-generating systems in acute renal allograft rejection in the rat. Co-induction of heme oxygenase and nitric oxide synthase. Transplantation 1996, 61, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.; Iyer, S.; Cornejo, M.-C.; Mori, N.; Gao, L.; Sipos, I.; Maines, M.; Buelow, R. Stress protein-induced immunosuppression: Inhibition of cellular immune effector functions following overexpression of haem oxygenase (HSP 32). Transpl. Immunol. 1998, 6, 84–93. [Google Scholar] [CrossRef]
- Yu, X.; Song, B.; Huang, C.; Xiao, Y.; Fang, M.; Feng, J.; Wang, P.; Zhang, G. Prolonged survival time of allografts by the oral administration of RDP58 linked to the cholera toxin B subunit. Transpl. Immunol. 2012, 27, 122–127. [Google Scholar] [CrossRef]
- Martins, P.N.; Reuzel-Selke, A.; Jurisch, A.; Atrott, K.; Pascher, A.; Pratschke, J.; Buelow, R.; Neuhaus, P.; Volk, H.-D.; Tullius, S. Induction of carbon monoxide in the donor reduces graft immunogenicity and chronic graft deterioration. Transplant. Proc. 2005, 37, 379–381. [Google Scholar] [CrossRef]
- Kirkby, K.A.; Adin, C.A. Products of heme oxygenase and their potential therapeutic applications. Am. J. Physiol. Physiol. 2006, 290, F563–F571. [Google Scholar] [CrossRef] [Green Version]
- Nakao, A.; Neto, J.S.; Kanno, S.; Stolz, D.B.; Kimizuka, K.; Liu, F.; Bach, F.H.; Billiar, T.R.; Choi, A.M.K.; Otterbein, L.E.; et al. Protection Against Ischemia/Reperfusion Injury in Cardiac and Renal Transplantation with Carbon Monoxide, Biliverdin and Both. Arab. Archaeol. Epigr. 2005, 5, 282–291. [Google Scholar] [CrossRef]
- Öllinger, R.; Pratschke, J. Role of heme oxygenase-1 in transplantation. Transpl. Int. 2010, 23, 1071–1081. [Google Scholar] [CrossRef]
- Zhou, Z.; Ma, D.; Liu, P.; Wang, P.; Wei, D.; Yu, K.; Li, P.; Fang, Q.; Wang, J. Deletion of HO-1 blocks development of B lymphocytes in mice. Cell. Signal. 2019, 63, 109378. [Google Scholar] [CrossRef]
- Hancock, W.W.; Buelow, R.; Sayegh, M.H.; Turka, L.A. Antibody-induced transplant arteriosclerosis is prevented by graft expression of anti-oxidant and anti-apoptotic genes. Nat. Med. 1998, 4, 1392–1396. [Google Scholar] [CrossRef]
- Bédard, E.L.R.; Jiang, J.; Parry, N.; Wang, H.; Liu, W.; Garcia, B.; Kim, P.; Chakrabarti, S.; Buelow, R.; Zhong, R. Peritransplant treatment with cobalt protoporphyrin attenuates chronic renal allograft rejection. Transpl. Int. 2005, 18, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Baan, C.; Peeters, A.; Lemos, F.; Uitterlinden, A.; Doxiadis, I.; Claas, F.; Ijzermans, J.; Roodnat, J.; Weimar, W. Fundamental Role for HO-1 in the Self-Protection of Renal Allografts. Arab. Archaeol. Epigr. 2004, 4, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, K.; Marques, G.; Nogueira, E.; Feitoza, R.; Cenedeze, M.; Franco, M.; Mazzali, M.; Soares, M.; Pacheco-Silva, A.; Câmara, N.O.S. Improved renal function after kidney transplantation is associated with heme oxygenase-1 polymorphism. Clin. Transplant. 2008, 22, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Katana, E.; Skoura, L.; Giakoustidis, D.; Takoudas, D.; Malisiovas, N.; Daniilidis, M. Association Between the Heme Oxygenase-1 Promoter Polymorphism and Renal Transplantation Outcome in Greece. Transplant. Proc. 2010, 42, 2479–2485. [Google Scholar] [CrossRef]
- Courtney, A.E.; McNamee, P.T.; Middleton, D.; Heggarty, S.; Patterson, C.C.; Maxwell, A.P. Association of Functional Heme Oxygenase-1 Gene Promoter Polymorphism with Renal Transplantation Outcomes. Arab. Archaeol. Epigr. 2007, 7, 908–913. [Google Scholar] [CrossRef]
- Ekser, B.; Bianchi, J.; Ball, S.; Iwase, H.; Walters, A.; Ezzelarab, M.B.; Veroux, M.; Gridelli, B.; Wagner, R.; Ayares, D.; et al. Comparison of hematologic, biochemical, and coagulation parameters in α1,3-galactosyltransferase gene-knockout pigs, wild-type pigs, and four primate species. Xenotransplantation 2012, 19, 342–354. [Google Scholar] [CrossRef] [Green Version]
- Iwase, H.; Ekser, B.; Satyananda, V.; Bhama, J.; Hara, H.; Ezzelarab, M.B.; Klein, E.; Wagner, R.; Long, C.; Thacker, J.; et al. Pig-to-baboon heterotopic heart transplantation - exploratory preliminary experience with pigs transgenic for human thrombomodulin and comparison of three costimulation blockade-based regimens. Xenotransplantation 2015, 22, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Petersen, B.; Ramackers, W.; Lucas-Hahn, A.; Lemme, E.; Hasse, P.; Queisser, A.-L.; Herrmann, D.; Barg-Kues, B.; Carnwath, J.W.; Klose, J.; et al. Transgenic expression of human hemeoxygenase-1 in pigs confers resistance against xenograft rejection during ex vivo perfusion of porcine kidneys. Xenotransplantation 2011, 18, 355–368. [Google Scholar] [CrossRef]
- Yeom, H.-J.; Koo, O.J.; Yang, J.; Cho, B.; Hwang, J.-I.; Park, S.J.; Hurh, S.; Kim, H.; Lee, E.M.; Ro, H.; et al. Generation and Characterization of Human Heme Oxygenase-1 Transgenic Pigs. PLoS ONE 2012, 7, e46646. [Google Scholar] [CrossRef]
- Ahrens, H.E.; Petersen, B.; Ramackers, W.; Petkov, S.; Herrmann, R.; Hauschild-Quintern, J.; Lucas-Hahn, A.; Hassel, P.; Ziegler, M.; Baars, W.; et al. Kidneys From α1,3-Galactosyltransferase Knockout/Human Heme Oxygenase-1/Human A20 Transgenic Pigs Are Protected From Rejection During Ex Vivo Perfusion With Human Blood. Transplant. Direct 2015, 1, 1–8. [Google Scholar] [CrossRef]
- Liang, G.-B.; Luo, G.-H.; Bao, D.-S.; Chen, A.-J.; Zhuang, Y.-X.; Guo, Y.-N.; Wang, X.; Wang, Y.-L.; Chen, Z.-P.; Lu, Y.; et al. Impact of immunosuppressive agents on the expression of indoleamine 2,3-dioxygenase, heme oxygenase-1 and interleukin-7 in mesangial cells. Mol. Med. Rep. 2015, 12, 2577–2583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, G.; Cenedeze, M.; Feitoza, C.; Wang, P.; Bertocchi, A.; Damião, M.; Pinheiro, H.; Teixeira, V.A.; Dos Reis, M.; Pacheco-Silva, A.; et al. The role of heme oxygenase 1 in rapamycin-induced renal dysfunction after ischemia and reperfusion injury. Kidney Int. 2006, 70, 1742–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezzani, R.; Rodella, L.F.; Bianchi, R.; Goodman, A.; Lianos, E. Protective Effects of Heme-Oxygenase Expression in Cyclosporine A - Induced Injury. Curr. Neurovascular Res. 2005, 2, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhu, P.; Fujino, M.; Isaka, Y.; Ito, H.; Takahashi, K.; Nakajima, M.; Tanaka, T.; Zhuang, J.; Li, X.-K. 5-aminolaevulinic acid (ALA), enhances heme oxygenase (HO)-1 expression and attenuates tubulointerstitial fibrosis and renal apoptosis in chronic cyclosporine nephropathy. Biochem. Biophys. Res. Commun. 2019, 508, 583–589. [Google Scholar] [CrossRef]
- Liu, C.; Fujino, M.; Zhu, S.; Isaka, Y.; Ito, H.; Takahashi, K.; Nakajima, M.; Tanaka, T.; Zhu, P.; Li, X.-K. 5-ALA/SFC enhances HO-1 expression through the MAPK/Nrf2 antioxidant pathway and attenuates murine tubular epithelial cell apoptosis. FEBS Open Bio 2019, 9, 1928–1938. [Google Scholar] [CrossRef]
- Thomas, R.A.B.; Czopek, A.; Bellamy, C.O.C.; McNally, S.J.; Kluth, D.C.; Marson, L.P. Hemin Preconditioning Upregulates Heme Oxygenase-1 in Deceased Donor Renal Transplant Recipients. Transplantation 2016, 100, 176–183. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corona, D.; Ekser, B.; Gioco, R.; Caruso, M.; Schipa, C.; Veroux, P.; Giaquinta, A.; Granata, A.; Veroux, M. Heme-Oxygenase and Kidney Transplantation: A Potential for Target Therapy? Biomolecules 2020, 10, 840. https://doi.org/10.3390/biom10060840
Corona D, Ekser B, Gioco R, Caruso M, Schipa C, Veroux P, Giaquinta A, Granata A, Veroux M. Heme-Oxygenase and Kidney Transplantation: A Potential for Target Therapy? Biomolecules. 2020; 10(6):840. https://doi.org/10.3390/biom10060840
Chicago/Turabian StyleCorona, Daniela, Burcin Ekser, Rossella Gioco, Massimo Caruso, Chiara Schipa, Pierfrancesco Veroux, Alessia Giaquinta, Antonio Granata, and Massimiliano Veroux. 2020. "Heme-Oxygenase and Kidney Transplantation: A Potential for Target Therapy?" Biomolecules 10, no. 6: 840. https://doi.org/10.3390/biom10060840