Bitter Melon (Momordica charantia L.) Rootstock Improves the Heat Tolerance of Cucumber by Regulating Photosynthetic and Antioxidant Defense Pathways


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effects of Bitter Melon Rootstock on Plant Growth under Heat Stress
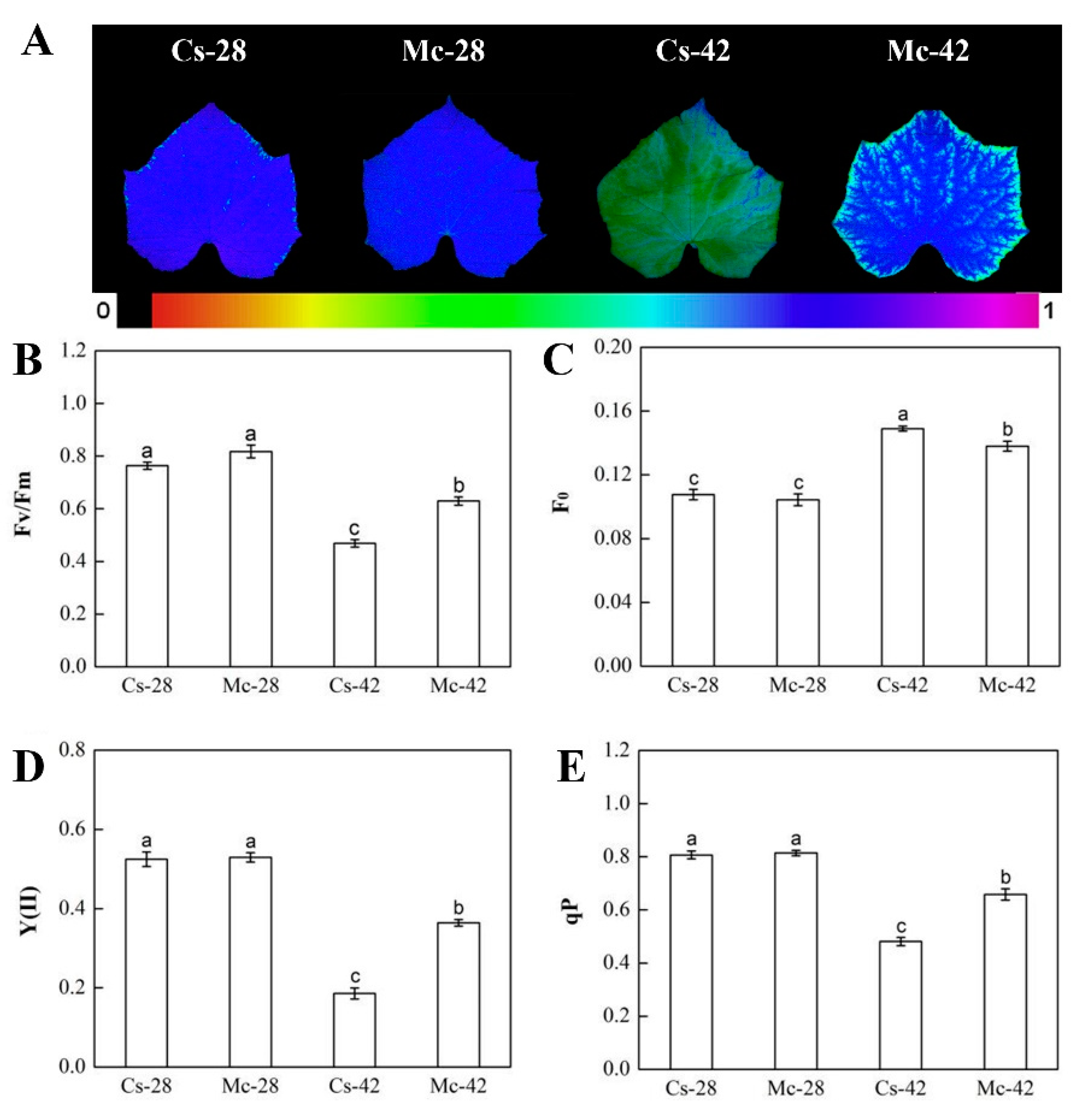
2.2. Effects of Bitter Melon Rootstock on Photosystem II (PSII) Photochemistry under Heat Stress Conditions
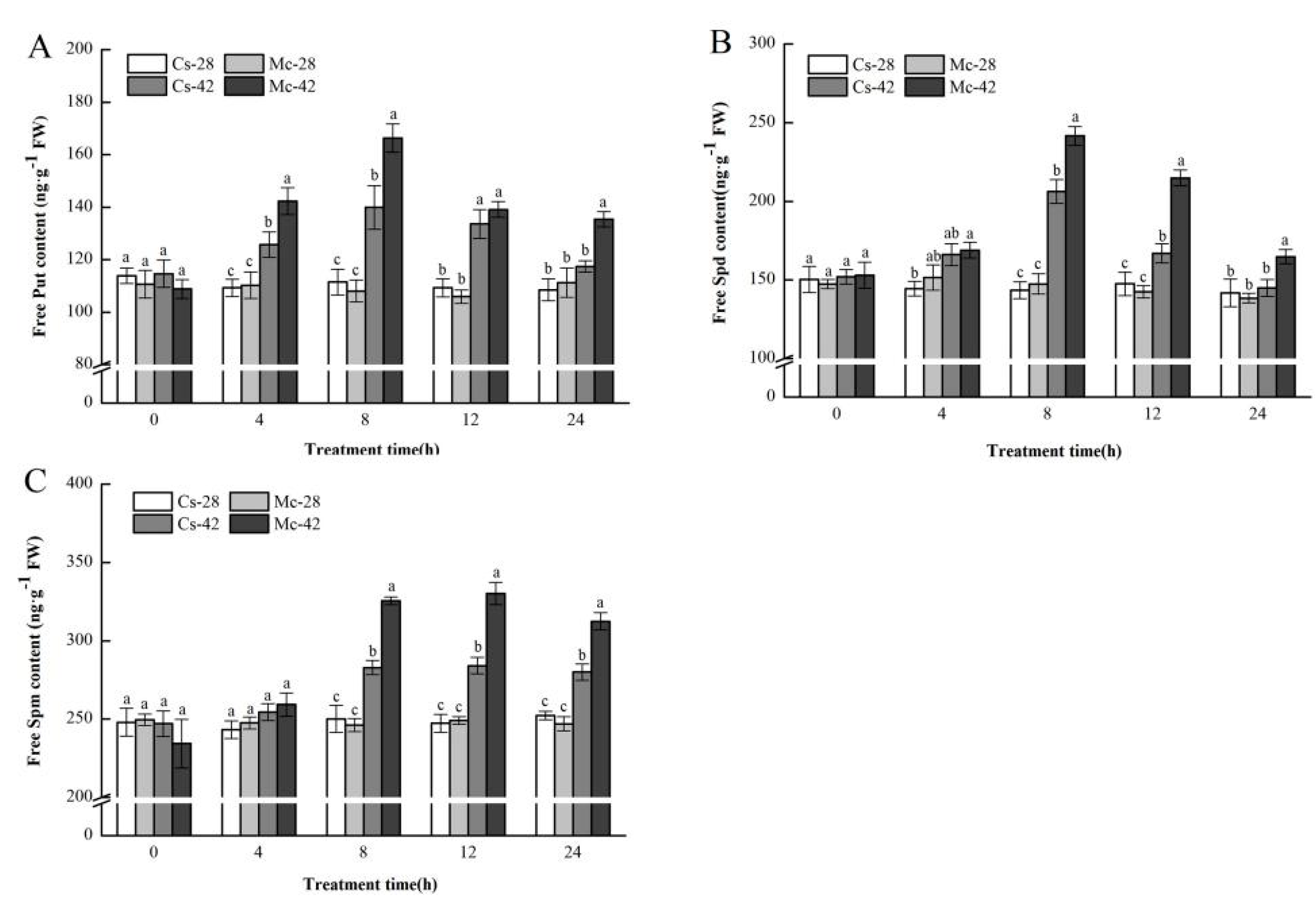
2.3. Effect of Bitter Melon Rootstock on the Endogenous Polyamine Content under Heat Stress Conditions
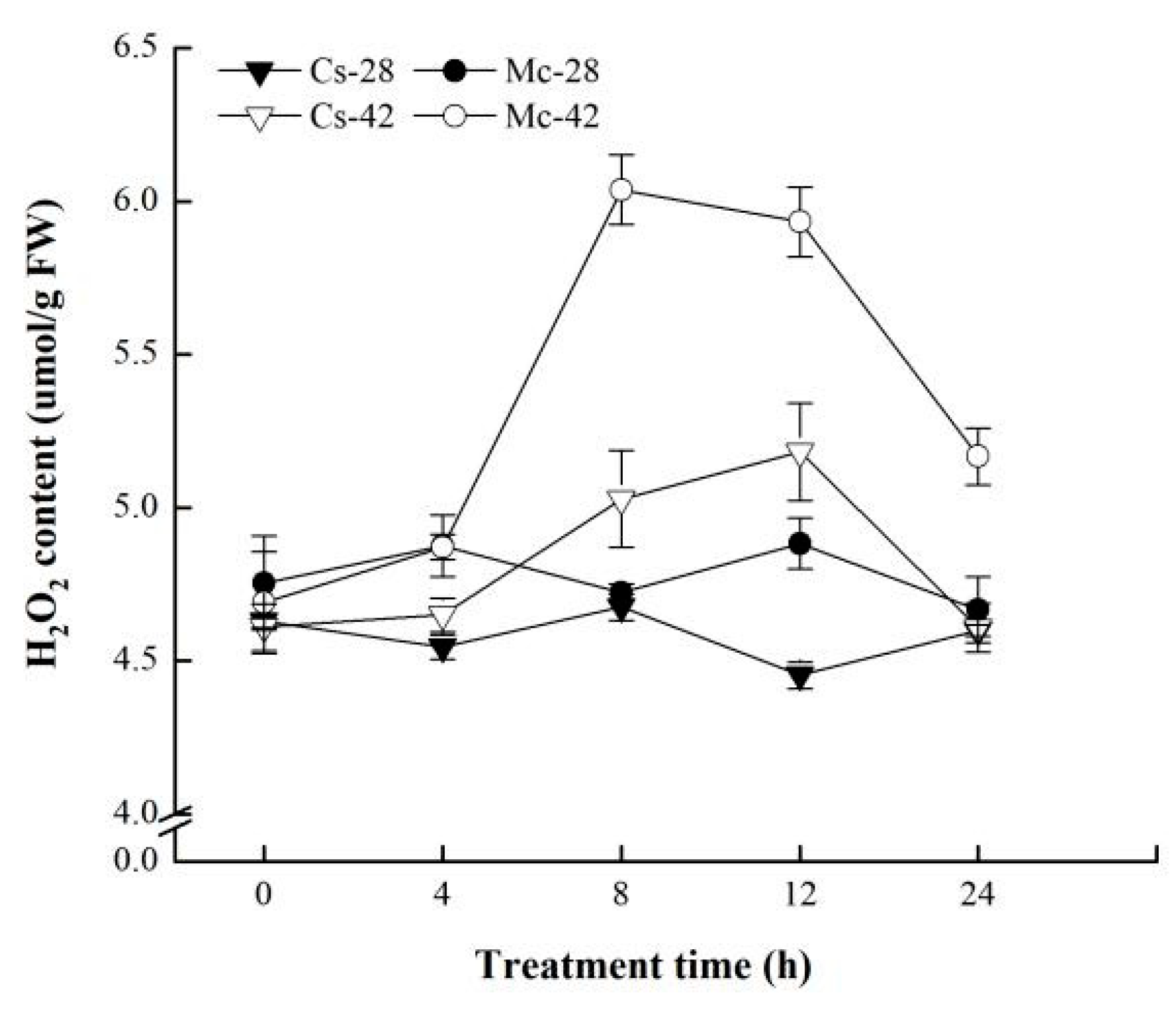
2.4. Effect of Bitter Melon Rootstock on the Accumulation of H2O2 under Heat Stress Conditions
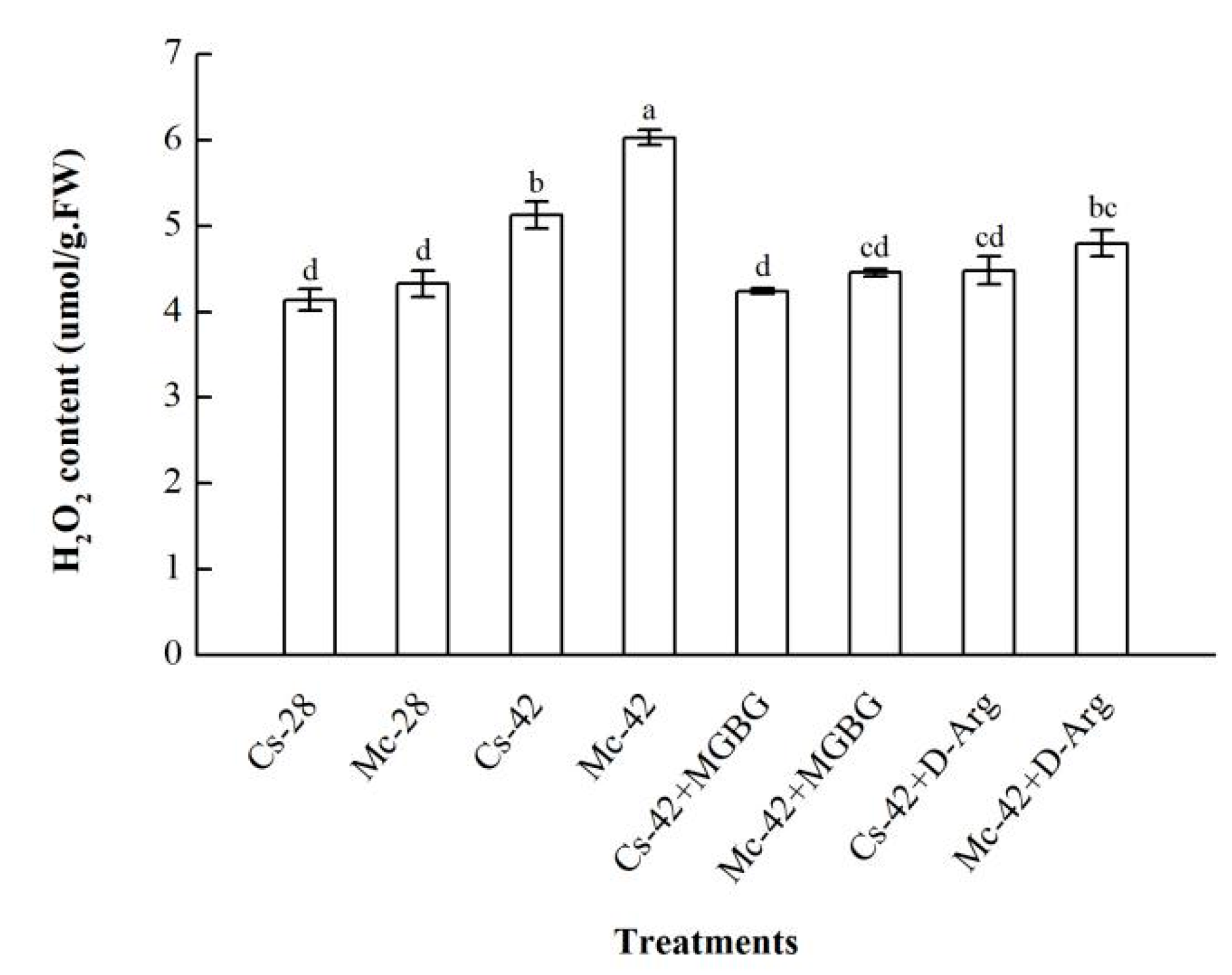
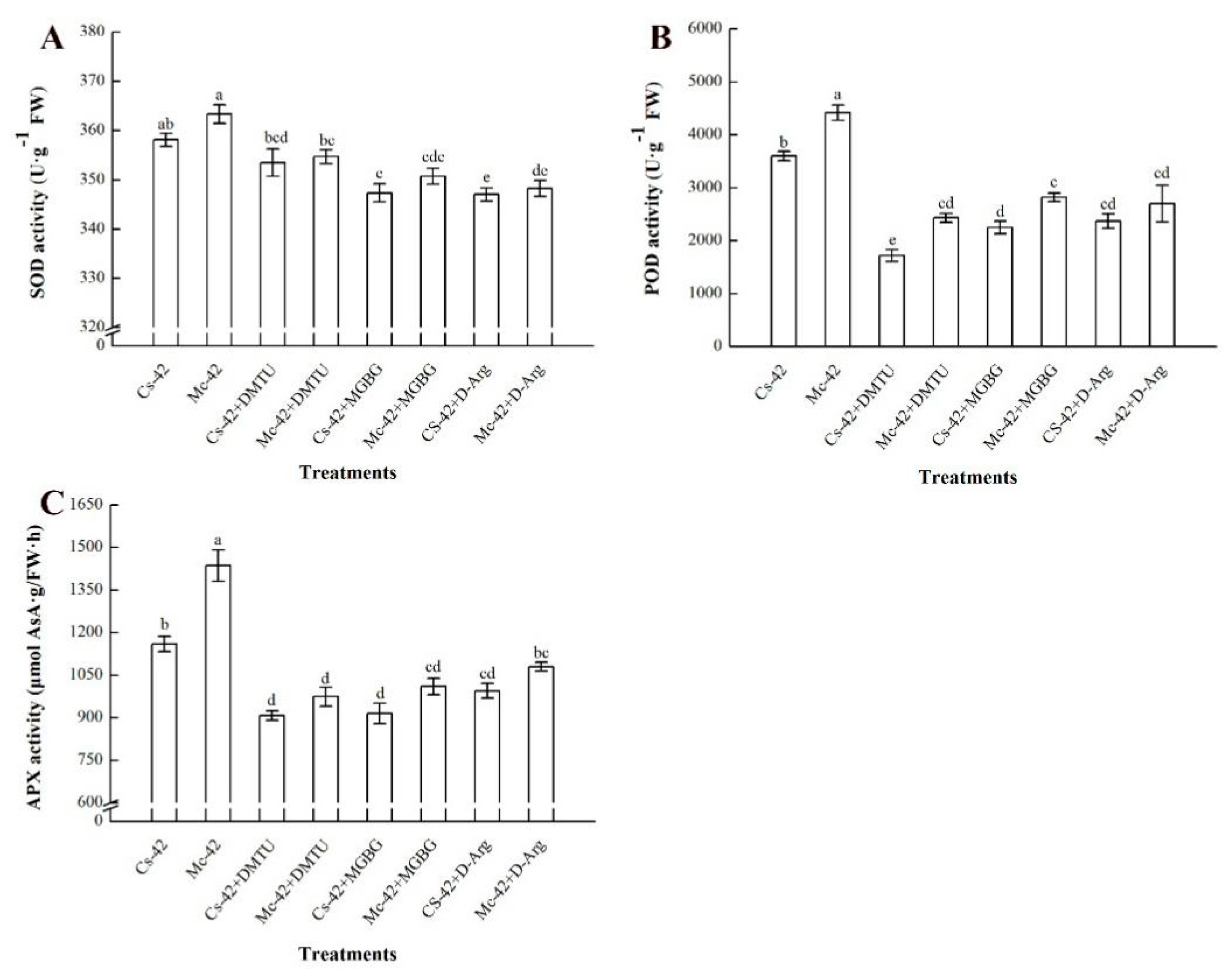
2.5. The Production of H2O2 is Regulated by Polyamines under Heat Stress Conditions
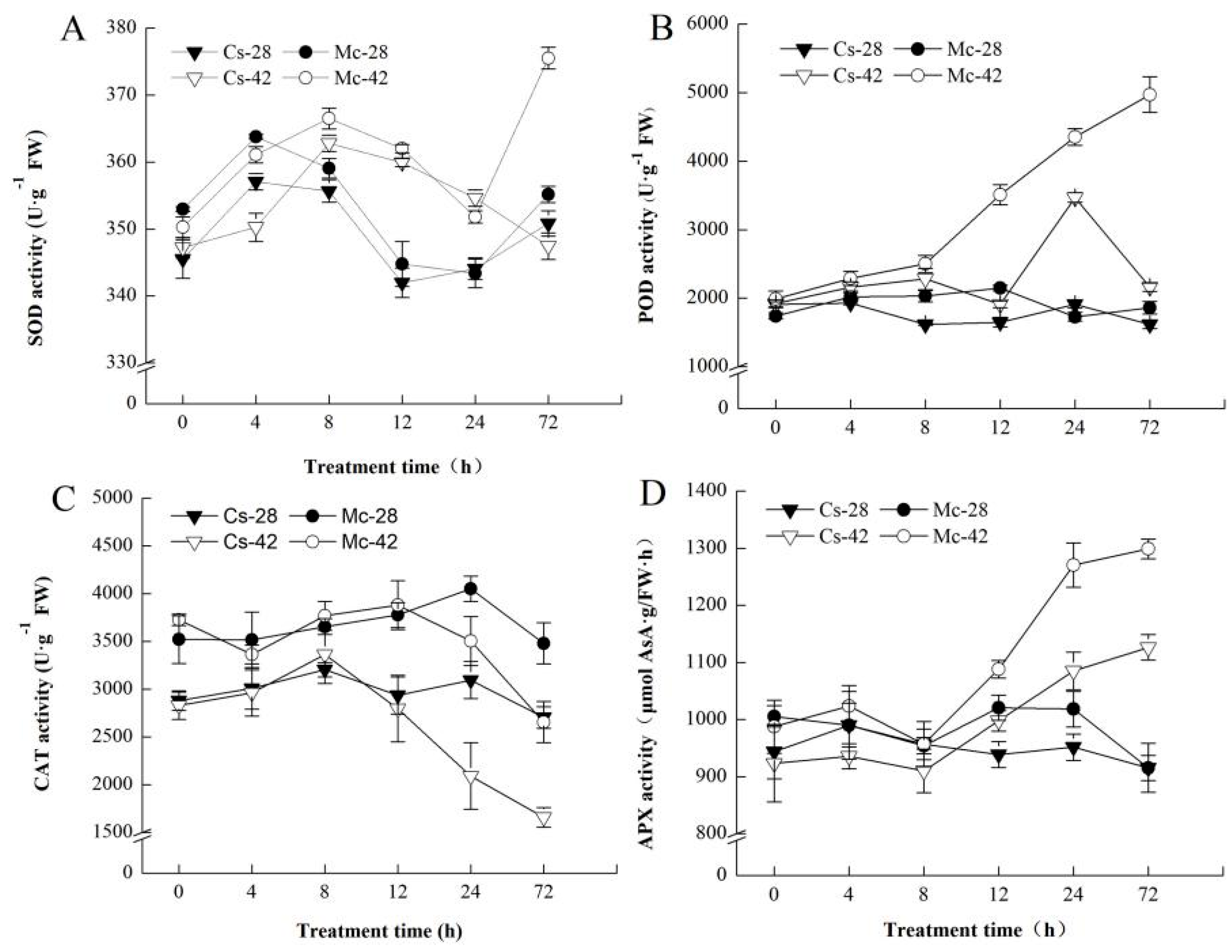
2.6. Polyamine and H2O2 Accumulation Enhanced the Antioxidant Defense System of Bitter-Melon-Grafted Seedling Leaves under Heat Stress Conditions
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Measurement of Plant Growth
4.3. Measurement of Chlorophyll Fluorescence
4.4. Quantification of Endogenous Polyamines
4.5. Determination of H2O2 Content
4.6. Assays of Antioxidant Enzyme Activity
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Dahal, K.; Li, X.-Q.; Tai, H.; Creelman, A.; Bizimungu, B. Improving Potato Stress Tolerance and Tuber Yield Under a Climate Change Scenario—A Current Overview. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Song, Q.; Yang, F.; Cui, B.; Li, J.; Zhang, Y.; Li, H.; Qiu, N.; Wang, F.; Gao, J. Physiological and molecular responses of two Chinese cabbage genotypes to heat stress. Biol. Plant. 2019, 63, 548–555. [Google Scholar] [CrossRef] [Green Version]
- Jahan, M.S.; Shu, S.; Wang, Y.; Chen, Z.; He, M.; Tao, M.; Sun, J.; Guo, S. Melatonin alleviates heat-induced damage of tomato seedlings by balancing redox homeostasis and modulating polyamine and nitric oxide biosynthesis. BMC Plant Biol. 2019, 19. [Google Scholar] [CrossRef]
- Jahan, M.S.; Wang, Y.; Shu, S.; Zhong, M.; Chen, Z.; Wu, J.; Sun, J.; Guo, S. Exogenous salicylic acid increases the heat tolerance in Tomato (Solanum lycopersicum L) by enhancing photosynthesis efficiency and improving antioxidant defense system through scavenging of reactive oxygen species. Sci. Hortic. 2019, 247, 421–429. [Google Scholar] [CrossRef]
- Xu, Y.; Yuan, Y.; Du, N.; Wang, Y.; Shu, S.; Sun, J.; Guo, S. Proteomic analysis of heat stress resistance of cucumber leaves when grafted onto Momordica rootstock. Hortic. Res. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Wang, Y.; Wu, X.; Shu, S.; Sun, J.; Guo, S. Redox and thylakoid membrane proteomic analysis reveals the Momordica (Momordica charantia L.) rootstock-induced photoprotection of cucumber leaves under short-term heat stress. Plant Physiol. Biochem. 2019, 136, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Jiang, Y.; Hao, T.; Jin, H.; Zhang, H.; He, L.; Zhou, Q.; Huang, D.; Hui, D.; Yu, J. Effects of Heat Shock on Photosynthetic Properties, Antioxidant Enzyme Activity, and Downy Mildew of Cucumber (Cucumis sativus L.). PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Shikanai, T. Physiological functions of cyclic electron transport around photosystem I in sustaining photosynthesis and plant growth. Annu. Rev. Plant Biol. 2016, 67, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Neves, L.H.; Nunes Santos, R.I.; dos Santos Teixeira, G.I.; de Araujo, D.G.; Duarte Silvestre, W.V.; Pinheiro, H.A. Leaf gas exchange, photochemical responses and oxidative damages in assai (Euterpe oleracea Mart.) seedlings subjected to high temperature stress. Sci. Hortic. 2019, 257. [Google Scholar] [CrossRef]
- Posch, B.C.; Kariyawasam, B.C.; Bramley, H.; Coast, O.; Richards, R.A.; Reynolds, M.P.; Trethowan, R.; Atkin, O.K. Exploring high temperature responses of photosynthesis and respiration to improve heat tolerance in wheat. J. Exp. Bot. 2019, 70, 5051–5069. [Google Scholar] [CrossRef]
- Rotundo, J.L.; Tang, T.; Messina, C.D. Response of maize photosynthesis to high temperature: Implications for for modeling the impact of global warming. Plant Physiol. Biochem. 2019, 141, 202–205. [Google Scholar] [CrossRef]
- Makonya, G.M.; Ogola, J.B.O.; Muasya, A.M.; Crespo, O.; Maseko, S.; Valentine, A.J.; Ottosen, C.-O.; Rosenqvist, E.; Chimphango, S.B.M. Chlorophyll fluorescence and carbohydrate concentration as field selection traits for heat tolerant chickpea genotypes. Plant Physiol. Biochem. 2019, 141, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Parvin, S.; Lee, O.R.; Sathiyaraj, G.; Khorolragchaa, A.; Kim, Y.-J.; Yang, D.-C. Spermidine alleviates the growth of saline-stressed ginseng seedlings through antioxidative defense system. Gene 2014, 537, 70–78. [Google Scholar] [CrossRef]
- Shen, J.-l.; Wang, Y.; Shu, S.; Jahan, M.S.; Zhong, M.; Wu, J.-Q.; Sun, J.; Guo, S.-R. Exogenous putrescine regulates leaf starch overaccumulation in cucumber under salt stress. Sci. Hortic. 2019, 253, 99–110. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Rahman, A.; Alam, M.M.; Jubayer-Al, M.; Suzuki, T.; Fujita, M. Polyamines Confer Salt Tolerance in Mung Bean (Vigna radiata L.) by Reducing Sodium Uptake, Improving Nutrient Homeostasis, Antioxidant Defense, and Methylglyoxal Detoxification Systems. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Yoshida, N.; Fujita, M. Spermidine pretreatment enhances heat tolerance in rice seedlings through modulating antioxidative and glyoxalase systems. Plant Growth Regul. 2014, 73, 31–44. [Google Scholar] [CrossRef]
- Song, Y.; Diao, Q.; Qi, H. Putrescine enhances chilling tolerance of tomato (Lycopersicon esculentum Mill.) through modulating antioxidant systems. Acta Physiol. Plant. 2014, 36, 3013–3027. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, E.; Romero, L.; Ruiz, J.M. Accumulation of free polyamines enhances the antioxidant response in fruits of grafted tomato plants under water stress. J. Plant Physiol. 2016, 190, 72–78. [Google Scholar] [CrossRef]
- Jia, Y.X.; Sun, J.; Guo, S.R.; Li, J.; Hu, X.H.; Wang, S.P. Effect of root-applied spermidine on growth and respiratory metabolism in roots of cucumber (Cucumis sativus) seedlings under hypoxia. Russ. J. Plant Physiol. 2010, 57, 648–655. [Google Scholar] [CrossRef]
- Reggiani, R.; Aurisano, N.; Mattana, M.; Bertani, A. Effect of K+ ions on polyamine levels in wheat seedlings under anoxia. J. Plant Physiol. 1993, 142, 94–98. [Google Scholar] [CrossRef]
- Pal, M.; Szalai, G.; Janda, T. Speculation: Polyamines are important in abiotic stress signaling. Plant Sci. 2015, 237, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Shu, S.; Li, C.; Sun, J.; Guo, S. Spermidine-mediated hydrogen peroxide signaling enhances the antioxidant capacity of salt-stressed cucumber roots. Plant Physiol. Biochem. 2018, 128, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Agurla, S.; Gayatri, G.; Raghavendra, A.S. Polyamines increase nitric oxide and reactive oxygen species in guard cells of Arabidopsis thaliana during stomatal closure. Protoplasma 2018, 255, 153–162. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Peng, D.; Wang, X.; Peng, Y.; He, X.; Zhang, X.; Ma, X.; Huang, L.; Yan, Y. Polyamine regulates tolerance to water stress in leaves of white clover associated with antioxidant defense and dehydrin genes via involvement in calcium messenger system and hydrogen peroxide signaling. Front. Physiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Gong, B.; Wan, X.; Wei, M.; Li, Y.; Wei, M.; Shi, Q. Overexpression of S-adenosylmethionine synthetase 1 enhances tomato callus tolerance to alkali stress through polyamine and hydrogen peroxide cross-linked networks. Plant Cell Tissue Organ Cult. 2016, 124, 377–391. [Google Scholar] [CrossRef]
- Zepeda-Jazo, I.; Maria Velarde-Buendia, A.; Enriquez-Figueroa, R.; Bose, J.; Shabala, S.; Muniz-Murguia, J.; Pottosin, I.I. Polyamines Interact with Hydroxyl Radicals in Activating Ca2+ and K+ Transport across the Root Epidermal Plasma Membranes. Plant Physiol. 2011, 157, 2167–2180. [Google Scholar] [CrossRef] [Green Version]
- Petropoulos, S.A.; Khah, E.M.; Passam, H.C. Evaluation of rootstocks for watermelon grafting with reference to plant development, yield and fruit quality. Int. J. Plant Prod. 2012, 6, 481–491. [Google Scholar]
- Rouphael, Y.; Cardarelli, M.; Rea, E.; Colla, G. Improving melon and cucumber photosynthetic activity, mineral composition, and growth performance under salinity stress by grafting onto Cucurbita hybrid rootstocks. Photosynthetica 2012, 50, 180–188. [Google Scholar] [CrossRef]
- Tripodi, G.; Condurso, C.; Cincotta, F.; Merlino, M.; Verzera, A. Aroma compounds in mini-watermelon fruits from different grafting combinations. J. Sci. Food Agric. 2020, 100, 1328–1335. [Google Scholar] [CrossRef]
- Mudge, K.; Janick, J.; Scofield, S.; Goldschmidt, E. A History of Grafting. Hortic. Rev. 2009, 35. [Google Scholar] [CrossRef]
- Li, H.; Ahammed, G.J.; Zhou, G.; Xia, X.; Zhou, J.; Shi, K.; Yu, J.; Zhou, Y. Unraveling Main Limiting Sites of Photosynthesis under Below- and Above-Ground Heat Stress in Cucumber and the Alleviatory Role of Luffa Rootstock. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Wang, F.; Chen, X.-J.; Shi, K.; Xia, X.-J.; Considine, M.J.; Yu, J.-Q.; Zhou, Y.-H. The sub/supra-optimal temperature-induced inhibition of photosynthesis and oxidative damage in cucumber leaves are alleviated by grafting onto figleaf gourd/luffa rootstocks. Physiol. Plant. 2014, 152, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, S.-S.; Yi, C.-Y.; Wang, F.; Zhou, J.; Xia, X.-J.; Shi, K.; Zhou, Y.-H.; Yu, J.-Q. Hydrogen peroxide mediates abscisic acid-induced HSP70 accumulation and heat tolerance in grafted cucumber plants. Plant Cell Environ. 2014, 37, 2768–2780. [Google Scholar] [CrossRef]
- Khraiwesh, B.; Zhu, J.-K.; Zhu, J. Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochim. Biophys. Acta Gene Regul. Mech. 2012, 1819, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Xiang, N.; Hu, J.; Wen, T.; Brennan, M.A.; Brennan, C.S.; Guo, X. Effects of temperature stress on the accumulation of ascorbic acid and folates in sweet corn (Zea mays L.) seedlings. J. Sci. Food Agric. 2019. [Google Scholar] [CrossRef]
- Hsuan, T.-P.; Jhuang, P.-R.; Wu, W.-C.; Lur, H.-S. Thermotolerance evaluation of Taiwan Japonica type rice cultivars at the seedling stage. Bot. Stud. 2019, 60. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Bie, Z.; Huang, Y.; Zhen, A.; Niu, M.; Lei, B. Rootstocks improve cucumber photosynthesis through nitrogen metabolism regulation under salt stress. Acta Physiol. Plant. 2013, 35, 2259–2267. [Google Scholar] [CrossRef]
- Li, L.; Xing, W.-W.; Shao, Q.-S.; Shu, S.; Sun, J.; Guo, S.-R. The effects of grafting on glycolysis and the tricarboxylic acid cycle in Ca(NO3)(2)-stressed cucumber seedlings with pumpkin as rootstock. Acta Physiol. Plant. 2015, 37. [Google Scholar] [CrossRef]
- Huve, K.; Bichele, I.; Tobias, M.; Niinemets, U. Heat sensitivity of photosynthetic electron transport varies during the day due to changes in sugars and osmotic potential. Plant Cell Environ. 2006, 29, 212–228. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Wang, Z.; Guo, X.; Wang, F.; Xia, X.-J.; Zhou, J.; Shi, K.; Yu, J.-Q.; Zhou, Y.-H. Microarray and genetic analysis reveals that csa-miR159b plays a critical role in abscisic acid-mediated heat tolerance in grafted cucumber plants. Plant Cell Environ. 2016, 39, 1790–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcazar, R.; Altabella, T.; Marco, F.; Bortolotti, C.; Reymond, M.; Koncz, C.; Carrasco, P.; Tiburcio, A.F. Polyamines: Molecules with regulatory functions in plant abiotic stress tolerance. Planta 2010, 231, 1237–1249. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Fujita, M. Exogenous Spermidine Alleviates Low Temperature Injury in Mung Bean (Vigna radiata L.) Seedlings by Modulating Ascorbate-Glutathione and Glyoxalase Pathway. Int. J. Mol. Sci. 2015, 16, 30117–30132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.-H.; Wang, W.; Wu, H.; Gong, X.; Moriguchi, T. Polyamines function in stress tolerance: From synthesis to regulation. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Saxena, I.; Srikanth, S.; Chen, Z. Cross Talk between H2O2 and Interacting Signal Molecules under Plant Stress Response. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, J.; Li, J.-L.; Ma, X.-R. Exogenous hydrogen peroxide enhanced the thermotolerance of Festuca arundinacea and Lolium perenne by increasing the antioxidative capacity. Acta Physiol. Plant. 2014, 36, 2915–2924. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Hu, X.; Zhang, J.; Zhang, J.; Du, Q.; Li, J. H2O2 mediates ALA-induced glutathione and ascorbate accumulation in the perception and resistance to oxidative stress in Solanum lycopersicum at low temperatures. BMC Plant Biol. 2018, 18. [Google Scholar] [CrossRef] [Green Version]
- Gupta, K.; Sengupta, A.; Chakraborty, M.; Gupta, B. Hydrogen Peroxide and Polyamines Actas Double Edged Swords in Plant Abiotic Stress Responses. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Diao, Q.; Song, Y.; Shi, D.; Qi, H. Interaction of Polyamines, Abscisic Acid, Nitric Oxide, and Hydrogen Peroxide under Chilling Stress in Tomato (Lycopersicon esculentum Mill.) Seedlings. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Shu, S.; Gao, P.; Li, L.; Yuan, Y.; Sun, J.; Guo, S. Abscisic Acid-Induced H2O2 Accumulation Enhances Antioxidant Capacity in Pumpkin-Grafted Cucumber Leaves under Ca(NO3)(2) Stress. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Jahan, M.S.; Guo, S.; Baloch, A.R.; Sun, J.; Shu, S.; Wang, Y.; Ahammed, G.J.; Kabir, K.; Roy, R. Melatonin alleviates nickel phytotoxicity by improving photosynthesis, secondary metabolism and oxidative stress tolerance in tomato seedlings. Ecotoxicol. Environ. Saf. 2020, 197, 110593. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, M.; Saleem, M.F.; Ullah, N.; Rizwan, M.; Ali, S.; Shahid, M.R.; Alamri, S.A.; Alyemeni, M.N.; Ahmad, P. Exogenously applied growth regulators protect the cotton crop from heat-induced injury by modulating plant defense mechanism. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Chen, X.; Li, L.; Niu, H.; Yi, Y. Effects of high temperature stress on antioxidant enzyme activities in leaves of different wheat varieties. Henan Agric. Sci. 2008, 38–40. [Google Scholar]
- Lu, C.M.; Qiu, N.W.; Wang, B.S.; Zhang, J.H. Salinity treatment shows no effects on photosystem II photochemistry, but increases the resistance of photosystem II to heat stress in halophyte Suaeda salsa. J. Exp. Bot. 2003, 54, 851–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, S.; Guo, S.-R.; Sun, J.; Yuan, L.-Y. Effects of salt stress on the structure and function of the photosynthetic apparatus in Cucumis sativus and its protection by exogenous putrescine. Physiol. Plant. 2012, 146, 285–296. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Tan, W.; Liu, J.; Dai, T.; Jing, Q.; Cao, W.; Jiang, D. Alterations in photosynthesis and antioxidant enzyme activity in winter wheat subjected to post-anthesis water-logging. Photosynthetica 2008, 46, 21–27. [Google Scholar] [CrossRef]
- Dhindsa, R.; Plumb-Dhindsa, P.; Reid, D. Leaf senescence and lipid peroxidation: Effects of some phytohormones, and scavengers of free radicals and singlet oxygen. Physiol. Plant. 2006, 56, 453–457. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1980, 22. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height (cm) | Stem Diameter (mm) | Fresh Weight (Plant g−1) | Dry Weight (Plant g−1) |
|---|---|---|---|---|
| Cs-28 | 18.37 ± 0.32 a | 5.03 ± 0.13 bc | 17.19 ± 0.01 a | 1.83 ± 0.03 a |
| Mc-28 | 16.27 ± 0.12 b | 5.62 ± 0.14 a | 16.62 ± 0.15 b | 1.69 ± 0.01 b |
| Cs-42 | 17.63 ± 0.23 a | 4.79 ± 0.12 c | 13.70 ± 0.21 d | 1.29 ± 0.04 d |
| Mc-42 | 16.23 ± 0.54 b | 5.39 ± 0.09 ab | 15.58 ± 0.21 c | 1.42 ± 0.02 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, M.-Q.; Jahan, M.S.; Hou, K.; Shu, S.; Wang, Y.; Sun, J.; Guo, S.-R. Bitter Melon (Momordica charantia L.) Rootstock Improves the Heat Tolerance of Cucumber by Regulating Photosynthetic and Antioxidant Defense Pathways. Plants 2020, 9, 692. https://doi.org/10.3390/plants9060692
Tao M-Q, Jahan MS, Hou K, Shu S, Wang Y, Sun J, Guo S-R. Bitter Melon (Momordica charantia L.) Rootstock Improves the Heat Tolerance of Cucumber by Regulating Photosynthetic and Antioxidant Defense Pathways. Plants. 2020; 9(6):692. https://doi.org/10.3390/plants9060692
Chicago/Turabian StyleTao, Mei-Qi, Mohammad Shah Jahan, Kun Hou, Sheng Shu, Yu Wang, Jin Sun, and Shi-Rong Guo. 2020. "Bitter Melon (Momordica charantia L.) Rootstock Improves the Heat Tolerance of Cucumber by Regulating Photosynthetic and Antioxidant Defense Pathways" Plants 9, no. 6: 692. https://doi.org/10.3390/plants9060692