Salmonella Heterogeneously Expresses Flagellin during Colonization of Plants

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Culture Conditions and Media Preparation
2.2. Fluorescent Labeling of Salmonella Strains
2.3. Plant Cultivation
2.4. Confocal Laser Scanning Microscopy (CLSM)
2.5. Persistence of Salmonella in Plants
2.6. Translocation of Salmonella within Tomato Plants
2.7. Competitive Bacterial Colonization Assays
2.8. Quantitative Real Time PCR Analysis
2.9. Salmonella Response to Plant Media
2.10. Western Blot Analysis
2.11. Quantification of Flagellin Protein in Different Media
2.12. Flow Cytometry and Confocal Laser Scanning Microscopy (CLSM)
3. Results
3.1. Salmonella Does Not Require Flagella to Persist and to Colonize Plants
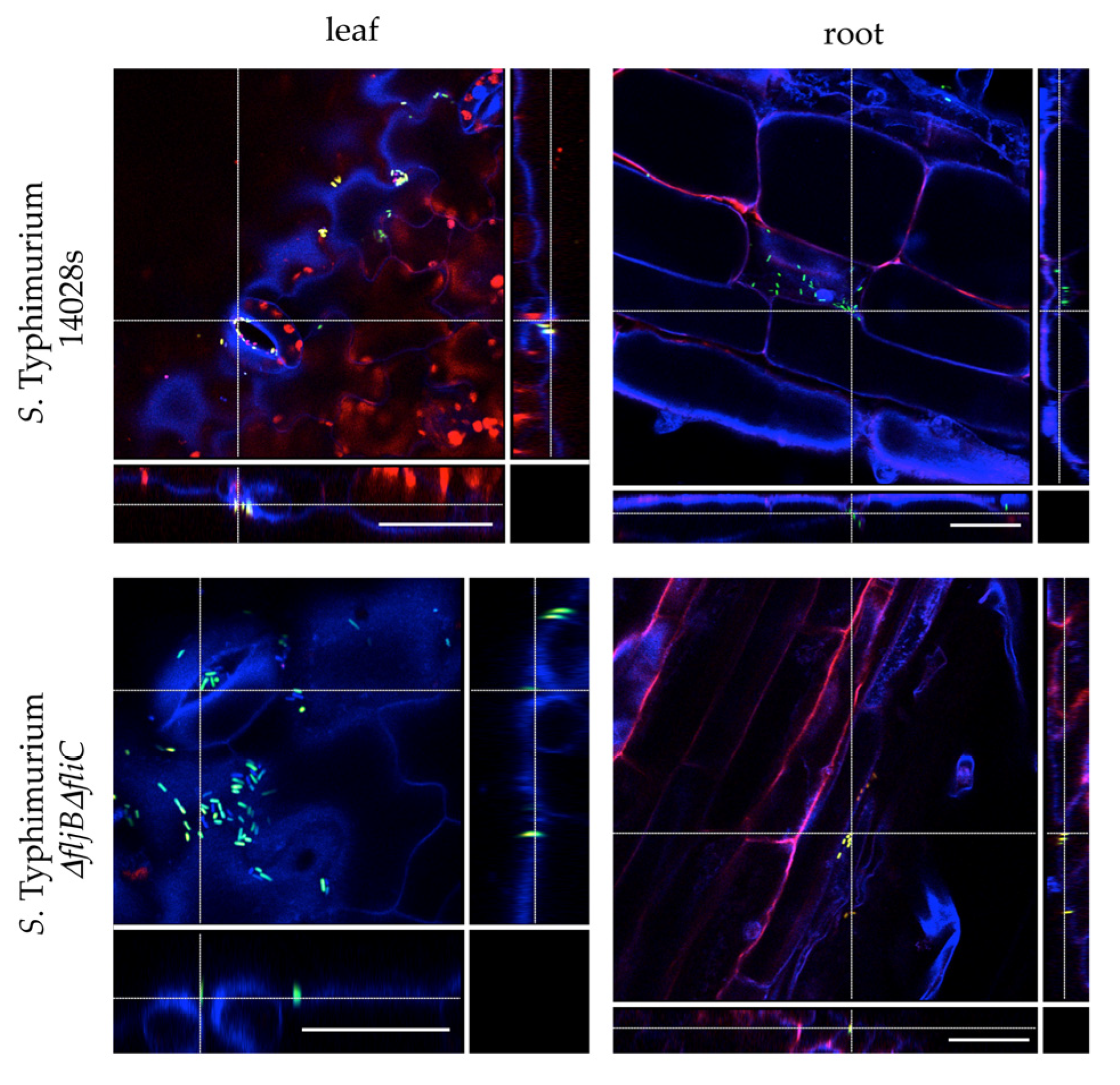
3.2. Salmonella Strain Lacking Flagellin Displays a Colonization Pattern on Tomato Leaves Similar to That of the Wild Type
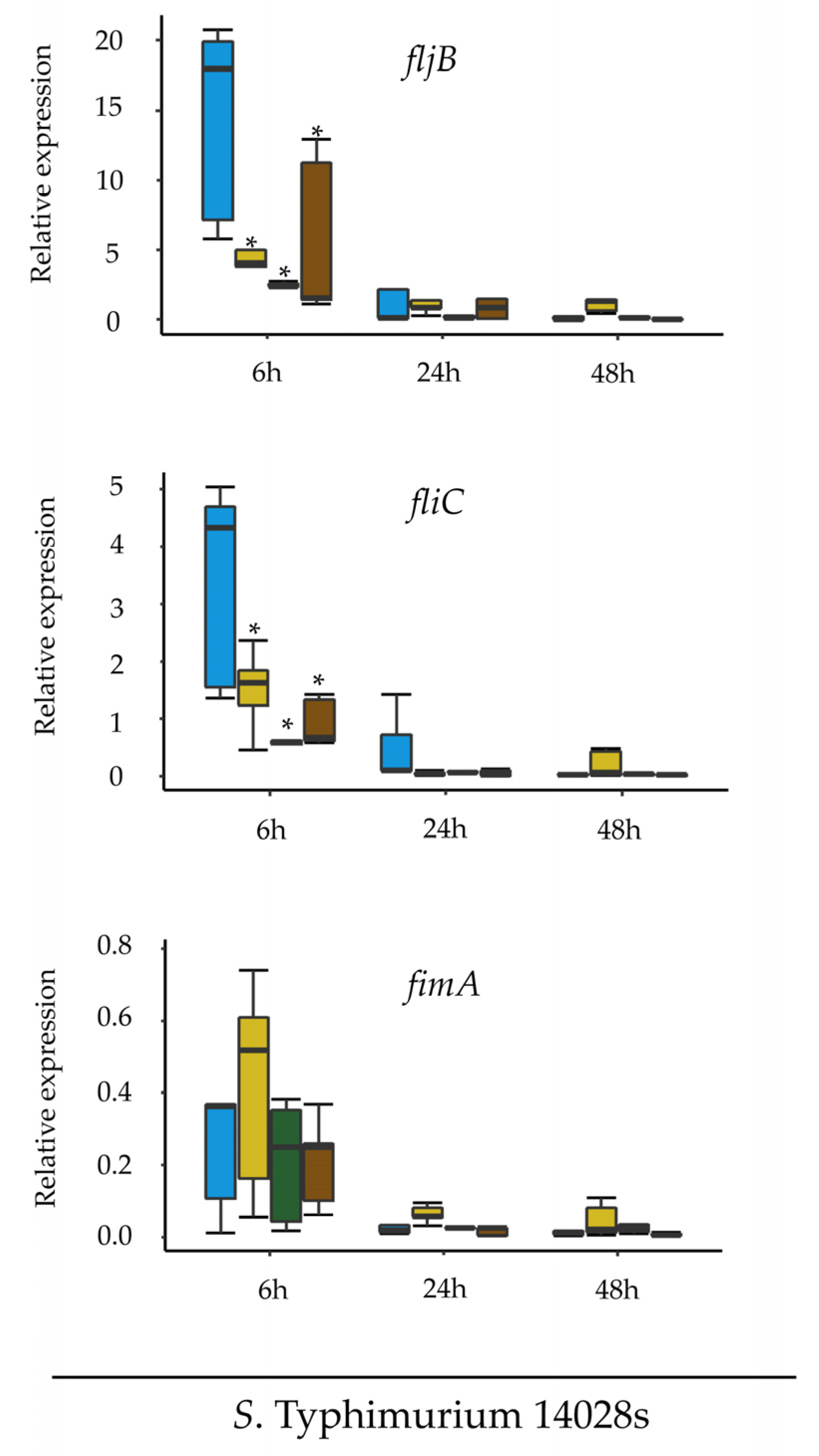
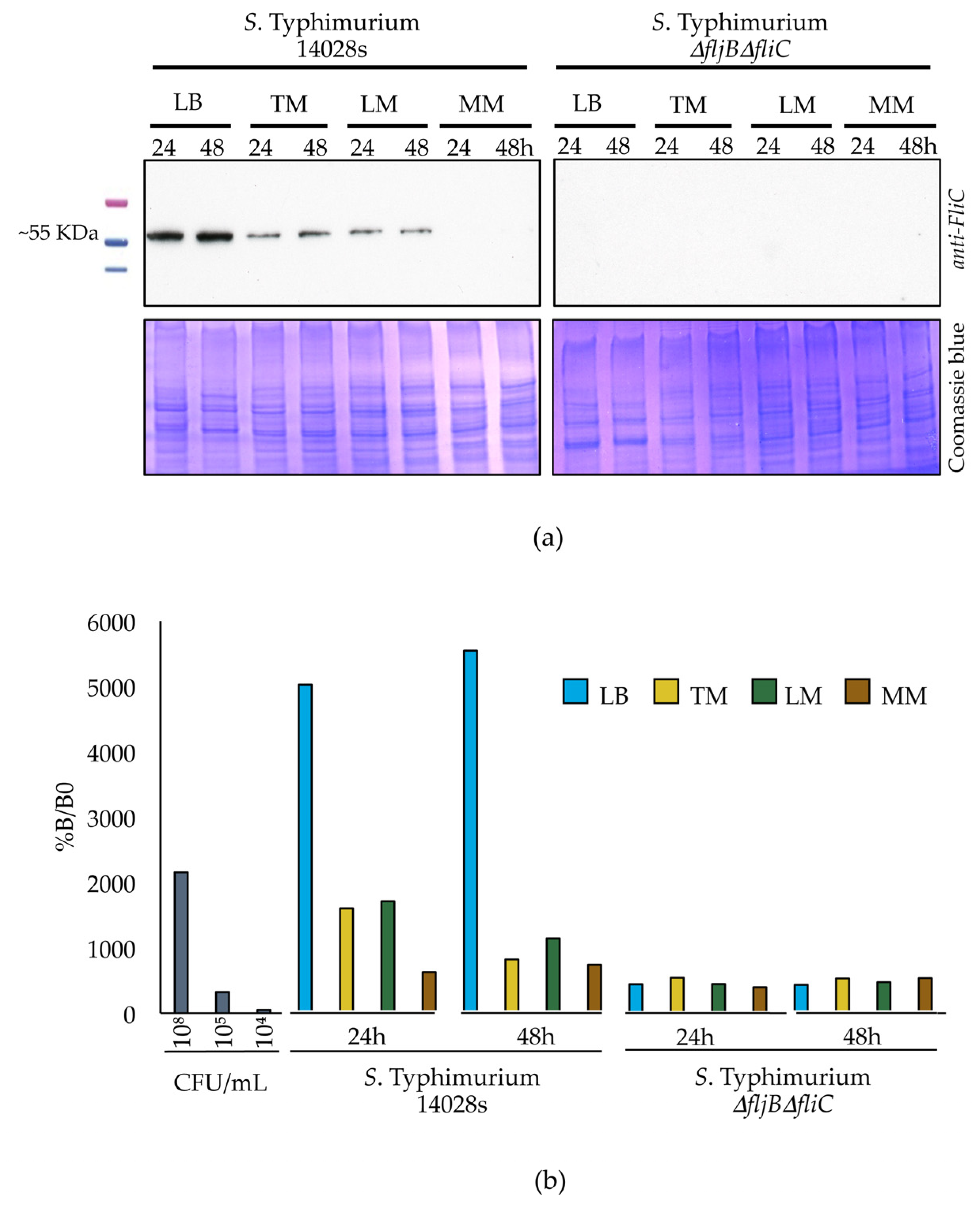
3.3. Expression of Flagellin-Coding Genes Is Down-Regulated in Plant-Mimicking Media
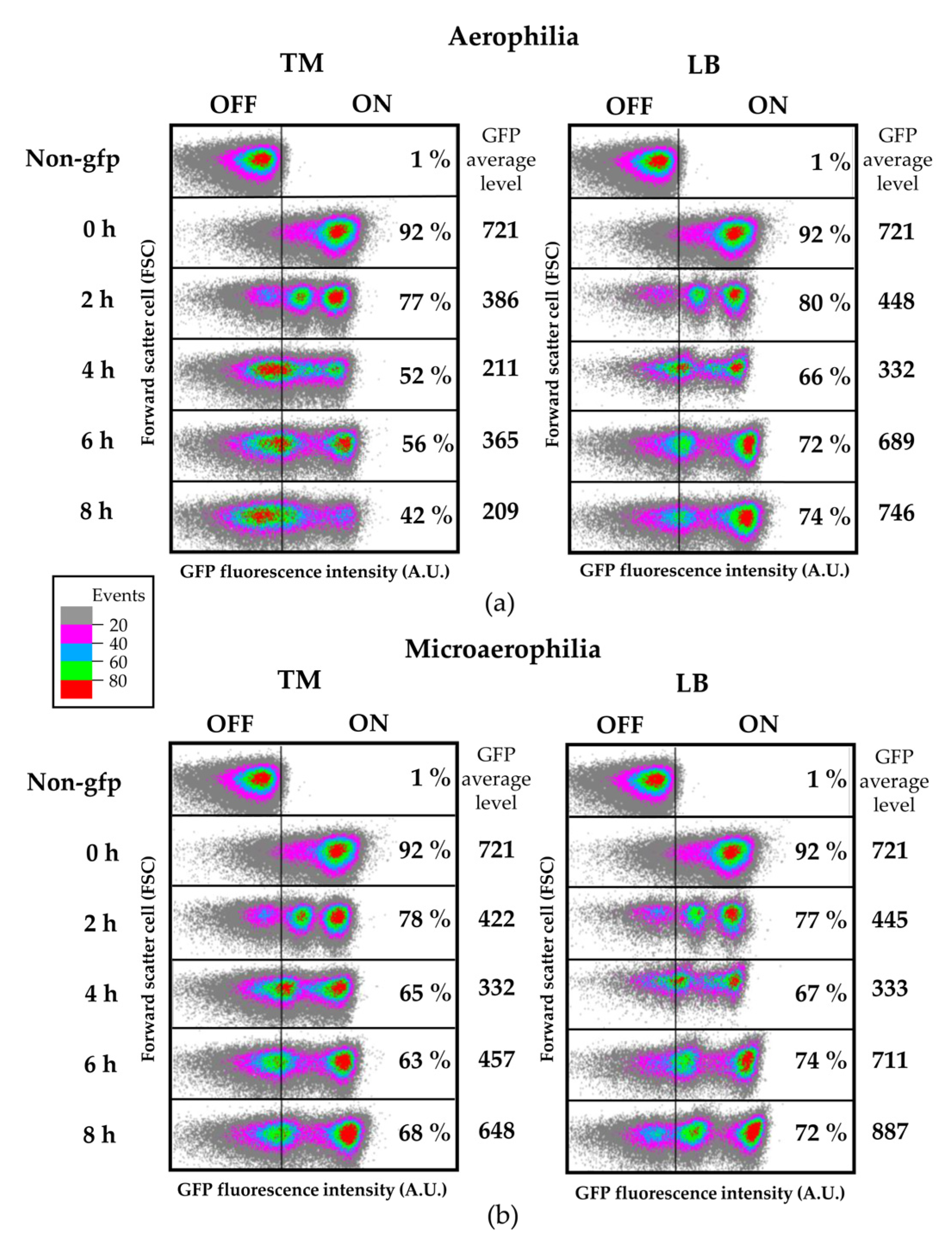
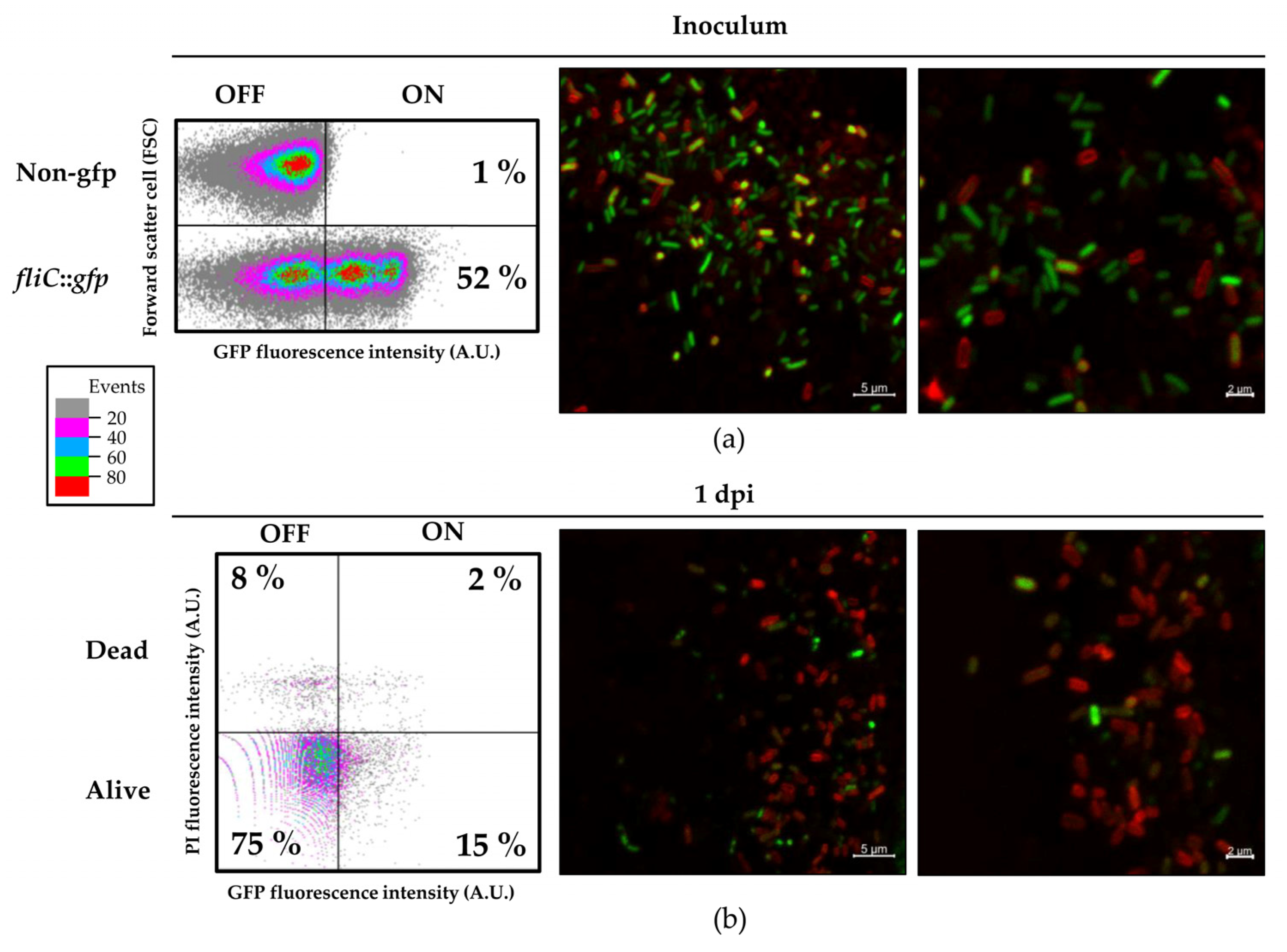
3.4. Flagellin Is Heterogeneously Expressed in the Plant Environment
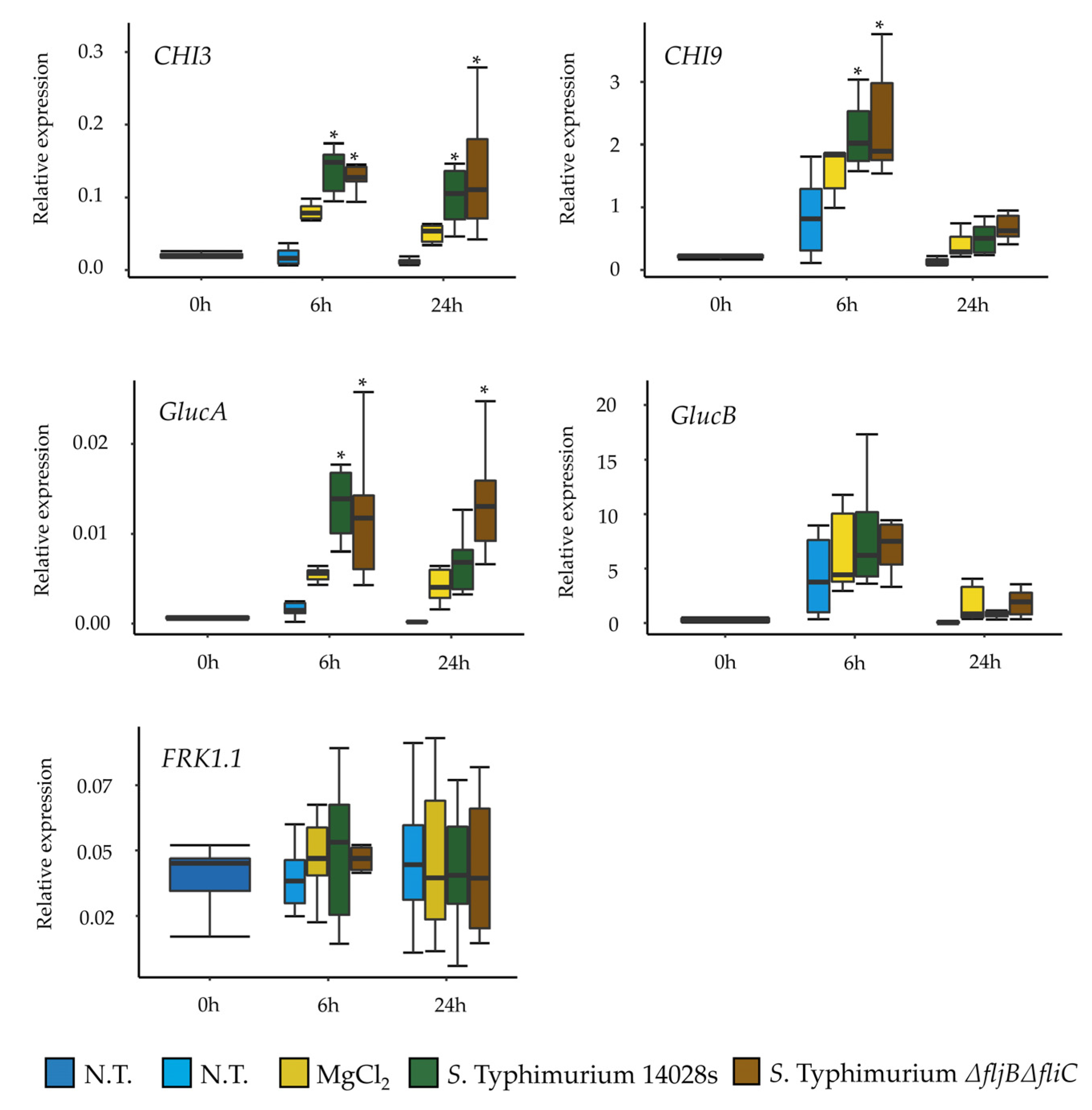
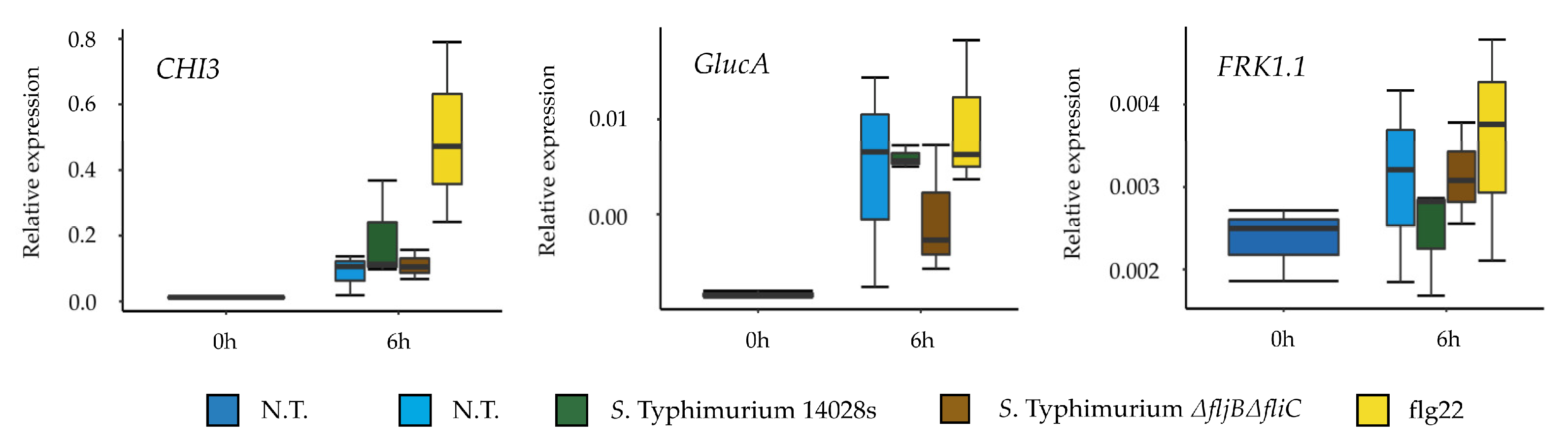
3.5. Recognition of Salmonella in Tomato Is Not Exclusively Based on Flagellin
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Available online: https://www.who.int/news-room/fact-sheets/detail/Salmonella-(non-typhoidal) (accessed on 28 April 2020).
- Zhang, S.; Kingsley, R.; Santos, R.; Andrews-Polymenis, H.; Raffatellu, M.; Figueiredo, J.; Nunes, J.; Tsolis, R.; Adams, G.; Bäumler, A.; et al. Molecular pathogenesis of Salmonella enterica serotype typhimurium-induced diarrhea. Infect. Immun. 2003, 71, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daoguo, Z.; Mark, S.; Jorge, E. Role of the S. typhimurium actin-binding protein SipA in bacterial internalization. Science 1999, 283, 2092. [Google Scholar] [CrossRef] [Green Version]
- Agnès, W.; Isabelle, V.; Anne-Marie, C.; Schikora, A.; Velge, P. Interactions of Salmonella with animals and plants. Front. Microbiol. 2014, 5, 791. [Google Scholar] [CrossRef]
- Heather, R.; Kelly, T. Flagellar phase variation in Salmonella enterica is mediated by a posttranscriptional control mechanism. J. Bacteriol. 2003, 185, 3567. [Google Scholar] [CrossRef] [Green Version]
- Bradley, D.; Nafisa, G.; Stanley, F. Salmonella typhimurium initiates murine infection by penetrating and destroying the specialized epithelial M cells of the Peyer’s patches. J. Exp. Med. 1994, 180, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Sansonetti, P. Host-pathogen interactions: The seduction of molecular cross talk. Gut 2002, 50 (Suppl. 3), III2–III8. [Google Scholar] [CrossRef]
- Erhardt, M.; Hirano, T.; Su, Y.; Wee, D.; Mizuno, S.; Aizawa, S.; Hughes, K. The role of the FliK molecular ruler in hook-length control in Salmonella enterica. Mol. Microbiol. 2010, 75, 1272–1284. [Google Scholar] [CrossRef] [Green Version]
- Erhardt, M.; Singer, H.; Wee, D.; Keener, J.; Hughes, K. An infrequent molecular ruler controls flagellar hook length in Salmonella enterica. EMBO J. 2011, 30, 2948–2961. [Google Scholar] [CrossRef] [Green Version]
- Horstmann, J.; Zschieschang, E.; Truschel, T.; Diego, J.; Lunelli, M.; Rohde, M.; May, T.; Strowig, T.; Stradal, T.; Kolbe, M.; et al. Flagellin phase-dependent swimming on epithelial cell surfaces contributes to productive Salmonella gut colonisation. Cell. Microbiol. 2017, 19. [Google Scholar] [CrossRef] [Green Version]
- Misselwitz, B.; Barrett, N.; Kreibich, S.; Vonaesch, P.; Andritschke, D.; Rout, S.; Weidner, K.; Sormaz, M.; Songhet, P.; Horvath, P.; et al. Near surface swimming of Salmonella Typhimurium explains target-site selection and cooperative invasion. PLoS Pathog. 2012, 8, e1002810. [Google Scholar] [CrossRef] [Green Version]
- Chaban, B.; Hughes, H.; Beeby, M. The flagellum in bacterial pathogens: For motility and a whole lot more. Semin. Cell Dev. Biol. 2015, 46, 91–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, F.; Smith, K.; Ozinsky, A.; Hawn, T.; Yi, E.; Goodlett, D.; Eng, J.; Akira, S.; Underhill, D.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gómez, L.; Boller, T. FLS2: An LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Mol. Cell 2000, 5, 1003–1011. [Google Scholar] [CrossRef]
- Meindl, T.; Boller, T.; Felix, G. The bacterial elicitor flagellin activates its receptor in tomato cells according to the address-message concept. Plant Cell 2000, 12, 1783–1794. [Google Scholar] [CrossRef] [Green Version]
- Felix, G.; Duran, J.; Volko, S.; Boller, T. Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J. 1999, 18, 265–276. [Google Scholar] [CrossRef]
- Sun, W.; Dunning, F.; Pfund, C.; Weingarten, R.; Bent, A. Within-species flagellin polymorphism in Xanthomonas campestris pv campestris and its impact on elicitation of Arabidopsis FLAGELLIN SENSING2-dependent defenses. Plant Cell 2006, 18, 764–779. [Google Scholar] [CrossRef] [Green Version]
- Cai, R.; Lewis, J.; Yan, S.; Liu, H.; Clarke, C.; Campanile, F.; Almeida, N.; Studholme, D.; Lindeberg, M.; Schneider, D.; et al. The plant pathogen Pseudomonas syringae pv. tomato is genetically monomorphic and under strong selection to evade tomato immunity. PLoS Pathog. 2011, 7, e1002130. [Google Scholar] [CrossRef]
- Clarke, C.; Chinchilla, D.; Hind, S.; Taguchi, F.; Miki, R.; Ichinose, Y.; Martin, G.; Leman, S.; Felix, G.; Vinatzer, B. Allelic variation in two distinct Pseudomonas syringae flagellin epitopes modulates the strength of plant immune responses but not bacterial motility. New Phytol. 2013, 200, 847–860. [Google Scholar] [CrossRef] [Green Version]
- Shaw, R.; Lasa, I.; García, B.; Pallen, M.; Hinton, J.; Berger, C.; Frankel, G. Cellulose mediates attachment of Salmonella enterica Serovar Typhimurium to tomatoes. Environ. Microbiol. Rep. 2011, 3, 569–573. [Google Scholar] [CrossRef]
- Berger, C.; Shaw, R.; Brown, D.; Ahmer, B.; Stone, J.; Triplett, E. Interaction of Salmonella enterica with basil and other salad leaves. ISME J. 2009, 3, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Iniguez, A.; Dong, D.; Carter, H.; Ahmer, B.M.; Stone, J.M.; Triplett, E.W. Regulation of enteric endophytic bacterial colonization by plant defenses. MPMI 2005, 18, 169–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elpers, L.; Kretzschmar, J.; Nuccio, S.; Bäumler, A.; Hensel, M. Factors Required for Adhesion of Salmonella enterica Serovar Typhimurium to Corn Salad (Valerianella locusta). Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Toma, S.; Terahara, N.; Miyata, T.; Ashihara, M.; Minamino, T.; Namba, K.; Kato, T. Structural and Functional Comparison of Salmonella Flagellar Filaments Composed of FljB and FliC. Biomolecules 2020, 10, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartz, J.; Marvasi, M.; Teplitski, M. Salmonella and Tomatoes. In The Produce Contamination Problem, 2nd ed.; Matthews, K., Sapers, J., Gerba, C., Eds.; Academic Press: San Diego, CA, USA, 2014; Chapter 12; pp. 269–289. [Google Scholar]
- Fornefeld, E.; Schierstaedt, J.; Jechalke, S.; Grosch, R.; Schikora, A.; Smalla, K. Persistence of Salmonella Typhimurium LT2 in soil enhanced after growth in lettuce medium. Front. Microbiol. 2017, 8, 757. [Google Scholar] [CrossRef]
- Zarkani, A.; Schierstaedt, J.; Becker, M.; Krumwiede, J.; Grimm, M.; Grosch, R.; Jechalke, S.; Schikora, A. Salmonella adapts to plants and their environment during colonization of tomatoes. FEMS Microbiol. Ecol. 2019, 95. [Google Scholar] [CrossRef]
- Christensen, B.; Haagensen, J.; Heydorn, A.; Molin, S. Metabolic commensalism and competition in a two- species microbial consortium. Appl. Environ. Microbiol. 2002, 68, 2495–2502. [Google Scholar] [CrossRef] [Green Version]
- Jechalke, S.; Schierstaedt, J.; Becker, M.; Flemer, B.; Grosch, R.; Smalla, K.; Schikora, A. Salmonella establishment in agricultural soil and colonization of crop plants depend on soil type and plant species. Front. Microbiol. 2019, 10, 967. [Google Scholar] [CrossRef] [Green Version]
- Richter, B.; Smalla, K. Screening of rhizosphere and soil bacteria for transformability. Environ. Biosaf. Res. 2007, 6, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Thorsted, P.; Macartney, D.; Akhtar, P.; Haines, A.; Ali, N.; Davidson, P.; Stafford, T.; Pocklington, M.; Pansegrau, W.; Wilkins, B.; et al. Complete sequence of the IncPbeta plasmid R751: Implications for evolution and organisation of the IncP backbone. J. Mol. Biol. 1998, 282, 969–990. [Google Scholar] [CrossRef]
- Hautefort, I.; Proença, M.; Hinton, J. Single-copy green fluorescent protein gene fusions allow accurate measurement of Salmonella gene expression in vitro and during infection of mammalian cells. Appl. Environ. Microbiol. 2003, 69, 7480. [Google Scholar] [CrossRef] [Green Version]
- Datsenko, K.; Wanner, B. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640. [Google Scholar] [CrossRef] [Green Version]
- Macho, A.; Rufián, J.; Ruiz-Albert, J.; Beuzón, C. Competitive Index: Mixed Infection-Based Virulence Assays for Genetic Analysis in Pseudomonas syringae-Plant Interactions. Methods Mol. Biol. 2016, 1363, 209–217. [Google Scholar] [CrossRef]
- Freter, R.; O’Brien, P.; Macsai, M. Role of chemotaxis in the association of motile bacteria with intestinal mucosa: In vitro studies. Infect. Immun. 1981, 34, 241–249. [Google Scholar]
- Taylor, R.; Miller, V.; Furlong, D. Use of phoA gene fusions to identify a pilus colonization factor coordinately regulated with cholera toxin. Proc. Natl. Acad. Sci. USA 1987, 84, 2833–2837. [Google Scholar] [CrossRef] [Green Version]
- Beuzon, C.; Holden, D. Use of mixed infections with Salmonella strains to study virulence genes and their interactions in vivo. Microbes Infect. 2001, 3, 1345–1352. [Google Scholar]
- Macho, A.; Zumaquero, A.; Ortiz-Martín, I.; Beuzón, C. Competitive index in mixed infections: A sensitive and accurate assay for the genetic analysis of Pseudomonas syringae–plant interactions. Mol. Plant Pathol. 2007, 8, 437–450. [Google Scholar] [CrossRef]
- Macho, A.; Guidot, A.; Barberis, P.; Beuzón, C.; Genin, S. A Competitive index assay identifies several Ralstonia solanacearum Type III effector mutant strains with reduced fitness in host plants. Mol. Plant Microbe Interact. 2010, 23, 1197–1205. [Google Scholar] [CrossRef] [Green Version]
- Zeiner, A.; Dwyer, B.; Clegg, S. FimA, FimF, and FimH Are Necessary for Assembly of Type 1 Fimbriae on Salmonella enterica Serovar Typhimurium. Infect. Immun. 2012, 80, 3289–3296. [Google Scholar] [CrossRef] [Green Version]
- Freed, N.; Silander, O.; Stecher, B.; Böhm, A.; Hardt, W.; Ackermann, M. A simple screen to identify promoters conferring high levels of phenotypic noise. PLoS Genet 2008, 4, e1000307. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.; Charrier, A.; Schikora, A.; Bigeard, J.; Pateyron, S.; de Tauzia-Moreau, M.; Evrard, A.; Mithöfer, A.; Martin-Magniette, M. Salmonella enterica flagellin is recognized via FLS2 and activates PAMP-Triggered Immunity in Arabidopsis thaliana. Mol. Plant 2014, 7, 657–674. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention (CDC). Outbreak of Salmonella Infections Linked to Cut Fruit. Available online: https://www.cdc.gov/salmonella/javiana-12-19/index.html (accessed on 29 April 2020).
- Melotto, M.; Underwood, W.; He, S. Role of stomata in plant innate immunity and foliar bacterial diseases. Annu. Rev. Phytopathol. 2008, 46, 101–122. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Daudi, A.; Butt, V.; Bolwell, G. Reactive oxygen species and their role in plant defence and cell wall metabolism. Planta 2012, 236, 765–779. [Google Scholar] [CrossRef]
- Sawinski, K.; Mersmann, S.; Robatzek, S.; Böhmer, M. Guarding the green: Pathways to stomatal immunity. Mol. Plant Microbe Interact. 2013, 26, 626–632. [Google Scholar] [CrossRef]
- Haiko, J.; Westerlund-Wikstrom, B. The role of the bacterial flagellum in adhesion and virulence. Biology 2013, 2, 1242–1267. [Google Scholar] [CrossRef] [Green Version]
- Schuster, S.; Khan, S. The bacterial flagellar motor. Annu. Rev. Biophys. Biomol. Struct. 1994, 23, 509–539. [Google Scholar]
- Zhang, Y.; Nandakumar, R.; Bartelt-Hunt, S.; Snow, D.; Hodges, L.; Li, X. Quantitative proteomic analysis of the Salmonella-lettuce interaction. Microb. Biotechnol. 2014, 7, 630–637. [Google Scholar] [CrossRef]
- Cooley, M.; Miller, W.; Mandrell, R. Colonization of Arabidopsis thaliana with Salmonella enterica and enterohemorrhagic Escherichia coli O157:H7 and competition by Enterobacter asburiae. Appl. Environ. Microbiol. 2003, 69, 4915–4926. [Google Scholar] [CrossRef] [Green Version]
- Kroupitski, Y.; Golberg, D.; Belausov, E.; Pinto, R.; Swartzberg, D.; Granot, D.; Sela, S. Internalization of Salmonella enterica in leaves is induced by light and involves chemotaxis and penetration through open stomata. Appl. Environ. Microbiol. 2009, 75, 6076–6086. [Google Scholar] [CrossRef] [Green Version]
- Schikora, A.; Virlogeux-Payant, I.; Bueso, E.; Garcia, A.; Nilau, T.; Charrier, A.; Pelletier, S.; Menanteau, P.; Baccarini, M.; Velge, P.; et al. Conservation of Salmonella infection mechanisms in plants and animals. PLoS ONE 2011, 6, e24112. [Google Scholar]
- Hattermann, D.; Ries, S. Motility of Pseudomonas syringae pv. glycinea and its role in infection. Phytopathology 1989, 79, 284–289. [Google Scholar] [CrossRef] [Green Version]
- Kamoun, S.; Kado, C. Phenotypic switching affecting chemotaxis, xanthan production, and virulence in Xanthomonas campestris. Appl. Environ. Microbiol. 1990, 56, 3855–3860. [Google Scholar] [CrossRef] [Green Version]
- Saini, S.; Slauch, J.; Phillip, D.; Rao, C. Role of cross talk in regulating the dynamic expression of the flagellar Salmonella pathogenicity island 1 and type 1 fimbrial genes. J. Bacteriol. 2010, 192, 5767–5777. [Google Scholar] [CrossRef] [Green Version]
- Prüß, B.; Besemann, C.; Denton, A.; Wolfe, A. A complex transcription network controls the early stages of biofilm development by Escherichia coli. J. Bacteriol. 2006, 188, 3731. [Google Scholar] [CrossRef] [Green Version]
- Heng, F.; Altier, C.; Martin, G. Salmonella colonization activates the plant immune system and benefits from association with plant pathogenic bacteria. Environ. Microbiol. 2013, 15, 2418–2430. [Google Scholar] [CrossRef]
- Hébrard, M.; Viala, J.; Méresse, S.; Barras, F.; Aussel, L. Redundant hydrogen peroxide scavengers contribute to Salmonella virulence and oxidative stress resistance. J. Bacteriol. 2009, 191, 4605–4614. [Google Scholar] [CrossRef] [Green Version]
- Wahlig, T.; Bixler, B.; Valdés-López, O.; Mysore, K.; Wen, J.; Ané, J.; Kaspar, C. Salmonella enterica serovar Typhimurium ATCC 14028s is tolerant to plant defenses triggered by the flagellin receptor FLS2. FEMS Microbiol. Lett. 2019, 366, fny296. [Google Scholar] [CrossRef]
- Stewart, M.; Cummings, L.; Johnson, M.; Berezow, A.; Cookson, B. Regulation of phenotypic heterogeneity permits Salmonella evasion of the host caspase-1 inflammatory response. Proc. Natl. Acad. Sci. USA 2011, 108, 20742. [Google Scholar] [CrossRef] [Green Version]
- Jarvik, T.; Smillie, C.; Groisman, E.; Ochman, H. Short-term signatures of evolutionary change in the Salmonella enterica serovar Typhimurium 14028s genome. J. Bacteriol. 2010, 192, 560. [Google Scholar] [CrossRef] [Green Version]
- Aime, S.; Alabouvette, C.; Steinberg, C.; Olivain, C. The endophytic strain Fusarium oxysporum Fo47: A good candidate for priming the defense responses in tomato roots. Mol. Plant Microbe Interact. 2013, 26, 918–926. [Google Scholar] [CrossRef] [Green Version]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zarkani, A.A.; López-Pagán, N.; Grimm, M.; Sánchez-Romero, M.A.; Ruiz-Albert, J.; Beuzón, C.R.; Schikora, A. Salmonella Heterogeneously Expresses Flagellin during Colonization of Plants. Microorganisms 2020, 8, 815. https://doi.org/10.3390/microorganisms8060815
Zarkani AA, López-Pagán N, Grimm M, Sánchez-Romero MA, Ruiz-Albert J, Beuzón CR, Schikora A. Salmonella Heterogeneously Expresses Flagellin during Colonization of Plants. Microorganisms. 2020; 8(6):815. https://doi.org/10.3390/microorganisms8060815
Chicago/Turabian StyleZarkani, Azhar A., Nieves López-Pagán, Maja Grimm, María Antonia Sánchez-Romero, Javier Ruiz-Albert, Carmen R. Beuzón, and Adam Schikora. 2020. "Salmonella Heterogeneously Expresses Flagellin during Colonization of Plants" Microorganisms 8, no. 6: 815. https://doi.org/10.3390/microorganisms8060815