Differentiation of Leishmania (L.) infantum, Leishmania (L.) amazonensis and Leishmania (L.) mexicana Using Sequential qPCR Assays and High-Resolution Melt Analysis


 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Leishmania Strains, Clinical Samples and DNA Extraction
2.2. ITS1-PCR RFLP
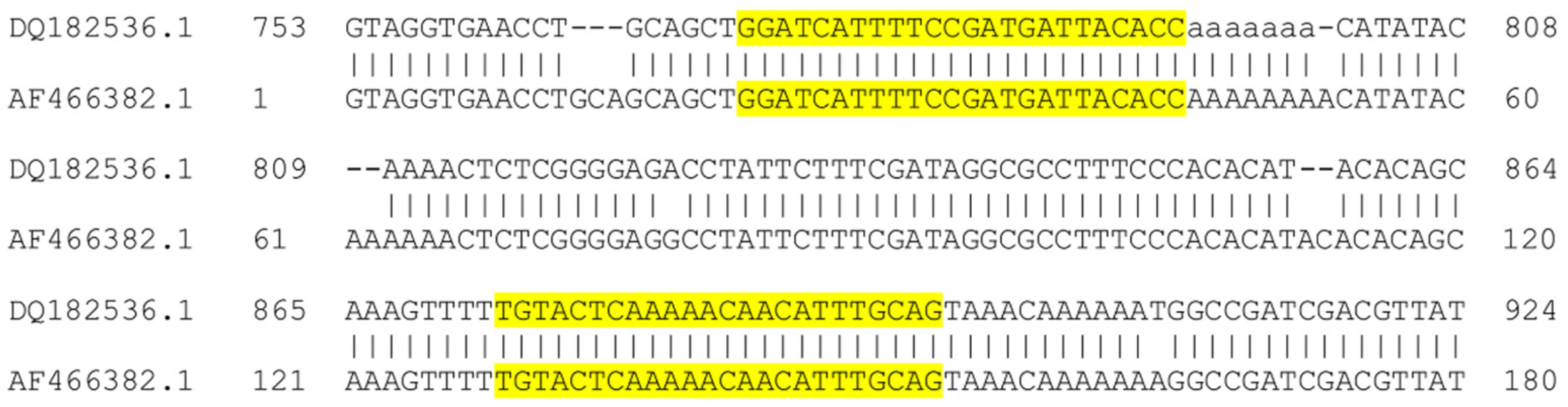
2.3. DNA Sequencing and Phylogenetic Analysis
2.4. qPCR Assays
2.5. High-Resolution Melt (HRM) Analysis
2.6. Ethics Approval
2.7. Statistical Analysis
3. Results
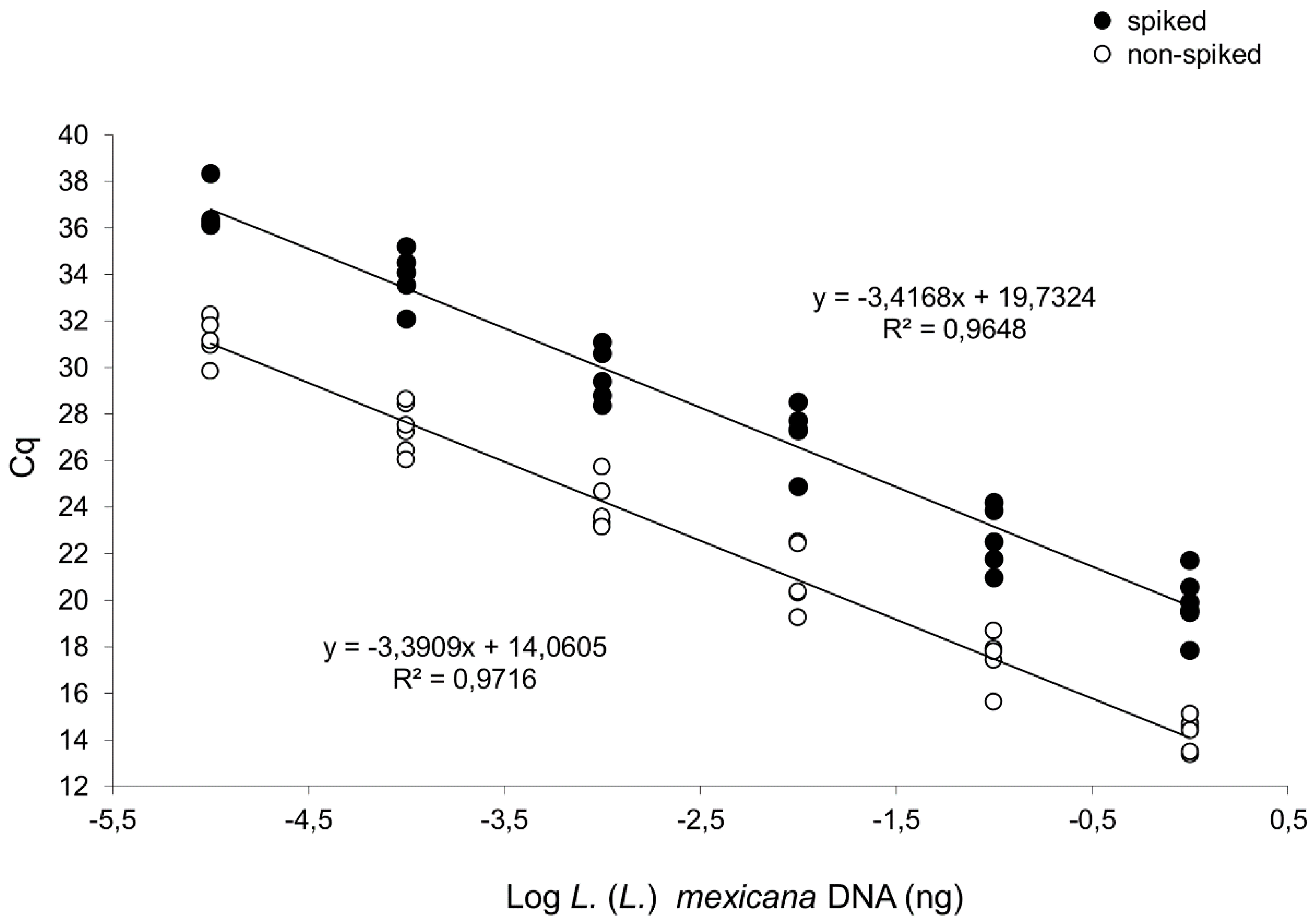
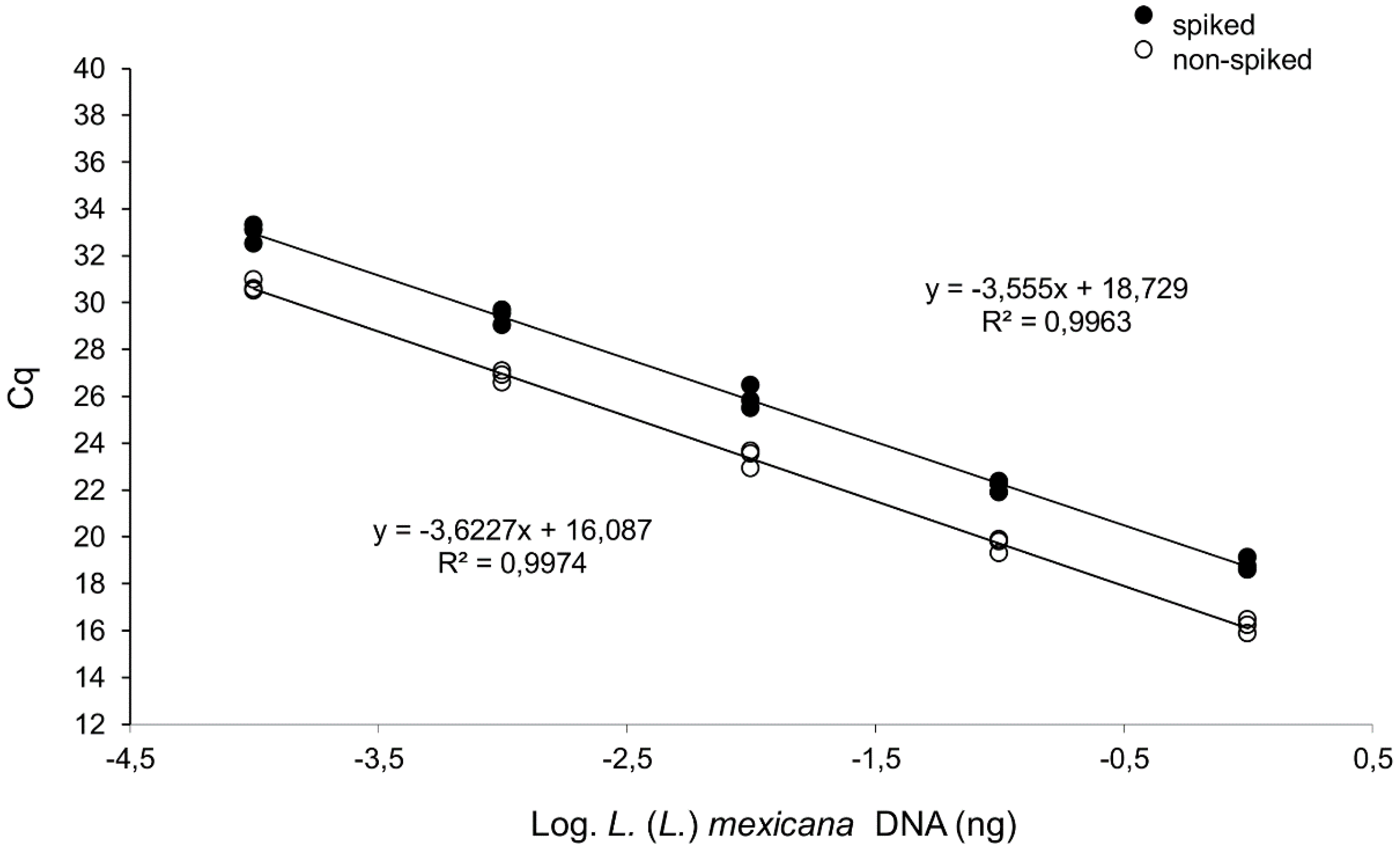
3.1. Both L. (L.) mexicana and L. (L.) amazonensis Can be Distinguished from L. (L.) infantum Exploiting A Differential qPCR Targeting Minicircle kDNA
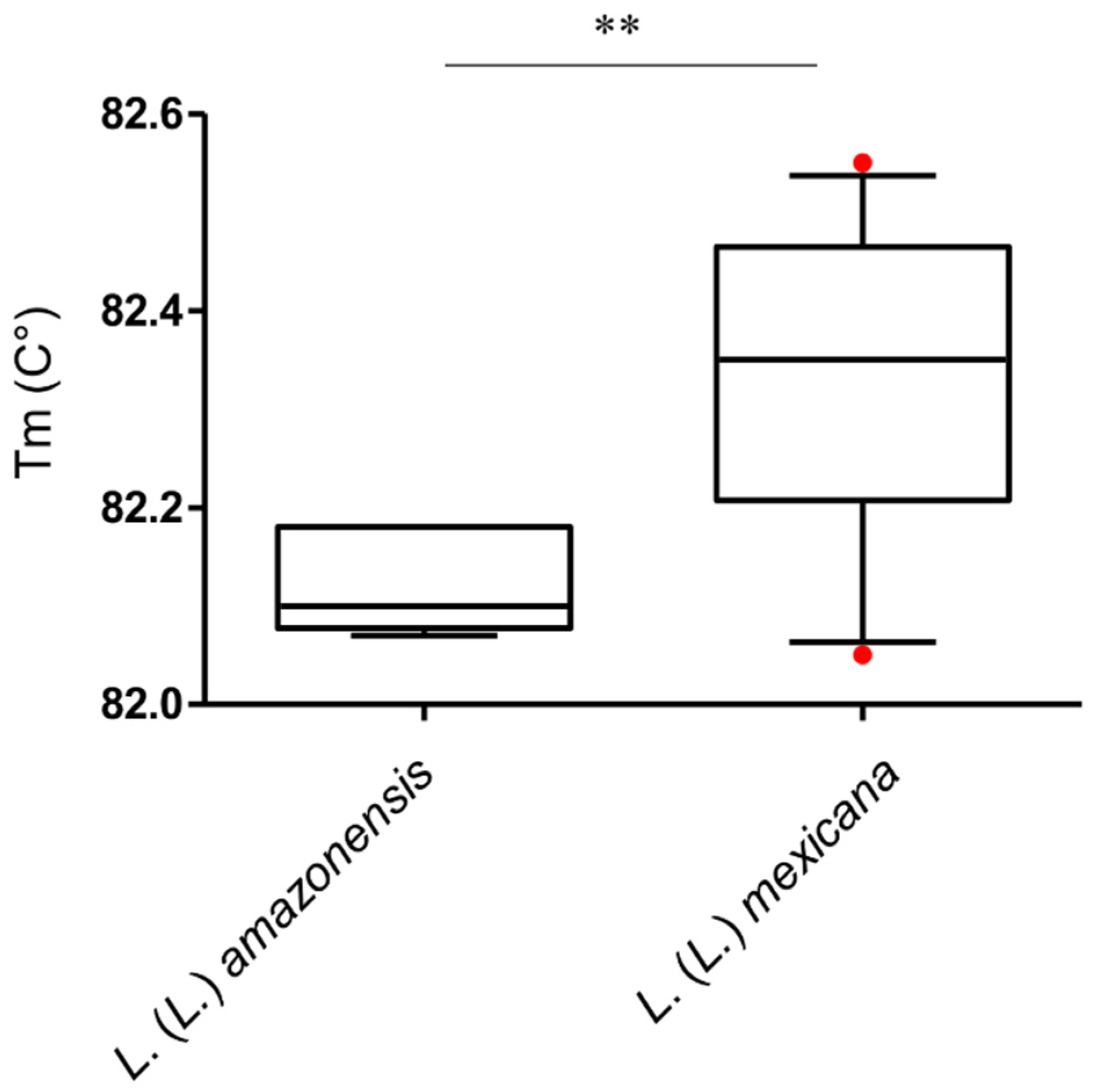
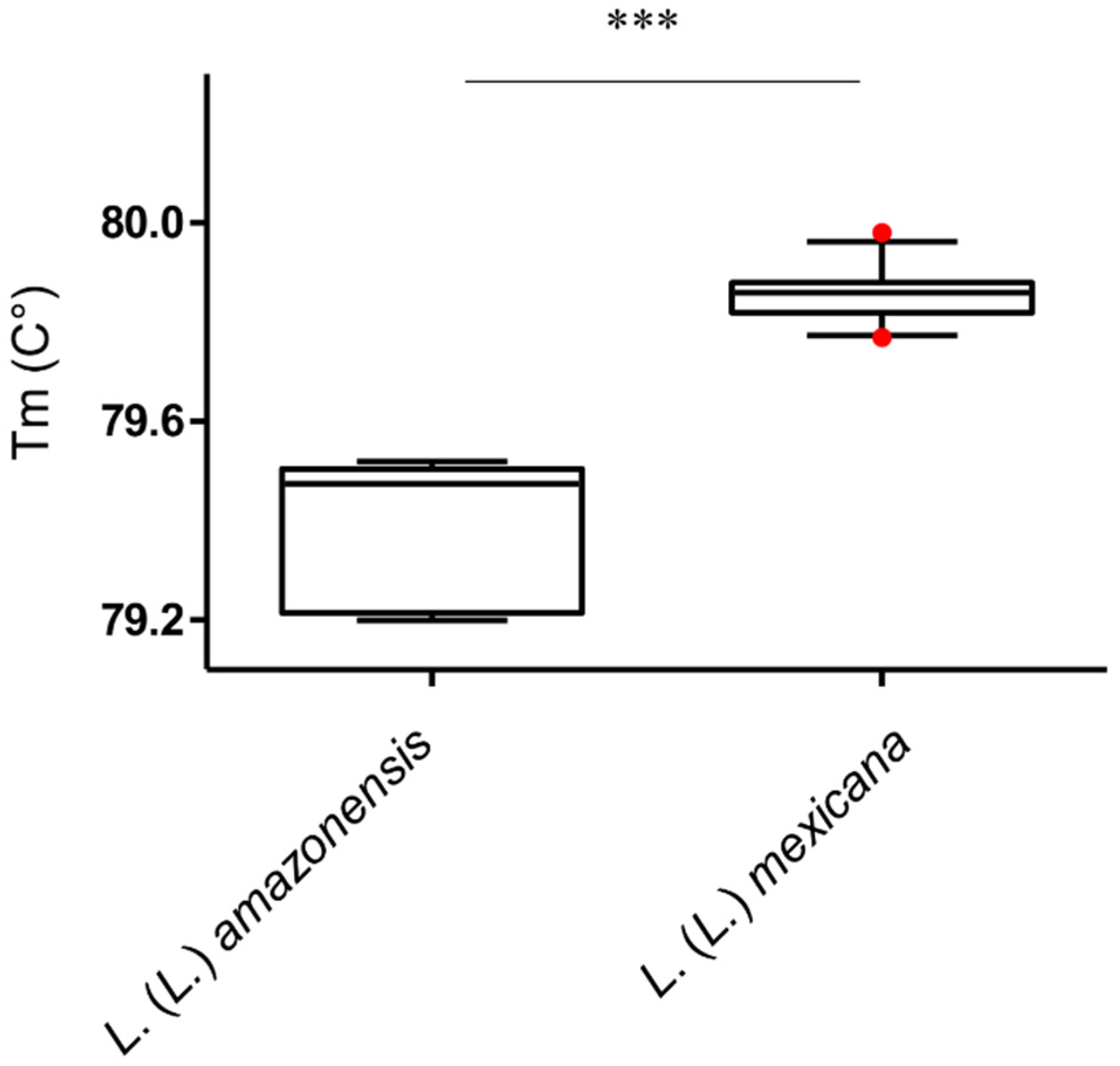
3.2. L. (L.) amazonensis Can be Differentiated from L. (L.) mexicana by qPCR-ITS1 HRM Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; den Boer, M. Leishmaniasis Worldwide and Global Estimates of Its Incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef] [PubMed]
- Hashiguchi, Y.; Velez, L.N.; Villegas, N.V.; Mimori, T.; Gomez, E.A.L.; Kato, H. Leishmaniases in Ecuador: Comprehensive review and current status. Acta Trop. 2017, 166, 299–315. [Google Scholar] [CrossRef] [PubMed]
- Goto, H.; Lindoso, J.A. Current diagnosis and treatment of cutaneous and mucocutaneous leishmaniasis. Expert Rev. Anti Infect. Ther. 2010, 8, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Akhoundi, M.; Downing, T.; Votýpka, J.; Kuhls, K.; Lukeš, J.; Cannet, A.; Ravel, C.; Marty, P.; Delaunay, P.; Kasbari, M.; et al. Leishmania infections: Molecular targets and diagnosis. Mol. Asp. Med. 2017, 57, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Recalde, O.D.S.; Brunelli, J.P.; Rolon, M.S.; De Arias, A.R.; Aldama, O.; Gómez, C.V. First molecular report of Leishmania (Leishmania) amazonensis and Leishmania (Viannia) guyanensis in paraguayan inhabitants using high-resolution melt-PCR. Am. J. Trop. Med. Hyg. 2019, 101, 780–788. [Google Scholar] [CrossRef]
- Galluzzi, L.; Ceccarelli, M.; Diotallevi, A.; Menotta, M.; Magnani, M. Real-time PCR applications for diagnosis of leishmaniasis. Parasites Vectors 2018, 11, 273. [Google Scholar] [CrossRef]
- Medkour, H.; Varloud, M.; Davoust, B.; Mediannikov, O. New Molecular Approach for the Detection of Kinetoplastida Parasites of Medical and Veterinary Interest. Microorganisms 2020, 8, 356. [Google Scholar] [CrossRef] [Green Version]
- Schonian, G.; Kuhls, K.; Mauricio, I.L. Molecular approaches for a better understanding of the epidemiology and population genetics of Leishmania. Parasitology 2011, 138, 405–425. [Google Scholar] [CrossRef] [Green Version]
- Jensen, R.E.; Englund, P.T. Network News: The Replication of Kinetoplast DNA. Annu. Rev. Microbiol. 2012, 66, 473–491. [Google Scholar] [CrossRef]
- Brewster, S.; Barker, D. Analysis of minicircle classes in Leishmania (Viannia) species. Trans. R. Soc. Trop. Med. Hyg. 2002, 96, S55–S63. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Lee, S.-T.; Chang, K.-P. Transkinetoplastidy-A Novel Phenomenon Involving Bulk Alterations of Mitochondrion-Kinetoplast DNA of a Trypanosomatid Protozoan 1, 2. J. Protozool. 1992, 39, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Simpson, L. The genomic organization of guide RNA genes in kinetoplastid protozoa: Several conundrums and their solutions. Mol. Biochem. Parasitol. 1997, 86, 133–141. [Google Scholar] [CrossRef]
- Kocher, A.; Valière, S.; Bañuls, A.-L.; Murienne, J. High-throughput sequencing of kDNA amplicons for the analysis of Leishmania minicircles and identification of Neotropical species. Parasitology 2017, 145, 1–8. [Google Scholar]
- Mary, C.; Faraut, F.; Lascombe, L.; Dumon, H. Quantification of Leishmania infantum DNA by a real-time PCR assay with high sensitivity. J. Clin. Microbiol. 2004, 42, 5249–5255. [Google Scholar] [CrossRef] [Green Version]
- Pita-Pereira, D.; Lins, R.; Oliveira, M.P.; Lima, R.B.; Pereira, B.A.S.; Moreira, O.C.; Brazil, R.P.; Britto, C. SYBR Green-based real-time PCR targeting kinetoplast DNA can be used to discriminate between the main etiologic agents of Brazilian cutaneous and visceral leishmaniases. Parasites Vectors 2012, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, M.; Galluzzi, L.; Migliazzo, A.; Magnani, M. Detection and Characterization of Leishmania (Leishmania) and Leishmania (Viannia) by SYBR Green-Based Real-Time PCR and High Resolution Melt Analysis Targeting Kinetoplast Minicircle DNA. PLoS ONE 2014, 9, e88845. [Google Scholar] [CrossRef] [Green Version]
- Losada-Barragán, M.; Cavalcanti, A.; Umaña-Pérez, A.; Porrozzi, R.; Cuervo-Escobar, S.; Vallejo, A.F.; Sánchez-Gómez, M.; Cuervo, P. Detection and quantification of Leishmania infantum in naturally and experimentally infected animal samples. Vet. Parasitol. 2016, 226, 57–64. [Google Scholar] [CrossRef]
- Ceccarelli, M.; Galluzzi, L.; Diotallevi, A.; Andreoni, F.; Fowler, H.; Petersen, C.; Vitale, F.; Magnani, M. The use of kDNA minicircle subclass relative abundance to differentiate between Leishmania (L.) infantum and Leishmania (L.) amazonensis. Parasites Vectors 2017, 10, 239. [Google Scholar] [CrossRef]
- Diotallevi, A.; Buffi, G.; Ceccarelli, M.; Neitzke-Abreu, H.C.; Gnutzmann, L.V.; da Costa Lima, M.S.; Di Domenico, A.; De Santi, M.; Magnani, M.; Galluzzi, L. Real-time PCR to differentiate among Leishmania (Viannia) subgenus, Leishmania (Leishmania) infantum and Leishmania (Leishmania) amazonensis: Application on Brazilian clinical samples. Acta Trop. 2020, 201, 105178. [Google Scholar] [CrossRef]
- Diotallevi, A.; Buffi, G.; Ceccarelli, M.; Di Domenico, A.; De Santi, M.; Magnani, M.; Galluzzi, L.; Neitzke-Abreu, H.C.; Gnutzmann, L.V.; da Costa Lima Junior, M.S. Data on the differentiation among Leishmania (Viannia) spp., Leishmania (Leishmania) infantum and Leishmania (Leishmania) amazonensis in Brazilian clinical samples using real-time PCR. Data Brief 2020, 28, 104914. [Google Scholar] [CrossRef]
- Tschoeke, D.A.; Nunes, G.L.; Jardim, R.; Lima, J.; Dumaresq, A.S.R.; Gomes, M.R.; De Mattos Pereira, L.; Loureiro, D.R.; Stoco, P.H.; De Matos Gliedes, H.L.; et al. The Comparative Genomics and Phylogenomics of Leishmania Amazonensis Parasite. Evol. Bioinform. 2014, 10, EBO.S13759. [Google Scholar] [CrossRef] [PubMed]
- Monroy-Ostria, A.; Nasereddin, A.; Monteon, V.M.; Guzmán-Bracho, C.; Jaffe, C.L. ITS1 PCR-RFLP Diagnosis and Characterization of Leishmania in Clinical Samples and Strains from Cases of Human Cutaneous Leishmaniasis in States of the Mexican Southeast. Interdiscip. Perspect. Infect. Dis. 2014, 2014, 607287. [Google Scholar] [CrossRef]
- Marco, J.D.; Barroso, P.A.; Locatelli, F.M.; Cajal, S.P.; Hoyos, C.L.; Nevot, M.C.; Lauthier, J.J.; Tomasini, N.; Juarez, M.; Estévez, J.O.; et al. Multilocus sequence typing approach for a broader range of species of Leishmania genus: Describing parasite diversity in Argentina. Infect. Genet. Evol. 2015, 30, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, M.; Diotallevi, A.; Andreoni, F.; Vitale, F.; Galluzzi, L.; Magnani, M. Exploiting genetic polymorphisms in metabolic enzymes for rapid screening of Leishmania infantum genotypes. Parasites Vectors 2018, 11, 572. [Google Scholar] [CrossRef] [PubMed]
- Quaresma, P.F.; Murta, S.M.F.; de Ferreira, E.C.; da Rocha-Lima, A.C.V.M.; Xavier, A.A.P.; Gontijo, C.M.F. Molecular diagnosis of canine visceral leishmaniasis: Identification of Leishmania species by PCR-RFLP and quantification of parasite DNA by real-time PCR. Acta Trop. 2009, 111, 289–294. [Google Scholar] [CrossRef]
- Nicolas, L.; Prina, E.; Lang, T.; Milon, G. Real-Time PCR for Detection and Quantitation of Leishmania in Mouse Tissues. J. Clin. Microbiol. 2002, 40, 1666–1669. [Google Scholar] [CrossRef] [Green Version]
- Hernández, C.; Alvarez, C.; González, C.; Ayala, M.S.; León, C.M.; Ramírez, J.D. Identification of Six New World Leishmania species through the implementation of a High-Resolution Melting (HRM) genotyping assay. Parasites Vectors 2014, 7, 501. [Google Scholar] [CrossRef]
- Zampieri, R.A.; Laranjeira-Silva, M.F.; Muxel, S.M.; Stocco de Lima, A.C.; Shaw, J.J.; Floeter-Winter, L.M. High Resolution Melting Analysis Targeting hsp70 as a Fast and Efficient Method for the Discrimination of Leishmania Species. PLoS Negl. Trop. Dis. 2016, 10, e0004485. [Google Scholar] [CrossRef] [Green Version]
- Fraga, J.; Montalvo, A.M.; De Doncker, S.; Dujardin, J.-C.; Van der Auwera, G. Phylogeny of Leishmania species based on the heat-shock protein 70 gene. Infect. Genet. Evol. 2010, 10, 238–245. [Google Scholar] [CrossRef]
- Van der Auwera, G.; Ravel, C.; Verweij, J.J.; Bart, A.; Schonian, G.; Felger, I. Evaluation of Four Single-Locus Markers for Leishmania Species Discrimination by Sequencing. J. Clin. Microbiol. 2014, 52, 1098–1104. [Google Scholar] [CrossRef] [Green Version]
- Schönian, G.; Nasereddin, A.; Dinse, N.; Schweynoch, C.; Schallig, H.D.F.; Presber, W.; Jaffe, C.L. PCR diagnosis and characterization of Leishmania in local and imported clinical samples. Diagn. Microbiol. Infect. Dis. 2003, 47, 349–358. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain/Isolate |
|---|---|
| L. (L.) infantum | MHOM/TN/80/IPT1 |
| L. (L.) infantum | MHOM/IT/86/ISS218 |
| L. (L.) infantum | MHOM/FR/78/LEM75 |
| L. (L.) amazonensis | MHOM/BR/00/LTB0016 |
| L. (L.) amazonensis | IFLA/BR/67/PH8 |
| L. (L.) amazonensis | Clinical isolate |
| L. (V.) braziliensis | MHOM/BR/75/M2904 |
| L. (L.) mexicana | MHOM/MX/2011/Lacandona |
| L. (L.) mexicana | Clinical isolate 2 |
| L. (L.) mexicana | Clinical isolate 3 |
| L. (L.) mexicana | Clinical isolate 5 |
| L. (L.) mexicana | Clinical isolate 14 |
| L. (L.) mexicana | Clinical isolate 17 |
| Sample | Species Identification (ITS1-PCR RFLP) | Species Identification (ALAT sequencing) | CL Form |
|---|---|---|---|
| Px1 | L. (L.) mexicana | n.a. 1 | LCL |
| Px2 | L. (L.) mexicana | L. (L.) mexicana | LCL |
| Px3 | L. (L.) mexicana | L. (L.) mexicana | LCL |
| Px4 | L. (L.) mexicana | n.a. 1 | LCL |
| Px5 | L. (L.) mexicana | n.a. 1 | DCL |
| Px7 | L. (L.) mexicana | n.a. 1 | LCL |
| Px9 | L. (L.) mexicana | L. (L.) mexicana | DCL |
| Px10 | L. (L.) mexicana | L. (L.) mexicana | LCL |
| PxGSF | L. (L.) mexicana | n.a. 1 | LCL |
| PxCMU | n.a. 1 | n.a. 1 | n.a. 1 |
| PxJLC | L. (L.) mexicana | L. (L.) mexicana | LCL |
| Leishmania Species, Strain/Isolate | DNA Template (ng) | qPCR-ML (Cq ± SD) | qPCR-ama (Cq ± SD) |
|---|---|---|---|
| L. (L.)mexicana MHOM/MX/2011/Lacandona | 1.0 | 31.61 ± 2.03 | 14.25 ± 0.69 |
| L. (L.)mexicana MHOM/MX/2011/Lacandona | 1.0 × 10−1 | 33.43 ± 2.09 | 17.48 ± 1.13 |
| L. (L.)mexicana MHOM/MX/2011/Lacandona | 1.0 × 10−2 | 37.53 ± 1.27 | 20.68 ± 1.46 |
| L. (L.)mexicana MHOM/MX/2011/Lacandona | 1.0 × 10−3 | n.d. 1 | 24.07 ± 1.09 |
| L. (L.)mexicana MHOM/MX/2011/Lacandona | 1.0 × 10−4 | n.d. 1 | 27.37 ± 1.05 |
| L. (L.)mexicana MHOM/MX/2011/Lacandona | 1.0 × 10−5 | n.d.1 | 31.36 ± 0.93 |
| L. (L.)mexicana Isolate 2 | 1.0 | 33.19 ± 1.34 | 16.78 ± 0.06 |
| L. (L.)mexicana Isolate 3 | 1.0 | 33.62 ± 2.14 | 18.62 ± 1.12 |
| L. (L.)mexicana Isolate 5 | 1.0 | 38.19 ± 1.01 | 20.14 ± 0.43 |
| L. (L.)mexicana Isolate 14 | 1.0 | 34.59 ± 0.51 | 16.54 ± 0.17 |
| L. (L.)mexicana Isolate 17 | 1.0 | 35.94 ± 1.20 | 19.15 ± 1.00 |
| L. (L.)amazonensis MHOM/BR/00/LTB0016 | 1.0 × 10−1 | 33.95 ± 0.34 | 21.1 ±1.02 |
| L. (L.)infantum MHOM/FR/78/LEM75 | 1.0 | 14.42 ± 0.75 | 28.02 ± 0.98 |
| Sample ID | qPCR-ML (Cq ± SD) | qPCR-ama (Cq ± SD) |
|---|---|---|
| Px1 | n.d. 1 | 27.71 ± 0.08 |
| Px2 | n.d. 1 | 25.79 ± 0.55 |
| Px3 | n.d. 1 | 24.57 ± 0.58 |
| Px4 | 36.21 ± 1.99 | 29.59 ± 0.80 |
| Px5 | n.d. 1 | 28.99 ± 1.57 |
| Px7 | 35.31 ± 1.47 | 28.04 ± 0.25 |
| Px9 | n.d. 1 | 28.35 ± 1.93 |
| Px10 | n.d. 1 | 24.92 ± 0.87 |
| PxGSF | 36.38 ± 1.70 | 29.49 ± 0.53 |
| PxCMU | 36.53 ± 1.36 | 31.48 ± 0.36 |
| PxJLC | n.d. 1 | 34.87 ± 1.93 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceccarelli, M.; Diotallevi, A.; Buffi, G.; De Santi, M.; Fernández-Figueroa, E.A.; Rangel-Escareño, C.; Muñoz-Montero, S.A.; Becker, I.; Magnani, M.; Galluzzi, L. Differentiation of Leishmania (L.) infantum, Leishmania (L.) amazonensis and Leishmania (L.) mexicana Using Sequential qPCR Assays and High-Resolution Melt Analysis. Microorganisms 2020, 8, 818. https://doi.org/10.3390/microorganisms8060818
Ceccarelli M, Diotallevi A, Buffi G, De Santi M, Fernández-Figueroa EA, Rangel-Escareño C, Muñoz-Montero SA, Becker I, Magnani M, Galluzzi L. Differentiation of Leishmania (L.) infantum, Leishmania (L.) amazonensis and Leishmania (L.) mexicana Using Sequential qPCR Assays and High-Resolution Melt Analysis. Microorganisms. 2020; 8(6):818. https://doi.org/10.3390/microorganisms8060818
Chicago/Turabian StyleCeccarelli, Marcello, Aurora Diotallevi, Gloria Buffi, Mauro De Santi, Edith A. Fernández-Figueroa, Claudia Rangel-Escareño, Said A. Muñoz-Montero, Ingeborg Becker, Mauro Magnani, and Luca Galluzzi. 2020. "Differentiation of Leishmania (L.) infantum, Leishmania (L.) amazonensis and Leishmania (L.) mexicana Using Sequential qPCR Assays and High-Resolution Melt Analysis" Microorganisms 8, no. 6: 818. https://doi.org/10.3390/microorganisms8060818