Novel Intact Polar and Core Lipid Compositions in the Pyrococcus Model Species, P. furiosus and P. yayanosii, Reveal the Largest Lipid Diversity Amongst Thermococcales

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Growth Conditions
2.2. IPL Extraction and HPLC-ESI-MS Analysis
2.3. CL Preparation and HPLC-APCI-MS Analysis
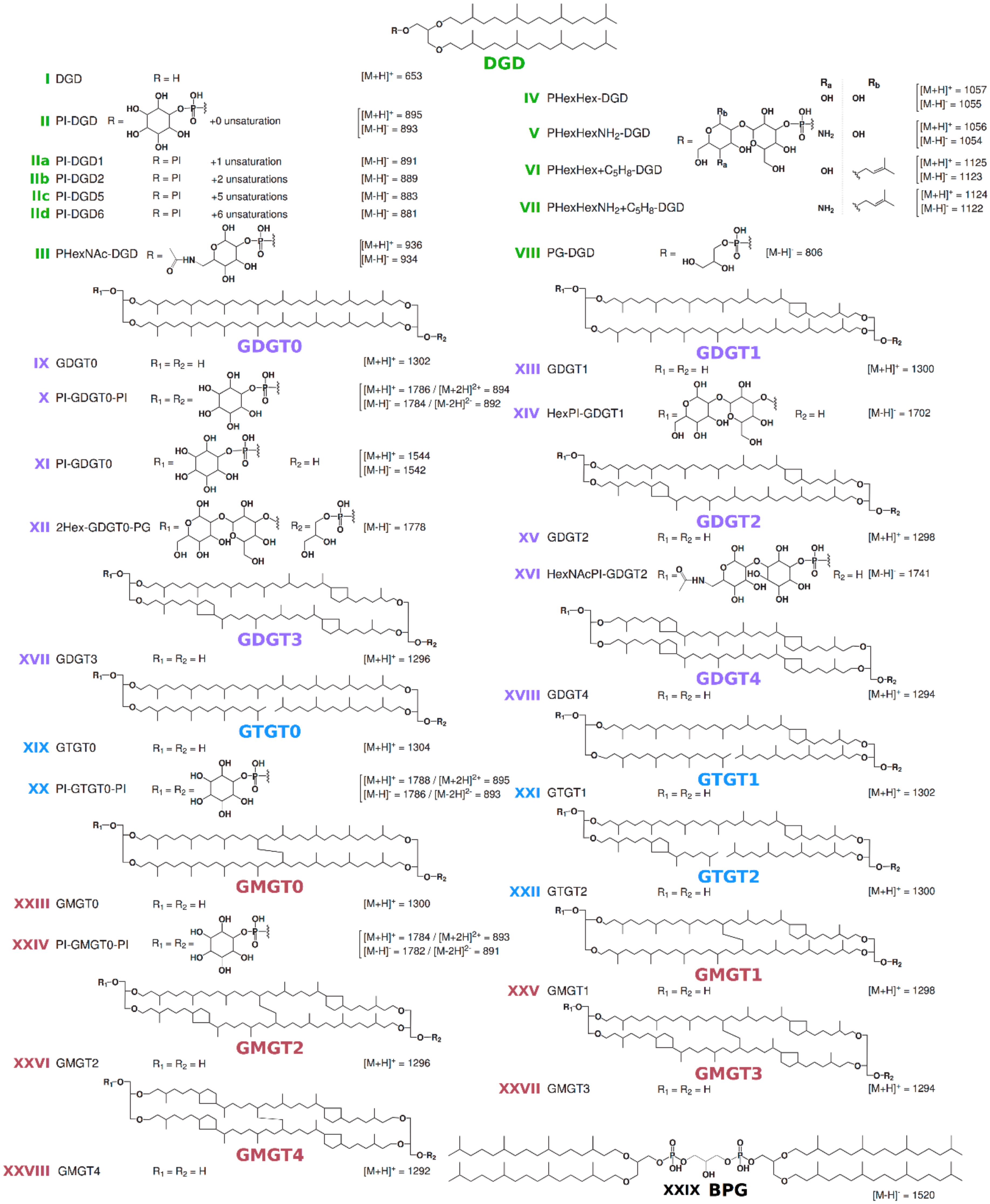
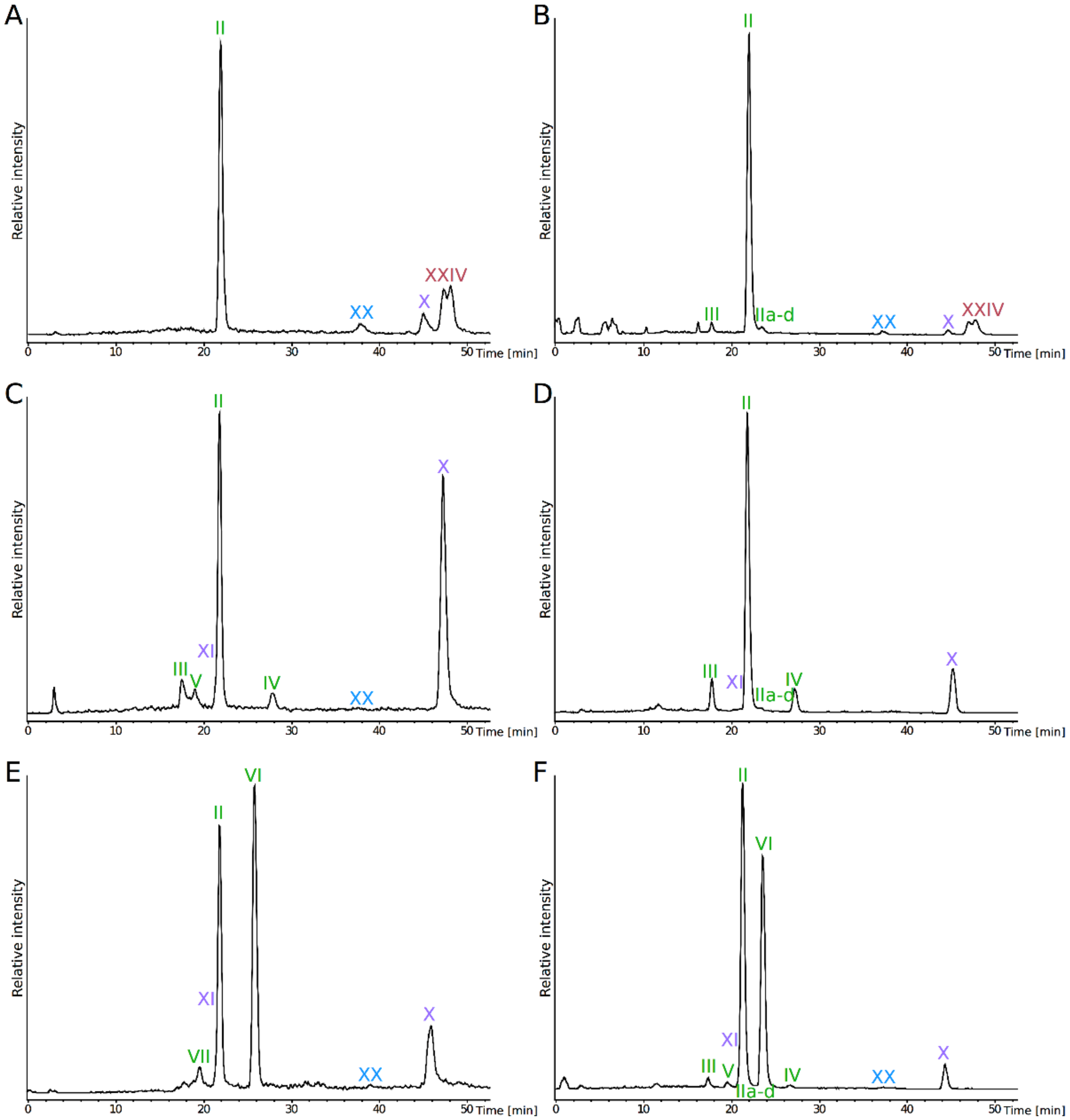
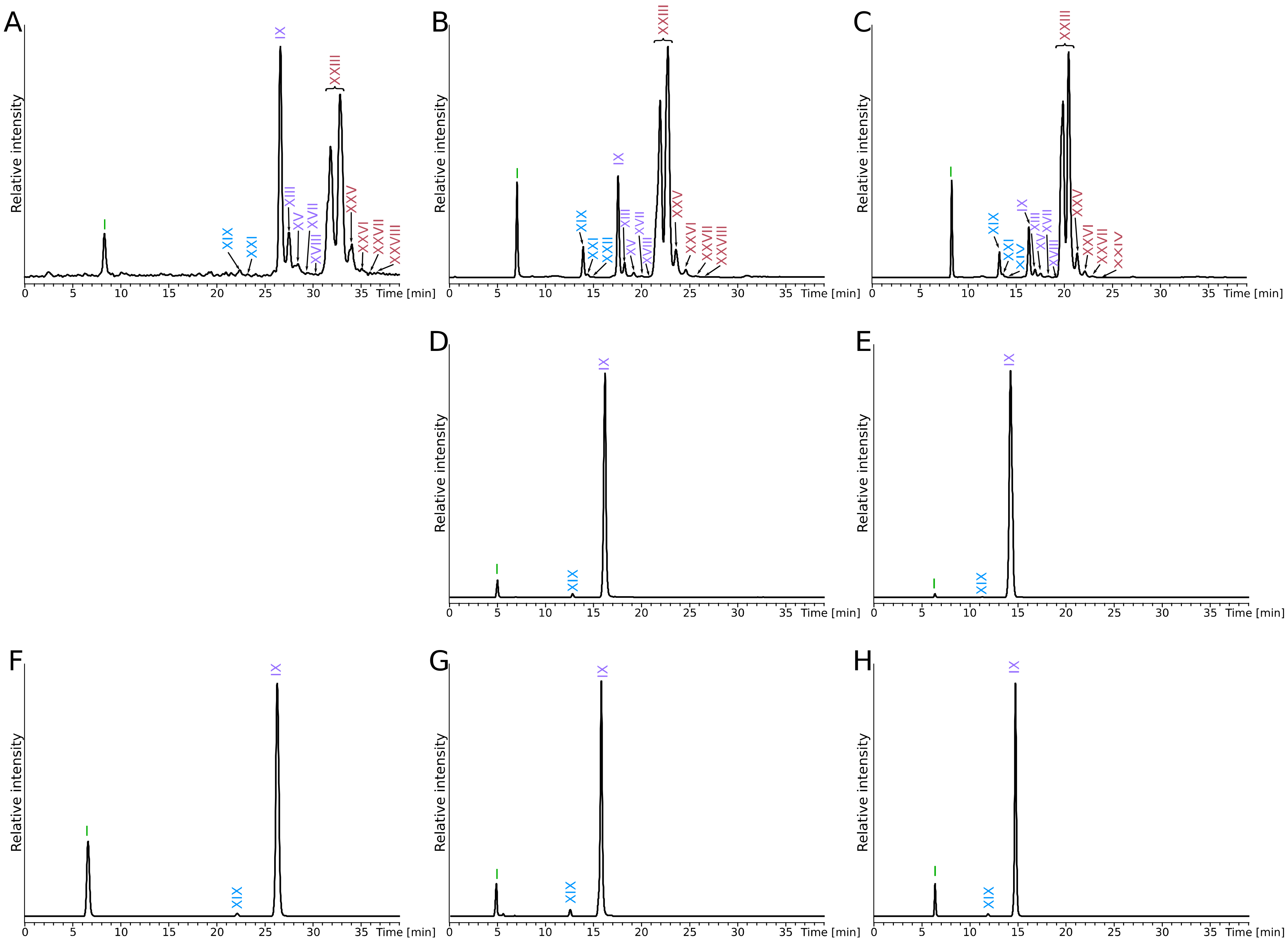
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| IPL | Intact polar lipid(s) |
| CL | Core lipid(s) |
| MeOH | Methanol |
| DCM | Dichloromethane |
| HPLC | High-performance liquid chromatography |
| MS | Mass spectrometry |
| ESI | Electrospray ionization |
| APCI | Atmospheric-pressure chemical ionization |
| MGD | Monoalkyl glycerol diethers |
| DGD | Dialkyl glycerol diethers |
| GDGT | Glycerol dialkyl glycerol tetraethers |
| GTGT | Glycerol trialkyl glycerol tetraethers |
| GMGT | Glycerol monoalkyl glycerol tetraethers |
| B&D | Bligh and Dyer |
| PI | Phosphatidyl inositol |
| PHexNAc | Phosphatidyl N-acetylhexosamine |
| PHexHex | Glycosylated phosphatidyl hexose |
| PG | Phosphatidyl glycerol |
| FAB | Fast atom bombardment |
| MALDI-TOF | Matrix-assisted laser desorption ionization-time of flight |
References
- Baba, T.; Toshima, Y.; Minamikawa, H.; Hato, M.; Suzuki, K.; Kamo, N. Formation and characterization of planar lipid bilayer membranes from synthetic phytanyl-chained glycolipids. BBA Biomembr. 1999, 1421, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, H.; Chong, P.L.-G. Low permeability of liposomal membranes composed of bipolar tetraether lipids from thermoacidophilic archaebacterium Sulfolobus acidocaldarius. Biochemestry 1998, 37, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Comita, P.B.; Gagosian, R.B.; Pang, H.; Costello, C.E. Structural elucidation of a unique macrocyclic membrane lipid from a new, extremely thermophilic, deep-sea hydrothermal vent archaebacterium, Methanococcus jannaschii. J. Biol. 1984, 259, 15234–15241. [Google Scholar]
- Knappy, C.S.; Nunn, C.E.M.; Morgan, H.W.; Keely, B. The major lipid cores of the archaeon Ignisphaera aggregans: Implications for the phylogeny and biosynthesis of glycerol monoalkyl glycerol tetraether isoprenoid lipids. Extremophiles 2011, 15, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Nichols, D.S.; Miller, M.R.; Davies, N.W.; Goodchild, A.; Raftery, M.; Cavicchioli, R. Cold adaptation in the Antarctic archaeon Methanococcoides burtonii involves membrane lipid unsaturation. J. Bacteriol. Res. 2004, 186, 8508–8515. [Google Scholar] [CrossRef] [Green Version]
- Gambacorta, A.; Trincone, A.; Nicolaus, B.; Lama, L.; De Rosa, M. Unique features of lipids of Archaea. Syst. Appl. Microbiol. 1993, 16, 518–527. [Google Scholar] [CrossRef]
- Becker, K.W.; Elling, F.J.; Yoshinaga, M.Y.; Söllinger, A.; Urich, T.; Hinrichs, K.-U. Unusual butane- and pentanetriol-based tetraether lipids in Methanomassiliicoccus luminyensis, a representative of the seventh order of methanogens. Appl. Environ. Microbiol. 2016, 82, 4505–4516. [Google Scholar] [CrossRef] [Green Version]
- Jahn, U.; Summons, R.; Sturt, H.; Grosjean, E.; Huber, H. Composition of the lipids of Nanoarchaeum equitans and their origin from its host Ignicoccus sp. strain KIN4/I. Arch. Microbiol. 2004, 182, 404–413. [Google Scholar] [CrossRef]
- Koga, Y.; Akagawa-Matsushita, M.; Ohga, M.; Nishihara, M. Taxonomic significance of the distribution of component parts of polar ether lipids in methanogens. Syst. Appl. Microbiol. 1993, 16, 342–351. [Google Scholar] [CrossRef]
- Huguet, C.; Martens-Habbena, W.; Urakawa, H.; Stahl, D.A.; Ingalls, A.E. Comparison of extraction methods for quantitative analysis of core and intact polar glycerol dialkyl glycerol tetraethers (GDGTs) in environmental samples. Limnol. Oceanogr. Meth. 2010, 8, 127–145. [Google Scholar] [CrossRef]
- Cario, A.; Grossi, V.; Schaeffer, P.; Oger, P.M. Membrane homeoviscous adaptation in the piezo-hyperthermophilic archaeon Thermococcus barophilus. Front. Microbiol. 2015, 6, 1152–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishihara, M.; Koga, Y. Extraction and Composition of polar lipids from the archaebacterium, Methanobacterium thermoautotrophicum: Effective extraction of tetraether lipids by an acidified solvent. J. Biochem. 1987, 101, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Yoshinaga, M.Y.; Gagen, E.J.; Wörmer, L.; Broda, N.K.; Meador, T.B.; Wendt, J.; Thomm, M.; Hinrichs, K.U. Methanothermobacter thermautotrophicus modulates its membrane lipids in response to hydrogen and nutrient availability. Front. Microbiol. 2015, 6, 5–14. [Google Scholar] [PubMed] [Green Version]
- Marteinsson, V.T.; Birrien, J.L.; Reysenbach, A.L.; Vernet, M.; Marie, D.; Gambacorta, A.; Messner, P.; Sleytr, U.B.; Prieur, D. Thermococcus barophilus sp. nov., a new barophilic and hyperthermophilic archaeon isolated under high hydrostatic pressure from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Micr. 1999, 49, 351–359. [Google Scholar]
- Fiala, G.; Stetter, K.O. Pyrococcus furiosus sp. nov. represents a novel genus of marine heterotrophic archaebacteria growing optimally at 100 C. Arch. Microbiol. 1986, 145, 56–61. [Google Scholar]
- Farkas, J.; Stirrett, K.; Lipscomb, G.L.; Nixon, W.; Scott, R.; Adams, M.W.W.; Westpheling, J. Recombinogenic properties of Pyrococcus furiosus strain COM1 enable rapid selection of targeted mutants. Appl. Environ. Microbiol. 2012, 78, 4669–4676. [Google Scholar] [CrossRef] [Green Version]
- Lipscomb, G.L.; Stirrett, K.; Schut, G.J.; Yang, F.; Jenney, F.E.; Scott, R.A.; Adams, M.W.; Westpheling, J. Natural competence in the hyperthermophilic archaeon Pyrococcus furiosus facilitates genetic manipulation: Construction of markerless deletions of genes encoding the two cytoplasmic hydrogenases. Appl. Environ. Microbiol. 2011, 77, 2232–2238. [Google Scholar] [CrossRef] [Green Version]
- Zillig, W.; Holz, I.; Klenk, H.-P.; Trent, J.; Wunderl, S.; Janekovic, D.; Imsel, E.; Haas, B. Pyrococcus woesei, sp. nov., an ultra-thermophilic marine archaebacterium, representing a novel order, Thermococcales. Syst. Appl. Microbiol. 1987, 9, 62–70. [Google Scholar] [CrossRef]
- Kanoksilapatham, W.; Gonzalez, J.M.; Maeder, D.L.; DiRuggiero, J.; Robb, F. A proposal to rename the hyperthermophile Pyrococcus woesei as Pyrococcus furiosus subsp. woesei. Archaea 2004, 1, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Lanzotti, V.; Trincone, A.; Nicolaus, B.; Zillig, W.; De Rosa, M.; Gambacorta, A. Complex lipids of Pyrococcus and AN1, thermophilic members of archaebcteria belonging to Thermococcales. Biochim. Biophys. Acta Lipids Lipid Metab. 1989, 1004, 44–48. [Google Scholar] [CrossRef]
- Sprott, G.D.; Agnew, B.J.; Patel, G.B. Structural features of ether lipids in the archaeobacterial thermophiles Pyrococcus furiosus, Methanopyrus kandleri, Methanothermus fervidus, and Sulfolobus acidocaldarius. Can. J. Microbiol. 1997, 43, 467–476. [Google Scholar] [CrossRef]
- Lobasso, S.; Lopalco, P.; Angelini, R.; Vitale, R.; Huber, H.; Müller, V.; Corcelli, A. Coupled TLC and MALDI-TOF/MS analyses of the lipid extract of the hyperthermophilic archaeon Pyrococcus furiosus. Archaea 2012, 2012, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reysenbach, A.-L.; Godfroy, A.; Meunier, J.-R.; Crump, B.C.; Baross, J.; Marteinsson, V.; Barbier, G.; Pace, N.; Prieur, D. Pyrococcus abyssi sp. nov., a new hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent. Arch. Microbiol. 1993, 160, 338–349. [Google Scholar] [CrossRef]
- Sugai, A.; Masuchi, Y.; Uda, I.; Itoh, T.; Itoh, Y.H. Core lipids of hyperthermophilic archaeon, Pyrococcus horikoshii OT3. JOSAP 2000, 49, 695–700. [Google Scholar] [CrossRef]
- Sugai, A.; Uda, I.; Itoh, Y.H.; Itoh, T. The core lipid composition of the 17 strains of hyperthermophilic archaea. J. Oleo Sci. 2004, 53, 41–44. [Google Scholar] [CrossRef] [Green Version]
- Tourte, M.; Schaeffer, P.; Grossi, V.; Oger, P.M. Functionalized membrane domains: An ancestral feature of Archaea? Front. Microbiol. 2020, 11, 526–540. [Google Scholar] [CrossRef]
- Callac, N.; Schaeffer, P.; Lesongeur, F.; Rattray, J.E.; Vannier, P.; Michoud, G.; Beauverger, M.; Gayet, N.; Rouxel, O.; Jebbar, M.; et al. Pyrococcus kukulkanii sp. nov., a hyperthermophilic, piezophilic archaeon isolated from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2016, 66, 3142–3149. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Birrien, J.-L.; Fouquet, Y.; Cherkashov, G.; Jebbar, M.; Querellou, J.; Oger, P.; Cambon-Bonavita, M.-A.; Xiao, X.; Prieur, D. Pyrococcus CH1, an obligate piezophilic hyperthermophile: Extending the upper pressure-temperature limits for life. ISME J. 2009, 3, 873–876. [Google Scholar] [CrossRef] [Green Version]
- Meador, T.B.; Gagen, E.J.; Loscar, M.E.; Goldhammer, T.; Yoshinaga, M.Y.; Wendt, J.; Thomm, M.; Hinrichs, K.U. Thermococcus kodakarensis modulates its polar membrane lipids and elemental composition according to growth stage and phosphate availability. Front. Microbiol. 2014, 5, 10–23. [Google Scholar] [CrossRef] [Green Version]
- Morikawa, M.; Izawa, Y.; Rashid, N.; Hoaki, T.; Imanaka, T. Purification and characterization of a thermostable thiol protease from a newly isolated hyperthermophilic Pyrococcus sp. Appl. Environ. Microbiol. 1994, 60, 4559–4566. [Google Scholar] [CrossRef] [Green Version]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damsté, J.S.S.; Rijpstra, W.I.C.; Hopmans, E.C.; Jung, M.-Y.; Kim, J.-G.; Rhee, S.-K.; Stieglmeier, M.; Schleper, C. Intact polar and core glycerol dibiphytanyl glycerol tetraether lipids of group, I.1a and I.1b Thaumarchaeota in soil. Appl. Environ. Microbiol. 2012, 78, 6866–6874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, L.M.; Taubner, R.S.; Bauersachs, T.; Steiner, M.; Schleper, C.; Peckmann, J.; Simon, K.M.; Birgel, D. Intact polar lipid and core lipid inventory of the hydrothermal vent methanogens Methanocaldococcus villosus and Methanothermococcus okinawensis. Org. Geochem. 2018, 126, 33–42. [Google Scholar] [CrossRef]
- Corcelli, A. The cardiolipin analogues of Archaea. Biochim. Biophys. Acta Biomembr. 2009, 1788, 2101–2106. [Google Scholar] [CrossRef] [Green Version]
- Hankin, J.A.; Murphy, R.C.; Barkley, R.M.; Gijón, M.A. Ion mobility and tandem mass spectrometry of phosphatidylglycerol and bis (monoacylglycerol) phosphate (BMP). Int. J. Mass Spectrom. 2015, 378, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Law, K.P.; Zhang, C.L. Current progress and future trends of mass spectrometry-based ar-chaeal lipidomics. Org. Geochem. 2019, 134, 45–61. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | MS Mode | Diethers * | Tetraethers * | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DGDsat+uns | DGD | GDGT0 | GTGT0 | GMGT0 | |||||||
| PI | PHexNAc | PHexHex | PHexHexNH2 | PHexHex+C5H8 | PHexHexNH2+C5H8 | PI+PI | PI | PI+PI | PI+PI | ||
| II+IIa+IIb+IIc+IId | III | IV | V | VI | VII | IX | X | XX | XXIV | ||
| P. furiosus | ESI+ | 59 | ND | ND | ND | ND | ND | 8 | ND | 5 | 28 |
| ESI- | 84 | 2 | ND | ND | ND | ND | 2 | ND | 1 | 11 | |
| P. yayanosii | ESI+ | 38 | 7 | 4 | ND | 5 | ND | 44 | 3 | Traces | ND |
| ESI- | 61 | 9 | 7 | ND | ND | ND | 22 | Traces | ND | ND | |
| T. kodakarensis | ESI+ | 31 | ND | ND | 41 | ND | 5 | 14 | 9 | Traces | ND |
| ESI- | 51 | 1 | Traces | 42 | Traces | ND | 3 | 2 | Traces | ND | |
| Species | Extract | Diether * | Tetraethers * | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DGD | GDGT0 | GDGT1 | GDGT2 | GDGT3 | GDGT4 | GTGT0 | GTGT1 | GTGT2 | GMGT0 | GMGT1 | GMGT2 | GMGT3 | GMGT4 | ||
| I | IX | XIII | XV | XVII | XVIII | XIX | XXI | XXII | XXIII | XXV | XXVI | XXVII | XXVIII | ||
| P. furiosus | totCL | 36 | 17 | 4 | 1 | Traces | Traces | Traces | Traces | Traces | 33 | 6 | 1 | Traces | Traces |
| CLfromIPL | 44 | 6 | Traces | Traces | Traces | Traces | 2 | Traces | Traces | 41 | 4 | Traces | Traces | Traces | |
| resCL | 45 | 4 | Traces | Traces | Traces | Traces | 2 | Traces | Traces | 43 | 4 | Traces | Traces | Traces | |
| P. yayanosii | CLfromIPL | 35 | 64 | ND | ND | ND | ND | Traces | ND | ND | ND | ND | ND | ND | ND |
| resCL | 7 | 92 | ND | ND | ND | ND | Traces | ND | ND | ND | ND | ND | ND | ND | |
| T. kodakarensis | totCL | 77 | 23 | ND | ND | ND | ND | Traces | ND | ND | ND | ND | ND | ND | ND |
| CLfromIPL | 56 | 43 | ND | ND | ND | ND | 1 | ND | ND | ND | ND | ND | ND | ND | |
| resCL | 50 | 49 | ND | ND | ND | ND | Traces | ND | ND | ND | ND | ND | ND | ND | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tourte, M.; Kuentz, V.; Schaeffer, P.; Grossi, V.; Cario, A.; Oger, P.M. Novel Intact Polar and Core Lipid Compositions in the Pyrococcus Model Species, P. furiosus and P. yayanosii, Reveal the Largest Lipid Diversity Amongst Thermococcales. Biomolecules 2020, 10, 830. https://doi.org/10.3390/biom10060830
Tourte M, Kuentz V, Schaeffer P, Grossi V, Cario A, Oger PM. Novel Intact Polar and Core Lipid Compositions in the Pyrococcus Model Species, P. furiosus and P. yayanosii, Reveal the Largest Lipid Diversity Amongst Thermococcales. Biomolecules. 2020; 10(6):830. https://doi.org/10.3390/biom10060830
Chicago/Turabian StyleTourte, Maxime, Vanessa Kuentz, Philippe Schaeffer, Vincent Grossi, Anais Cario, and Philippe M. Oger. 2020. "Novel Intact Polar and Core Lipid Compositions in the Pyrococcus Model Species, P. furiosus and P. yayanosii, Reveal the Largest Lipid Diversity Amongst Thermococcales" Biomolecules 10, no. 6: 830. https://doi.org/10.3390/biom10060830