Black Soldier Fly (Hermetia illucens) Larvae Protein Derivatives: Potential to Promote Animal Health


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Raw Materials
2.3. Protein Quantification
2.4. DPPH Assay
2.5. ABTS Assay
2.6. Myeloperoxidase (MPO) Activity Using Specific Immunological Extraction Followed by Enzymatic Detection (SIEFED) Assay
2.7. Myeloperoxidase (MPO) Activity Using Classical Measurement
2.8. Cellular Antioxidant Activity
2.9. Statistical Analyses
3. Results
3.1. Protein Quantification
3.2. DPPH Assay
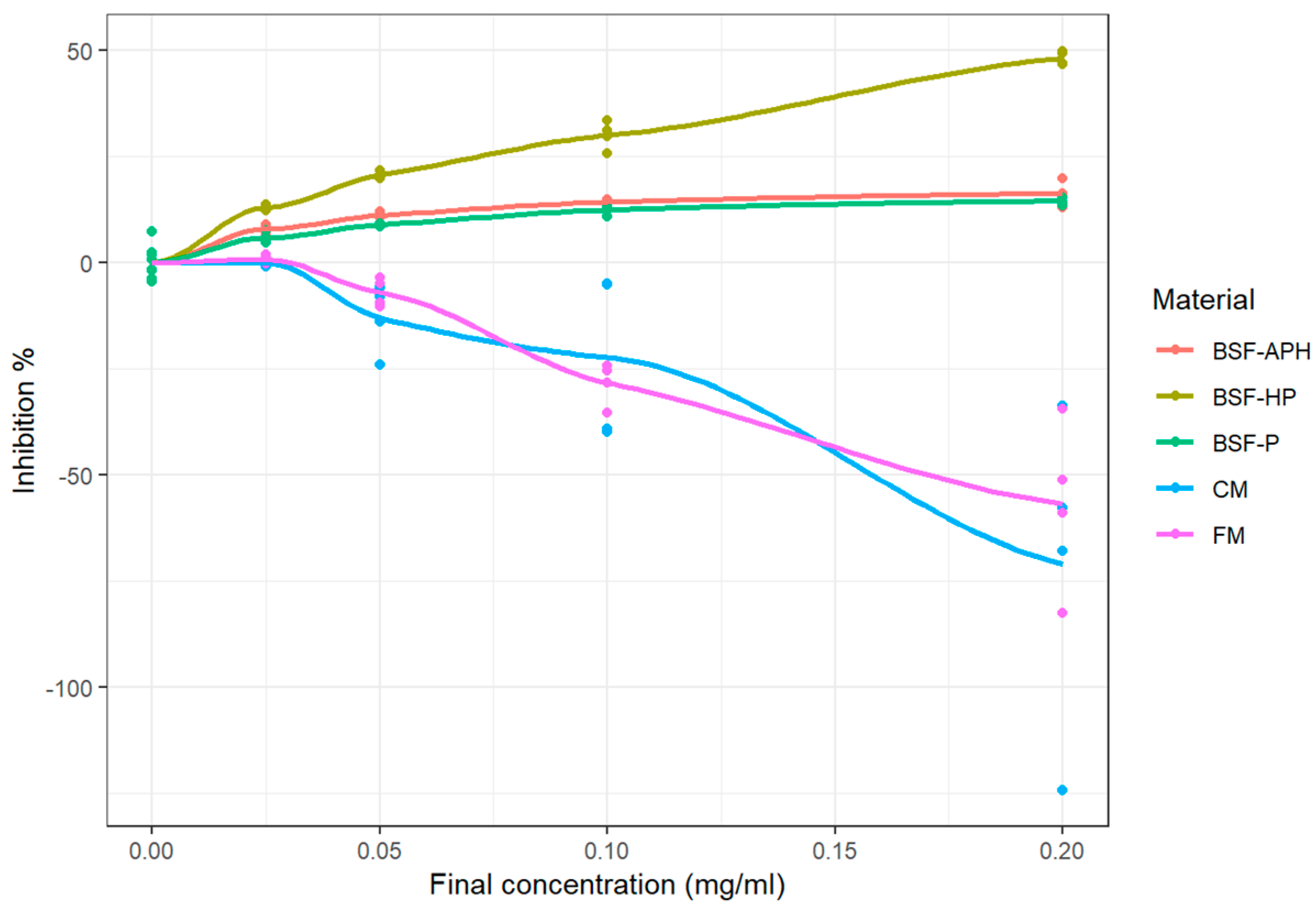
3.3. ABTS Assay
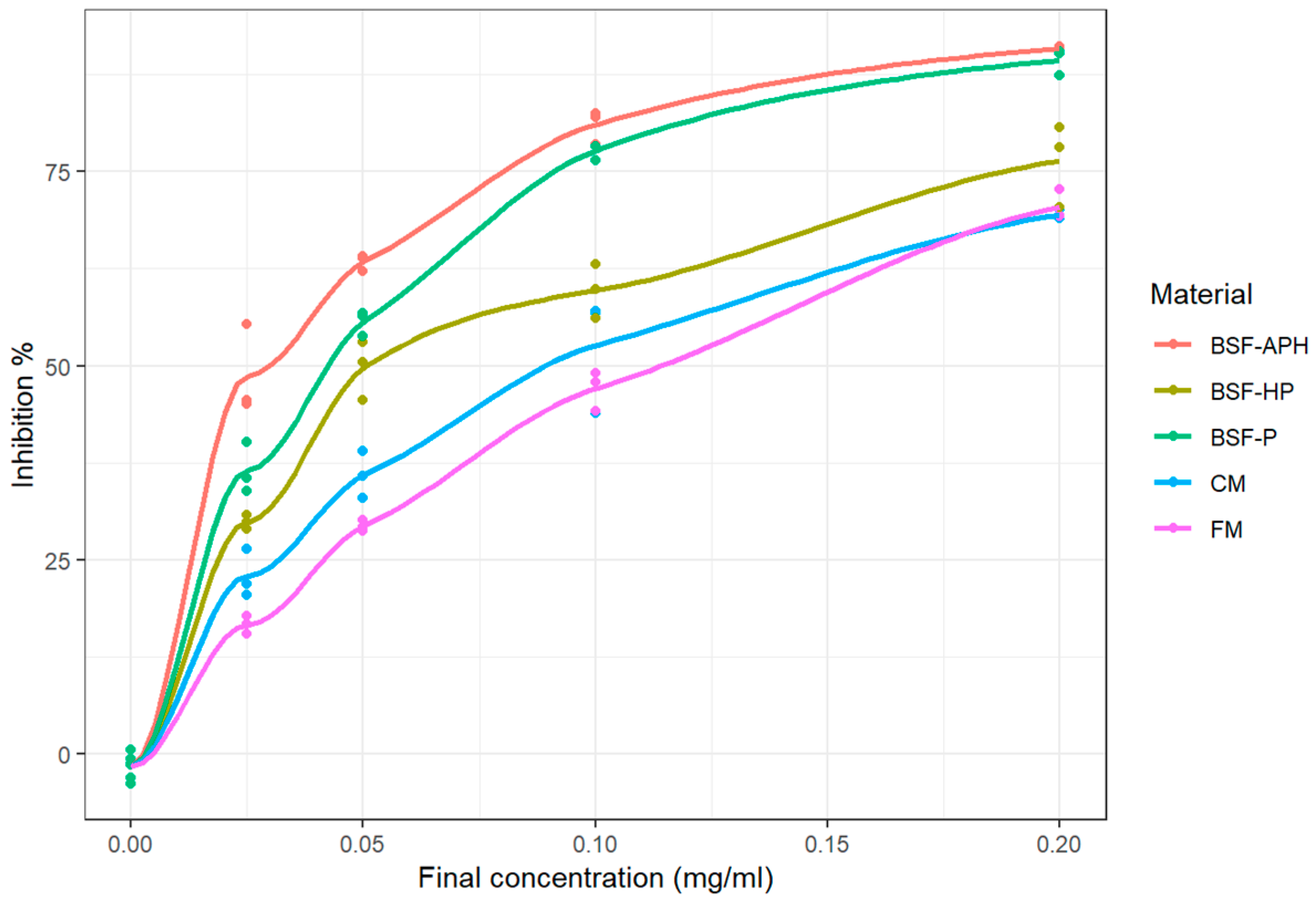
3.4. Myeloperoxidase (MPO) Activity Using Specific Immunological Extraction Followed by Enzymatic Detection (SIEFED) Assay
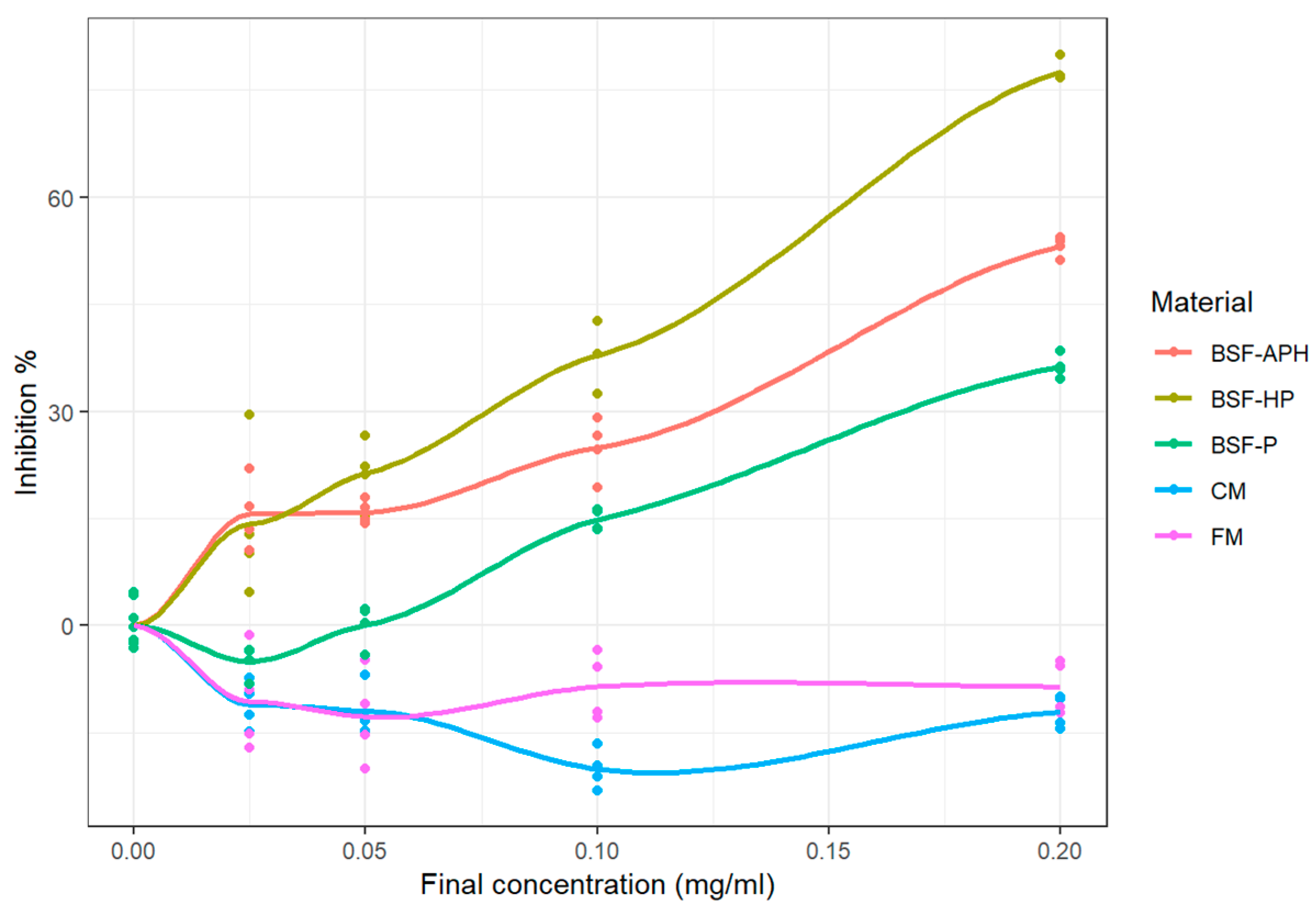
3.5. Myeloperoxidase (MPO) Activity Using Classical Assay
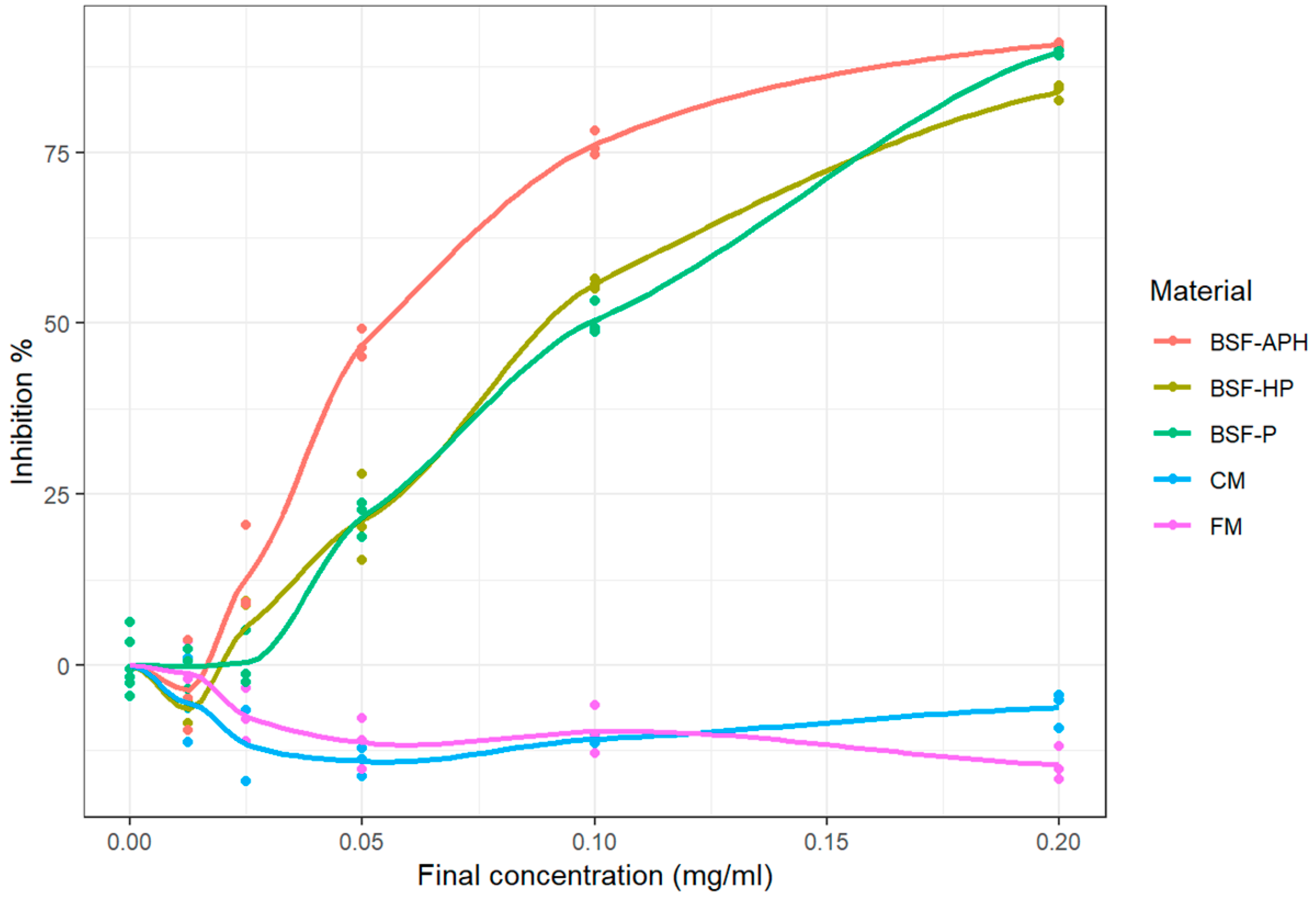
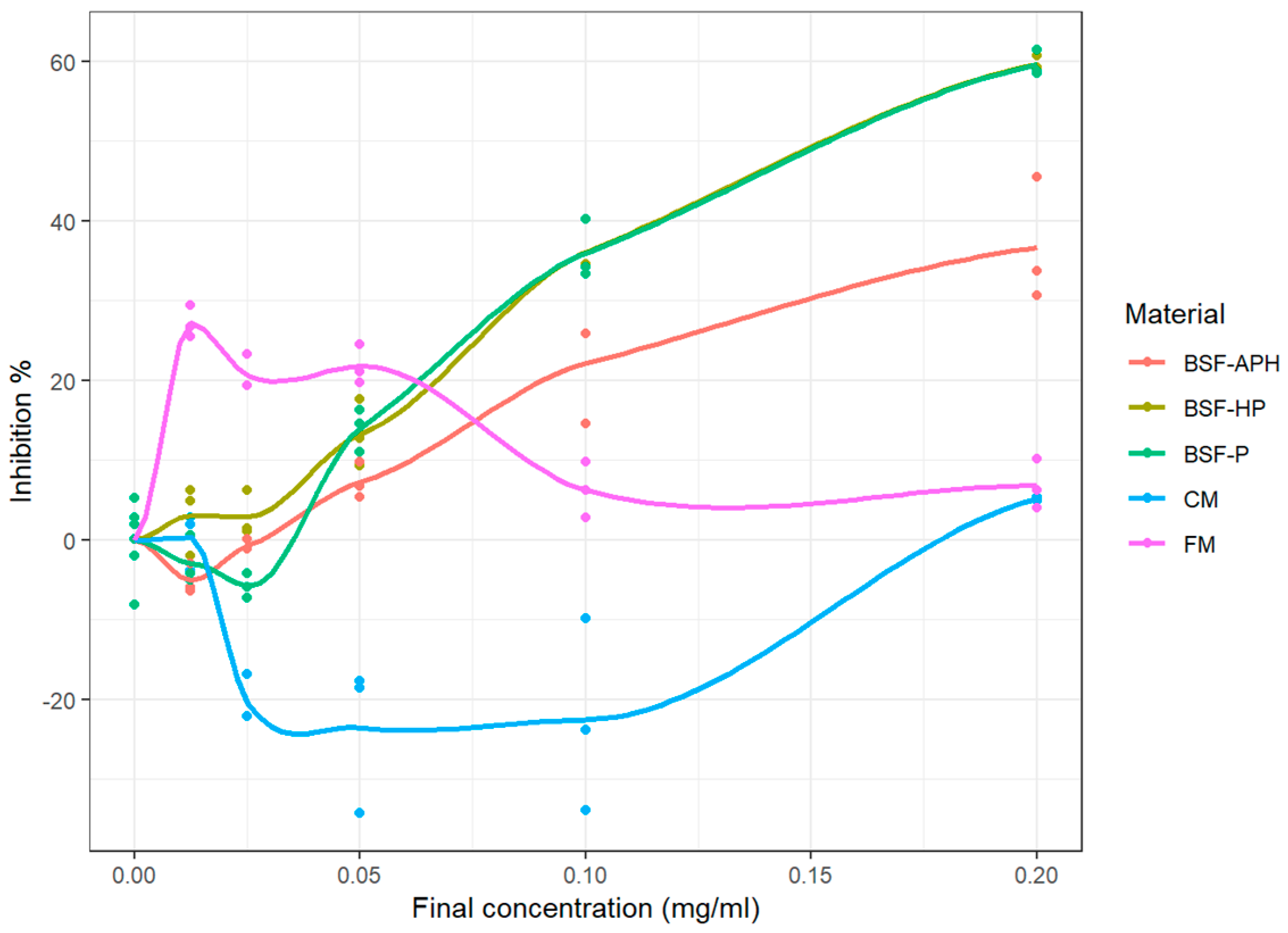
3.6. Cellular Antioxidant Activity
4. Discussion
4.1. Protein Quantification
4.2. DPPH Radical Scavenging Activity
4.3. ABTS Cation Radical Scavening Activity
4.4. Neutrophil Response Modulation Activity
4.5. MPO Response Modulation Activity (SIEFED and Classical Assay)
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Paul, A.; Frederich, M.; Uyttenbroeck, R.; Hatt, S.; Malik, P.; Lebecque, S.; Hamaidia, M.; Miazek, K.; Goffin, D.; Willems, L.; et al. Grasshoppers as a food source? A review. Biotechnol. Agron. Soc. Environ. 2016, 20, 337–352. [Google Scholar]
- Paul, A.; Frederich, M.; Uyttenbroeck, R.; Malik, P.; Filocco, S.; Richel, A.; Heuskin, S.; Alabi, T.; Megido, R.C.; Franck, T.; et al. Nutritional composition and rearing potential of the meadow grasshopper (Chorthippus parallelus Zetterstedt). J. Asia-Pac. Entomol. 2016, 19, 1111–1116. [Google Scholar] [CrossRef]
- Paul, A.; Frederich, M.; Megido, R.C.; Alabi, T.; Malik, P.; Uyttenbroeck, R.; Francis, F.; Blecker, C.; Haubruge, E.; Lognay, G.; et al. Insect fatty acids: A comparison of lipids from three Orthopterans and Tenebrio molitor L. larvae. J. Asia-Pac. Entomol. 2017, 20, 337–340. [Google Scholar] [CrossRef]
- IPIFF. Promoting Insects for Human Consumption & Animal Feed—IPIFF. Available online: http://ipiff.org/ (accessed on 5 February 2020).
- Gasco, L.; Gai, F.; Maricchiolo, G.; Genovese, L.; Ragonese, S.; Bottari, T.; Caruso, G. Fishmeal Alternative Protein Sources for Aquaculture Feeds. In Feeds for the Aquaculture Sector: Current Situation and Alternative Sources; Gasco, L., Gai, F., Maricchiolo, G., Genovese, L., Ragonese, S., Bottari, T., Caruso, G., Eds.; Springer Briefs in Molecular Science; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–28. ISBN 978-3-319-77941-6. [Google Scholar]
- Deng, P.; Utterback, P.L.; Parsons, C.M.; Hancock, L.; Swanson, K.S. Chemical composition, true nutrient digestibility, and true metabolizable energy of novel pet food protein sources using the precision-fed cecectomized rooster assay. J. Anim. Sci. 2016, 94, 3335–3342. [Google Scholar] [CrossRef]
- Star, L.; Arsiwalla, T.; Molist, F.; Leushuis, R.; Dalim, M.; Paul, A. Gradual Provision of Live Black Soldier Fly (Hermetia illucens) Larvae to Older Laying Hens: Effect on Production Performance, Egg Quality, Feather Condition and Behavior. Animals 2020, 10, 216. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, E.; Belghit, I.; Johansen, J.; Leushuis, R.; Lock, E.J.; Melsen, D.; Kathirampatti Ramasamy Shanmugam, R.; Van Loon, J.; Paul, A. Growth and Safety Assessment of Feed Streams for Black Soldier Fly Larvae: A Case Study with Aquaculture Sludge. Animals 2019, 9, 189. [Google Scholar] [CrossRef] [Green Version]
- Belghit, I.; Liland, N.S.; Gjesdal, P.; Biancarosa, I.; Menchetti, E.; Li, Y.; Waagbø, R.; Krogdahl, Å.; Lock, E.J. Black soldier fly larvae meal can replace fish meal in diets of sea-water phase Atlantic salmon (Salmo salar). Aquaculture 2018. [Google Scholar] [CrossRef]
- Belghit, I.; Waagbø, R.; Lock, E.J.; Liland, N.S. Insect-based diets high in lauric acid reduce liver lipids in freshwater Atlantic salmon. Aquac. Nutr. 2019, 25, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Kortner, T.M.; Chikwati, E.M.; Munang’andu, H.M.; Lock, E.J.; Krogdahl, Å. Gut health and vaccination response in pre-smolt Atlantic salmon (Salmo salar) fed black soldier fly (Hermetia illucens) larvae meal. Fish Shellfish Immunol. 2018, 86, 1106–1113. [Google Scholar] [CrossRef]
- Bosch, G.; Zhang, S.; Oonincx, D.G.A.B.; Hendriks, W.H. Protein quality of insects as potential ingredients for dog and cat foods. J. Nutr. Sci. 2014, 3. [Google Scholar] [CrossRef] [Green Version]
- Meyer, L.F.; Kolln, M.; Kamphues, J. Dog food containing insects? Studies on compound feed with larvae of the black soldier fly as the main protein source. Kleintierpraxis 2019, 64, 124–135. [Google Scholar] [CrossRef]
- Head, E.; Rofina, J.; Zicker, S. Oxidative Stress, Aging, and Central Nervous System Disease in the Canine Model of Human Brain Aging. Vet. Clin. N. Am. 2008, 38, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Blier, P. Fish Health: An Oxidative Stress Perspective. Fish Aquac. J. 2014, 5, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Tsumbu, C.N.; Deby-Dupont, G.; Tits, M.; Angenot, L.; Frederich, M.; Kohnen, S.; Mouithys-Mickalad, A.; Serteyn, D.; Franck, T. Polyphenol content and modulatory activities of some tropical dietary plant extracts on the oxidant activities of neutrophils and myeloperoxidase. Int. J. Mol. Sci. 2012, 13, 628–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, A. Field Border Flowering Strips As a Source of Valuable Compounds. Ph.D. Thesis, Gembloux Agro-Bio Tech University of Liège, Gembloux, Belgique, 2017. [Google Scholar]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharm. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant Activity of Proteins and Peptides. Crit. Rev. Food Sci. Nutr. 2008, 48, 430–441. [Google Scholar] [CrossRef]
- Zielińska, E.; Karaś, M.; Jakubczyk, A. Antioxidant activity of predigested protein obtained from a range of farmed edible insects. Int. J. Food Sci. Technol. 2017, 52, 306–312. [Google Scholar] [CrossRef]
- Zielińska, E.; Baraniak, B.; Karaś, M. Antioxidant and Anti-Inflammatory Activities of Hydrolysates and Peptide Fractions Obtained by Enzymatic Hydrolysis of Selected Heat-Treated Edible Insects. Nutrients 2017, 9, 970. [Google Scholar] [CrossRef] [Green Version]
- Zielińska, E.; Baraniak, B.; Karaś, M. Identification of antioxidant and anti-inflammatory peptides obtained by simulated gastrointestinal digestion of three edible insects species (Gryllodes sigillatus, Tenebrio molitor, Schistocerca gragaria). Int. J. Food Sci. Technol. 2018, 53, 2542–2551. [Google Scholar] [CrossRef]
- Vercruysse, L.; Smagghe, G.; Herregods, G.; Van Camp, J. ACE Inhibitory Activity in Enzymatic Hydrolysates of Insect Protein. J. Agric. Food Chem. 2005, 53, 5207–5211. [Google Scholar] [CrossRef]
- Paul, A. EU Patent Application: N2023838—Hydrolysate of Water Soluble Insect Proteins.
- Arsiwalla, T.; Aarts, K.W.P. United States Patent Application: 0150223508—Method to Convert Insects or Worms into Nutrient Streams and Compositions Obtained Thereby A1. Available online: https://patents.google.com/patent/US20150223508A1/en (accessed on 27 March 2020).
- Zhu, D.; Huang, X.; Tu, F.; Wang, C.; Yang, F. Preparation, antioxidant activity evaluation, and identification of antioxidant peptide from black soldier fly (Hermetia illucens L.) larvae. J. Food Biochem. 2020, e13186. [Google Scholar] [CrossRef] [PubMed]
- Firmansyah, M.; Abduh, M.Y. Production of protein hydrolysate containing antioxidant activity from Hermetia illucens. Heliyon 2019, 5, e02005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Arnao, M.B.; Cano, A.; Acosta, M. The hydrophilic and lipophilic contribution to total antioxidant activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Franck, T.; Kohnen, S.; Boudjeltia, K.Z.; Van Antwerpen, P.; Bosseloir, A.; Niesten, A.; Gach, O.; Nys, M.; Deby-Dupont, G.; Serteyn, D. A new easy method for specific measurement of active myeloperoxidase in human biological fluids and tissue extracts. Talanta 2009, 80, 723–729. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Holden-Wiltse, J.; Wang, J.; Liang, H. A Strategy to Model Nonmonotonic Dose-Response Curve and Estimate IC50. PLoS ONE 2013, 8, e69301. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Chang, S.K.C.; Zhang, Y. Protein Analysis. In Food Analysis; Nielsen, S.S., Ed.; Food Science Text Series; Springer International Publishing: Cham, Switzerland, 2017; pp. 315–331. ISBN 978-3-319-45776-5. [Google Scholar]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Rojas, O.J.; Stein, H.H. Effects of replacing fish, chicken, or poultry by-product meal with fermented soybean meal in diets fed to weanling pigs. Rev. Colomb. Cienc. Pecu. 2015, 28, 22–41. [Google Scholar]
- Li, P.; Wu, G. Composition of amino acids and related nitrogenous nutrients in feedstuffs for animal diets. Amino Acids 2020, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Zou, T.B.; He, T.P.; Li, H.B.; Tang, H.W.; Xia, E.Q. The Structure-Activity Relationship of the Antioxidant Peptides from Natural Proteins. Molecules 2016, 21, 72. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.R.; Zhang, L.; Ding, D.G.; Chi, C.F.; Wang, B.; Huo, J.C. Preparation, Identification, and Activity Evaluation of Eight Antioxidant Peptides from Protein Hydrolysate of Hairtail (Trichiurus japonicas) Muscle. Mar. Drugs 2019, 17, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.; Wu, X.; Zhang, Y.; Li, X.; Ma, S.; Li, J. Purification and characterization of three antioxidant peptides from protein hydrolysate of grass carp (Ctenopharyngodon idella) skin. J. Funct. Foods 2015, 16, 234–242. [Google Scholar] [CrossRef]
- Esfandi, R.; Walters, M.E.; Tsopmo, A. Antioxidant properties and potential mechanisms of hydrolyzed proteins and peptides from cereals. Heliyon 2019, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. The Production of Fish Meal and Oil; FAO: Rome, Italy, 1986. [Google Scholar]
- Diedrich, D.M. For Pet’s Sake, Do Something! Book Two: How to Heal You Sick, Overfed, and Bored Pets with Nutrition, Supplements, Herbs, and Exercise (Mom’s Choice Award Recipient); Two Paws Up Press: Anaheim, CA, USA, 2007; ISBN 978-0-9713812-8-5. [Google Scholar]
- Nygaard, H. Standard Norwegian fishmeal- and Fishoil Process. Heat Treatment Requirements; Nofima: Tromsø, Norway, 2010; p. 30. [Google Scholar]
- Bartosz, G. Food Oxidants and Antioxidants: Chemical, Biological, and Functional Properties; CRC Press: Boca Raton, FL, USA, 2013; ISBN 978-1-4398-8242-9. [Google Scholar]
- Yu, T.Y.; Morton, J.D.; Clerens, S.; Dyer, J.M. Cooking-Induced Protein Modifications in Meat. Compr. Rev. Food Sci. Food Saf. 2017, 16, 141–159. [Google Scholar] [CrossRef] [Green Version]
- Celi, P.; Gabai, G. Oxidant/Antioxidant Balance in Animal Nutrition and Health: The Role of Protein Oxidation. Front. Vet. Sci. 2015, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallegos, M.E.H.; Zannatha, M.M.I.; Osornio, E.G.; Sánchez, A.S.; del Rio, F.A.P. The Activities of Six Exo- and Endopeptidases in the Substantia Nigra, Neostriatum, and Cortex of the Rat Brain. Neurochem. Res. 1999, 24, 1557–1561. [Google Scholar] [CrossRef]
- Perobelli, S.M.; Galvani, R.G.; Gonçalves-Silva, T.; Xavier, C.R.; Nóbrega, A.; Bonomo, A. Plasticity of neutrophils reveals modulatory capacity. Braz. J. Med. Biol. Res. 2015, 48, 665–675. [Google Scholar] [CrossRef] [Green Version]
- Swain, S.D.; Rohn, T.T.; Quinn, M.T. Neutrophil Priming in Host Defense: Role of Oxidants as Priming Agents. Antioxid. Redox Signal. 2002, 4, 69–83. [Google Scholar] [CrossRef]
- Zholobenko, A.; Mouithys-Mickalad, A.; Modriansky, M.; Serteyn, D.; Franck, T. Polyphenols from Silybum marianum inhibit in vitro the oxidant response of equine neutrophils and myeloperoxidase activity. J. Vet. Pharmacol. Ther. 2016, 39, 592–601. [Google Scholar] [CrossRef]
- Yamka, R.M.; Jamikorn, U.; True, A.D.; Harmon, D.L. Evaluation of low-ash poultry meal as a protein source in canine foods. J. Anim. Sci. 2003, 81, 2279–2284. [Google Scholar] [CrossRef] [PubMed]
- Ben Hamed, S.; Tavares Ranzani-Paiva, M.J.; Tachibana, L.; de Carla Dias, D.; Ishikawa, C.M.; Esteban, M.A. Fish pathogen bacteria: Adhesion, parameters influencing virulence and interaction with host cells. Fish Shellfish Immunol. 2018, 80, 550–562. [Google Scholar] [CrossRef] [PubMed]
- Washabau, R.J.; Day, M.J. Canine and Feline Gastroenterology—E-Book; Elsevier Health Sciences: Oxford, UK, 2012; ISBN 978-1-4377-0302-3. [Google Scholar]
- Wang, B.; Xie, N.; Li, B. Influence of peptide characteristics on their stability, intestinal transport, and in vitro bioavailability: A review. J. Food Biochem. 2019, 43, e12571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, R.W.; Wyse, G.A.; Anderson, M. Animal Physiology, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2013; ISBN 9780878935598. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrients | Chickenmeal | Fishmeal |
|---|---|---|
| Moisture (g/kg) | 60.0 | 100.0 |
| Crude protein (g/kg) | 700.0 | 710.0 |
| Crude fat (g/kg) | 120.0 | 120.0 |
| Added antioxidant | No | Yes (E324 *) |
| Form | Powder | |
| Nutrients | BSF-P 1 | BSF-HP 2 | BSF-APH 3 |
|---|---|---|---|
| Moisture (g/kg) | 700.0 a | 700.0 a | 55 a |
| Crude protein (g/kg) | 120 a | 120 a | 455 a |
| Crude fat (g/kg) | 122.5 a | 122.5 a | 35 a |
| Added antioxidant | No | No | No |
| % of total proteins <1000 Da | >6 | >24 | >98 |
| Form | Frozen minced meat | Powder | |
| Product | Product Used for Testing in all the Assays | Mean Optical Density | Protein Concentration (mg/mL) |
|---|---|---|---|
| BSF-P 1 | Water-soluble extract | 0.486 | 1.013 |
| BSF-HP 2 | Water-soluble extract | 0.365 | 0.648 |
| BSF-APH 3 | Product as provided by supplier | 0.383 | 0.702 |
| FM 4 | Water-soluble extract | 0.425 | 0.829 |
| CM 5 | Water-soluble extract | 0.481 | 0.998 |
| Assay | BSF-P 1 | BSF-HP 2 | BSF-APH 3 | FM 4 | CM 5 |
|---|---|---|---|---|---|
| DPPH | NE c | NE c | NE c | NE c | PO d |
| ABTS | 0.04 | 0.05 | 0.03 | 0.11 | 0.09 |
| MPO a SIEFED | NE c | 0.14 | 0.18 | PO d | PO d |
| MPO a Classical | 0.10 | 0.09 | 0.05 | PO d | PO d |
| CAA b | 0.15 | 0.15 | NE c | NE c | NE c |
| Assay | Parameter | BSF-P 1 | BSF-HP 2 | BSF-APH 3 | FM 4 | CM 5 |
|---|---|---|---|---|---|---|
| DPPH | Emax (%) | 14.52 | 48.09 | 16.26 | 0.75 | PO c |
| C * (mg/mL) | 0.20 | 0.20 | 0.20 | 0.03 | - | |
| ABTS | Emax (%) | 89.33 | 76.32 | 90.81 | 70.40 | 69.39 |
| C * (mg/mL) | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | |
| MPO a SIEFED | Emax (%) | 36.23 | 77.58 | 53.08 | PO c | PO c |
| C * (mg/mL) | 0.20 | 0.20 | 0.20 | - | - | |
| MPO a Classical | Emax (%) | 89.66 | 83.82 | 90.86 | PO c | PO c |
| C * (mg/mL) | 0.20 | 0.20 | 0.20 | - | - | |
| CAA b | Emax (%) | 59.57 | 59.64 | 36.62 | 21.81 | 5.08 |
| C * (mg/mL) | 0.20 | 0.20 | 0.20 | 0.05 | 0.20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mouithys-Mickalad, A.; Schmitt, E.; Dalim, M.; Franck, T.; Tome, N.M.; van Spankeren, M.; Serteyn, D.; Paul, A. Black Soldier Fly (Hermetia illucens) Larvae Protein Derivatives: Potential to Promote Animal Health. Animals 2020, 10, 941. https://doi.org/10.3390/ani10060941
Mouithys-Mickalad A, Schmitt E, Dalim M, Franck T, Tome NM, van Spankeren M, Serteyn D, Paul A. Black Soldier Fly (Hermetia illucens) Larvae Protein Derivatives: Potential to Promote Animal Health. Animals. 2020; 10(6):941. https://doi.org/10.3390/ani10060941
Chicago/Turabian StyleMouithys-Mickalad, Ange, Eric Schmitt, Monika Dalim, Thierry Franck, Nuria Martin Tome, Michel van Spankeren, Didier Serteyn, and Aman Paul. 2020. "Black Soldier Fly (Hermetia illucens) Larvae Protein Derivatives: Potential to Promote Animal Health" Animals 10, no. 6: 941. https://doi.org/10.3390/ani10060941