The Secretory Apparatus of Tabernaemontana ventricosa Hochst. ex A.DC. (Apocynaceae): Laticifer Identification, Characterization and Distribution




Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
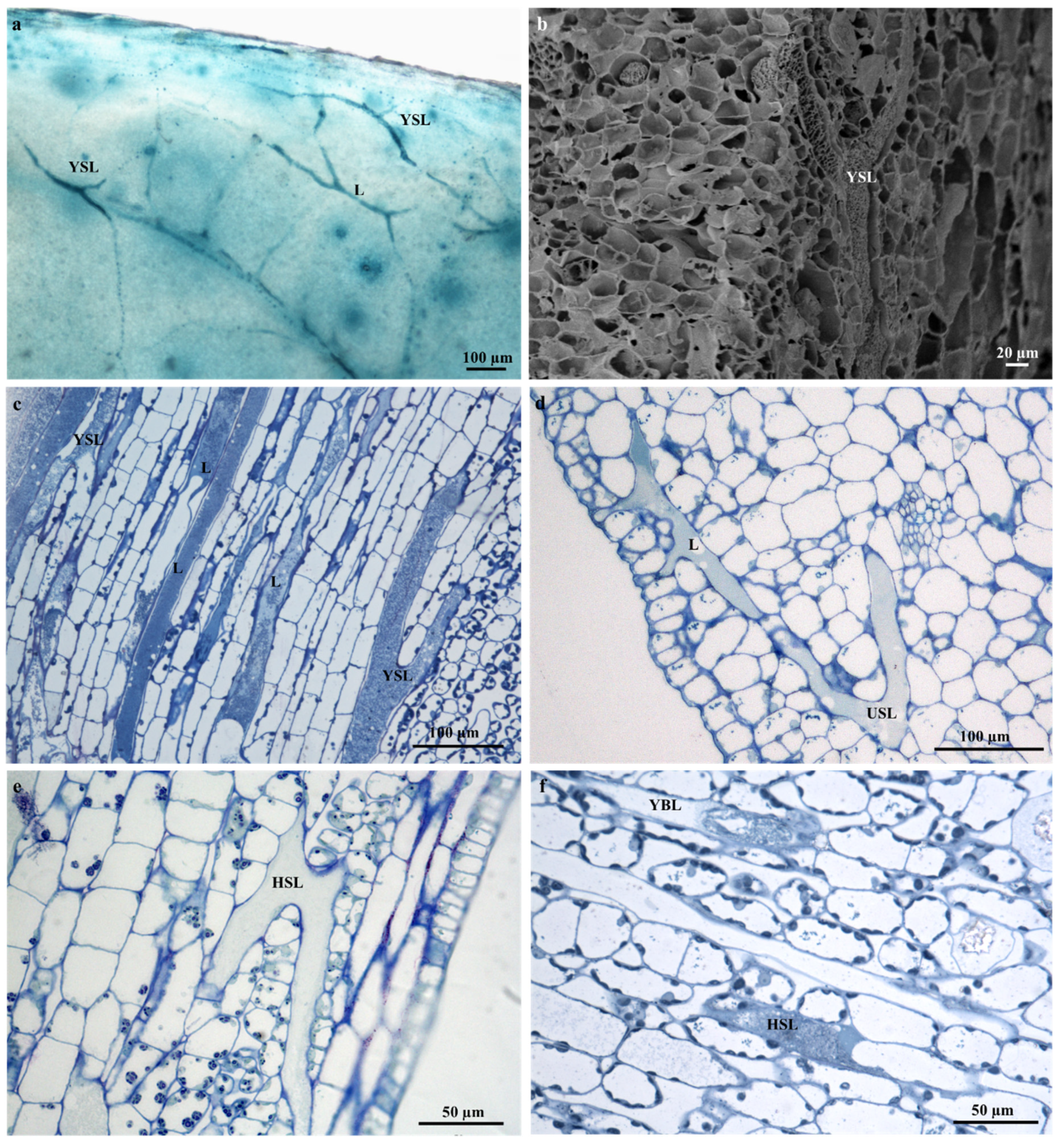
2.1. Leaf Micromorphology
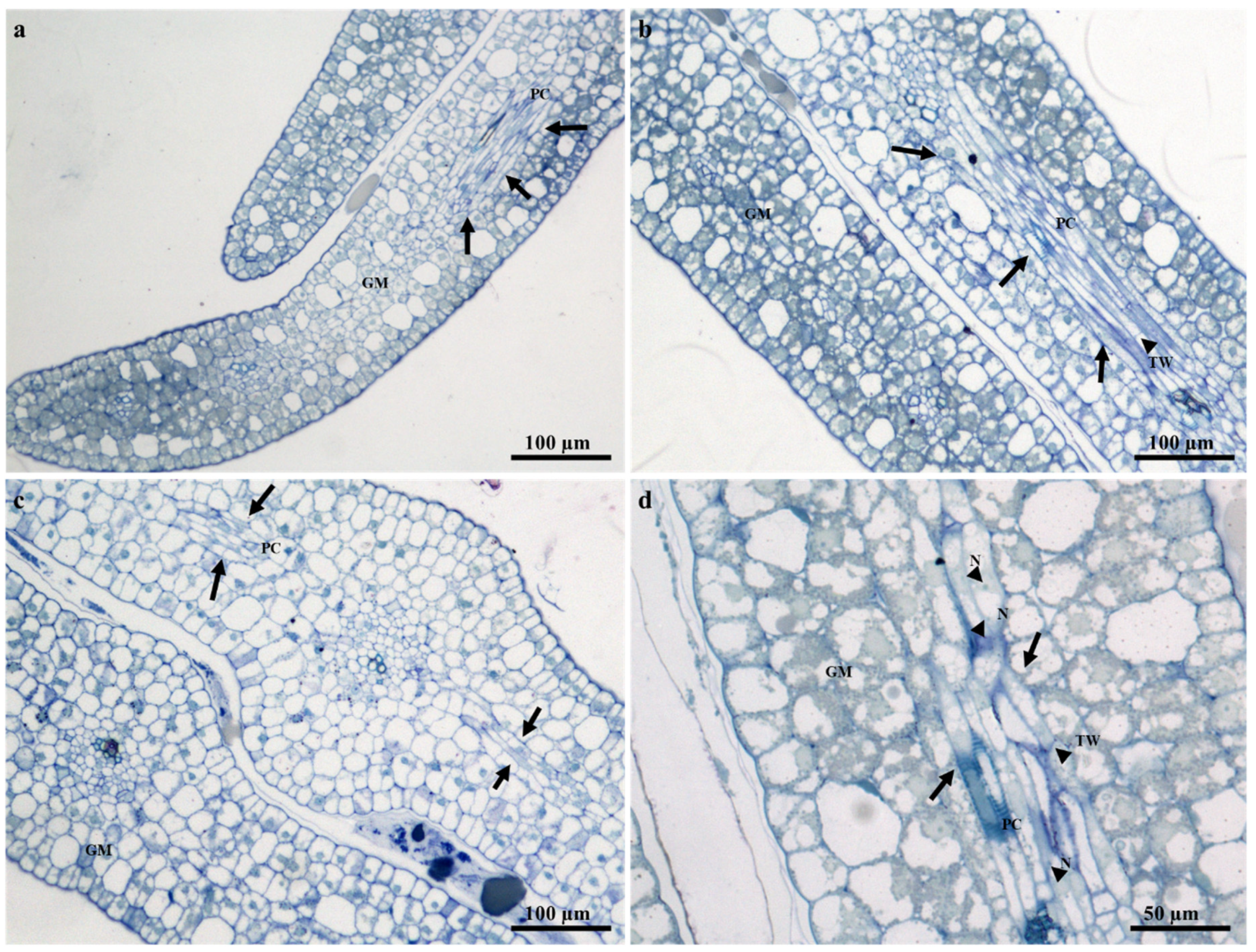
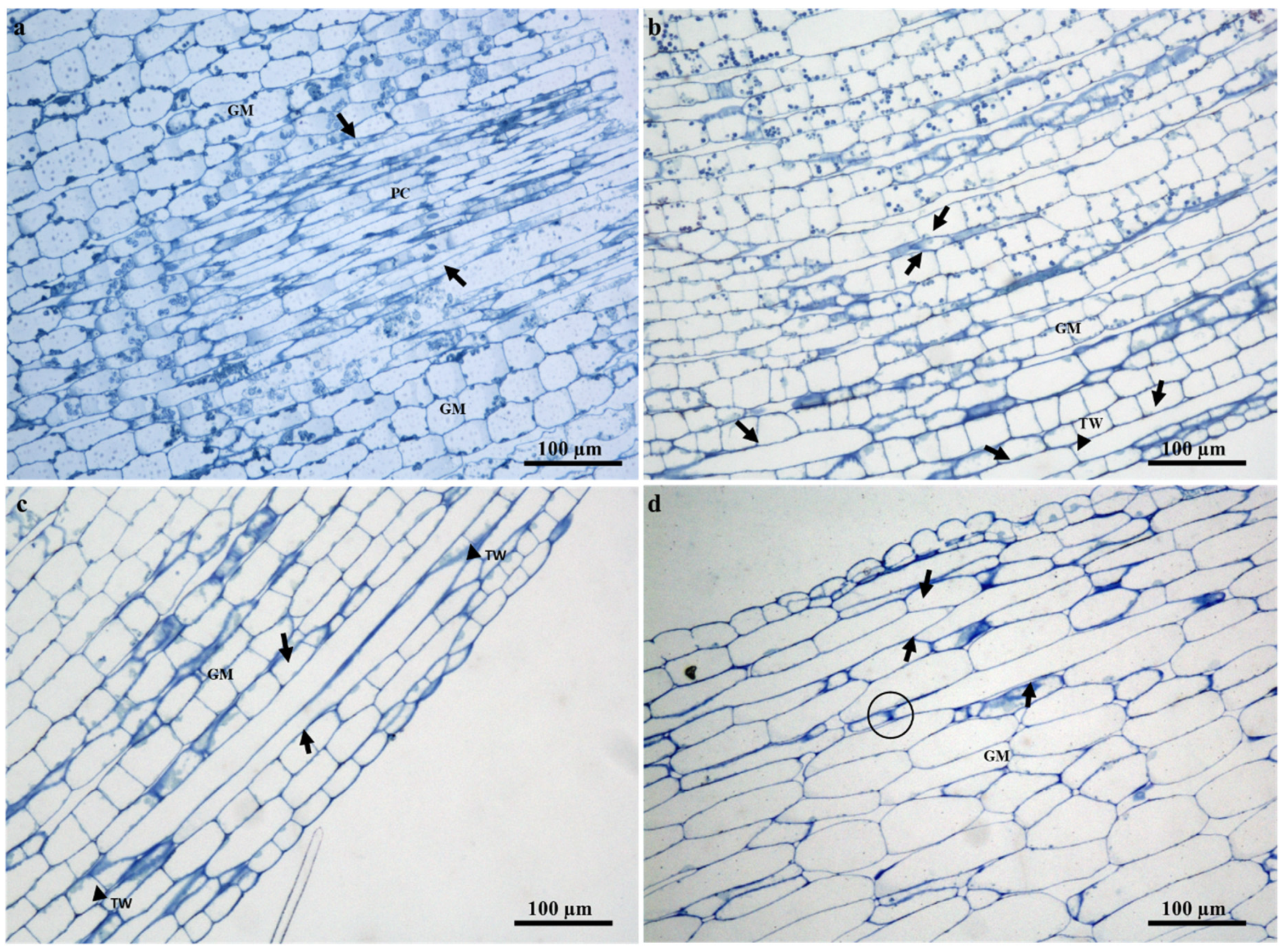
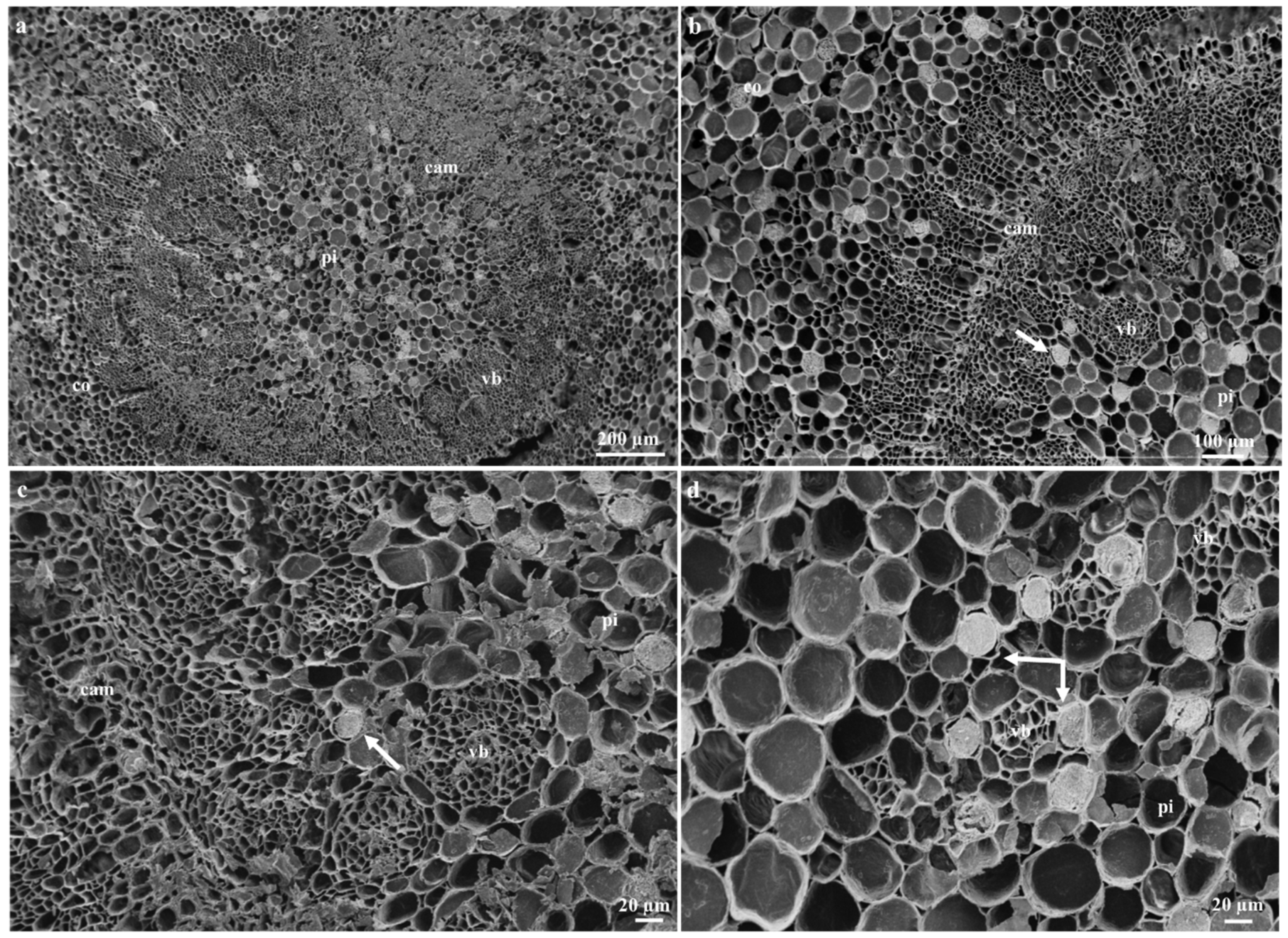
2.2. Ontogeny and Structure of Laticifers
2.3. Distribution of Laticifers
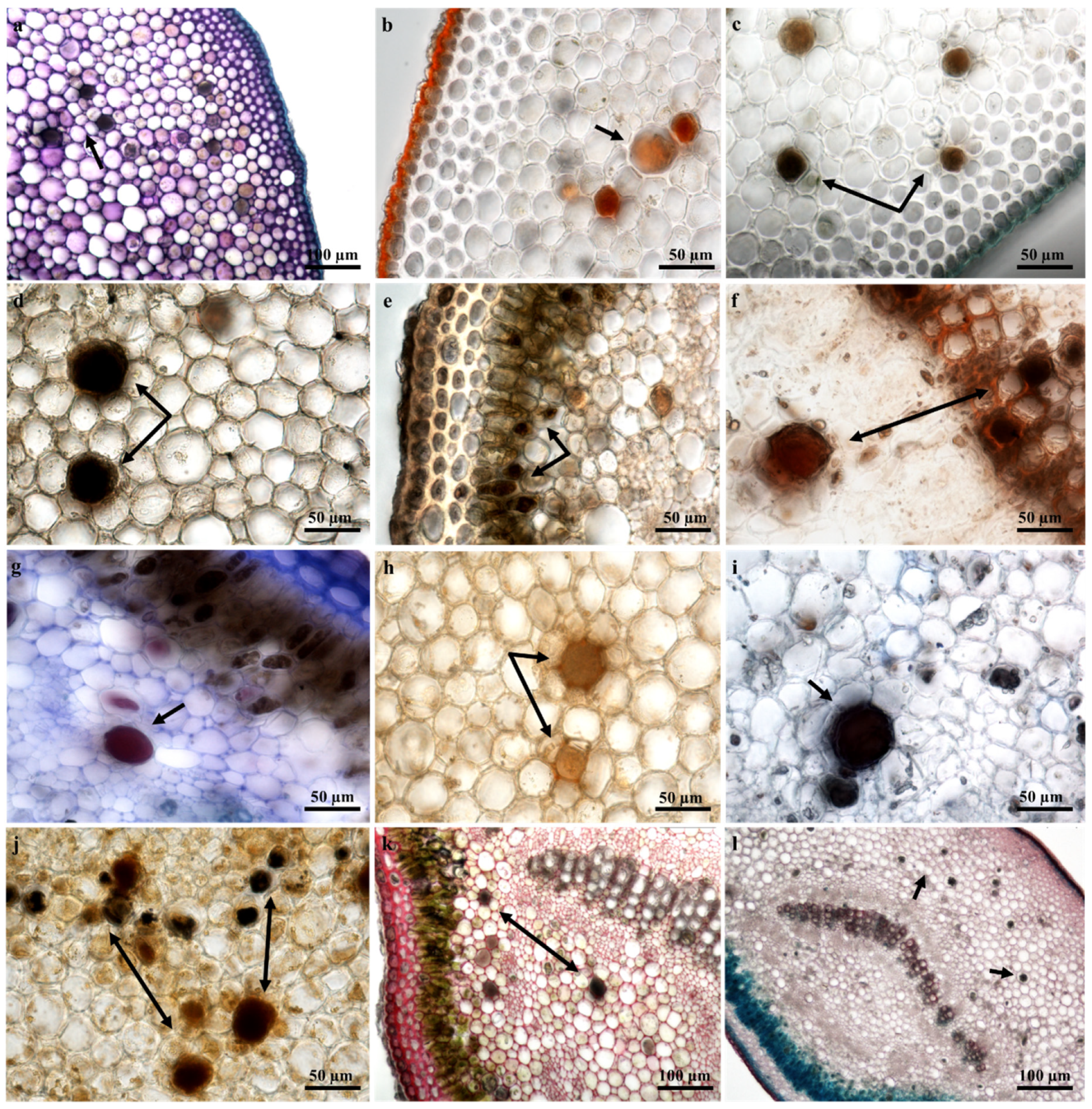
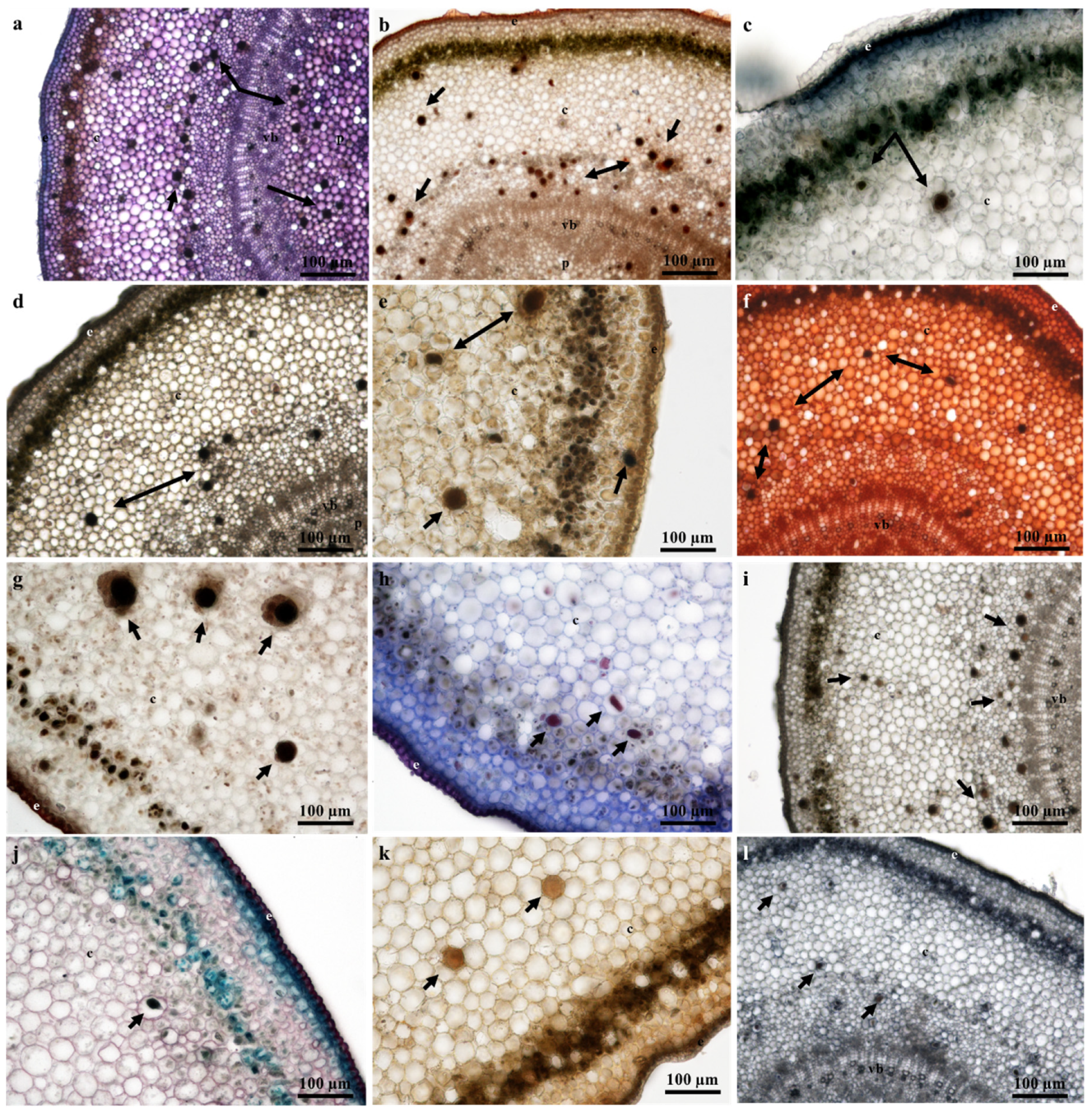
2.4. Laticifer Histochemical Characterization
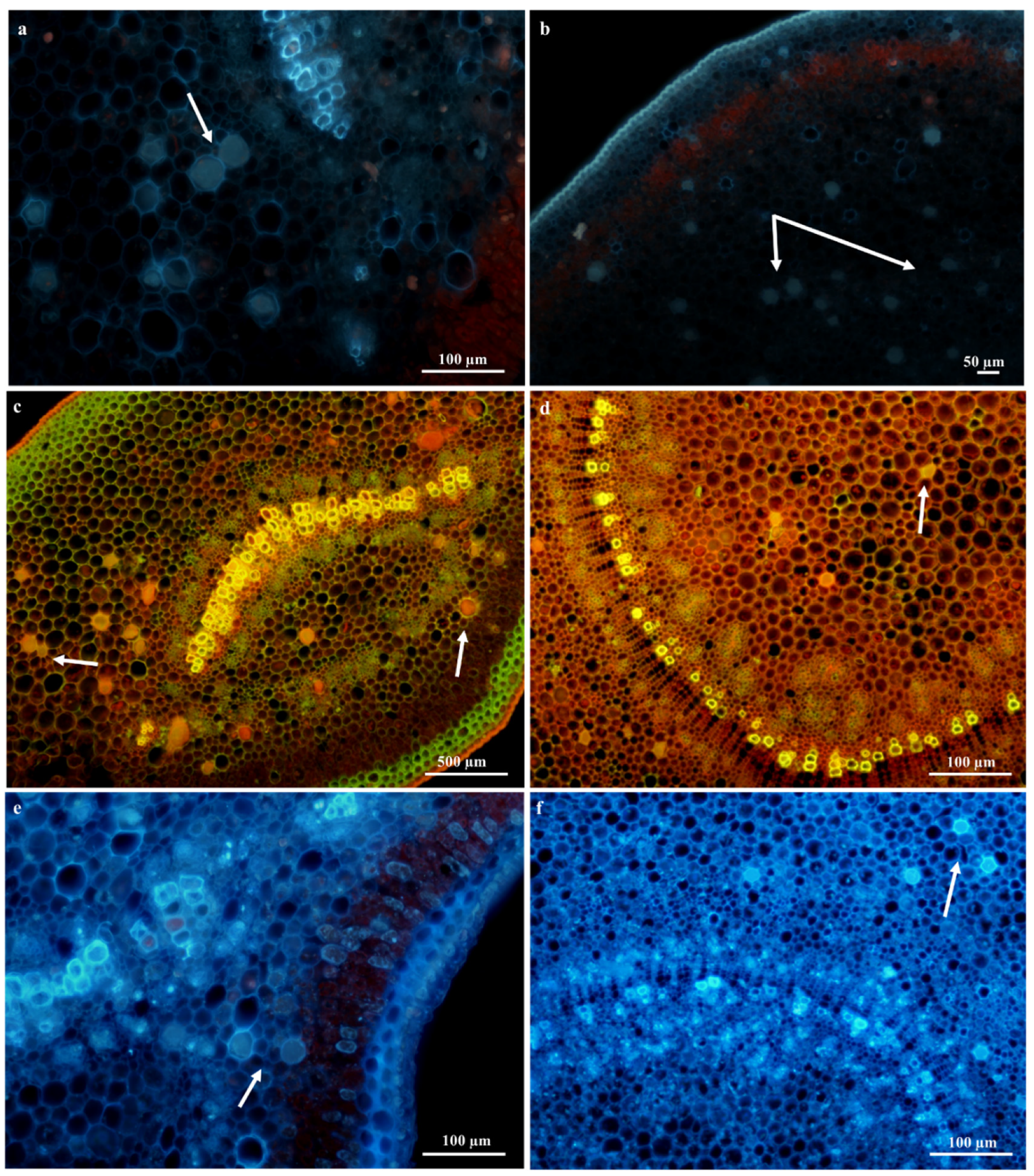
Laticifer Fluorescence Microscopy
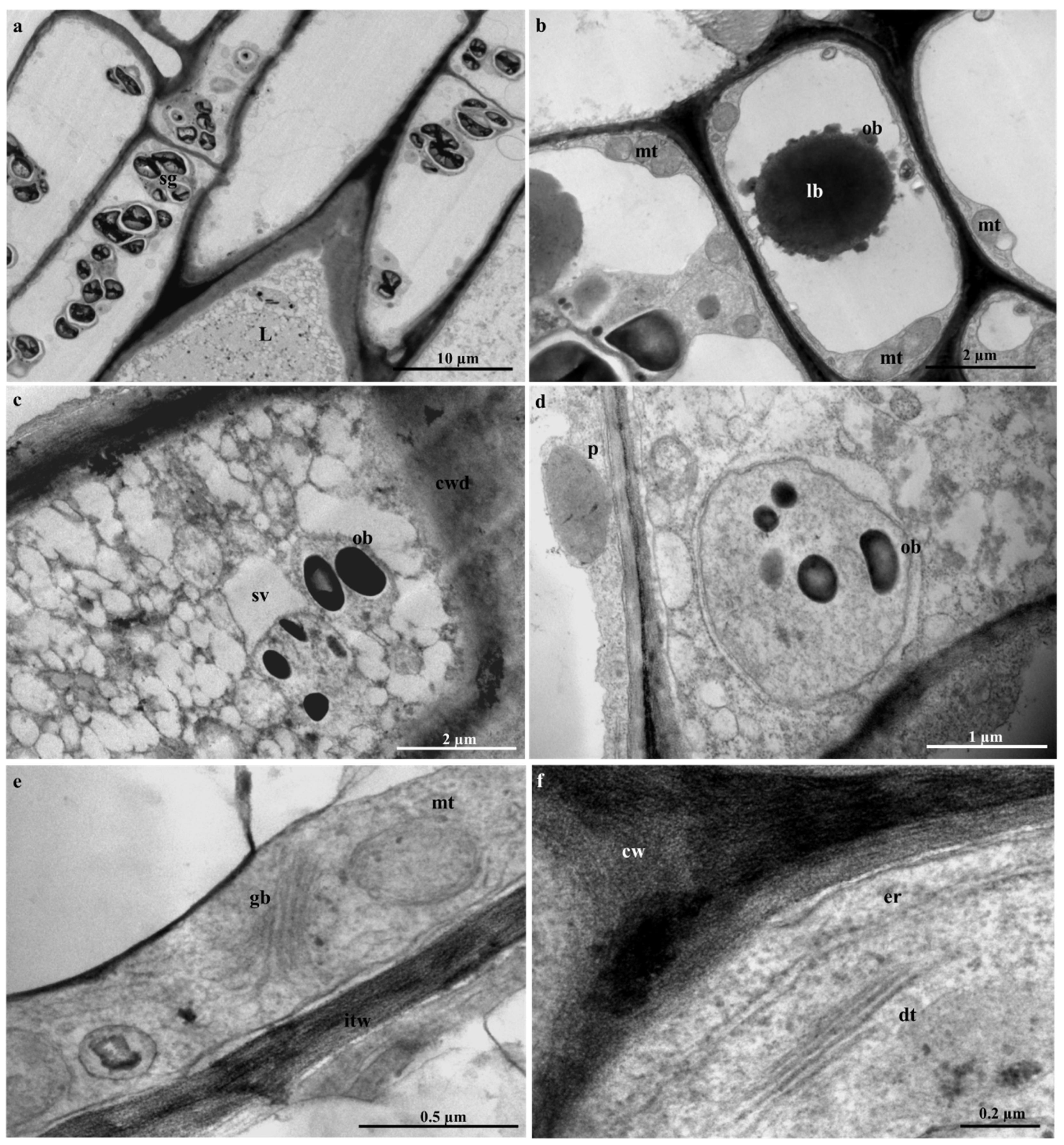
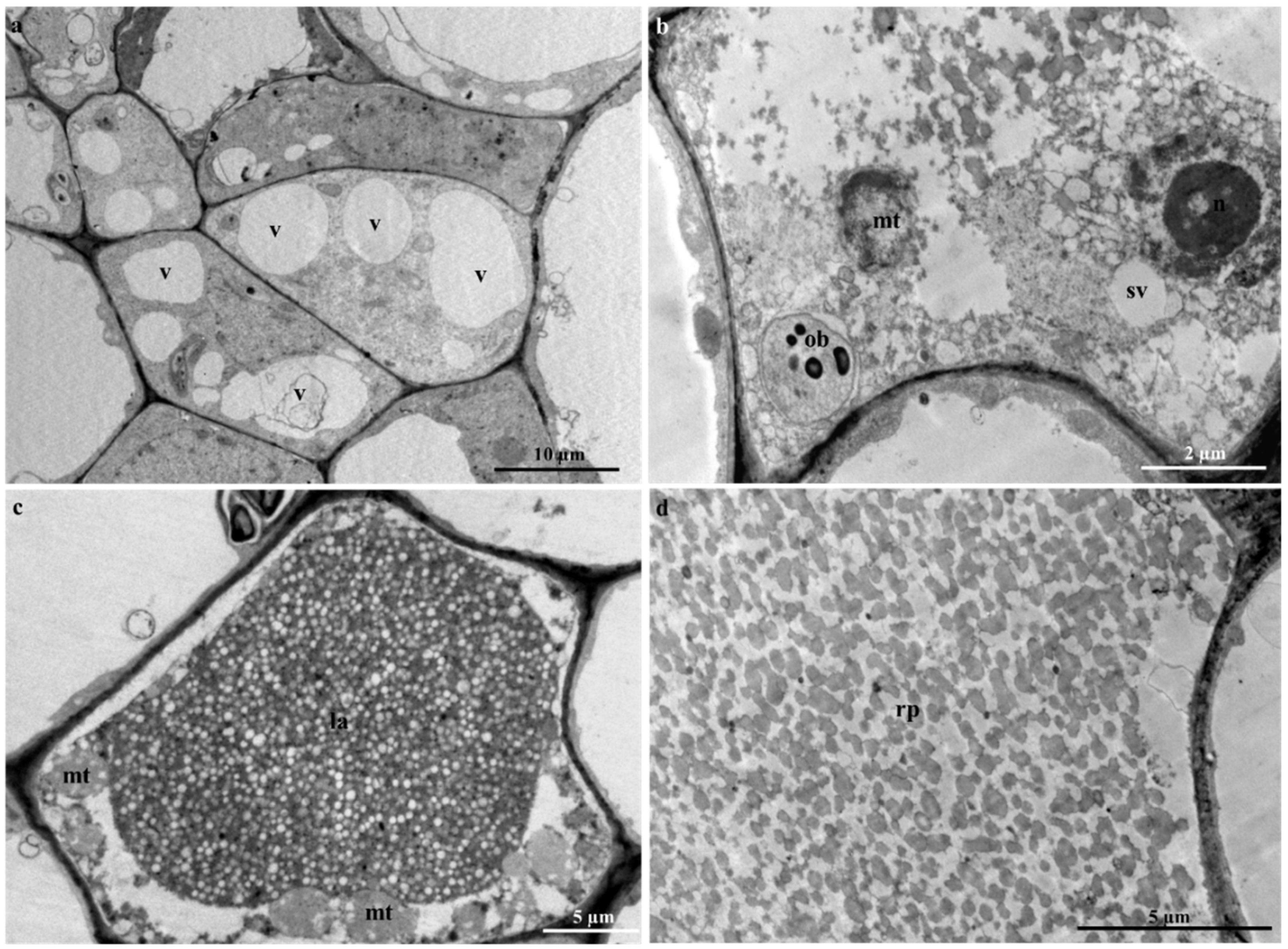
2.5. Laticifer Ultrastructure of Adult Plant Leaves
3. Materials and Methods
3.1. Collection of Leaf and Stem Samples
3.2. Stereomicroscopy
3.3. Scanning Electron Microscopy (SEM)
3.3.1. Chemical Fixation
3.3.2. Freeze-Fracture
3.4. Transmission Electron Microscopy (TEM)
3.5. Histochemistry
3.6. Fluorescence Microscopy
3.7. Whole Mount Staining
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Evert, R.F. Esau’s Plant Anatomy, 3rd ed.; Wiley-Interscience: New York, NY, USA, 2006; pp. 483–495. [Google Scholar]
- Castelblanque, L.; Balaguer, B.; Marti, C.; Rodríguez, J.J.; Orozco, M.; Vera, P. Multiple facets of laticifer cells. Plant Signal. Behav. 2017, 12, e1300743. [Google Scholar] [CrossRef]
- Castelblanque, L.; Balaguer, B.; Marti, C.; Rodriguez, J.J.; Orozco, M.; Vera, P. Novel Insights into the Organization of Laticifer Cells: A Cell Comprising a Unified Whole System. Plant Physiol. 2016, 172, 1032–1044. [Google Scholar] [CrossRef] [PubMed]
- Dussourd, D.; Eisner, T. Vein-cutting behavior: Insect counterploy to the latex defense of plants. Science 1987, 237, 898–901. [Google Scholar] [CrossRef] [PubMed]
- Bauer, G.; Gorb, S.N.; Klein, M.-C.; Nellesen, A.; Von Tapavicza, M.; Speck, T. Comparative Study on Plant Latex Particles and Latex Coagulation in Ficus benjamina, Campanula glomerata and Three Euphorbia species. PLoS ONE 2014, 9, e113336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, M.-L.; A Morgan, J. Metabolic flux analysis of secondary metabolism in plants. Metab. Eng. Commun. 2020, 10, e00123. [Google Scholar] [CrossRef]
- Tiwari, P.; Sangwan, N.S.; Sangwan, N.S. Plant secondary metabolism linked glycosyltransferases: An update on expanding knowledge and scopes. Biotechnol. Adv. 2016, 34, 714–739. [Google Scholar] [CrossRef]
- Pott, D.M.; Osorio, S.; Vallarino, J. From Central to Specialized Metabolism: An Overview of Some Secondary Compounds Derived from the Primary Metabolism for Their Role in Conferring Nutritional and Organoleptic Characteristics to Fruit. Front. Plant Sci. 2019, 10, 835. [Google Scholar] [CrossRef] [Green Version]
- Aharoni, A.; Galili, G. Metabolic engineering of the plant primary–secondary metabolism interface. Curr. Opin. Biotechnol. 2011, 22, 239–244. [Google Scholar] [CrossRef]
- Wink, M. Introduction: Biochemistry, Physiology and Ecological Functions of Secondary Metabolites. Biochem. Plant Second. Metab. 2010, 40, 1–19. [Google Scholar] [CrossRef]
- Esau, K. Plant Anatomy, 3rd ed.; McGraw-Hill: New York, NY, USA, 1965; pp. 483–490. [Google Scholar]
- Metcalfe, C.R. Distribution of latex in the plant kingdom. Econ. Bot. 1967, 21, 115–127. [Google Scholar] [CrossRef]
- Pickard, W.F. Laticifers and secretory ducts: Two other tube systems in plants. New Phytol. 2008, 177, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Ramos, M.V.; Demarco, D.; Souza, I.C.D.C.; De Freitas, C.D.T. Laticifers, Latex, and Their Role in Plant Defense. Trends Plant Sci. 2019, 24, 553–567. [Google Scholar] [CrossRef] [PubMed]
- Fahn, A. Functions and location of secretory tissues in plants and their possible evolutionary trends. Isr. J. Plant Sci. 2002, 50, 302. [Google Scholar] [CrossRef]
- Hagel, J.M.; Yeung, E.C.; Facchini, P.J. Got milk? The secret life of laticifers. Trends Plant Sci. 2008, 13, 631–639. [Google Scholar] [CrossRef]
- Mahlberg, P.G. Laticifers: An historical perspective. Bot. Rev. 1993, 59, 1–23. [Google Scholar] [CrossRef]
- Canaveze, Y.; Machado, S.R. The Occurrence of Intrusive Growth Associated with Articulated Laticifers in Tabernaemontana catharinensis A.DC., a New Record for Apocynaceae. Int. J. Plant Sci. 2016, 177, 458–467. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, K.V.; Venkatasubramanian, P.; Lalitha, S.; Bahadur, B.; Sujatha, M.; Carels, N. Laticifers of Jatropha. In Jatropha; Challenges for a New Energy crop; Springer: New York, NY, USA, 2013; Volume 2, pp. 3–10. [Google Scholar] [CrossRef]
- Demarco, D.; Castro, M.D.M. Laticíferos articulados anastomosados em espécies de Asclepiadeae (Asclepiadoideae, Apocynaceae) e suas implicações ecológicas. Braz. J. Bot. 2008, 31, 701–713. [Google Scholar] [CrossRef] [Green Version]
- Canaveze, Y.; Machado, S.R. Leaf colleters in Tabernaemontana catharinensis (Apocynaceae, Rauvolfioideae): Structure, ontogenesis, and cellular secretion. Botany 2015, 93, 287–296. [Google Scholar] [CrossRef]
- Leeuwenberg, A.J.M.; Kupicha, F.K. Apocynaceae; Launert, E., Ed.; Flora Zambesiana: London, UK, 1985; pp. 395–503. [Google Scholar]
- Schmelzer, G.B.; Gurib-Fakim, A. Medicinal Plants; plant resources of tropical Africa PROTA Foundation, 1st ed.; Backhuys Publishers: Wageningen, The Netherlands, 2008; Volume 1, pp. 597–598. [Google Scholar]
- Schmidt, E.; Lotter, M.; McCleland, W. Trees and Shrubs of Mpumalanga and Kruger National Park; Illustrated, Ed.; Jacana Media: Johannesburg, South Africa, 2002; pp. 566–569. [Google Scholar]
- Mehrbod, P.; Abdalla, M.; Njoya, E.M.; Ahmed, A.S.; Fotouhi, F.; Farahmand, B.; Gado, D.; Tabatabaian, M.; Fasanmi, O.; Jn, E.; et al. South African medicinal plant extracts active against influenza A virus. BMC Complement. Altern. Med. 2018, 18, 112. [Google Scholar] [CrossRef] [Green Version]
- Hanley, M.E.; Lamont, B.; Fairbanks, M.M.; Rafferty, C. Plant structural traits and their role in anti-herbivore defence. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 157–178. [Google Scholar] [CrossRef]
- Konno, K. Plant latex and other exudates as plant defence systems: Roles of various defence chemicals and proteins contained therein. Phytochemistry 2011, 72, 1510–1530. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.A.; Konno, K. Latex: A model for understanding mechanisms, ecology, and evolution of plant defence against herbivory. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 311–331. [Google Scholar] [CrossRef] [Green Version]
- Lopes, K.L.B.; Thadeo, M.; Azevedo, A.A.; Soares, A.A.; Meira, R.M.S.A. Articulated laticifers in the vegetative organs of Mandevilla atroviolacea (Apocynaceae, Apocynoideae). Botany. 2009, 87, 202–209. [Google Scholar] [CrossRef]
- Pirolla-Souza, A.; Arruda, R.C.O.; Pace, M.R.; Farinaccio, M.A. Leaf anatomical characters of Rhabdadenia (Rhabdadenieae, Apocynaceae), their taxonomic implications, and notes on the presence of articulated laticifers in the genus. Plant Syst. Evol. 2019, 305, 797–810. [Google Scholar] [CrossRef]
- Gama, T.D.S.S.; Rubiano, V.S.; Demarco, D. Laticifer development and its growth mode in Allamanda blanchetii A. DC. (Apocynaceae). J. Torrey Bot. Soc. 2017, 144, 303–312. [Google Scholar] [CrossRef]
- Serpe, M.D.; Muir, A.; Driouich, A. Immunolocalization of β-D-glucans, pectins, and arabinogalactan-proteins during intrusive growth and elongation of nonarticulated laticifers in Asclepias speciosa Torr. Planta 2002, 215, 357–370. [Google Scholar] [CrossRef]
- Demarco, D.; Castro, M.D.M.; Ascensão, L. Two laticifer systems in Sapium haematospermum—New records for Euphorbiaceae. Botany 2013, 91, 545–554. [Google Scholar] [CrossRef]
- Marinho, C.; Teixeira, S.P. Novel reports of laticifers in Moraceae and Urticaceae: Revisiting synapomorphies. Plant Syst. Evol. 2018, 305, 13–31. [Google Scholar] [CrossRef]
- Fineran, B.A. Distribution and Organization of Non-articulated Laticifers in Mature Tissues of Poinsettia (Euphorbia pulcherrima Willd.). Ann. Bot. 1982, 50, 207–220. [Google Scholar] [CrossRef]
- Rudall, P. Laticifers in Crotonoideae (Euphorbiaceae): Homology and Evolution. Ann. Missouri Bot. Gard. 1994, 81, 270. [Google Scholar] [CrossRef]
- Kumar, G.; Karthik, L.; Rao, K.V.B. A review on pharmacological and phytochemical profile of Calotropis gigantea Linn. Pharmacol. Online 2011, 1, 1–8. [Google Scholar]
- Nazar, N.; Goyder, D.J.; Clarkson, J.J.; Mahmood, T.; Chase, M.W. The taxonomy and systematics of Apocynaceae: Where we stand in 2012. Bot. J. Linn. Soc. 2013, 171, 482–490. [Google Scholar] [CrossRef] [Green Version]
- Rapini, A.; Chase, M.W.; Goyder, D.J.; Griffiths, J.; Vanderpoorten, A.; Żarnowiec, J. Asclepiadeae classification: Evaluating the phylogenetic relationships of New World Asclepiadoideae (Apocynaceae). TAXON 2003, 52, 33–50. [Google Scholar] [CrossRef]
- Gonçalves, M.P.; Mercadante-Simões, M.O.; Ribeiro, L.M. Ontogeny of anastomosed laticifers in the stem apex of Hancornia speciosa (Apocynaceae): A topographic approach. Protoplasma 2018, 255, 1713–1724. [Google Scholar] [CrossRef]
- De, M.C.M.; Demarco, D. Phenolic Compounds Produced by Secretory Structures in Plants: A Brief Review. Nat. Prod. Commun. 2008, 3, 1273–1284. [Google Scholar] [CrossRef] [Green Version]
- Silveira, D.; De Melo, A.F.; Magalhães, P.; Fonseca, Y. Tabernaemontana Species: Promising Sources of New Useful Drugs. In Bioactive Natural Products; Elsevier BV: Amsterdam, The Netherlands, 2017; Volume 54, pp. 227–289. [Google Scholar]
- Van Beek, T.; Verpoorte, R.; Svendsen, A.; Leeuwenberg, A.; Bisset, N. Tabernaemontana L. (Apocynaceae): A review of its taxonomy, phytochemistry, ethnobotany and pharmacology. J. Ethnopharmacol. 1984, 10, 1–156. [Google Scholar] [CrossRef]
- Marinho, F.F.; Simões, A.O.; Barcellos, T.; Moura, S. Brazilian Tabernaemontana genus: Indole alkaloids and phytochemical activities. Fitoterapia 2016, 114, 127–137. [Google Scholar] [CrossRef]
- Athipornchai, A. A Review on Tabernaemontana spp.: Multipotential Medicinal Plant. Asian J. Pharm. Clin. Res. 2018, 11, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Schripsema, J.; Hermans-Lokkerbol, A.; Van Der Heijden, R.; Verpoorte, R.; Svendsen, A.B.; Van Beek, T.A. Alkaloids of Tabernaemontana ventricosa. J. Nat. Prod. 1986, 49, 733–735. [Google Scholar] [CrossRef]
- Pallant, C.; Cromarty, A.; Steenkamp, V.; Cromarty, A.D. Effect of an alkaloidal fraction of Tabernaemontana elegans (Stapf.) on selected micro-organisms. J. Ethnopharmacol. 2012, 140, 398–404. [Google Scholar] [CrossRef]
- Mahlberg, P.G.; Davis, D.G.; Galitz, D.S.; Manners, G.D. Laticifers and the classification of Euphorbia: The chemotaxonomy of Euphorbia esula L. Bot. J. Linn. Soc. 1987, 94, 165–180. [Google Scholar] [CrossRef]
- Farrell, B.D.; Dussourd, D.E.; Mitter, C. Escalation of Plant Defense: Do Latex and Resin Canals Spur Plant Diversification? Am. Nat. 1991, 138, 881–900. [Google Scholar] [CrossRef]
- Pereira, C.G.; Marques, M.; Barreto, A.S.; Siani, A.C.; Fernandes, E.C.; Meireles, M.A. Extraction of indole alkaloids from Tabernaemontana catharinensis using supercritical CO2+ethanol: An evaluation of the process variables and the raw material origin. J. Supercrit. Fluids 2004, 30, 51–61. [Google Scholar] [CrossRef]
- Thombre, R.; Jagtap, R.; Patil, N. Evaluation of phytoconstituents, anti-bacterial, anti-oxidant and cytotoxic activity of Vitex negundo L. and Tabernaemontana divaricata L. Int. J. Pharma. Bio. Sci. 2013, 4, 389–396. [Google Scholar]
- Demarco, D. Histochemical Analysis of Plant Secretory Structures; Humana Press: New York, NY, USA, 2017; pp. 31–330. [Google Scholar] [CrossRef]
- Wilson, K.J.; Mahlberg, P.G. Ultrastructure of non-articulated laticifers in mature embryos and seedlings of Asclepias syriaca L (Asclepiadaceae). Am. J. Bot. 1978, 65, 98–109. [Google Scholar] [CrossRef]
- Wilson, K.J.; Mahlberg, P.G. Ultrastructure of developing and mature non-articulated laticifers in the milkweed Asclepias syriaca L. (Asclepiadaceae). Am. J. Bot. 1980, 67, 1160–1170. [Google Scholar] [CrossRef]
- Roy, A.T.; De, D.N. Studies on Differentiation of Laticifers through Light and Electron Microscopy in Calotropis gigantea (Linn.) R.Br. Ann. Bot. 1992, 70, 443–449. [Google Scholar] [CrossRef]
- Demarco, D. Micromorfologı´a y histoquı´mica de los laticı ‘feros de’ organos vegetativos de especies de Asclepiadoideae (Apocynaceae). Acta Biol. Colomb. 2015, 20, 57–65. [Google Scholar] [CrossRef]
- O’Brien, T.P.; Feder, N.; McCully, M.E. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 1964, 59, 368–373. [Google Scholar] [CrossRef]
- McCracken, E.; Johansen, D.A. Plant Microtechnique. Trans. Am. Microsc. Soc. 1940, 59, 405. [Google Scholar] [CrossRef]
- Mazia, D.; Brewer, P.A.; Alfert, M. The cytochemical staining and measurement of protein with mercuric bromphenol blue. Boil. Bull. 1953, 104, 57–67. [Google Scholar] [CrossRef]
- Furr, M.; Mahlberg, P.G. Histochemical Analyses of Laticifers and Glandular Trichomes in Cannabis sativa. J. Nat. Prod. 1981, 44, 153–159. [Google Scholar] [CrossRef]
- Periasamy, K. A technique of staining sections of paraffin-embedded plant materials without employing a graded ethanol series. J. R. Microsc. Soc. 1967, 87, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Ascensão, L.; Pais, M.S.S. Glandular Trichomes of Artemisia campestris (ssp. Maritima): Ontogeny and Histochemistry of the Secretory Product. Int. J. Plant Sci. 1987, 148, 221–227. [Google Scholar] [CrossRef]











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naidoo, C.; Naidoo, Y.; Dewir, Y.H. The Secretory Apparatus of Tabernaemontana ventricosa Hochst. ex A.DC. (Apocynaceae): Laticifer Identification, Characterization and Distribution. Plants 2020, 9, 686. https://doi.org/10.3390/plants9060686
Naidoo C, Naidoo Y, Dewir YH. The Secretory Apparatus of Tabernaemontana ventricosa Hochst. ex A.DC. (Apocynaceae): Laticifer Identification, Characterization and Distribution. Plants. 2020; 9(6):686. https://doi.org/10.3390/plants9060686
Chicago/Turabian StyleNaidoo, Clarissa, Yougasphree Naidoo, and Yaser Hassan Dewir. 2020. "The Secretory Apparatus of Tabernaemontana ventricosa Hochst. ex A.DC. (Apocynaceae): Laticifer Identification, Characterization and Distribution" Plants 9, no. 6: 686. https://doi.org/10.3390/plants9060686